
Comparative Whole-Genome Analysis of Russian Foodborne Multidrug-Resistant Salmonella Infantis Isolates

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Isolation, Identification and Antibiotic Susceptibility Testing
2.2. DNA Isolation and Whole Genome Sequencing
2.3. Genome Assembly and Sequence Analysis
2.4. CRISPR Typing and Analysis
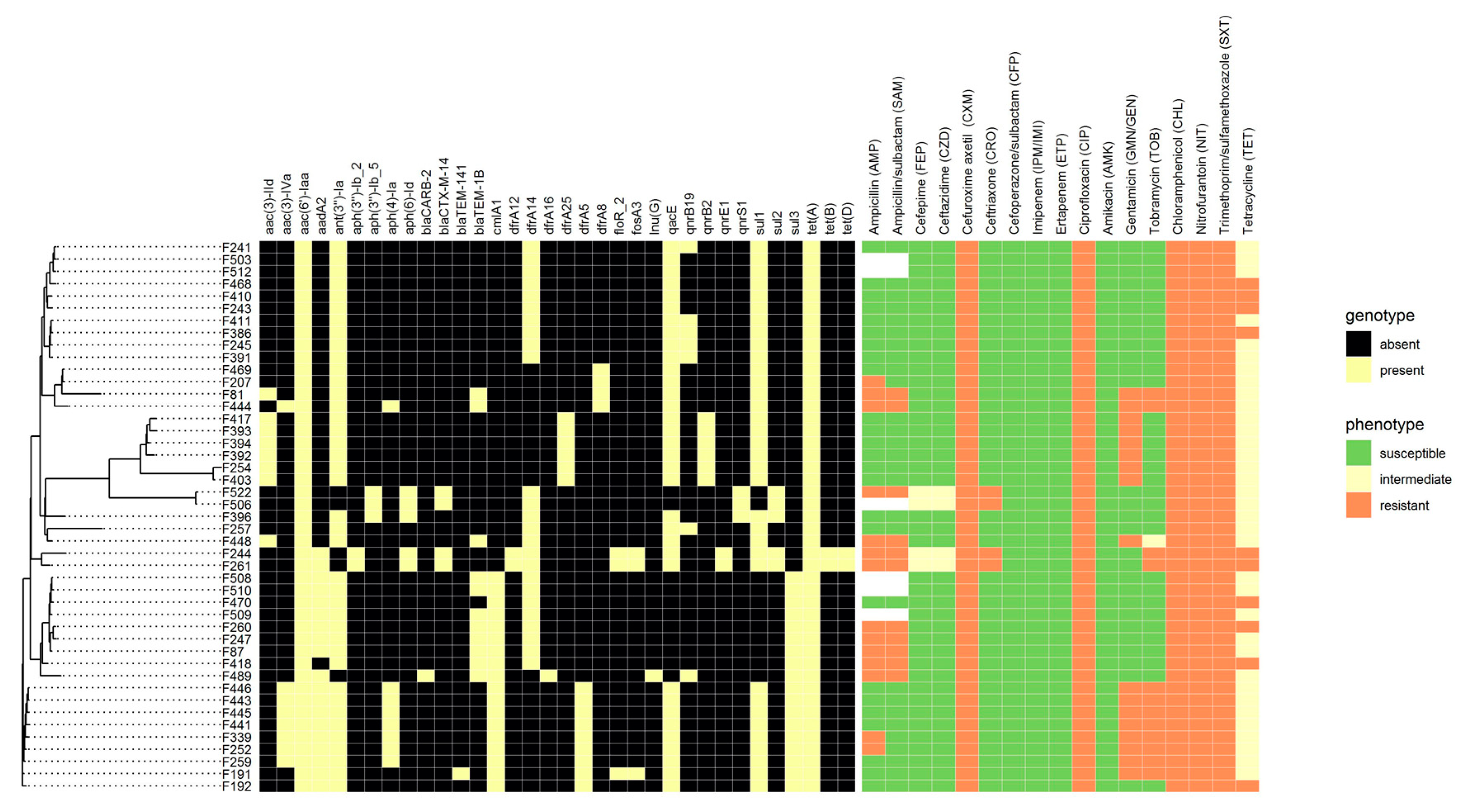
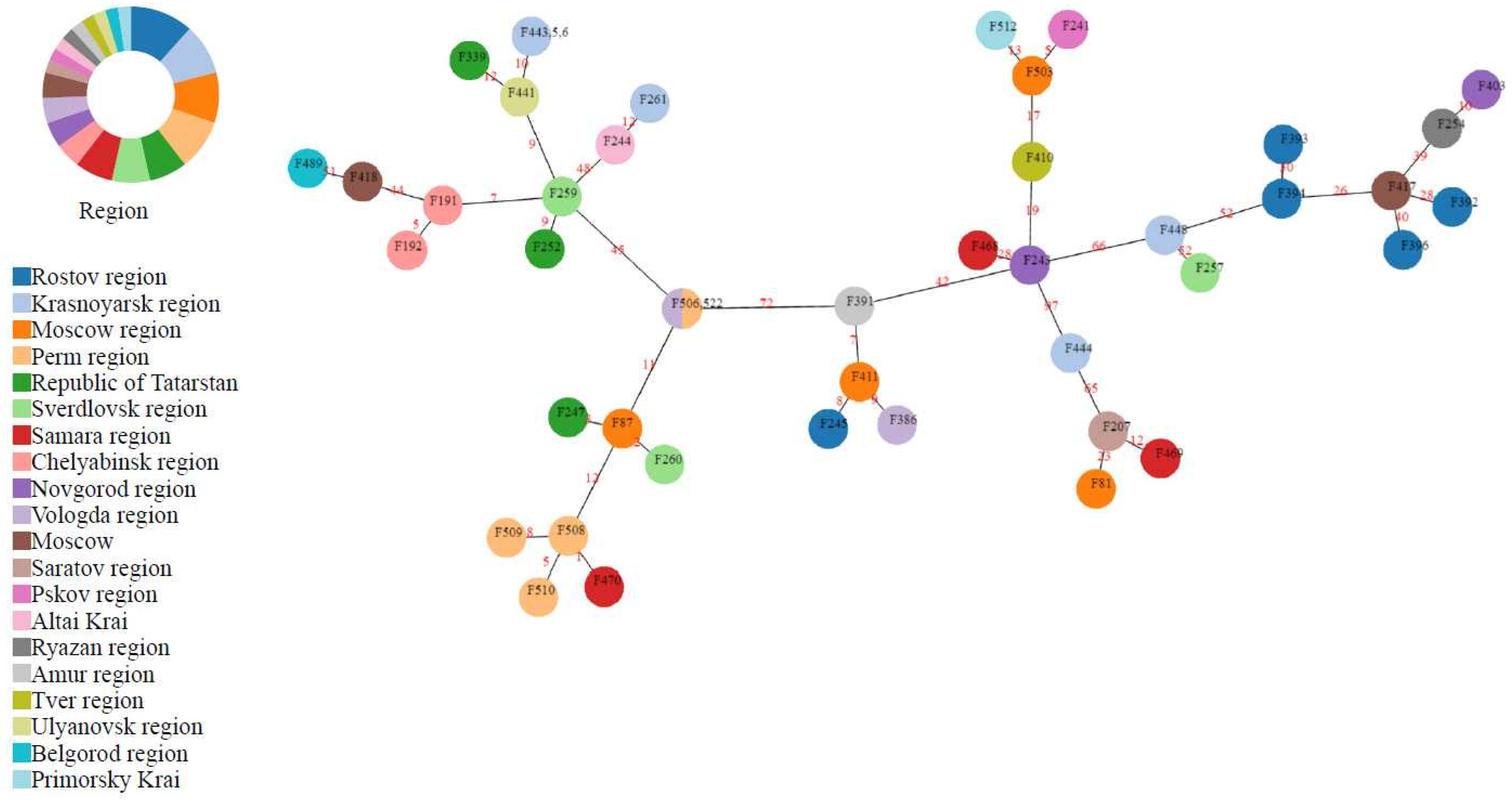
3. Results
3.1. Isolation and Typing
3.2. Antibiotic Resistance Determination
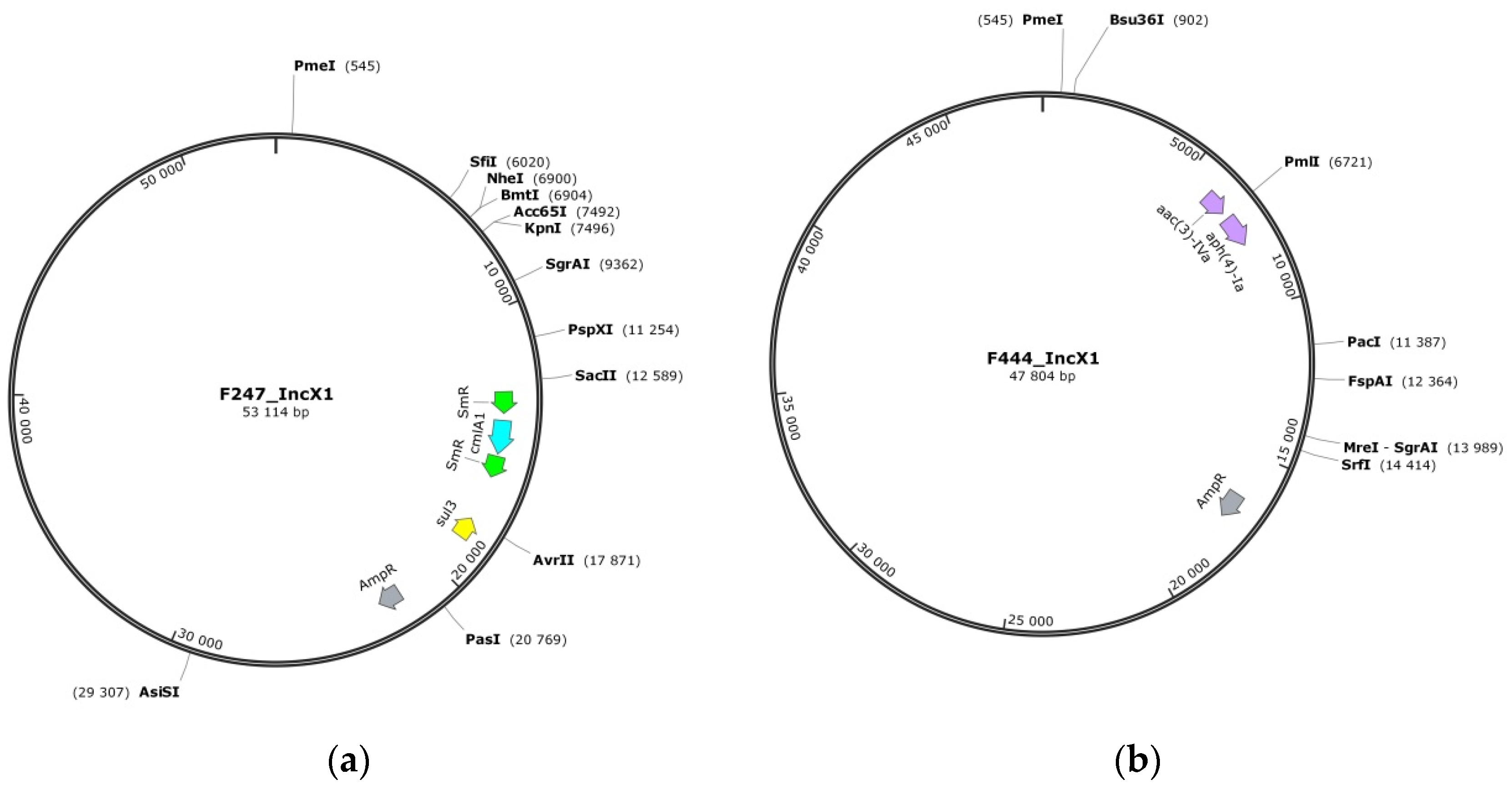
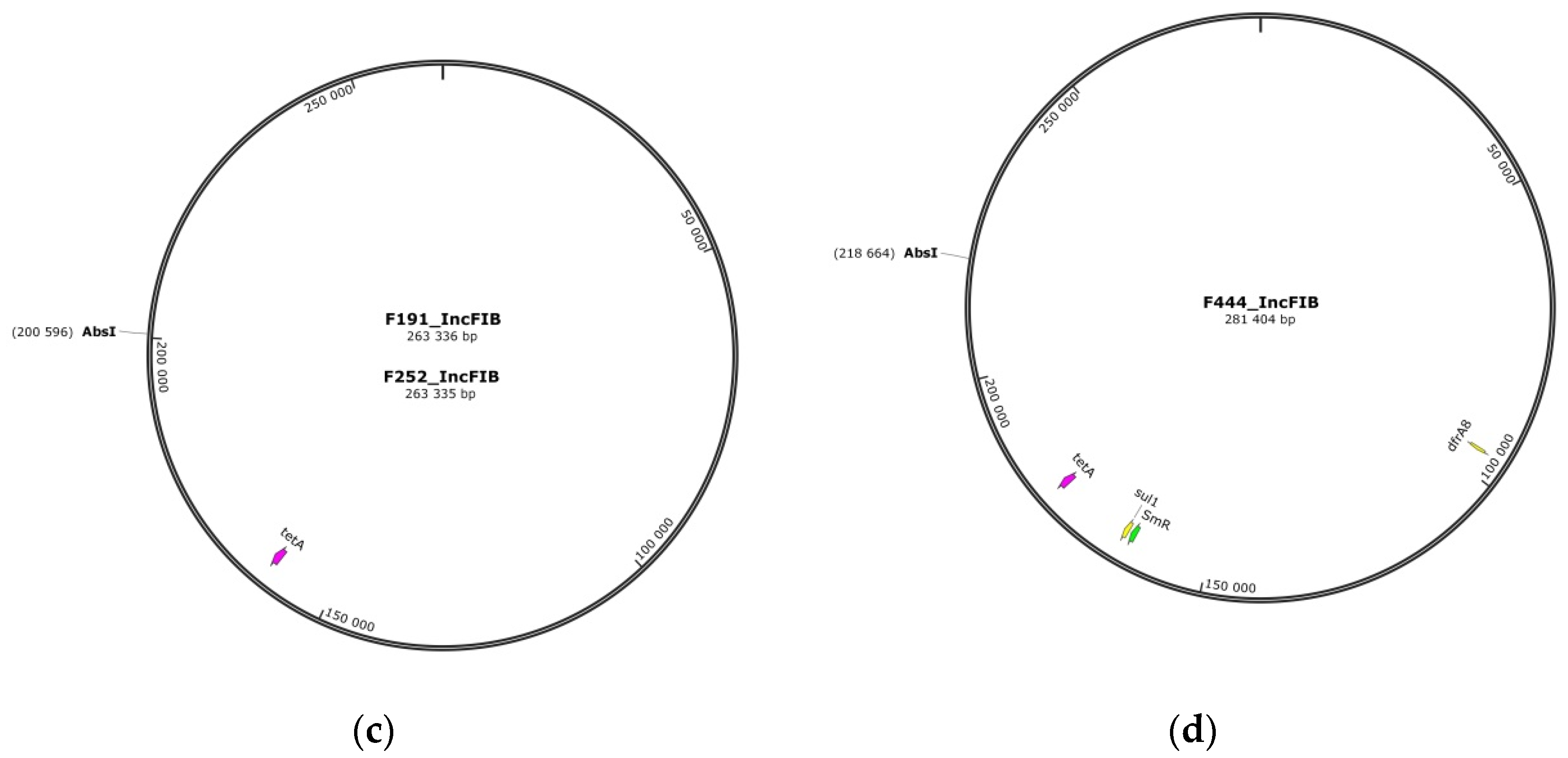
3.3. Plasmids
3.4. Virulence Genes
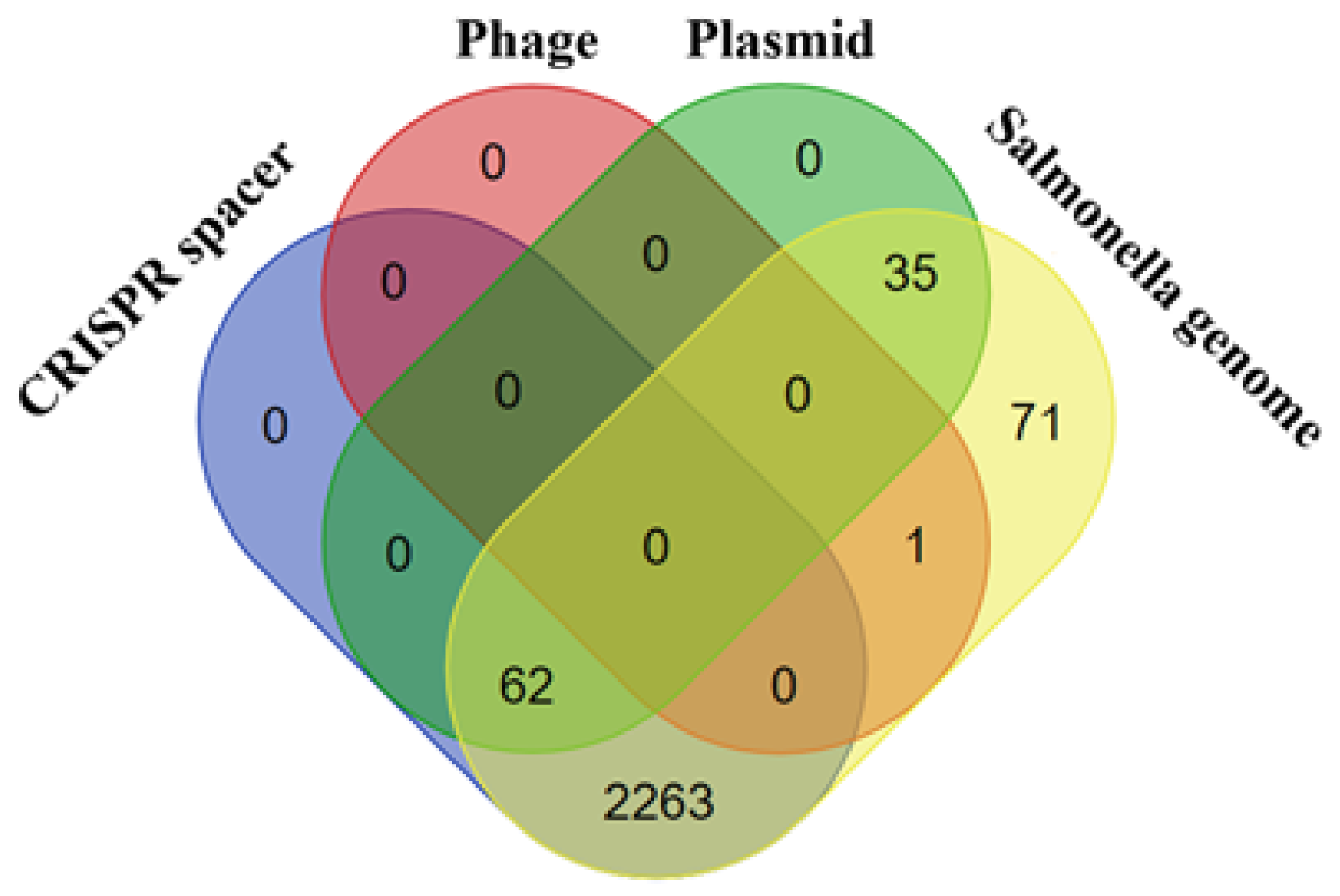
