
The Campylobacter jejuni Response Regulator and Cyclic-Di-GMP Binding CbrR Is a Novel Regulator of Flagellar Motility

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Routine Growth Conditions
2.2. cbrR− Mutant Construction
2.3. Complementation of cbrR− Mutant
2.4. Purification of CbrR Protein
2.5. Motility Assays
2.6. Transmission Electron Microscopy
2.7. Autoagglutination Assay
2.8. Biofilm Assays and Scanning Electron Microscopy
2.9. Site-Directed Mutagenesis
2.10. Differential Radial Capillary Action of Ligand Assay
2.11. INT-407 Adherence/Invasion Assay
2.12. Nucleotide Extractions for c-di-GMP Detection via LC-MS/MS
3. Results
3.1. C. jejuni CbrR Is Annotated as a DGC Similar to C. crescentus PleD
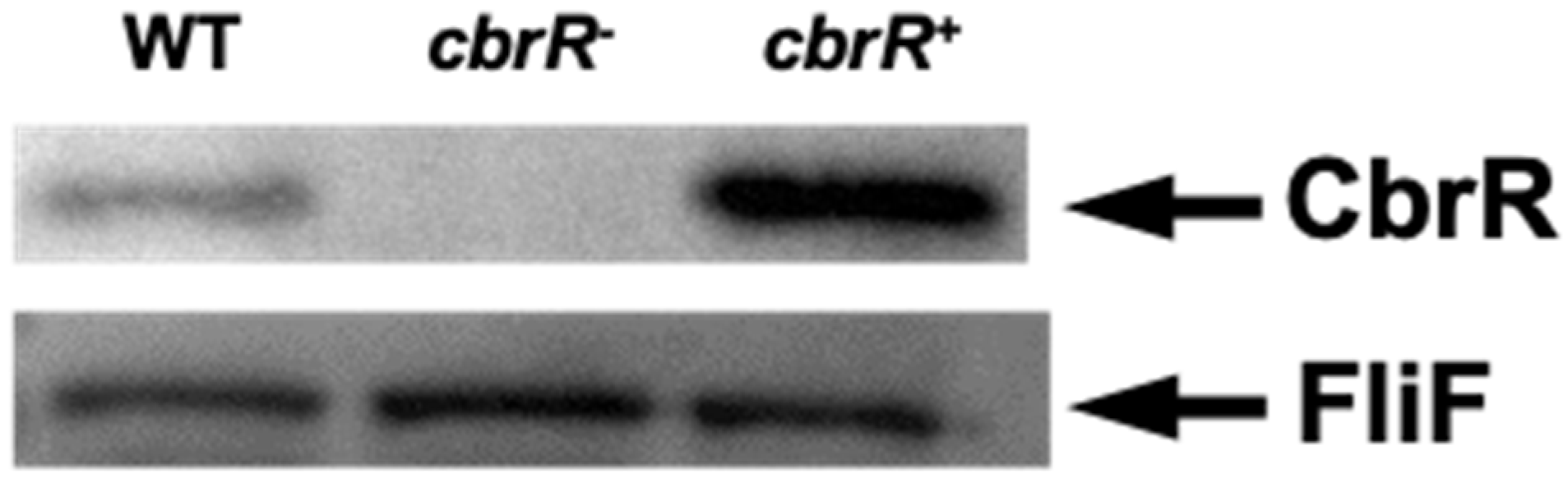
3.2. CbrR Is Overexpressed in cbrR+ Complement
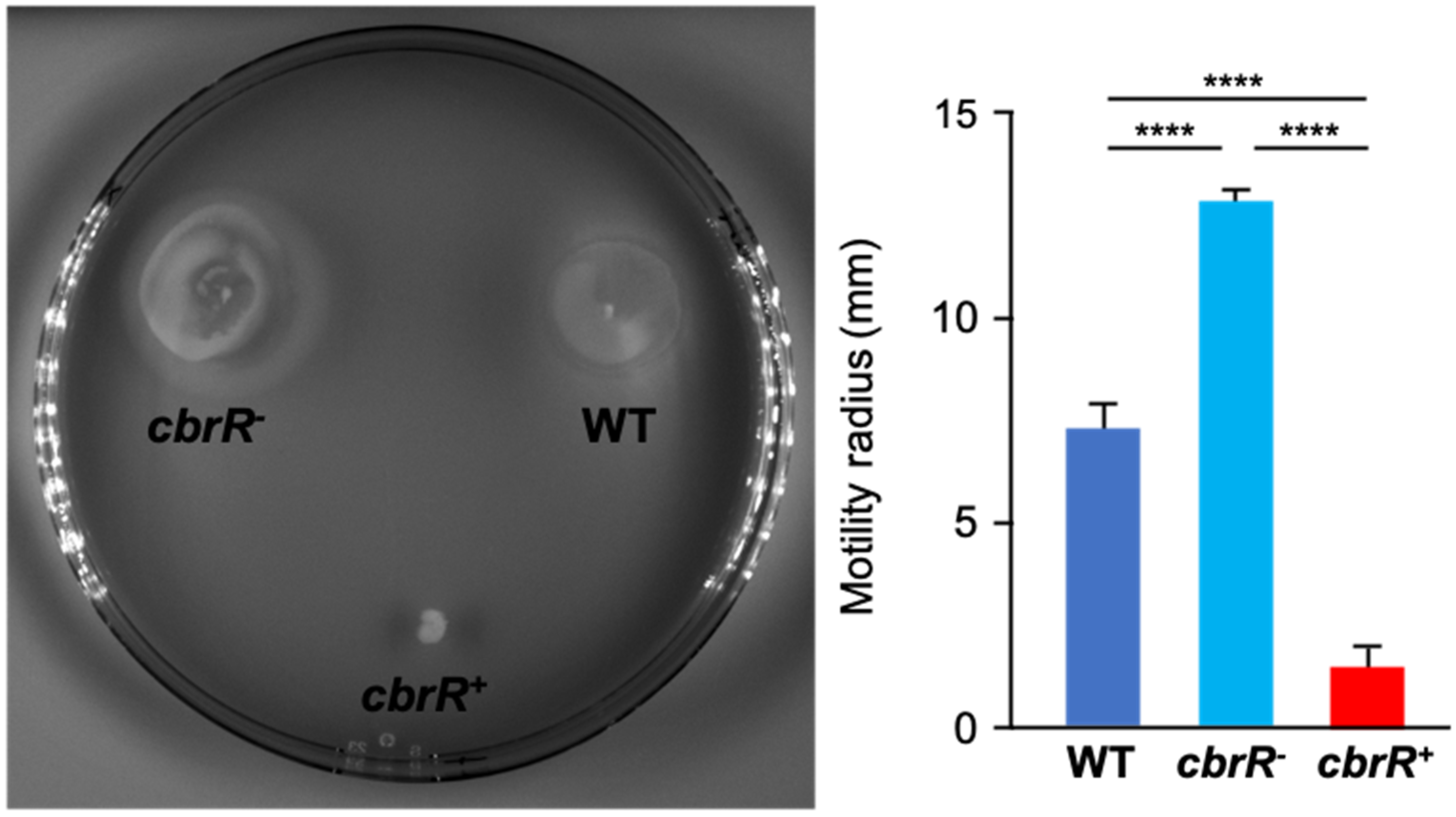
3.3. Motility Is Enhanced in the Absence of CbrR
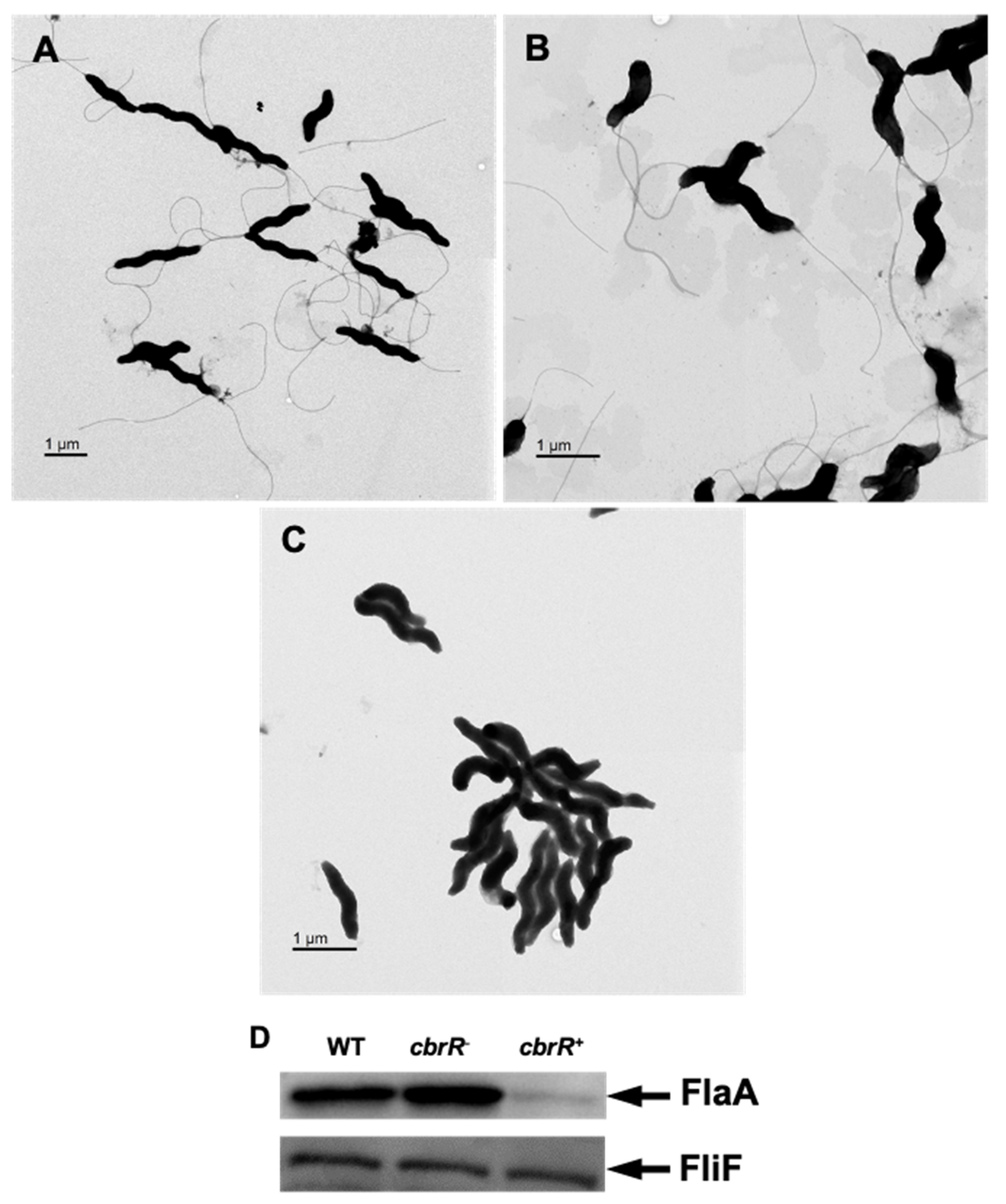
3.4. Higher Amounts of CbrR Lead to Loss of Flagellar Expression
3.5. CbrR Affects the Ability of C. jejuni to Attach to and Invade Human INT407 Cells
3.6. CbrR Negatively Regulates Biofilm Formation
3.7. Purified CbrR Is Able to Bind Both GTP and c-di-GMP
3.8. Mutation of the Autoinhibitory Site Leads to Loss of c-di-GMP Binding
3.9. Low Levels of c-di-GMP Were Detected in C. jejuni and in E. coli
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- US Department of Health and Human Services. FoodNet Fast: Pathogen Surveillance Tool. 2019. Available online: https://wwwn.cdc.gov/FoodNetFast/PathogenSurveillance.aspx (accessed on 27 January 2020).
- Mehla, K.; Ramana, J. Surface proteome mining for identification of potential vaccine candidates against Campylobacter jejuni: An in silico approach. Funct. Integr. Genom. 2016, 17, 27–37. [Google Scholar] [CrossRef]
- GBD GBDDDC. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 coun-tries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [Green Version]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Wang, N.; Ren, F.; Tang, H.; Jiao, X.; Huang, J. cj0371: A Novel Virulence-Associated Gene of Campylobacter jejuni. Front. Microbiol. 2016, 7, 1094. [Google Scholar] [CrossRef]
- Campagnolo, E.R.; Philipp, L.M.; Long, J.M.; Hanshaw, N.L. Pet-associated Campylobacteriosis: A persisting public health concern. Zoonoses Public Health 2017, 65, 304–311. [Google Scholar] [CrossRef]
- Burnham, P.M.; Hendrixson, D.R. Campylobacter jejuni: Collective components promoting a successful enteric lifestyle. Nat. Rev. Genet. 2018, 16, 551–565. [Google Scholar] [CrossRef]
- Jeon, B.; Muraoka, W.T.; Zhang, Q. Advances in Campylobacter biology and implications for biotechnological applications. Microb. Biotechnol. 2010, 3, 242–258. [Google Scholar] [CrossRef] [Green Version]
- Kovac, J.; Stessl, B.; Cadez, N.; Gruntar, I.; Cimerman, M.; Stingl, K.; Lušicky, M.; Ocepek, M.; Wagner, M.; Smole Možinaet, S. Population structure and attribution of human clinical Campylobacter jejuni isolates from central Europe to livestock and environmental sources. Zoonoses Public Health 2018, 65, 51–58. [Google Scholar] [CrossRef]
