
Broad Antiviral Effects of Echinacea purpurea against SARS-CoV-2 Variants of Concern and Potential Mechanism of Action

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Material
Cells and Viruses
2.2. SARS-CoV-2 Pseudoparticle Generation
2.3. Cell Viability Assays
2.4. Virucidal Activity
2.4.1. Plaque Reduction Assay
2.4.2. Cytopathic Effect
2.5. Pretreatment of Human Airway Epithelial Cells
2.6. Immunofluorescence Staining
2.7. Molecular Modeling and Molecular Docking Studies
2.8. Statistics
2.9. Biosafety
3. Results
3.1. Cell Viability Tests
3.2. Pretreatment of VOCs Prevents Viral Propagation at Low, Non-Toxic EF Concentrations
3.2.1. Pretreatment of Pseudoviruses Expressing Wild Type S Protein Prevents Viral Infection at Low, Non-Toxic Concentrations
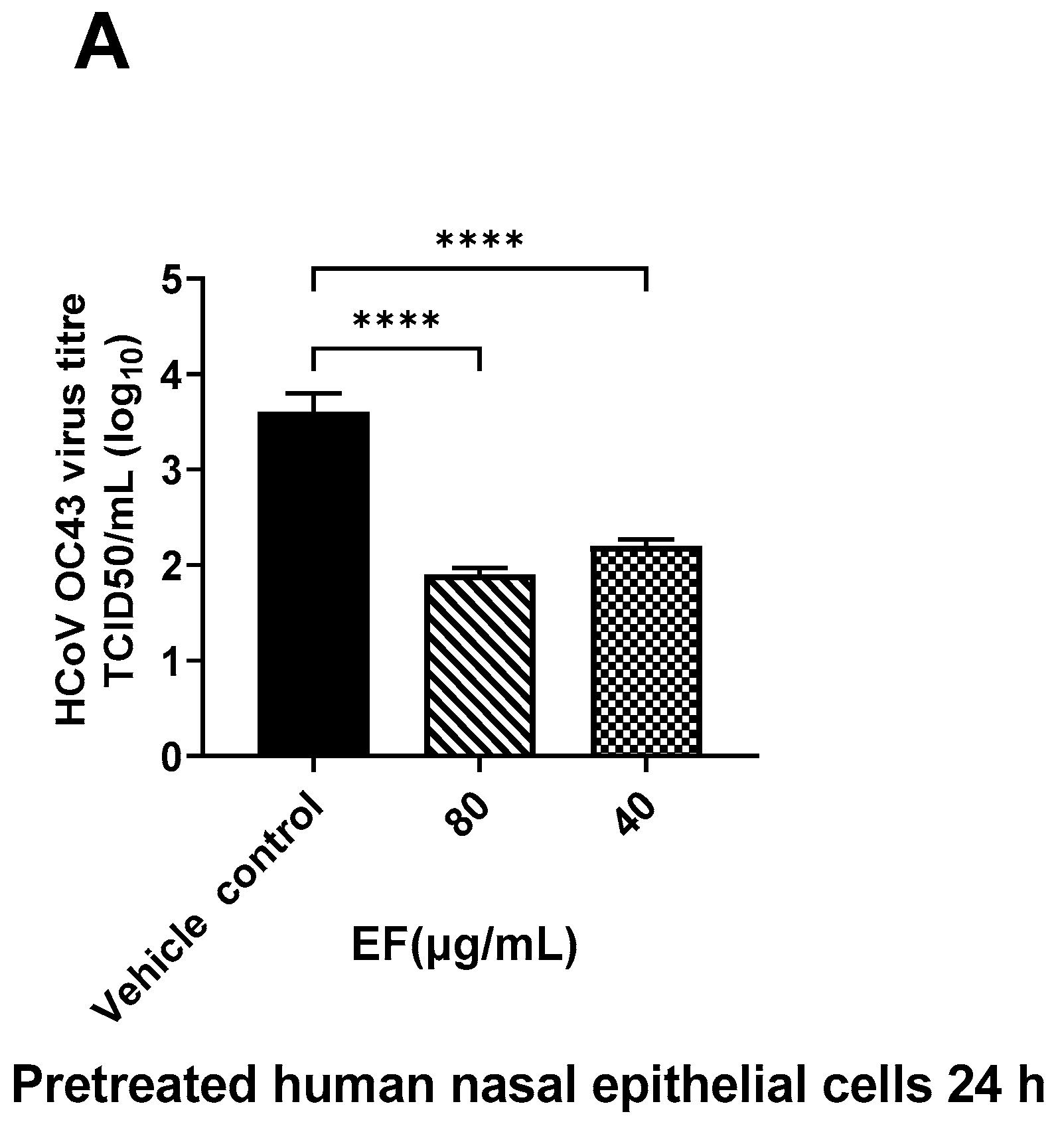
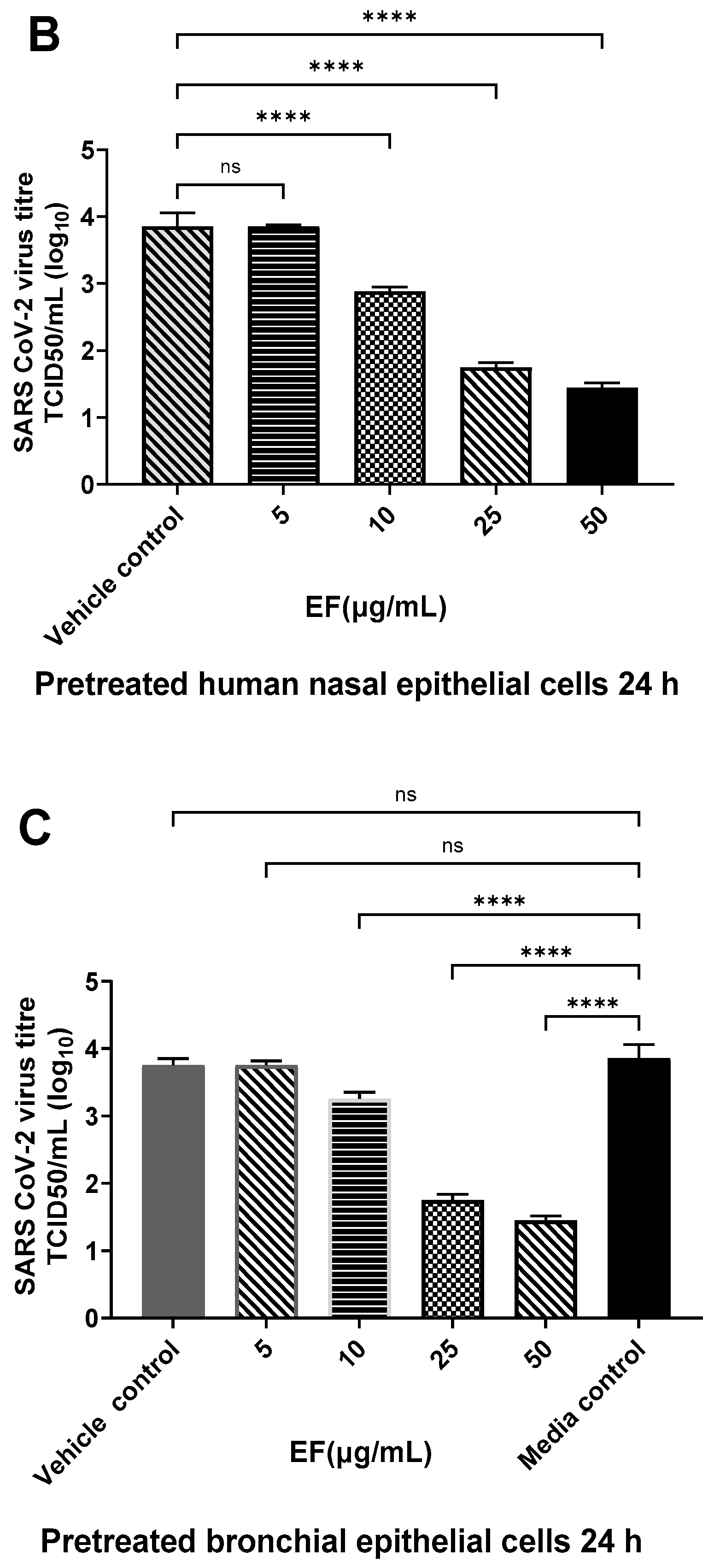
3.2.2. Pretreatment of Primary HNEpC and HBEpC Impairs Infection with OC-43 and SARS-CoV-2
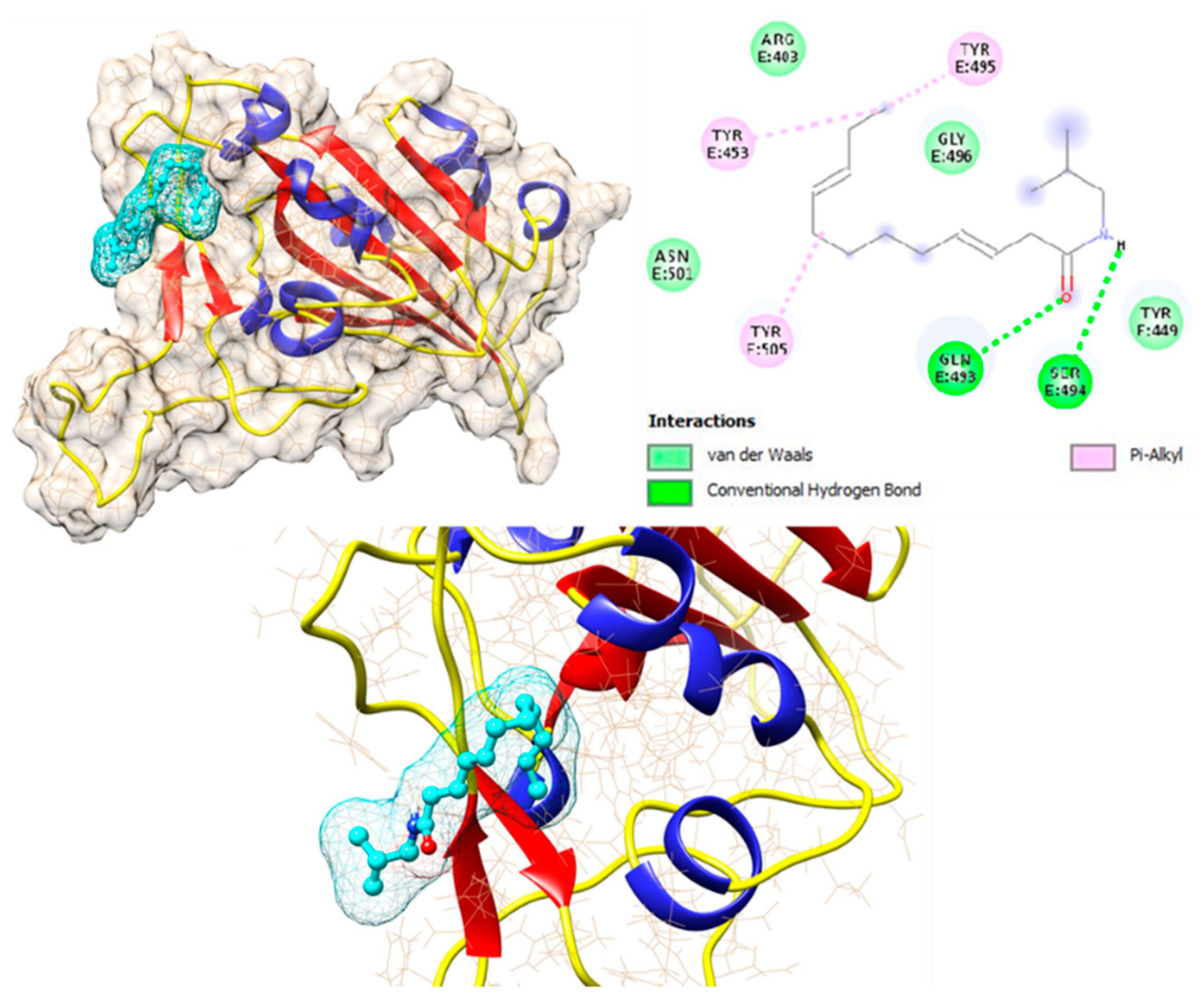
3.3. Molecular Docking (MD)
3.4. Echinacea’s Interaction with Pharmaceutical Targets for CoV Infection Process
3.5. Effect of EF on TMPRSS-2 Expression
4. Discussions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lauring, A.S.; Hodcroft, E.B. Genetic Variants of SARS-CoV-2—What Do They Mean? JAMA 2021, 325, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Rampersad, S.; Tennant, P. Replication and Expression Strategies of Viruses. Viruses 2018, 55–82. [Google Scholar] [CrossRef]
- Jackson, B.; Boni, M.F.; Bull, M.J.; Colleran, A.; Colquhoun, R.M.; Darby, A.C.; Haldenby, S.; Hill, V.; Lucaci, A.; McCrone, J.T.; et al. Generation and Transmission of Interlineage Recombinants in the SARS-CoV-2 Pandemic. Cell 2021, 184, 5179.e8–5188.e8. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Mossman, K.; Grandvaux, N. Molecular Determinants of SARS-CoV-2 Variants. Trends Microbiol. 2021, 29, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.H.; Morris, C.P.; Sachithanandham, J.; Amadi, A.; Gaston, D.; Li, M.; Swanson, N.J.; Schwartz, M.; Klein, E.Y.; Pekosz, A.; et al. Infection with the SARS-CoV-2 Delta Variant Is Associated with Higher Infectious Virus Loads Compared to the Alpha Variant in Both Unvaccinated and Vaccinated Individuals. medRxiv 2021. [Google Scholar] [CrossRef]
