
Lactobacillus reuteri MJM60668 Prevent Progression of Non-Alcoholic Fatty Liver Disease through Anti-Adipogenesis and Anti-Inflammatory Pathway

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Identification by 16S rDNA Sequence and Phylogenetic Analysis of MJM60668
2.2. Cell Culture and Treatment
2.3. Cell Viability Assay (MTT)
2.4. Oil Red O Staining
2.5. Safety Test
2.5.1. D-Lactate Production
2.5.2. Bile Salts Deconjugation
2.5.3. Antimicrobial Susceptibility Test (MIC)
2.5.4. Hemolytic Activity
2.5.5. Biogenic Amine Production Test
2.5.6. Mucin Degradation Activity
2.5.7. Antimicrobial Assay
2.5.8. Oro-Gastrointestinal Transit Assay
2.5.9. Cell Adhesion Assay
2.6. Animal Assay
2.6.1. Animal Monitoring and Treatment
2.6.2. Serum Biochemical Analysis
2.6.3. Liver, Kidney, Intestine Sectioning, and Histopathological Assessment
2.6.4. Western Blot Analysis
2.6.5. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
2.6.6. Fecal Sample Analysis
2.7. Statistical Analysis
3. Results
3.1. Phylogenic Analysis of MJM60668
3.2. Effect of MJM60668 on Viability of HepG2 Cells
3.3. Anti-Lipogenic Effect of MJM60668 on HepG2 Cells
3.4. Safety Assessment of MJM60668
3.4.1. D-Lactic Production, Bile Salt Deconjugation, Hemolytic Activity, Mucin Degradation Activity, and Antibiotic Susceptibility
3.4.2. Antibacterial Activity of MJM60668
3.4.3. Adherence of MJM60668 to HT-29 Cells
3.4.4. OGI Transit Assay
3.5. Animal Study
