
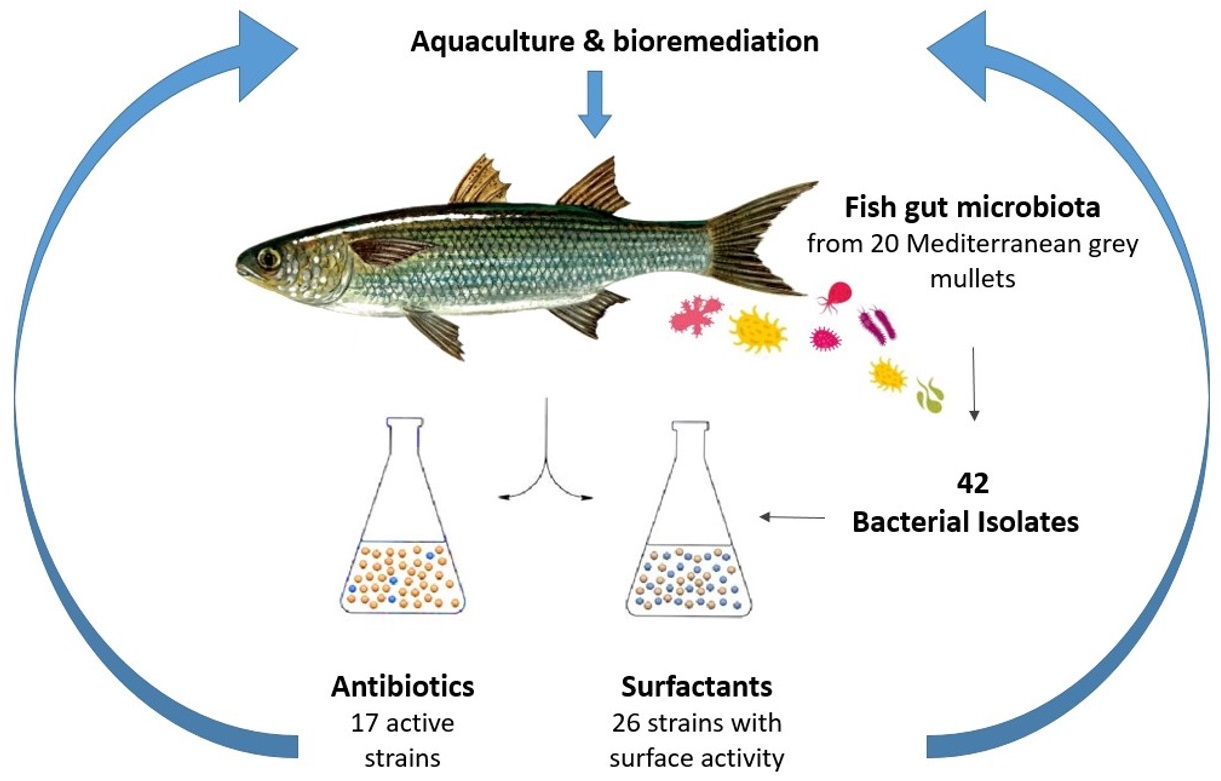
Isolation and Identification of Bacteria with Surface and Antibacterial Activity from the Gut of Mediterranean Grey Mullets

 ,
,  ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area, Sampling and Microbiological Analysis
2.2. Screening of Bacteria for Biosurfactant Production
2.2.1. Detection of the BSs in the Culture Broth
2.2.2. Emulsification Index (E-24)
2.2.3. Drop-Collapse Assay
2.2.4. Surface Tension Measurement
2.3. Biosurfactant Extraction and Thin Layer Chromatography (TLC)
2.4. Bacterial Identification
2.5. BS Antibacterial Activity
2.5.1. Bacterial Pathogens
2.5.2. Antibacterial Activity
- Cell-Free Supernatants
- Crude Extracts
3. Results
3.1. Enumeration of Bacteria and Colony Isolation
3.2. Screening of Bacteria for BS Production
3.3. Bacterial Identification
3.4. BSs Extracts and Thin Layer Chromatography (TLC)
3.5. Antibacterial Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- De Pascale, D.; De Santi, C.; Fu, J.; Landfald, B. The microbial diversity of Polar environments is a fertile ground for bioprospecting. Mar. Genom. 2012, 8, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Bălașa, A.F.; Chircov, C.; Grumezescu, A.M. Marine Biocompounds for Neuroprotection—A Review. Mar. Drugs 2020, 18, 290. [Google Scholar] [CrossRef]
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef] [PubMed]
- Floris, R.; Rizzo, C.; Giudice, A.L. Biosurfactants from Marine Microorganisms. In Metabolomics—New Insights into Biology and Medicine; IntechOpen: London, UK, 2020. [Google Scholar]
- Dusane, D.H.; Matkar, P.; Venugopalan, V.P.; Kumar, A.R.; Zinjarde, S.S. Cross-Species Induction of Antimicrobial Compounds, Biosurfactants and Quorum-Sensing Inhibitors in Tropical Marine Epibiotic Bacteria by Pathogens and Biofouling Microorganisms. Curr. Microbiol. 2011, 62, 974–980. [Google Scholar] [CrossRef]
- Floris, R.; Scanu, G.; Fois, N.; Rizzo, C.; Malavenda, R.; Spanò, N.; Giudice, A.L. Intestinal bacterial flora of Mediterranean gilthead sea bream (Sparus aurata Linnaeus) as a novel source of natural surface active compounds. Aquac. Res. 2018, 49, 1262–1273. [Google Scholar] [CrossRef]
- Rizzo, C.; Gugliandolo, C.; Giudice, A.L. Exploring Mediterranean and Arctic Environments as a Novel Source of Bacteria Producing Antibacterial Compounds to be Applied in Aquaculture. Appl. Sci. 2020, 10, 4006. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, O.; de Vivo, L.; Sulpizio, R.; Gioacchini, G.; Olivotto, I.; Silvi, S.; Cresci, A. Growth improvement by probiotic in European sea bass juveniles (Dicentrarchus labrax, L.), with particular attention to IGF-1, myostatin and cortisol gene expression. Aquaculture 2006, 258, 430–438. [Google Scholar] [CrossRef]