3.5. CRISPR/Cas Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coburn, B.; Grassl, G.A.; Finlay, B.B. Salmonella, the host and disease: A brief review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef]
- Santos, R.L.; Tsolis, R.M.; Bäumler, A.J.; Adams, L.G. Pathogenesis of Salmonella-induced enteritis. Braz. J. Med. Biol. Res. 2003, 36, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, C.G.; Marcus, S.L.; Steele-Mortimer, O.; Knodler, L.A.; Finlay, B.B. Salmonella Typhimurium virulence genes are induced upon bacterial invasion into phagocytic and nonphagocytic cells. Infect. Immun. 1999, 67, 5690–5698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langridge, G.C.; Fookesa, M.; Connora, T.R.; Feltwella, T.; Feaseya, N.; Parsonsb, B.N.; Seth-Smitha, H.M.B.; Barquista, L.; Stedmana, A.; Humphreyd, T.; et al. Patterns of genome evolution that have accompanied host adaptation in Salmonella. Proc. Natl. Acad. Sci. USA 2015, 112, 863–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Ngoi, S.T.; Ju Teh, C.S.; Chai, L.C.; Thong, K.L. Overview of Molecular Typing Tools for the Characterization of Salmonella enterica in Malaysia. Biomed. Environ. Sci. 2015, 28, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Ben Hassena, A.; Haendiges, J.; Zormati, S.; Guermazi, S.; Gdoura, R.; Gonzalez-Escalona, N.; Siala, M. Virulence and resistance genes profiles and clonal relationships of non-typhoidal food-borne Salmonella strains isolated in Tunisia by whole genome sequencing. Int. J. Food Microbiol. 2021, 337, 108941. [Google Scholar] [CrossRef] [PubMed]
- Medalla, F.; Gu, W.; Mahon, B.E.; Judd, M.; Folster, J.; Griffin, P.M.; Hoekstra, R.M. Estimated incidence of antimicrobial drug-resistant nontyphoidal Salmonella infections, United States, 2004–2012. Emerg. Infect. Dis. 2017, 23, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nüesch-Inderbinen, M. Salmonella enterica serovar Infantis from food and human infections, Switzerland, 2010-2015: Poultry-related multidrug resistant clones and an emerging ESBL producing clonal lineage. Front. Microbiol. 2017, 8, 1322. [Google Scholar] [CrossRef] [Green Version]
- ECDC Annual Epidemiological Report 2016 [2014 data]. EFSA J. 2016, 2016–2017. Available online: https://www.ecdc.europa.eu/en/publications-data/malaria-annual-epidemiological-report-2016 (accessed on 29 November 2021).