- Szczepanska, B.; Andrzejewska, M.; Spica, D.; Klawe, J.J. Prevalence and antimicrobial resistance of Campylobacter jejuni and Campylobacter coli isolated from children and environmental sources in urban and suburban areas. BMC Microbiol. 2017, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Havelaar, A.H.; Swart, A. Impact of waning acquired immunity and asymptomatic infections on case-control studies for enteric pathogens. Epidemics 2016, 17, 56–63. [Google Scholar] [CrossRef]
- Ekmekciu, I.; Fiebiger, U.; Stingl, K.; Bereswill, S.; Heimesaat, M.M. Amelioration of intestinal and systemic sequelae of murine Campylobacter jejuni infection by probiotic VSL#3 treatment. Gut Pathog. 2017, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, S.; Yuki, N. Axonal Guillain-Barré syndrome: Concepts and controversies. Lancet Neurol. 2013, 12, 1180–1188. [Google Scholar] [CrossRef]
- Hobb, R.I.; Fields, J.A.; Burns, C.M.; Thompson, S.A. Evaluation of procedures for outer membrane isolation from Campylobacter jejuni. Microbiology 2009, 155, 979–988. [Google Scholar] [CrossRef] [Green Version]
- Madan Babu, M.; Teichmann, S.A. Evolution of transcription factors and the gene regulatory network in Escherichia coli. Nucleic Acids Res. 2003, 31, 1234–1244. [Google Scholar] [CrossRef]
- Shimada, T.; Tanaka, K.; Ishihama, A. The whole set of the constitutive promoters recognized by four minor sigma subunits of Escherichia coli RNA polymerase. PLoS ONE 2017, 12, e0179181. [Google Scholar] [CrossRef] [PubMed]
- Wösten, M.M.S.M.; van Mourik, A.; van Putten, J.P.M. Regulation of Genes in Campylobacter jejuni. In Campylobacter, 3rd ed.; American Society of Microbiology: Washington, DC, USA, 2008. [Google Scholar]
- Raphael, B.H.; Pereira, S.; Flom, G.A.; Zhang, Q.; Ketley, J.M.; Konkel, M.E. The Campylobacter jejuni response regulator, CbrR, mod-ulates sodium deoxycholate resistance and chicken colonization. J. Bacteriol. 2005, 187, 3662–3670. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-H.; Köseoğlu, V.K.; Güvener, Z.T.; Myers-Morales, T.; Reed, J.M.; D’Orazio, S.E.F.; Miller, K.W.; Gomelsky, M. Cyclic di-GMP-dependent Signaling Pathways in the Pathogenic Firmicute Listeria monocytogenes. PLoS Pathog. 2014, 10, e1004301. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.; Abel, S.; Wassmann, P.; Beck, A.; Heerklotz, H.; Jenal, U. Activation of the diguanylate cyclase PleD by phosphoryla-tion-mediated dimerization. J. Biol. Chem. 2007, 282, 29170–29177. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.; Weiser, S.; Amiot, N.C.; Chan, C.; Schirmer, T.; Giese, B.; Jenal, U. Cell cycle-dependent dynamic localization of a bacterial re-sponse regulator with a novel di-guanylate cyclase output domain. Genes Dev. 2004, 18, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Sharma, I.M.; Petchiappan, A.; Chatterji, D. Quorum sensing and biofilm formation in mycobacteria: Role of c-di-GMP and methods to study this second messenger. IUBMB Life 2014, 66, 823–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Yin, K.; Qian, H.; Zhao, Y.; Wang, W.; Chou, S.