- Floris, R.; Manca, S.; Fois, N. Microbial ecology of intestinal tract of gilthead sea bream (Sparus aurata Linnaeus, 1758) from two coastal lagoons of Sardinia (Italy). Trans. Water Bullet 2013, 7, 4–12. [Google Scholar] [CrossRef]
- Floris, R.; Sanna, G.; Satta, C.; Piga, C.; Sanna, F.; Lugliè, A.; Fois, N. Intestinal Microbial Ecology and Fillet Metal Chemistry of Wild Grey Mullets Reflect the Variability of the Aquatic Environment in a Western Mediterranean Coastal Lagoon (Santa Giusta, Sardinia, Italy). Water 2021, 13, 879. [Google Scholar] [CrossRef]
- Li, J.; Ni, J.; Wang, C.; Li, X.; Wu, S.; Zhang, T.; Yu, Y.; Yan, Q. Comparative study on gastrointestinal microbiota of eight fish species with different feeding habits. J. Appl. Microbiol. 2014, 117, 1750–1760. [Google Scholar] [CrossRef]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: Factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Burtseva, O.; Baulina, O.; Zaytseva, A.; Fedorenko, T.; Chekanov, K.; Lobakova, E. In vitro Biofilm Formation by Bioluminescent Bacteria Isolated from the Marine Fish Gut. Microb. Ecol. 2021, 81, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Malavenda, R.; Rizzo, C.; Michaud, L.; Gerçe, B.; Bruni, V.; Syldatk, C.; Hausmann, R.; Lo Giudice, A. Biosurfactant production by Arctic and Antarctic bacteria growing on hydrocarbons. Pol. Biol. 2015, 38, 1565–1574. [Google Scholar] [CrossRef]
- Satpute, S.K.; Banat, I.; Dhakephalkar, P.K.; Banpurkar, A.G.; Chopade, B.A. Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnol. Adv. 2010, 28, 436–450. [Google Scholar] [CrossRef]
- Panjiar, N.; Sachan, S.G.; Sachan, A. Screening of bioemulsifier-producing micro-organisms isolated from oil-contaminated sites. Ann. Microbiol. 2015, 65, 753–764. [Google Scholar] [CrossRef]
- Rizzo, C.; Michaud, L.; Hörmann, B.; Gerçe, B.; Syldatk, C.; Hausmann, R.; De Domenico, E.; Giudice, A.L. Bacteria associated with sabellids (Polychaeta: Annelida) as a novel source of surface active compounds. Mar. Pollut. Bull. 2013, 70, 125–133. [Google Scholar] [CrossRef]
- Giri, S.S.; Kim, H.J.; Kim, S.; Kim, S.; Kwon, J.; Bin Lee, S.; Park, S.C. Immunomodulatory Role of Microbial Surfactants, with Special Emphasis on Fish. Int. J. Mol. Sci. 2020, 21, 7004. [Google Scholar] [CrossRef]
- Mukherjee, A.; Banerjee, G.; Mukherjee, P.; Ray, A.K.; Chandra, G.; Ghosh, K. Antibacterial substances produced by pathogen inhibitory gut bacteria in Labeo rohita: Physicochemical characterization, purification and identification through MALDI-TOF mass spectrometry. Microb. Pathog. 2019, 130, 146–155. [Google Scholar] [CrossRef]