- Zhou, D.; Dejnirattisai, W.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.E.; Tuekprakhon, A.; Nutalai, R.; et al. Evidence of Escape of SARS-CoV-2 Variant B.1.351 from Natural and Vaccine-Induced Sera. Cell 2021, 184, 2348.e6–2361.e6. [Google Scholar] [CrossRef]
- Collier, D.A.; Ferreira, I.A.T.M.; Kotagiri, P.; Datir, R.P.; Lim, E.Y.; Touizer, E.; Meng, B.; Abdullahi, A.; The CITIID-NIHR BioResource COVID-19 Collaboration; Elmer, A.; et al. Age-Related Immune Response Heterogeneity to SARS-CoV-2 Vaccine BNT162b2. Nature 2021, 596, 417–422. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, Á.; Hill, V.; Pybus, O.G.; Watts, A.; Bogoch, I.I.; Khan, K.; Messina, J.P.; Tegally, H.; Lessells, R.R.; Giandhari, J.; et al. Tracking the International Spread of SARS-CoV-2 Lineages B.1.1.7 and B.1.351/501Y-V2 with Grinch. Wellcome Open Res. 2021, 6, 121. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated Transmissibility and Impact of SARS-CoV-2 Lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Kidd, M.; Richter, A.; Best, A.; Cumley, N.; Mirza, J.; Percival, B.; Mayhew, M.; Megram, O.; Ashford, F.; White, T.; et al. S-Variant SARS-CoV-2 Lineage B1.1.7 Is Associated With Significantly Higher Viral Load in Samples Tested by TaqPath Polymerase Chain Reaction. J. Infect. Dis. 2021, 223, 1666–1670. [Google Scholar] [CrossRef]
- Monel, B.; Planas, D.; Grzelak, L.; Smith, N.; Robillard, N.; Staropoli, I.; Goncalves, P.; Porrot, F.; Guivel-Benhassine, F.; Guinet, N.D.; et al. Release of Infectious Virus and Cytokines in Nasopharyngeal Swabs from Individuals Infected with Non-Alpha or Alpha SARS-CoV-2 Variants: An Observational Retrospective Study. EBioMedicine 2021, 73, 103637. [Google Scholar] [CrossRef] [PubMed]