- Rozhnova, S.S.; Kuleshov, K.V.; Pavlova, A.S.; Guseva, A.N.; Kozhakhmetova, T.A.; Akulova, N.K.; Podkolzin, A.T. Heterogeneity of Salmonella isolates obtained from various sources in Russia 2010–2019. Epidemiol. Infect. Dis. 2020, 25, 26–34. [Google Scholar] [CrossRef]
- Federal Service for Consumer Protection and Human Welfare Federal Report “On the State of the Sanitary-Epidemiological Well-Being of the Population in the Russian Federation in 2018.” 2019, 254. Available online: https://www.rospotrebnadzor.ru/documents/details.php?ELEMENT_ID=12053 (accessed on 4 June 2019).
- Rozhnova, S.H.; Akulova, N.K.; Khristukhina, O.A. Prospects for the organization of an expanded surveillance system for salmonellosis in Russia. Epidemiol. Vaccine 2015, 14, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Bogomazova, A.N.; Gordeeva, V.D.; Krylova, E.V.; Soltynskaya, I.V.; Davydova, E.E.; Ivanova, O.E.; Komarov, A.A. Mega-plasmid found worldwide confers multiple antimicrobial resistance in Salmonella Infantis of broiler origin in Russia. Int. J. Food Microbiol. 2020, 319, 108497. [Google Scholar] [CrossRef]
- Acar, S.; Bulut, E.; Durul, B.; Uner, I.; Kur, M.; Avsaroglu, M.D.; Kirmaci, H.A.; Tel, Y.O.; Zeyrek, F.Y.; Soyer, Y. Phenotyping and genetic characterization of Salmonella enterica isolates from Turkey revealing arise of different features specific to geography. Int. J. Food Microbiol. 2017, 241, 98–107. [Google Scholar] [CrossRef]
- Aviv, G.; Tsyba, K.; Steck, N.; Salmon-Divon, M.; Cornelius, A.; Rahav, G.; Grassl, G.A.; Gal-Mor, O. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ. Microbiol. 2014, 16, 977–994. [Google Scholar] [CrossRef] [PubMed]
- The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2017/2018. EFSA J. 2020, 18. Available online: https://efsa.onlinelibrary.wiley.com/doi/full/10.2903/j.efsa.2020.6007 (accessed on 29 November 2021).
- García-Soto, S.; Abdel-Glil, M.Y.; Tomaso, H.; Linde, J.; Methner, U. Emergence of Multidrug-Resistant Salmonella enterica Subspecies enterica Serovar Infantis of Multilocus Sequence Type 2283 in German Broiler Farms. Front. Microbiol. 2020, 11, 1741. [Google Scholar] [CrossRef]
- Karacan Sever, N.; Akan, M. Molecular analysis of virulence genes of Salmonella Infantis isolated from chickens and turkeys. Microb. Pathog. 2019, 126, 199–204. [Google Scholar] [CrossRef]
- Kudaka, J.; Itokazu, K.; Taira, K.; Iwai, A.; Kondo, M.; Susa, T.; Iwanaga, M. Characterization of Salmonella Isolated in Okinawa, Japan. Jpn. J. Infect. Dis 2006, 59, 15–19. [Google Scholar]
- Kürekci, C.; Sahin, S.; Iwan, E.; Kwit, R.; Bomba, A.; Wasyl, D. Whole-genome sequence analysis of Salmonella Infantis isolated from raw chicken meat samples and insights into pESI-like megaplasmid. Int. J. Food Microbiol. 2021, 337, 108956. [Google Scholar] [CrossRef]
- McDermott, P.F.; Tyson, G.H.; Kabera, C.; Chen, Y.; Li, C.; Folster, J.P.; Ayers, S.L.; Lam, C.; Tate, H.P.; Zhao, S. Whole-genome sequencing for detecting antimicrobial resistance in nontyphoidal Salmonella. Antimicrob. Agents Chemother. 2016, 60, 5515–5520. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.M.; White, D.G.; McDermott, P.F. The US National Antimicrobial Resistance Monitoring System. Future Microbiol. 2007, 2, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; White, D.G.; McDermott, P.F.; Walker, R.D.; Rhodes, B.; Fedorka-Cray, P.J.; Simjee, S.; Zhao, S. Comparison of subtyping methods for differentiating Salmonella enterica serovar Typhimurium isolates obtained from food animal sources. J. Clin. Microbiol. 2006, 44, 3569–3577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punina, N.V.; Makridakis, N.M.; Remnev, M.A.; Topunov, A.F. Whole-genome sequencing targets drug-resistant bacterial infections. Hum. Genom. 2015, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Egorova, S.; Kuleshov, K.; Kaftyreva, L. Modern Salmonella subtyping methods in outbreak investigations. Immunopathol. Allergol. Infectology 2019, 36–42. [Google Scholar] [CrossRef]
- Ashton, P.M.; Nair, S.; Dallman, T.; Rubino, S.; Rabsch, W.; Mwaigwisya, S.; Wain, J.; O’Grady, J. MinION nanopore sequencing identifies the position and structure of a bacterial antibiotic resistance island. Nat. Biotechnol. 2015, 33, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Madoui, M.A.; Engelen, S.; Cruaud, C.; Belser, C.; Bertrand, L.; Alberti, A.; Lemainque, A.; Wincker, P.; Aury, J.M. Genome assembly using Nanopore-guided long and error-free DNA reads. BMC Genom. 2015, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Parry, C.M.; Threlfall, E.J. Antimicrobial resistance in typhoidal and nontyphoidal Salmonellae. Curr. Opin. Infect. Dis. 2008, 21, 531–538. [Google Scholar] [CrossRef]