-H.; Fu, Y.; He, J. Cyclic di-GMP contributes to adaption and virulence of Bacillus thuringiensis through a riboswitch-regulated collagen adhesion protein. Sci. Rep. 2016, 6, 28807. [Google Scholar] [CrossRef]
- Chou, S.H.G.N.; Lee, V.T.; Romling, U. (Eds.) Cyclic di-GMP Regulation of Gene Expression. In Microbial Cyclic di Nucleotide Signaling; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Orr, M.W.; Lee, V.T. A PilZ domain protein for chemotaxis adds another layer to c-di-GMP-mediated regulation of flagellar motility. Sci. Signal. 2016, 9, fs16. [Google Scholar] [CrossRef]
- Taveirne, M.E.; Theriot, C.M.; Livny, J.; DiRita, V.J. The Complete Campylobacter jejuni Transcriptome during Colonization of a Natural Host Determined by RNAseq. PLoS ONE 2013, 8, e73586. [Google Scholar] [CrossRef]
- Johnson, J.G.; Livny, J.; DiRita, V.J. High-Throughput Sequencing of Campylobacter jejuni Insertion Mutant Libraries Reveals mapA as a Fitness Factor for Chicken Colonization. J. Bacteriol. 2014, 196, 1958–1967. [Google Scholar] [CrossRef]
- Townsley, L.; Yildiz, F.H. Temperature affects c-di-GMP signalling and biofilm formation inVibrio cholerae. Environ. Microbiol. 2015, 17, 4290–4305. [Google Scholar] [CrossRef] [Green Version]
- Parrish, J.R.; Yu, J.; Liu, G.; Hines, J.A.; Chan, J.E.; Mangiola, B.A.; Zhang, H.; Pacifico, S.; Fotouhi, F.; DiRita, V.J.; et al. A proteome-wide protein interaction map for Campylobacter jejuni. Genome Biol. 2007, 8, R130. [Google Scholar] [CrossRef] [Green Version]
- Morooka, T.; Umeda, A.; Amako, K. Motility as an Intestinal Colonization Factor for Campylobacter jejuni. Microbiology 1985, 131, 1973–1980. [Google Scholar] [CrossRef] [Green Version]
- Hendrixson, D.R.; DiRita, V.J. Identification of Campylobacter jejuni genes involved in commensal colonization of the chick gastrointestinal tract. Mol. Microbiol. 2004, 52, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Nachamkin, I.; Yang, X.H.; Stern, N.J. Role of Campylobacter jejuni flagella as colonization factors for three-day-old chicks: Analysis with flagellar mutants. Appl. Environ. Microbiol. 1993, 59, 1269–1273. [Google Scholar] [CrossRef] [Green Version]
- Wassenaar, T.M.; Van der Zeijst, B.A.; Ayling, R.; Newell, D.G. Colonization of chicks by motility mutants of Campylobacter jejuni demonstrates the importance of flagellin A expression. J. Gen. Microbiol. 1993, 139, 1171–1175. [Google Scholar] [CrossRef] [Green Version]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Hughes, T.P.; Blaser, M.J. Experimental Campylobacter Jejuni Infection in Humans. J. Infect. Dis. 1988, 157, 472. [Google Scholar] [CrossRef]