- Greaney, A.J.; Starr, T.N.; Gilchuk, P.; Zost, S.J.; Binshtein, E.; Loes, A.N.; Hilton, S.K.; Huddleston, J.; Eguia, R.; Crawford, K.H.D.; et al. Complete Mapping of Mutations to the SARS-CoV-2 Spike Receptor-Binding Domain That Escape Antibody Recognition. Cell Host Microbe 2021, 29, 44.e9–57.e9. [Google Scholar] [CrossRef] [PubMed]
- Castonguay, N.; Zhang, W.; Langlois, M.A. Meta-Analysis and Structural Dynamics of the Emergence of Genetic Variants of SARS-CoV-2. Front. Microbiol. 2021, 12, 676314. [Google Scholar] [CrossRef]
- Cele, S.; Gazy, I.; Jackson, L.; Hwa, S.H.; Tegally, H.; Lustig, G.; Giandhari, J.; Pillay, S.; Wilkinson, E.; Naidoo, Y.; et al. Escape of SARS-CoV-2 501Y.V2 from Neutralization by Convalescent Plasma. Nature 2021, 593, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.E.; Zhang, X.; Case, J.B.; Winkler, E.S.; Liu, Y.; VanBlargan, L.A.; Liu, J.; Errico, J.M.; Xie, X.; Suryadevara, N.; et al. Resistance of SARS-CoV-2 Variants to Neutralization by Monoclonal and Serum-Derived Polyclonal Antibodies. Nat. Med. 2021, 27, 717–726. [Google Scholar] [CrossRef]
- Salvatore, M.; Bhattacharyya, R.; Purkayastha, S.; Zimmermann, L.; Ray, D.; Hazra, A.; Kleinsasser, M.; Mellan, T.; Whittaker, C.; Flaxman, S.; et al. Resurgence of SARS-CoV-2 in India: Potential Role of the B.1.617.2 (Delta) Variant and Delayed Interventions. medRxiv 2021. [Google Scholar] [CrossRef]
- Christensen, P.A.; Olsen, R.J.; Long, S.W.; Subedi, S.; Davis, J.J.; Hodjat, P.; Walley, D.R.; Kinskey, J.C.; Saavedra, M.O.; Pruitt, L.; et al. Delta Variants of SARS-CoV-2 Cause Significantly Increased Vaccine Breakthrough COVID-19 Cases in Houston, Texas. Am. J. Pathol. 2021, 192, 320–331. [Google Scholar] [CrossRef]
- Chia, P.Y.; Ong, S.W.X.; Chiew, C.J.; Ang, L.W.; Chavatte, J.M.; Mak, T.M.; Cui, L.; Kalimuddin, S.; Chia, W.N.; Tan, C.W.; et al. Virological and Serological Kinetics of SARS-CoV-2 Delta Variant Vaccine-Breakthrough Infections: A Multi-Center Cohort Study. Clin. Microbiol. Infect. 2021, 28, 612.e1–612.e7. [Google Scholar] [CrossRef]