- Achtman, M.; Wain, J.; Weill, F.X.; Nair, S.; Zhou, Z.; Sangal, V.; Krauland, M.G.; Hale, J.L.; Harbottle, H.; Uesbeck, A.; et al. Multilocus Sequence Typing as a Replacement for Serotyping in Salmonella enterica. PLOS Pathog. 2012, 8, e1002776. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carriço, J.A.; Vaz, C. PHYLOViZ 2.0: Providing scalable data integration and visualization for multiple phylogenetic inference methods. Bioinformatics 2017, 33, 128–129. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Zhou, Z.; Sergeant, M.J.; Achtman, M. A genomic overview of the population structure of Salmonella. PLOS Genet. 2018, 14, e1007261. [Google Scholar] [CrossRef] [Green Version]
- Dolejska, M.; Villa, L.; Minoia, M.; Guardabassi, L.; Carattoli, A. Complete sequences of IncHI1 plasmids carrying blaCTX-M-1 and qnrS1 in equine Escherichia coli provide new insights into plasmid evolution. J. Antimicrob. Chemother. 2014, 69, 2388–2393. [Google Scholar] [CrossRef] [Green Version]
- Dobiasova, H.; Dolejska, M. Prevalence and diversity of IncX plasmids carrying fluoroquinolone and β-lactam resistance genes in Escherichia coli originating from diverse sources and geographical areas. J. Antimicrob. Chemother. 2016, 71, 2118–2124. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, T.; Dashti, A.A.; Albaksami, O.; Udo, E.E.; Jadaon, M.M.; Albert, M.J. Ciprofloxacin-resistant Salmonella enterica serovar Typhi from Kuwait with novel mutations in gyrA and parC genes. J. Clin. Microbiol. 2009, 47, 208–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, S.L.; Brumell, J.H.; Pfeifer, C.G.; Finlay, B.B. Salmonella pathogenicity islands: Big virulence in small packages. Microbes Infect. 2000, 2, 145–156. [Google Scholar] [CrossRef]
- Riquelme, S.; Varas, M.; Valenzuela, C.; Velozo, P.; Chahin, N.; Aguilera, P.; Sabag, A.; Labra, B.; Álvarez, S.A.; Chávez, F.P.; et al. Relevant genes linked to virulence are required for Salmonella Typhimurium to survive intracellularly in the social amoeba Dictyostelium discoideum. Front. Microbiol. 2016, 7, 1305. [Google Scholar] [CrossRef] [PubMed]
- Kombade, S.; Kaur, N. Pathogenicity Island in Salmonella. In Salmonella-a Challenge From Farm to Fork; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Bugarel, M.; den Bakker, H.; Grout, J.; Vignaud, M.L.; Loneragan, G.H.; Fach, P.; Brisabois, A. CRISPR-based assay for the molecular identification of highly prevalent Salmonella serotypes. Food Microbiol. 2018, 71, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Bratčikov, M.; Mauricas, M. The use of high-resolution melting analysis for Salmonella spp. CRISPR sequence genotyping. Acta Med. Litu. 2009, 16, 98–102. [Google Scholar] [CrossRef]
- Lindqvist, N.; Pelkonen, S. Genetic surveillance of endemic bovine Salmonella Infantis infection. Acta Vet. Scand. 2007, 49, 1–9. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Nógrády, N.; Király, M.; Davies, R.; Nagy, B. Multidrug resistant clones of Salmonella Infantis of broiler origin in Europe. Int. J. Food Microbiol. 2012, 157, 108–112. [Google Scholar] [CrossRef]
- Sakano, C.; Kuroda, M.; Sekizuka, T.; Ishioka, T.; Morita, Y.; Ryo, A.; Tsukagoshi, H.; Kawai, Y.; Inoue, N.; Takada, H.; et al. Genetic analysis of non-hydrogen sulfide-producing Salmonella enterica serovar Typhimurium and S. enterica Serovar Infantis isolates in Japan. J. Clin. Microbiol. 2013, 51, 328–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Kuang, D.; Xu, X.; González-Escalona, N.; Erickson, D.L.; Brown, E.; Meng, J. Genomic analyses of multidrug-resistant Salmonella Indiana, Typhimurium, and Enteritidis isolates using MinION and MiSeq sequencing technologies. PLoS ONE 2020, 15, e0235641. [Google Scholar] [CrossRef]