- Dasti, J.I.; Tareen, A.M.; Lugert, R.; Zautner, A.E.; Groß, U. Campylobacter jejuni: A brief overview on pathogenicity-associated factors and disease-mediating mechanisms. Int. J. Med. Microbiol. 2010, 300, 205–211. [Google Scholar] [CrossRef]
- Krause-Gruszczynska, M.; Rohde, M.; Hartig, R.; Genth, H.; Schmidt, G.; Keo, T.; König, W.; Miller, W.G.; Konkel, M.E.; Backert, S. Role of the small Rho GTPases Rac1 and Cdc42 in host cell invasion of Campylobacter jejuni. Cell. Microbiol. 2007, 9, 2431–2444. [Google Scholar] [CrossRef]
- Kalmokoff, M.; Lanthier, P.; Tremblay, T.-L.; Foss, M.; Lau, P.C.; Sanders, G.; Austin, J.; Kelly, J.; Szymanski, C.M. Proteomic Analysis of Campylobacter jejuni 11168 Biofilms Reveals a Role for the Motility Complex in Biofilm Formation. J. Bacteriol. 2006, 188, 4312–4320. [Google Scholar] [CrossRef] [Green Version]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- Guerry, P. Campylobacter flagella: Not just for motility. Trends Microbiol. 2007, 15, 456–461. [Google Scholar] [CrossRef]
- Fauchere, J.L.; Rosenau, A.; Veron, M.; Moyen, E.N.; Richard, S.; Pfister, A. Association with HeLa cells of Campylobacter jejuni and Campylobacter coli isolated from human feces. Infect. Immun. 1986, 54, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Buswell, C.M.; Herlihy, Y.M.; Lawrence, L.M.; McGuiggan, J.T.; Marsh, P.D.; Keevil, C.W.; Leach, S.A. Extended survival and persistence of Campylobacter spp. in water and aquatic biofilms and their detection by immunofluorescent-antibody and -rRNA staining. Appl Environ. Microbiol. 1998, 64, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.; Oh, E.; Jeon, B. Enhanced Transmission of Antibiotic Resistance in Campylobacter jejuni Biofilms by Natural Transformation. Antimicrob. Agents Chemother. 2014, 58, 7573–7575. [Google Scholar] [CrossRef] [Green Version]
- Reuter, M.; Mallett, A.; Pearson, B.M.; van Vliet, A.H. Biofilm formation by Campylobacter jejuni is increased under aerobic condi-tions. Appl. Environ. Microbiol. 2010, 76, 2122–2128. [Google Scholar] [CrossRef] [Green Version]
- Fux, C.A.; Costerton, J.W.; Stewart, P.S.; Stoodley, P. Survival strategies of infectious biofilms. Trends Microbiol. 2005, 13, 34–40. [Google Scholar] [CrossRef]
- Zimmer, M.; Barnhart, H.; Idris, U.; Lee, M.D. Detection of Campylobacter jejuni Strains in the Water Lines of a Commercial Broiler House and Their Relationship to the Strains That Colonized the Chickens. Avian Dis. 2003, 47, 101–107. [Google Scholar] [CrossRef]
- Haddock, G.; Mullin, M.; MacCallum, A.; Sherry, A.; Tetley, L.; Watson, E.; Dagleish, M.; Smith, D.G.E.; Everest, P. Campylobacter jejuni 81-176 forms distinct micro-colonies on in vitro-infected human small intestinal tissue prior to biofilm formation. Microbiology 2010, 156, 3079–3084. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-S.; Park, C.; Kim, Y.-J. Role of flgA for Flagellar Biosynthesis and Biofilm Formation of Campylobacter jejuni NCTC11168. J. Microbiol. Biotechnol. 2015, 25, 1871–1879. [Google Scholar] [CrossRef]