- Schmidt, F.; Muecksch, F.; Weisblum, Y.; Da Silva, J.; Bednarski, E.; Cho, A.; Wang, Z.; Gaebler, C.; Caskey, M.; Nussenzweig, M.C.; et al. Plasma Neutralization Properties of the SARS-CoV-2 Omicron Variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Moyo, S.; Amoako, D.G.; Baxter, C.; et al. Continued Emergence and Evolution of Omicron in South Africa: New BA.4 and BA.5 Lineages. medRxiv 2022. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.-M.; et al. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Krüger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron Variant Is Highly Resistant against Antibody-Mediated Neutralization: Implications for Control of the COVID-19 Pandemic. Cell 2022, 185, 447.e11–456.e11. [Google Scholar] [CrossRef] [PubMed]
- Van der Straten, K.; Guerra, D.; van Gils, M.J.; Bontjer, I.; Caniels, T.G.; van Willigen, H.D.G.; Wynberg, E.; Poniman, M.; Burger, J.A.; Bouhuijs, J.H.; et al. Antigenic Cartography Using Sera from Sequence-Confirmed SARS-CoV-2 Variants of Concern Infections Reveals Antigenic Divergence of Omicron. Immunity 2022, 55, 1725.e4–1731.e4. [Google Scholar] [CrossRef] [PubMed]
- Cevik, M.; Grubaugh, N.D.; Iwasaki, A.; Openshaw, P. COVID-19 Vaccines: Keeping Pace with SARS-CoV-2 Variants. Cell 2021, 184, 5077–5081. [Google Scholar] [CrossRef]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E.; et al. Efficacy of the ChAdOx1 NCoV-19 Covid-19 Vaccine against the B.1.351 Variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef]
- Vimalanathan, S.; Schoop, R.; Hudson, J. High-Potency Anti-Influenza Therapy by a Combination of Echinacea Purpurea Fresh Herb and Root Tinctures. J. Appl. Pharm. Sci. 2013, 3, 1–5. [Google Scholar] [CrossRef]
- Pleschka, S.; Stein, M.; Schoop, R.; Hudson, J.B. Anti-Viral Properties and Mode of Action of Standardized Echinacea Purpurea Extract against Highly Pathogenic Avian Influenza Virus (H5N1, H7N7) and Swine-Origin H1N1 (S-OIV). Virol. J. 2009, 6, 197. [Google Scholar] [CrossRef] [Green Version]