- Crosetti, D. Biology, Ecology and Culture of Grey Mullet (Mugilidae); Version Date 20151214; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2016; pp. 42–127. [Google Scholar]
- Thomson, J.M. The Mugilidae of the world. Mem. Queens. Mus. 1997, 41, 457–562. [Google Scholar]
- Caredda, M.; Addis, M.; Pes, M.; Fois, N.; Sanna, G.; Piredda, G.; Sanna, G. Physicochemical, colorimetric, rheological parameters and chemometric discrimination of the origin of Mugil cephalus’ roes during the manu-facturing process of Bottarga. Food Res. Int. 2018, 108, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.-D.; Blel, H.; Shen, K.; Koutrakis, E.; Guinand, B. Population genetic structure of Mugil cephalus in the Mediterranean and Black Seas: A single mitochondrial clade and many nuclear barriers. Mar. Ecol. Prog. Ser. 2013, 474, 243–261. [Google Scholar] [CrossRef] [Green Version]
- Turan, C.M.; Gürlek, D.; Ergüden, D.; Yağlıoğlu, D.; Öztürk, B. Systematic status of nine mullet species (Mugilidae) in the Mediterranean Sea. Turk. J. Fish. Aquat. Sci. 2011, 11, 315–321. [Google Scholar] [CrossRef]
- Kumar, R.S.; Kanmani, P.; Yuvaraj, N.; Paari, K.; Pattukumar, V.; Arul, V. Purification and characterization of enterocin MC13 produced by a potential aquaculture probiont Enterococcus faeciumMC13 isolated from the gut of Mugil cephalus. Can. J. Microbiol. 2011, 57, 993–1001. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chen, Y.-S.; Wu, H.-C.; Pan, S.-F.; Yu, B.; Chiang, C.-M.; Chiu, C.-M.; Yanagida, F. Screening and characterization of LAB-produced bacteriocin-like substances from the intestine of grey mullet (Mugil cephalus L.) as potential biocontrol agents in aquaculture. J. Appl. Microbiol. 2013, 114, 299–307. [Google Scholar] [CrossRef]
- Cui, J.; Xiao, M.; Liu, M.; Wang, Z.; Liu, F.; Guo, L.; Meng, H.; Zhang, H.; Yang, J.; Deng, D.; et al. Coupling metagenomics with cultivation to select host-specific probiotic micro-organisms for subtropical aquaculture. J. Appl. Microbiol. 2017, 123, 1274–1285. [Google Scholar] [CrossRef]
- Sechi, N.; Fiocca, F.; Sannio, A.; Lugliè, A. Santa Giusta Lagoon (Sardinia): Phytoplankton and nutrients before and after waste water diversion. J. Limnol. 2001, 60, 194–200. [Google Scholar] [CrossRef]
- Techaoei, S.; Leelapornpisid, P.; Santiarwarn, D.; Lumyong, S. Preliminary screenings of biosurfac-tants-producing microorganisms isolated from hot spring and garages in northern Thailand. Curr. Appl. Sci. Technol. 2007, 7, 38–43. [Google Scholar]