- Reeser, R.J.; Medler, R.T.; Billington, S.J.; Jost, B.H.; Joens, L.A. Characterization of Campylobacter jejuni Biofilms under Defined Growth Conditions. Appl. Environ. Microbiol. 2007, 73, 1908–1913. [Google Scholar] [CrossRef] [Green Version]
- Moe, K.K.; Mimura, J.; Ohnishi, T.; Wake, T.; Yamazaki, W.; Nakai, M.; Misawa, N. The Mode of Biofilm Formation on Smooth Surfaces by Campylobacter jejuni. J. Vet. Med. Sci. 2010, 72, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Crouzet, M.; Le Senechal, C.; Brözel, V.S.; Costaglioli, P.; Barthe, C.; Bonneu, M.; Garbay, B.; Vilain, S. Exploring early steps in biofilm formation: Set-up of an experimental system for molecular studies. BMC Microbiol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rathbun, K.M.; Hall, J.E.; Thompson, S.A. Cj0596 is a periplasmic peptidyl prolyl cis-trans isomerase involved in Campylobacter jejuni motility, invasion, and colonization. BMC Microbiol. 2009, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Dailidiene, D.; Dailide, G.; Kersulyte, D.; Berg, D.E. Contraselectable streptomycin susceptibility determinant for genetic manipu-lation and analysis of Helicobacter pylori. Appl Environ. Microbiol. 2006, 72, 5908–5914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.F.; Dower, W.J.; Tompkins, L.S. High-voltage electroporation of bacteria: Genetic transformation of Campylobacter jejuni with plasmid DNA. Proc. Natl. Acad. Sci. USA 1988, 85, 856–860. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Alm, R.A.; Trust, T.J.; Guerry, P. Construction of new Campylobacter cloning vectors and a new mutational cat cassette. Gene 1993, 130, 127–130. [Google Scholar] [CrossRef]
- Wang, Y.E.; Taylor, D. Natural transformation in Campylobacter species. J. Bacteriol. 1990, 172, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Misawa, N.; Blaser, M.J. Detection and Characterization of Autoagglutination Activity by Campylobacter jejuni. Infect. Immun. 2000, 68, 6168–6175. [Google Scholar] [CrossRef] [PubMed]
- Fields, J.A.; Thompson, S.A. Campylobacter jejuni CsrA Mediates Oxidative Stress Responses, Biofilm Formation, and Host Cell Invasion. J. Bacteriol. 2008, 190, 3411–3416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelofs, K.G.; Wang, J.; Sintim, H.O.; Lee, V.T. Differential radial capillary action of ligand assay for high-throughput detection of protein-metabolite interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 15528–15533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, I.H.; Bagnall, M.C.; Browning, D.D.; Thompson, S.A.; Manning, G.; Newell, D.G. Gamma-glutamyl transpeptidase has a role in the persistent colonization of the avian gut by Campylobacter jejuni. Microb. Pathog. 2007, 43, 198–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massie, J.P.; Reynolds, E.L.; Koestler, B.J.; Cong, J.P.; Agostoni, M.; Waters, C.M. Quantification of high-specificity cyclic diguanylate signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 12746–12751. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Nikolskaya, A.N.; Koonin, E.V. Novel domains of the prokaryotic two-component signal transduction systems. FEMS Microbiol. Lett. 2001, 203, 11–21. [Google Scholar] [CrossRef]