- Signer, J.; Jonsdottir, H.R.; Albrich, W.C.; Strasser, M.; Zust, R.; Ryter, S.; Ackermann-Gaumann, R.; Lenz, N.; Siegrist, D.; Suter, A.; et al. In Vitro Virucidal Activity of Echinaforce((R)), an Echinacea Purpurea Preparation, against Coronaviruses, Including Common Cold Coronavirus 229E and SARS-CoV-2. Virol. J. 2020, 17, 172. [Google Scholar] [CrossRef]
- Nicolussi, S.; Ardjomand-Woelkart, K.; Stange, R.; Gancitano, G.; Klein, P.; Ogal, M. Echinacea as a Potential Force against Coronavirus Infections? A Mini-Review of Randomized Controlled Trials in Adults and Children. Microorganisms 2022, 10, 211. [Google Scholar] [CrossRef]
- Thakur, N.; Gallo, G.; Elreafey, A.M.E.; Bailey, D. Production of Recombinant Replication-Defective Lentiviruses Bearing the SARS-CoV or SARS-CoV-2 Attachment Spike Glycoprotein and Their Application in Receptor Tropism and Neutralisation Assays. Bio-Protocol 2021, 11, e4249. [Google Scholar] [CrossRef]
- Conceicao, C.; Thakur, N.; Human, S.; Kelly, J.T.; Logan, L.; Bialy, D.; Bhat, S.; Stevenson-Leggett, P.; Zagrajek, A.K.; Hollinghurst, P.; et al. The SARS-CoV-2 Spike Protein Has a Broad Tropism for Mammalian ACE2 Proteins. PLoS Biol. 2020, 18, e3001016. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein Structure and Function Prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. Protein-Ligand Binding Site Recognition Using Complementary Binding-Specific Substructure Comparison and Sequence Profile Alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed Atlas of Surface Topography of Proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef] [Green Version]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein-Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Schoop, R.; Hudson, J.B. Echinacea as an Antiinflammatory Agent: The Influence of Physiologically Relevant Parameters. Phytother. Res. 2009, 23, 863–867. [Google Scholar] [CrossRef]