- Shelenkov, A.; Mikhaylova, Y.; Yanushevich, Y.; Samoilov, A.; Petrova, L.; Fomina, V.; Gusarov, V.; Zamyatin, M.; Shagin, D.; Akimkin, V. Molecular typing, characterization of antimicrobial resistance, virulence profiling and analysis of whole-genome sequence of clinical Klebsiella pneumoniae isolates. Antibiotics 2020, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Villa, L.; García-Fernández, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Fernández, A.; Chiaretto, G.; Bertini, A.; Villa, L.; Fortini, D.; Ricci, A.; Carattoli, A. Multilocus sequence typing of IncI1 plasmids carrying extended-spectrum β-lactamases in Escherichia coli and Salmonella of human and animal origin. J. Antimicrob. Chemother. 2008, 61, 1229–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Bielak, E.M.; Fortini, D.; Hansen, L.H.; Hasman, H.; Debroy, C.; Nolan, L.K.; Carattoli, A. Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 2012, 68, 43–50. [Google Scholar] [CrossRef]
- Ahmed, D.; Nahid, M.A.; Sami, A.B.; Halim, F.; Akter, N.; Sadique, T.; Rana, M.S.; Elahi, M.S.B.; Rahman, M.M. Bacterial etiology of bloodstream infections and antimicrobial resistance in Dhaka, Bangladesh, 2005–2014. Antimicrob. Resist. Infect. Control 2017, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Cheng, H.; Liang, Y.; Yu, S.; Yu, T.; Fang, J.; Zhu, C. Diverse Mobile Genetic Elements and Conjugal Transferability of Sulfonamide Resistance Genes (sul1, sul2, and sul3) in Escherichia coli Isolates From Penaeus vannamei and Pork From Large Markets in Zhejiang, China. Front. Microbiol. 2019, 10, 1787. [Google Scholar] [CrossRef] [Green Version]
- Conner, C.P.; Heithoff, D.M.; Julio, S.M.; Sinsheimer, R.L.; Mahan, M.J. Differential patterns of acquired virulence genes distinguish Salmonella strains. Proc. Natl. Acad. Sci. USA 1998, 95, 4641. [Google Scholar] [CrossRef] [Green Version]
- Groisman, E.A.; Ochman, H. Pathogenicity islands: Bacterial evolution in quantum leaps. Cell 1996, 87, 791–794. [Google Scholar] [CrossRef] [Green Version]
- Aviv, G.; Elpers, L.; Mikhlin, S.; Cohen, H.; Vitman Zilber, S.; Grassl, G.A.; Rahav, G.; Hensel, M.; Gal-Mor, O. The plasmid-encoded Ipf and Klf fimbriae display different expression and varying roles in the virulence of Salmonella enterica serovar Infantis in mouse vs. avian hosts. PLOS Pathog. 2017, 13, e1006559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaidullina, E.; Shelenkov, A.; Yanushevich, Y.; Mikhaylova, Y.; Shagin, D.; Alexandrova, I.; Ershova, O.; Akimkin, V.; Kozlov, R.; Edelstein, M. Antimicrobial resistance and genomic characterization of OXA-48-and CTX-M-15-co-producing hypervirulent Klebsiella pneumoniae st23 recovered from nosocomial outbreak. Antibiotics 2020, 9, 862. [Google Scholar] [CrossRef] [PubMed]
- McGinn, J.; Marraffini, L.A. Molecular mechanisms of CRISPR–Cas spacer acquisition. Nat. Rev. Microbiol. 2019, 17, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, F.; Beisel, C.L. CRISPR-Cas Systems and the Paradox of Self-Targeting Spacers. Front. Microbiol. 2020, 10, 3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Liu, J.; Du, B.; Ruan, H.-H.; Huo, Y.-X.; Du, Y.; Qiao, J. Whole-Genome-Based Survey for Polyphyletic Serovars of Salmonella enterica subsp. Enterica Provides New Insights into Public Health Surveillance. Int. J. Mol. Sci. 2020, 21, 5226. [Google Scholar] [CrossRef]
- Rakov, A.V.; Kuznetsova, N.A.; Yakovlev, A.A. Genetic diversity of Salmonella enterica subsp. enterica serovar Enteritidis in the Siberia and Far East of Russia based on plasmid profiles. AIMS Microbiol. 2020, 6, 106–120. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Year Collected | Region | Source |
|---|---|---|---|
| Crie-F81 | 2018 | Moscow region | Chicken |
| Crie-F87 | 2018 | Moscow region | Chicken |
| Crie-F191 | 2019 | Chelyabinsk region | Ready-to-cook chicken |
| Crie-F192 | 2019 | Chelyabinsk region | Chicken |
| Crie-F207 | 2019 | Saratov region | Turkey |
| Crie-F241 | 2019 | Pskov region | Ready-to-cook chicken cutlets |
| Crie-F243 | 2019 | Novgorod region | Meat |
| Crie-F244 | 2019 | Altai Krai | Chicken |
| Crie-F245 | 2019 | Rostov region | Ready-to-cook meat |
| Crie-F247 | 2019 | Republic of Tatarstan | Ready-to-cook meat |
| Crie-F252 | 2019 | Republic of Tatarstan | Chicken |
| Crie-F254 | 2019 | Ryazan region | Ready-to-cook chicken cutlets |
| Crie-F257 | 2019 | Sverdlovsk region | Ready-to-cook meat cutlets |
| Crie-F259 | 2019 | Sverdlovsk region | Chicken |
| Crie-F260 | 2019 | Sverdlovsk region | Ready-to-cook meat |
| Crie-F261 | 2019 | Krasnoyarsk region | Ready-to-cook meat cutlets |
| Crie-F339 | 2019 | Republic of Tatarstan | Chicken |
| Crie-F386 | 2019 | Vologda region | Ready-to-cook meat cutlets |
| Crie-F391 | 2019 | Amur region | Ready-to-cook chicken cutlets |
| Crie-F392 | 2019 | Rostov region | Chicken |
| Crie-F393 | 2019 | Rostov region | Chicken |