- Bodour, A.A.; Drees, K.P.; Maier, R.M. Distribution of Biosurfactant-Producing Bacteria in Undisturbed and Contaminated Arid Southwestern Soils. Appl. Environ. Microbiol. 2003, 69, 3280–3287. [Google Scholar] [CrossRef] [Green Version]
- Samadi, N.; Fazel, M.R.; Abadian, N.; Akhavan, A.; Tahzibi, A.; Jamalifar, H. Biosurfactant Production by the Strain Isolated from Contaminated Soil. J. Biol. Sci. 2007, 7, 1266–1269. [Google Scholar] [CrossRef] [Green Version]
- Jamal, P.; Wan Nawawi, W.M.F.; Alam, M.Z. Optimum medium components for biosurfactant production by Klebsiella pneumoniae WMF02 utilizing sludge palm oil as a substrate. Austral. J. Basic Appl. Sci. 2012, 6, 100–108. [Google Scholar]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Gomila, M.; Penã, A.; Mulet, M.; Lalucat, J.; Garcia-Valdes, E. Phylogenomics and systematics in Pseudomonas. Front. Microbiol. 2015, 6, 214. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Strimmer, K.; von Haeseler, A. Likelihood-mapping: A simple method to visualize phylogenetic content of a sequence alignment. Proc. Natl. Acad. Sci. USA 1997, 94, 6815–6819. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Quang Minh, B.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Meth. 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Simonetta, A.C.; de Velasco, L.G.M.; Frisón, L.N. Antibacterial activity of enterococci strains against Vibrio cholerae. Lett. Appl. Microbiol. 1997, 24, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Council of Europe. Pharmacopoeia, 3rd ed.; Council of Europe: Strasbourg, France, 1997; p. 121. [Google Scholar]
- Twigg, M.S.; Baccile, N.; Banat, I.M.; Déziel, E.; Marchant, R.; Roelants, S.; Van Bogaert, I.N.A. Microbial biosurfactant research: Time to improve the rigour in the reporting of synthesis, functional characterization and process development. Microb. Biotechnol. 2021, 14, 147–170. [Google Scholar] [CrossRef]
- Rizzo, C.; Michaud, L.; Syldatk, C.; Hausmann, R.; De Domenico, E.; Giudice, A.L. Influence of salinity and temperature on the activity of biosurfactants by polychaete-associated isolates. Environ. Sci. Pollut. Res. 2013, 21, 2988–3004. [Google Scholar] [CrossRef]
- Rizzo, C.; Syldatk, C.; Hausmann, R.; Gerçe, B.; Longo, C.; Papale, M.; Conte, A.; De Domenico, E.; Luigi Michaud, L.; Lo Giudice, A. The demosponge Halichondria (Halichondria) panicea (Pallas, 1766) as a novel source of bio-surfactant-producing bacteria. J. Basic. Microbiol. 2018, 58, 532–542. [Google Scholar] [CrossRef]