- Schirmer, T. C-di-GMP Synthesis: Structural Aspects of Evolution, Catalysis and Regulation. J. Mol. Biol. 2016, 428, 3683–3701. [Google Scholar] [CrossRef] [Green Version]
- Dahlstrom, K.M.; Giglio, K.M.; Sondermann, H.; O’Toole, G.A. The Inhibitory Site of a Diguanylate Cyclase Is a Necessary Element for Interaction and Signaling with an Effector Protein. J. Bacteriol. 2016, 198, 1595–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldridge, P.; Paul, R.; Goymer, P.; Rainey, P.; Jenal, U. Role of the GGDEF regulator PleD in polar development of Caulobacter crescentus. Mol. Microbiol. 2003, 47, 1695–1708. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.; Paul, R.; Samoray, D.; Amiot, N.C.; Giese, B.; Jenal, U.; Schirmer, T. Structural basis of activity and allosteric control of diguanylate cyclase. Proc. Natl. Acad. Sci. USA 2004, 101, 17084–17089. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capra, J.A.; Singh, M. Predicting functionally important residues from sequence conservation. Bioinformatics 2007, 23, 1875–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugar, G.; Herbig, A.; Förstner, K.U.; Heidrich, N.; Reinhardt, R.; Nieselt, K.; Sharma, C.M. High-Resolution Transcriptome Maps Reveal Strain-Specific Regulatory Features of Multiple Campylobacter jejuni Isolates. PLoS Genet. 2013, 9, e1003495. [Google Scholar] [CrossRef]
- Karlyshev, A.V.; Wren, B.W. Development and application of an insertional system for gene delivery and expression in Cam-pylobacter jejuni. Appl. Environ. Microbiol. 2005, 71, 4004–4013. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.J.; Cagliero, C.; Zhou, Y.N. Growth rate regulation in Escherichia coli. FEMS Microbiol. Rev. 2012, 36, 269–287. [Google Scholar] [CrossRef] [Green Version]
- Römling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The First 25 Years of a Universal Bacterial Second Messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef] [Green Version]
- Nesper, J.; Hug, I.; Kato, S.; Hee, C.S.; Habazettl, J.M.; Manfredi, P.; Grzesiek, S.; Schirmer, T.; Emonet, T.; Jenal, U. Cyclic di-GMP differentially tunes a bacterial flagellar motor through a novel class of CheY-like regulators. Elife 2017, 6, e28842. [Google Scholar] [CrossRef]
- Crushell, E.; Harty, S.; Sharif, F.; Bourke, B. Enteric campylobacter: Purging its secrets? Pediatr Res. 2004, 55, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a Foodborne Pathogen: A Review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekhar, K.; Kassem, I.I.; Rajashekara, G. Campylobacter jejuni transducer like proteins: Chemotaxis and beyond. Gut Microbes. 