| Crie-F394 | 2019 | Rostov region | Ready-to-cook chicken cutlets |
| Crie-F396 | 2019 | Rostov region | Chicken |
| Crie-F403 | 2019 | Novgorod region | Chicken |
| Crie-F410 | 2019 | Tver region | Ready-to-cook chicken cutlets |
| Crie-F411 | 2019 | Moscow region | Chicken |
| Crie-F417 | 2019 | Moscow | Chicken |
| Crie-F418 | 2019 | Moscow | Chicken |
| Crie-F441 | 2020 | Ulyanovsk region | Chicken |
| Crie-F443 | 2020 | Krasnoyarsk region | Chicken |
| Crie-F444 | 2020 | Krasnoyarsk region | Chicken |
| Crie-F445 | 2020 | Krasnoyarsk region | Chicken |
| Crie-F446 | 2020 | Krasnoyarsk region | Chicken |
| Crie-F448 | 2020 | Krasnoyarsk region | Chicken |
| Crie-F468 | 2020 | Samara region | Turkey |
| Crie-F469 | 2020 | Samara region | Pork |
| Crie-F470 | 2020 | Samara region | Ready-to-cook meat |
| Crie-F489 | 2020 | Belgorod region | Chicken |
| Crie-F503 | 2020 | Moscow region | Ready-to-cook beef cutlets |
| Crie-F506 | 2020 | Perm region | Ready-to-cook chicken cutlets |
| Crie-F508 | 2020 | Perm region | Chicken |
| Crie-F509 | 2020 | Perm region | Chicken |
| Crie-F510 | 2020 | Perm region | Chicken |
| Crie-F512 | 2020 | Primorsky Krai | Chicken |
| Crie-F522 | 2020 | Vologda region | Chicken |
| Isolate | Plasmid Replicon | |||
|---|---|---|---|---|
| Col | IncF | IncX | Other | |
| Crie-F81 | - | IncFIB | IncX1 | - |
| Crie-F87 | - | IncFIB | IncX1 | - |
| Crie-F191 | - | IncFII | - | - |
| Crie-F192 | - | - | - | - |
| Crie-F207 | Col440I | - | - | - |
| Crie-F241 | - | IncFIB | - | - |
| Crie-F243 | - | IncFIB | - | - |
| Crie-F244 | - | IncFIB | - | IncHI2, IncHI2A, IncN |
| Crie-F245 | - | IncFIB | - | - |
| Crie-F247 | - | IncFIB | IncX1 | - |
| Crie-F252 | - | IncFIB | - | - |
| Crie-F254 | - | IncFIB | - | - |
| Crie-F257 | - | IncFIB | - | - |
| Crie-F259 | - | IncFIB | - | - |
| Crie-F260 | - | IncFIB | IncX1 | - |
| Crie-F261 | - | IncFIB | - | IncHI2A, IncHI2, IncN |
| Crie-F339 | - | IncFIB | - | - |
| Crie-F386 | ColpVC | IncFIB | - | - |
| Crie-F391 | - | IncFIB | - | - |
| Crie-F392 | - | IncFIB | - | - |
| Crie-F393 | - | IncFIB | - | - |
| Crie-F394 | - | IncFIB | - | - |
| Crie-F396 | - | IncFIB | - | - |
| Crie-F403 | - | IncFIB | - | - |
| Crie-F410 | - | IncFIB | IncX1 | - |
| Crie-F411 | - | IncFIB | - | - |
| Crie-F417 | - | IncFIB | - | - |
| Crie-F418 | - | IncFIB | IncX1 | - |
| Crie-F441 | - | IncFIB | - | - |
| Crie-F443 | - | IncFIB | - | - |
| Crie-F444 | - | IncFIB | IncX1 | - |
| Crie-F445 | - | IncFIB | - | - |
| Crie-F446 | - | IncFIB | - | - |
| Crie-F448 | - | IncFIB | IncX1 | - |
| Crie-F468 | - | IncFIB | IncX4 | - |
| Crie-F469 | - | IncFIB | - | - |
| Crie-F470 | - | IncFIB | IncX1 | - |
| Crie-F489 | - | IncFIB | IncX1 | - |
| Crie-F503 | ColRNAI, ColpVC | IncFIB | - | - |
| Crie-F506 | - | IncFIB | - | - |
| Crie-F508 | - | IncFIB | IncX1 | - |
| Crie-F509 | - | IncFIB | IncX1 | - |
| Crie-F510 | Col(pHAD28), ColRNAI | IncFIB | IncX1 | - |
| Crie-F512 | - | IncFIB | - | - |
| Crie-F522 | - | IncFIB | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, A.; Mikhaylova, Y.; Saenko, S.; Tyumentseva, M.; Tyumentsev, A.; Karbyshev, K.; Chernyshkov, A.; Manzeniuk, I.; Akimkin, V.; Shelenkov, A. Comparative Whole-Genome Analysis of Russian Foodborne Multidrug-Resistant Salmonella Infantis Isolates. Microorganisms 2022, 10, 89. https://doi.org/10.3390/microorganisms10010089
Egorova A, Mikhaylova Y, Saenko S, Tyumentseva M, Tyumentsev A, Karbyshev K, Chernyshkov A, Manzeniuk I, Akimkin V, Shelenkov A. Comparative Whole-Genome Analysis of Russian Foodborne Multidrug-Resistant Salmonella Infantis Isolates. Microorganisms. 2022; 10(1):89. https://doi.org/10.3390/microorganisms10010089
Chicago/Turabian StyleEgorova, Anna, Yulia Mikhaylova, Stepan Saenko, Marina Tyumentseva, Aleksandr Tyumentsev, Konstantin Karbyshev, Aleksey Chernyshkov, Igor Manzeniuk, Vasiliy Akimkin, and Andrey Shelenkov. 2022. "Comparative Whole-Genome Analysis of Russian Foodborne Multidrug-Resistant Salmonella Infantis Isolates" Microorganisms 10, no. 1: 89. https://doi.org/10.3390/microorganisms10010089
APA StyleEgorova, A., Mikhaylova, Y., Saenko, S., Tyumentseva, M., Tyumentsev, A., Karbyshev, K., Chernyshkov, A., Manzeniuk, I., Akimkin, V., & Shelenkov, A. (2022). Comparative Whole-Genome Analysis of Russian Foodborne Multidrug-Resistant Salmonella Infantis Isolates. Microorganisms, 10(1), 89. https://doi.org/10.3390/microorganisms10010089