- Tuleva, B.K.; Ivanov, G.R.; Christova, N.E. Biosurfactant Production By A New Pseudomonas Putida Strain. Z. Für Nat. C 2002, 57, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Rahman, P.; Gakpe, E. Production, Characterisation and Applications of Biosurfactants-Review. Biotechnology 2008, 7, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Meliani, A.; Bensoltane, A. The ability of some Pseudomonas strains to produce biosurfactants. Poult. Fish. Wildl. Sci. 2014, 2, 112. [Google Scholar] [CrossRef] [Green Version]
- Twigg, M.S.; Tripathi, L.; Zompra, A.; Salek, K.; Irorere, V.; Gutierrez, T.; Spyroulias, G.A.; Marchant, R.; Banat, I.M. Identification and characterisation of short chain rhamnolipid production in a previously uninvestigated, non-pathogenic marine pseudomonad. Appl. Microbiol. Biotechnol. 2018, 102, 8537–8549. [Google Scholar] [CrossRef] [Green Version]
- Nikouli, E.; Meziti, A.; Antonopoulou, E.; Mente, E.; Kormas, K.A. Gut Bacterial Communities in Geographically Distant Populations of Farmed Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax). Microorganisms 2018, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Shu, Q.; Lou, H.; Wei, T.; Liu, X.; Chen, Q. Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review. Pharmaceutics 2021, 13, 227. [Google Scholar] [CrossRef] [PubMed]
- Chrzanowski, L.; Lawniczak, L.; Czaczyk, K. Why do microorganisms produce rhamnolipids? World J. Microbiol. Biotechnol. 2011, 28, 401–419. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [Green Version]
- Soberón-Chávez, G.; Hausmann, R.; Déziel, E. Editorial: Biosurfactants: New Insights in Their Biosynthesis, Production and Applications. Front. Bioeng. Biotechnol. 2021, 9, 769899. [Google Scholar] [CrossRef] [PubMed]
- Soberón-Chávez, G.; González-Valdez, A.; Soto-Aceves, M.P.; Cocotl-Yañez, M. Rhamnolipids produced by Pseudomonas: From molecular genetics to the market. Microb. Biotechnol. 2021, 14, 136–146. [Google Scholar] [CrossRef]
- Uzoigwe, C.; Burgess, J.G.; Ennis, C.J.; Rahman, P.K.S.M. Bioemulsifiers are not biosurfactants and require different screening approaches. Front. Microbiol. 2015, 6, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. The State of World Fisheries and Aquaculture 2010; Food and Agriculture Organization: Rome, Italy, 2010; p. 179. [Google Scholar] [CrossRef]
- Ramírez, C.; Gutiérrez, M.S.; Venegas, L.; Sapag, C.; Araya, C.; Caruffo, M.; López, P.; Reyes-Jara, A.; Toro, M.; González-Rocha, G.; et al. Microbiota composition and susceptibility to florfenicol and oxytetracycline of bacterial isolates from mussels (Mytilus spp.) reared on different years and distance from salmon farms. Environ. Res. 2021, 204, 112068. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Koshio, S.; Esteban, M. Ángeles Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Costa-Pierce, B.A.; Bartley, D.M.; Hasan, M.; Yusoff, F.; Kaushik, S.J.; Rana, K.; Lemos, D.; Bueno, P.; Ya-kupitiyage, A. 2012. Responsible use of resources for sustainable aquaculture. In Farming the Waters for People and Food, Proceedings of the Global Conference on Aquaculture, Phuket, Thailand, 22–25 September 2010; Subasinghe, R.P., Arthur, J.R., Bartley, D.M., De Silva, S.S., Halwart, M., Hishamunda, N., Mohan, C.V., Sorgeloos, P., Eds.; FAO: Rome, Italy; NACA: Bangkok, Thailand, 2010; pp. 113–147. [Google Scholar]
- Bledsoe, G.; Bledsoe, C.; Rasco, B. Caviars and Fish Roe Products. Crit. Rev. Food Sci. Nutr. 2003, 43, 317–356. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Fish Species | Bacterial Affiliation | GeneBank Accession Number | Drop Collapse | E-24 (%) | Surface TensionmN·m−1 | BS Type |
|---|---|---|---|---|---|---|---|
| 1 | CR | Pseudomonas aeruginosa | MW369461 | +++ | 70.5 ± 9.1 | 36.5 ± 0.1 | Rhamnolipid |
| 3 | CR | Pseudomonas aeruginosa | OK342256 | +++ | 68.0 ± 12.7 | 37.1 ± 0.1 | Rhamnolipid |
| 5 | CR | Pseudomonas aeruginosa | OK342257 | +++ | 77.0 ± 0.0 | 36.9 ± 0.4 | Rhamnolipid |
| 6 | CR | Pseudomonas aeruginosa | MW369462 | +++ | 56.4 ± 0.0 | 37.1 ± 0.1 | Less polar compound |
| 8 | CR | Pseudomonas aeruginosa | OK342258 | ++ | 0.0 ± 0.0 | 37.1 ± 0.1 | nd |
| 9 | CR | Pseudomonas aeruginosa | OK342259 | +++ | 57.7 ± 1.8 | 37.2 ± 0.3 | nd |
| 10 | CR | Pseudomonas alcaligenes | MW369463 | + | 50.0 ± 1.8 | 37.2 ± 0.3 | nd |
| 11 | CR | Aeromonas caviae | MW369464 | - | 0.0 ± 0.0 | 43.0 ± 0.1 | nd |
| 12 | CR | - | - | +++ | 15.4 ± 21.8 | 36.9 ± 0.1 | nd |
| 13 | CR | Pseudomonas aeruginosa | MW369465 | +++ | 51.3 ± 3.6 | 36.9 ± 0.1 | Rhamnolipid |
| 15 | CR | Pseudomonas aeruginosa | MW369466 | +++ | 59.0 ± 3.6 | 36.6 ± 0.6 | Rhamnolipid |
| 16 | CR | - | - | weak | 0.0 ± 0.0 | 35.35 ± 0.6 | nd |
| 17 | CR | Pseudomonas mendocina | MW369467 | - | 20.5 ± 0 | nd | nd |
| 18 | CR | Pseudomonas putida | OK342260 | weak | 33.3 ± 3.6 | 36.1 ± 0.1 | Less polar compound |
| 19 | MC | Pseudomonas sp. | OK342261 | + | 28.2 ± 3.6 | 35.2 ± 0.0 | Less polar compound |
| 20 | MC | Pseudomonas alcaliphila | MW369468 | weak | 0.0 ± 0.0 | 35.0 ± 0.4 | nd |
| 21 | MC | - | - | weak | 25.6 ± 14.5 | nd | nd |
| 22 | MC | Pseudomonas sp. | OK342262 | + | 25.6 ± 0.0 | 35.1 ± 0.2 | Less polar compound |
| 23 | MC | Pseudomonas sp. | OK342263 | weak | 0.0 ± 0.0 | 36.5 ± 0.1 | nd |
| 24 | MC | Pseudomonas khazarica | MW369469 | weak | 0.0 ± 0.0 | nd | nd |