- Jawad, M.; Schoop, R.; Suter, A.; Klein, P.; Eccles, R. Safety and Efficacy Profile of Echinacea Purpurea to Prevent Common Cold Episodes: A Randomized, Double-Blind, Placebo-Controlled Trial. Evid.-Based Complement. Altern. Med. 2012, 2012, 841315. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Echinacea Purpurea Compounds | 3CLpro | PLpro | RdRp | S-Protein | NSP9 | NSP13 | NSP14 | NSP15 | NSP16 | ACE2 | TMPRSS-2 | JAK-Janus Kianse |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Undeca-2E,4Z-dien-8,10-diinsaure-isobutylamid | −5.7 | −4.6 | −5 | −5.4 | −4.6 | −5.5 | −6.5 | −5.9 | −5.6 | −4.3 | −5 | −5.8 |
| Undeca-2Z,4E-dien-8,10-diinsaure-isobutylamid | −5.7 | −4.7 | −5 | −5.3 | −4.4 | −5.6 | −7 | −4.8 | −5.8 | −5.9 | −5.2 | −5.8 |
| Dodeca-2E,4Z-dien-8,10-diinsaure-isobutylamid | −5.1 | −5.6 | −5.2 | −5.2 | −4.5 | −5.8 | −7.4 | −5.8 | −6 | −5.6 | −5 | −6.1 |
| Undeca-2E,4Z-dien-8,10-diinsaure-2-methybutylamid | −5.3 | −5.3 | −5.1 | −5.2 | −4.5 | −5.7 | −6.7 | −5.6 | −6.1 | −5.9 | −5.5 | −5.8 |
| Dodeca-2E,4E,10E-trien-8-insaure-isobutylamid | −5.2 | −5.4 | −5.4 | −5.3 | −4.9 | −5.9 | −7.3 | −6.1 | −5.8 | −5.7 | −5.6 | −6.1 |
| Trideca-2E,7Z-dien-10,12-diinasaure-isobutylamid | −4.8 | −4.4 | −5.2 | −5.3 | −4.5 | −5.6 | −6.6 | −5.8 | −5.6 | −4.4 | −5.3 | −5.9 |
| Dodeca-2E,4Z-dien-8,10-diinasaure-2-methybutylamid | −5.2 | −5.4 | −5.2 | −6.9 | −4.1 | −6.4 | −7.1 | −5.5 | −5.8 | −6.3 | −5 | −6.3 |
| Dodeca-2E,4E,8Z,10E-tetraensaure-isobutylamid | −5.2 | −4.7 | −5.2 | −7.6 | −4.7 | −5.9 | −7.6 | −6.2 | −6.1 | −4.7 | −5.2 | −5.8 |
| Dodeca-2E,4E,8Z,10Z-tetraensaure-isobutylamid | −5 | −5.3 | −5.1 | −6.0 | −4.6 | −5.9 | −6.6 | −6.2 | −5.8 | −5.5 | −5.4 | −5.9 |
| Dodeca-2E,4E,8Z-triensaure-isobutylamid | −4.9 | −5.3 | −5.3 | −6.0 | −4.3 | −5.7 | −6.3 | −5.9 | −5.6 | −4.4 | −5.3 | −5.5 |
| Dodeca-2E,4E-diensaure-isobutylamid | −4.7 | −4.3 | −4.8 | −5.0 | −4.3 | −5.5 | −6.5 | −5.1 | −5.6 | −5.6 | −5.3 | −5.7 |
| 7-Hydroxy-Dodeca-2E,4E,8Z,10E-tetraensaure-isobutylamid | −5.4 | −4.8 | −5.7 | −4.8 | −4.5 | −6 | −7.1 | −6.5 | −6.2 | −4.8 | −5.8 | −5.9 |