2017, 8, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekhar, K.; Gangaiah, D.; Pina-Mimbela, R.; Kassem, I.I.; Jeon, B.H.; Rajashekara, G. Transducer like proteins of Campylobacter jejuni 81-176: Role in chemotaxis and colonization of the chicken gastrointestinal tract. Front. Cell. Infect. Microbiol. 2015, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekhar, K.; Srivastava, V.; Hwang, S.; Jeon, B.; Ryu, S.; Rajashekara, G. Transducer-Like Protein in Campylobacter jejuni with a Role in Mediating Chemotaxis to Iron and Phosphate. Front. Microbiol. 2018, 9, 2674. [Google Scholar] [CrossRef]
- Marchant, J.; Wren, B.; Ketley, J. Exploiting genome sequence: Predictions for mechanisms of Campylobacter chemotaxis. Trends Microbiol. 2002, 10, 155–159. [Google Scholar] [CrossRef]
- Rahman, H.; King, R.M.; Shewell, L.K.; Semchenko, E.A.; Hartley-Tassell, L.E.; Wilson, J.C.; Day, C.; Korolik, V. Characterisation of a Multi-ligand Binding Chemoreceptor CcmL (Tlp3) of Campylobacter jejuni. PLoS Pathog. 2014, 10, e1003822. [Google Scholar] [CrossRef] [Green Version]
- Burrough, E.R.; Wu, Z.; Sahin, O.; Zhang, Q.; Yaeger, M.J. Spatial Distribution of Putative Growth Factors in the Guinea Pig Placenta and the Effects of These Factors, Plasma, and Bile on the Growth and Chemotaxis of Campylobacter jejuni. Vet. Pathol. 2012, 49, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Vegge, C.S.; Brøndsted, L.; Li, Y.-P.; Bang, D.D.; Ingmer, H. Energy Taxis Drives Campylobacter jejuni toward the Most Favorable Conditions for Growth. Appl. Environ. Microbiol. 2009, 75, 5308–5314. [Google Scholar] [CrossRef] [Green Version]
- Hugdahl, M.B.; Beery, J.T.; Doyle, M.P. Chemotactic behavior of Campylobacter jejuni. Infect. Immun. 1988, 56, 1560–1566. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Sarenko, O.; Klauck, G.; Wilke, F.M.; Pfiffer, V.; Richter, A.M.; Herbst, S.; Kaever, V.; Hengge, R. More than Enzymes That Make or Break Cyclic Di-GMP—Local Signaling in the Interactome of GGDEF/EAL Domain Proteins of Escherichia coli. MBio 2017, 8, e01639-17. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.E.; Webster, S.S.; Diepold, A.; Kuchma, S.L.; Bordeleau, E.; Armitage, J.P.; O’Toole, G.A. Flagellar Stators Stimulate c-di-GMP Production by Pseudomonas aeruginosa. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontaine, B.M.; Duggal, Y.; Weinert, E.E. Exploring the Links between Nucleotide Signaling and Quorum Sensing Pathways in Regulating Bacterial Virulence. ACS Infect. Dis. 2018, 4, 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Hengge, R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 2009, 7, 263–273. [Google Scholar] [CrossRef]
- Navarro, M.V.A.S.; De, N.; Bae, N.; Wang, Q.; Sondermann, H. Structural Analysis of the GGDEF-EAL Domain-Containing c-di-GMP Receptor FimX. Structure 2009, 17, 1104–1116. [Google Scholar] [CrossRef] [Green Version]