| 25 | MC | Pseudomonas sp. | OK342264 | + | 0.0 ± 0.0 | 35.3 ± 0.1 | Less polar compounds |
| 26 | MC | Pseudomonas aeruginosa | MW369470 | ++ | 33.3 ± 0.0 | 37.6 ± 0.3 | Less polar compound |
| 28 | CS | Enterobacter ludwigii | MW369471 | + | 0.0 ± 0.0 | 37.9 ± 0.1 | nd |
| 30 | CS | Aeromonas media | MW369472 | weak | 0.0 ± 0.0 | 39.4 ± 0.9 | nd |
| 35 | CS | Aeromonas taiwanensis | MW369473 | weak | 0.0 ± 0.0 | 43.2 ± 0.1 | nd |
| 37 | CL | Aeromonas media | MW369474 | - | 0.0 ± 0.0 | 35.9 ± 0.1 | nd |
| 40 | CL | Aeromonas media | MW369476 | - | 0.0 ± 0.0 | 46.1 ± 0.3 | nd |
| 41 | CL | Pseudomonas anguilliseptica | MW369477 | + | 32.1 ± 5.4 | 35.2 ± 0.6 | nd |
| 45 | CL | Pseudomonas stutzeri | OK342265 | + | 0.0 ± 0.0 | 36.3 ± 0.1 | nd |
| 47 | CL | Pseudomonas protegens | MW369478 | weak | 0.0 ± 0.0 | 40.5 ± 0.4 | nd |
| 51 | CL | Pseudomonas protegens | OK342266 | + | 0.0 ± 0.0 | 35.5 ± 0.1 | nd |
| 55 | CL | Pseudomonas protegens | MW369480 | - | 0.0 ± 0.0 | 37.7 ± 0.1 | nd |
| 56 | CL | Pseudomonas sp. | OK342267 | + | 0.0 ± 0.0 | 39.9 ± 0.1 | nd |
| Cell-Free Supernatants (CFSs) and Crude Extracts (CEs) (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Test | S. aureus H1610 | P. mirabilis H1643 | K. pneumoniae H1637 | A. hydrophila H1563 | ||||
| CFSs | CEs | CFSs | CEs | CFSs | CEs | CFSs | CEs | |
| Pseudomonas aeruginosa 1 | 13.5 ± 0.7 | 5.5 ± 0.7 | 15 ± 0.0 | - | - | 17.5 ± 0.7 | - | - |
| Pseudomonas aeruginosa 3 | 10.5 ± 2.1 | 5.5 ± 0.7 | 18.5 ± 0.7 | - | - | 8.0 ± 0.0 | - | - |
| Pseudomonas aeruginosa 5 | - | 5.5 ± 0.7 | - | 15.5 ± 0.7 | - | 6.0 ± 0.0 | - | - |
| Pseudomonas aeruginosa 6 | + | 8.5 ± 0.7 | - | 12.5 ± 0.7 | - | 11.5 ± 0.7 | - | - |
| Pseudomonas aeruginosa 8 | - | 6.5 ± 0.7 | + | - | - | 7.0 ± 0.0 | - | 6.5 ± 0.7 |
| Pseudomonas aeruginosa 9 | - | 14.5 ± 0.7 | - | - | - | 12.0 ± 0.0 | - | - |
| Pseudomonas alcaligenes 10 | 12 ± 0.0 | 7.5 ± 0.7 | - | 16.0 ± 0.0 | - | + | - | - |
| Aeromonas caviae 11 | - | 8.5 ± 0.7 | - | - | - | + | - | - |
| Unidentified 12 | 9 ± 1.4 | 7.5 ± 0.7 | 17.0 ± 1.4 | - | - | 7.5 ± 0.7 | - | - |
| Pseudomonas aeruginosa 13 | 12.5 ± 0.7 | 12.5 ± 0.7 | 7.0 ± 0.0 | - | - | 12.5 ± 0.7 | - | 7.0 ± 0.0 |
| Pseudomonas aeruginosa 15 | 14 ± 0.0 | 12.5 ± 0.7 | - | 14.0 ± 0.0 | - | 12.5 ± 0.7 | - | - |
| Unidentified 16 | - | - | 10.0 ± 0.0 | - | - | - | - | - |
| Pseudomonas putida 18 | - | - | + | - | - | - | - | - |
| Pseudomonas sp. 19 | - | 7.5 ± 0.7 | - | - | - | 8.0 ± 0.0 | - | + |
| Pseudomonas alcaliphila 20 | - | - | - | - | - | - | - | - |
| Pseudomonas sp. 22 | - | 7.5 ± 0.7 | 13.0 ± 0.0 | - | - | 5.5 ± 0.7 | - | - |
| Pseudomonas sp. 23 | - | + | + | - | - | - | - | - |