| Undeca-2E,4Z-dien-8,10-diinsäure-isobutylamide | −5.2 | −5 | −5 | −5.6 | −4.8 | −5.7 | −6.9 | −5.9 | −5.7 | −5.6 | −5.3 | −5.8 |
| Pentadeca-2E,9Z-dien-12,14 diinsaure-isobutylamide and hydroxylated derivates | −5.1 | −4.3 | −4.6 | −4.9 | −4.3 | −5.8 | −6.8 | −5.2 | −5.4 | −4.8 | −4.9 | −5.7 |
| Trideca-2E,7Z-dien-10,12-diinsäure-isobutylamid | −4.9 | −5.3 | −4.8 | −5.5 | −4.6 | −6.3 | −7.3 | −5.6 | −5.5 | −6 | −5.5 | −6 |
| Caftaric acid | −6.8 | −6.4 | −6.6 | −5.1 | −5.1 | −7.1 | −7.4 | −6.6 | −7.3 | −6.4 | −6.6 | −6.8 |
| 2-0-feruloly-tartaric acid | −6.4 | −6.3 | −6.6 | −5.7 | −4.6 | −7.1 | −7.6 | −6.2 | −7.4 | −5.9 | −6.4 | −6.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vimalanathan, S.; Shehata, M.; Sadasivam, K.; Delbue, S.; Dolci, M.; Pariani, E.; D’Alessandro, S.; Pleschka, S. Broad Antiviral Effects of Echinacea purpurea against SARS-CoV-2 Variants of Concern and Potential Mechanism of Action. Microorganisms 2022, 10, 2145. https://doi.org/10.3390/microorganisms10112145
Vimalanathan S, Shehata M, Sadasivam K, Delbue S, Dolci M, Pariani E, D’Alessandro S, Pleschka S. Broad Antiviral Effects of Echinacea purpurea against SARS-CoV-2 Variants of Concern and Potential Mechanism of Action. Microorganisms. 2022; 10(11):2145. https://doi.org/10.3390/microorganisms10112145
Chicago/Turabian StyleVimalanathan, Selvarani, Mahmoud Shehata, Kannan Sadasivam, Serena Delbue, Maria Dolci, Elena Pariani, Sarah D’Alessandro, and Stephan Pleschka. 2022. "Broad Antiviral Effects of Echinacea purpurea against SARS-CoV-2 Variants of Concern and Potential Mechanism of Action" Microorganisms 10, no. 11: 2145. https://doi.org/10.3390/microorganisms10112145
APA StyleVimalanathan, S., Shehata, M., Sadasivam, K., Delbue, S., Dolci, M., Pariani, E., D’Alessandro, S., & Pleschka, S. (2022). Broad Antiviral Effects of Echinacea purpurea against SARS-CoV-2 Variants of Concern and Potential Mechanism of Action. Microorganisms, 10(11), 2145. https://doi.org/10.3390/microorganisms10112145