- Karaolis, D.K.R.; Rashid, M.H.; Chythanya, R.; Luo, W.; Hyodo, M.; Hayakawa, Y. c-di-GMP (3′-5′-Cyclic Diguanylic Acid) Inhibits Staphylococcus aureus Cell-Cell Interactions and Biofilm Formation. Antimicrob. Agents Chemother. 2005, 49, 1029–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.; Qu, T.; Zhao, H.; Su, L.; Yu, Q.; Gao, J.; Wu, B. The effect of c-di-GMP (3′–5′-cyclic diguanylic acid) on the biofilm formation and adherence of Streptococcus mutans. Microbiol. Res. 2010, 165, 87–96. [Google Scholar] [CrossRef]
- Brouillette, E.; Hyodo, M.; Hayakawa, Y.; Karaolis, D.K.; Malouin, F. 3',5'-cyclic diguanylic acid reduces the virulence of bio-film-forming Staphylococcus aureus strains in a mouse model of mastitis infection. Antimicrob. Agents Chemother. 2005, 49, 3109–3113. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | c-di-GMP (nM) in Cellular Extracts |
|---|---|
| WT Plate | 0.4 |
| cbrR− Plate | 0 |
| cbrR+ Plate | 0 |
| WT Biphasic | 0 |
| cbrR− Biphasic | 0 |
| cbrR+ Biphasic | 0 |
| WT Broth | 0 |
| cbrR− Broth | 0 |
| cbrR+ Broth | 0 |
| QrgB (pCMW75) 4 h expression | 30.6 |
| Inactive QrgB (pCMW98) 4 h expression | 0 |
| WT CbrR (pCAF107) 4 h expression | 3.4 |
| Active site mutant CbrR (pCAF109) 4 h expression | 27.6 |
| I-site mutant CbrR (pCAF110) 4 h expression | 0 |
| QrgB (pCMW75) O/N expression | 816.0 |
| Inactive QrgB (pCMW98) O/N expression | 27.4 |
| WT CbrR (pCAF107) O/N expression | 10.9 |
| Active site mutant CbrR (pCAF109) O/N expression | 6.8 |
| I-site mutant CbrR (pCAF110) O/N expression | 4.0 |
| c-di-GMP Standard (1.953 nM) | 3.3 |
| c-di-GMP Standard (3.906 nM) | 4.2 |
| c-di-GMP Standard (7.813 nM) | 7.1 |
| c-di-GMP Standard (15.625 nM) | 15.6 |
| c-di-GMP Standard (31.250 nM) | 28.5 |
| c-di-GMP Standard (62.500 nM) | 63.8 |
| c-di-GMP Standard (125.000 nM) | 125.4 |
| Blank | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cox, C.A.; Bogacz, M.; El Abbar, F.M.; Browning, D.D.; Hsueh, B.Y.; Waters, C.M.; Lee, V.T.; Thompson, S.A. The Campylobacter jejuni Response Regulator and Cyclic-Di-GMP Binding CbrR Is a Novel Regulator of Flagellar Motility. Microorganisms 2022, 10, 86. https://doi.org/10.3390/microorganisms10010086
Cox CA, Bogacz M, El Abbar FM, Browning DD, Hsueh BY, Waters CM, Lee VT, Thompson SA. The Campylobacter jejuni Response Regulator and Cyclic-Di-GMP Binding CbrR Is a Novel Regulator of Flagellar Motility. Microorganisms. 2022; 10(1):86. https://doi.org/10.3390/microorganisms10010086
Chicago/Turabian StyleCox, Claudia A., Marek Bogacz, Faiha M. El Abbar, Darren D. Browning, Brian Y. Hsueh, Chris M. Waters, Vincent T. Lee, and Stuart A. Thompson. 2022. "The Campylobacter jejuni Response Regulator and Cyclic-Di-GMP Binding CbrR Is a Novel Regulator of Flagellar Motility" Microorganisms 10, no. 1: 86. https://doi.org/10.3390/microorganisms10010086
APA StyleCox, C. A., Bogacz, M., El Abbar, F. M., Browning, D. D., Hsueh, B. Y., Waters, C. M., Lee, V. T., & Thompson, S. A. (2022). The Campylobacter jejuni Response Regulator and Cyclic-Di-GMP Binding CbrR Is a Novel Regulator of Flagellar Motility. Microorganisms, 10(1), 86. https://doi.org/10.3390/microorganisms10010086