| Pseudomonas sp. 25 | 9 ± 0.0 | + | - | 12.0 ± 0.0 | - | 6.5 ± 0.7 | - | - |
| Pseudomonas aeruginosa 26 | - | - | 10 ± 0.0 | - | - | - | - | - |
| Enterococcus ludwigii 28 | - | 5.5 ± 0.7 | 9.0 ± 0.0 | - | + | + | - | - |
| Aeromonas media 30 | - | 5.5 ± 0.7 | - | - | - | 6.0 ± 0.0 | - | - |
| Aeromonas taiwanensis 35 | - | 5.5 ± 0.7 | - | - | - | 6.0 ± 0.0 | - | - |
| Pseudomonas protegens 37 | - | - | 10.0 ± 0.0 | - | - | - | - | - |
| Aeromonas media 40 | - | 7.5 ± 0.7 | - | 15.5 ± 0.7 | - | + | - | + |
| Pseudomonas anguilliseptica 41 | - | + | - | - | - | - | - | 8.0 ± 0.0 |
| Pseudomonas stutzeri 45 | - | - | + | - | - | - | - | - |
| Pseudomonas protegens 47 | - | 9.5 ± 0.7 | 8.0 ± 0.0 | - | - | - | - | 6.5 ± 0.7 |
| Pseudomonas protegens 51 | - | 7.0 ± 0.7 | - | - | - | 7.5 ± 0.7 | - | - |
| Pseudomonas protegens 55 | 10 ± 0.0 | 9.5 ± 0.7 | - | - | - | - | - | - |
| Pseudomonas sp. 56 | 13.5 ± 0.7 | 5.5 ± 0.7 | 8.0 ± 0.0 | - | - | + | - | - |
| Negative control | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | ||||
| Chloramphenicol | 21 ± 0.0 | - | + | 30.0 ± 0.0 | ||||
| Gentamycin CN30 | - | 14 | 8.0 ± 0.0 | 18.0 ± 0.0 | ||||
| Amoxycillin | - | - | - | - | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floris, R.; Sanna, G.; Mura, L.; Fiori, M.; Culurgioni, J.; Diciotti, R.; Rizzo, C.; Lo Giudice, A.; Laganà, P.; Fois, N. Isolation and Identification of Bacteria with Surface and Antibacterial Activity from the Gut of Mediterranean Grey Mullets. Microorganisms 2021, 9, 2555. https://doi.org/10.3390/microorganisms9122555
Floris R, Sanna G, Mura L, Fiori M, Culurgioni J, Diciotti R, Rizzo C, Lo Giudice A, Laganà P, Fois N. Isolation and Identification of Bacteria with Surface and Antibacterial Activity from the Gut of Mediterranean Grey Mullets. Microorganisms. 2021; 9(12):2555. https://doi.org/10.3390/microorganisms9122555
Chicago/Turabian StyleFloris, Rosanna, Gabriele Sanna, Laura Mura, Myriam Fiori, Jacopo Culurgioni, Riccardo Diciotti, Carmen Rizzo, Angelina Lo Giudice, Pasqualina Laganà, and Nicola Fois. 2021. "Isolation and Identification of Bacteria with Surface and Antibacterial Activity from the Gut of Mediterranean Grey Mullets" Microorganisms 9, no. 12: 2555. https://doi.org/10.3390/microorganisms9122555
APA StyleFloris, R., Sanna, G., Mura, L., Fiori, M., Culurgioni, J., Diciotti, R., Rizzo, C., Lo Giudice, A., Laganà, P., & Fois, N. (2021). Isolation and Identification of Bacteria with Surface and Antibacterial Activity from the Gut of Mediterranean Grey Mullets. Microorganisms, 9(12), 2555. https://doi.org/10.3390/microorganisms9122555