3.5.1. Effects of MJM60668 on Body Weight, Food Intake, Various Tissue Weights on HFD-Induced NAFLD Model in Mice
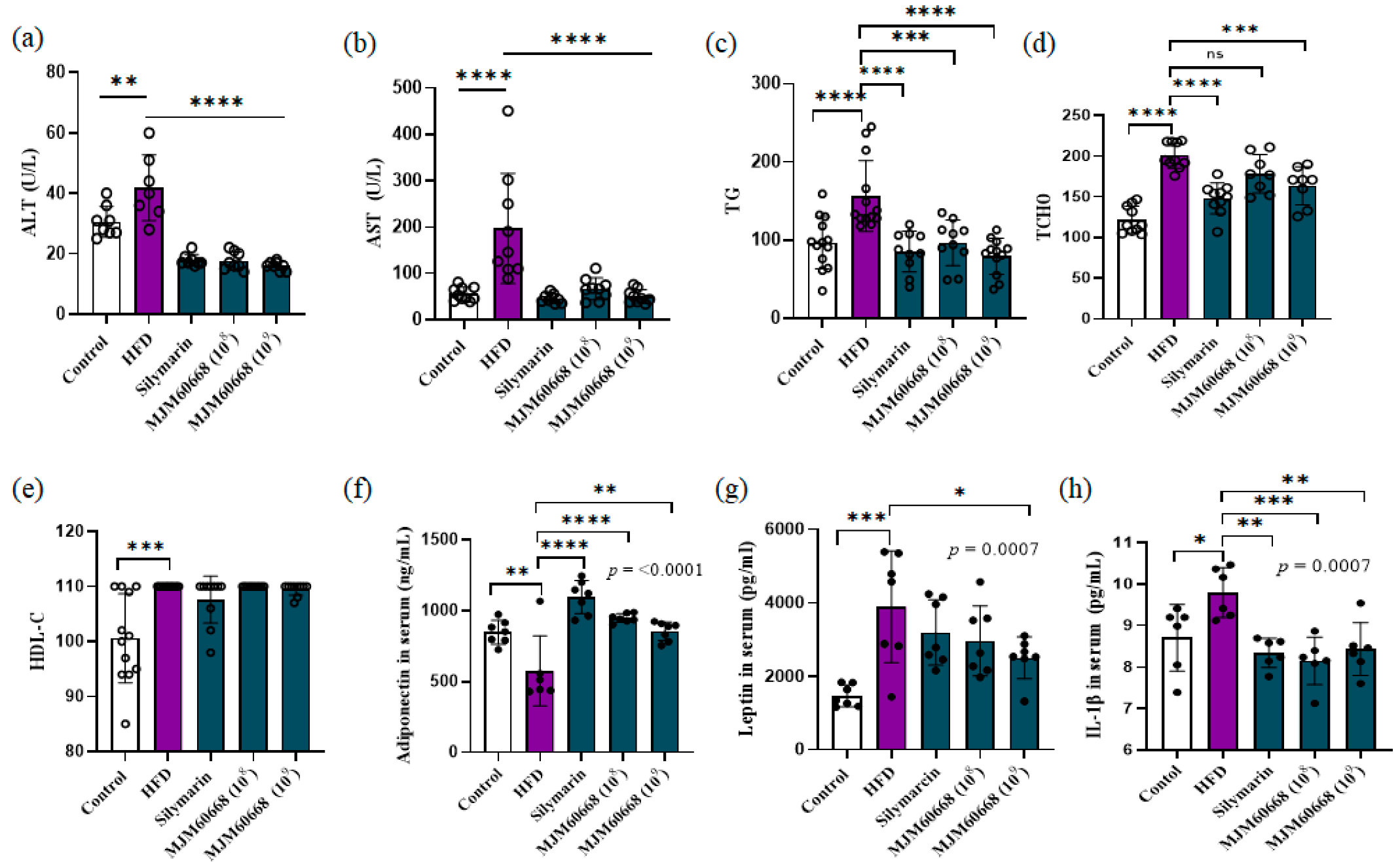
3.5.2. Effects of MJM60668 on Serum and Hepatic Lipid Profile in HFD-Induce NAFLD Mice Model
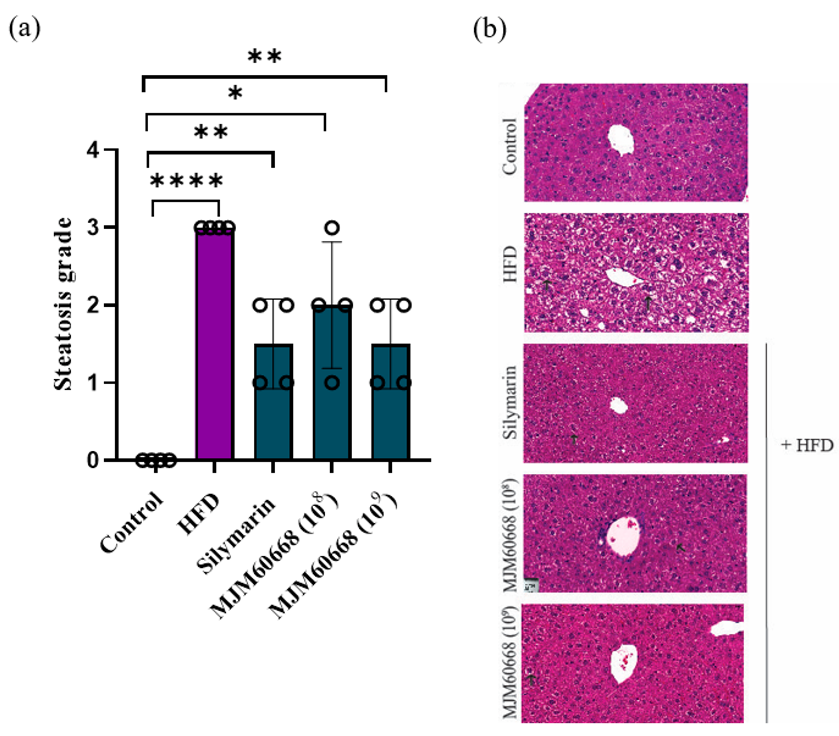
3.5.3. MJM60668 Can Improve Liver Regeneration under Steatosis Based on Histological Analysis
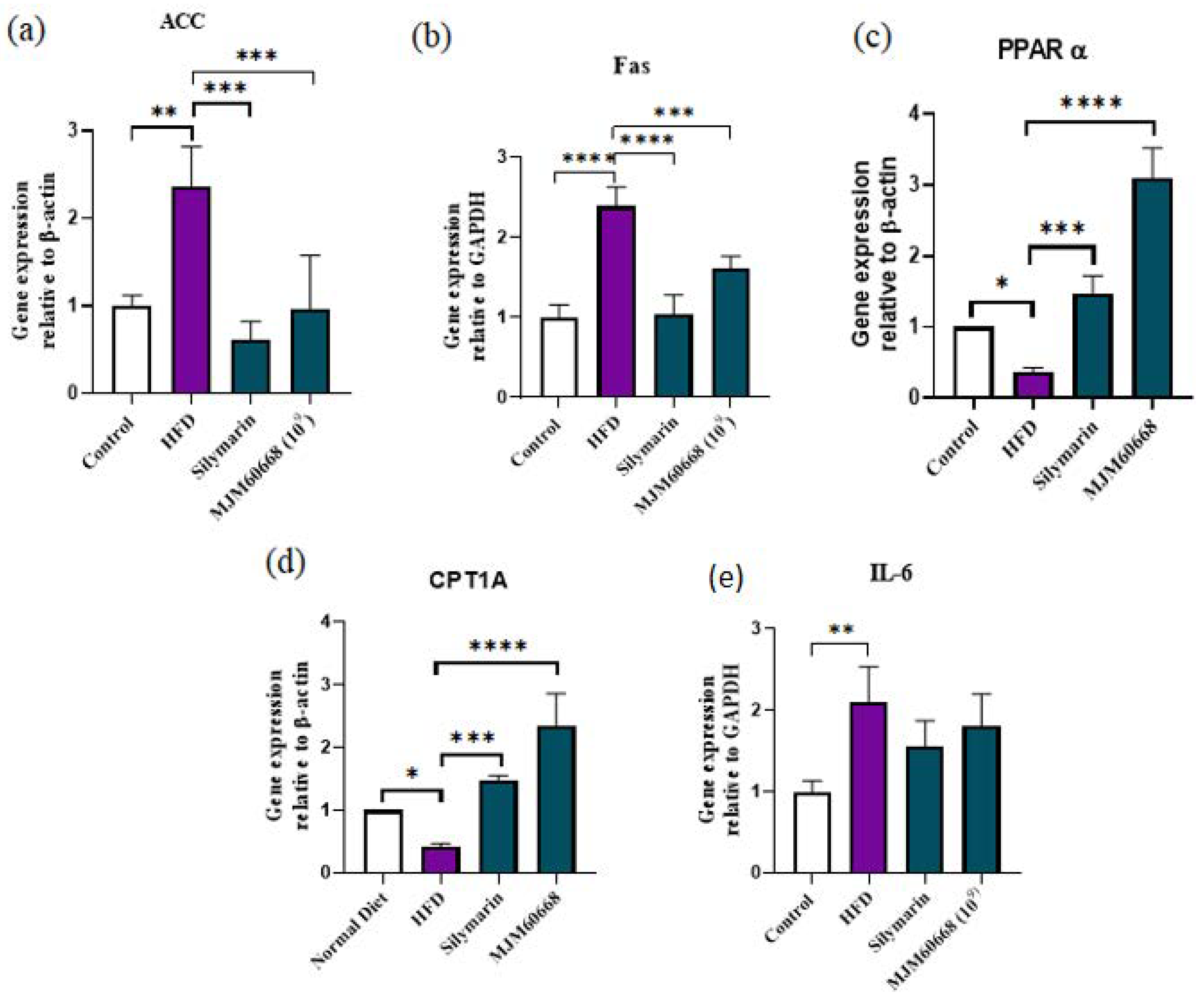
3.5.4. Effects of MJM60668 on ACC, Fas, PPARα, CPT1A, and IL-6 Gene Expression in HFD Group of Mice
3.5.5. Effects of MJM60668 on Proteins Involved in Fatty Acid Synthesis and Lipid Metabolism
3.5.6. MJM60668 Affects Fatty Acids Metabolism by Directly Affecting Adiponectin and Leptin Expression with an Anti-Inflammatory Effect on Liver
3.5.7. Effects of MJM60668 on Intestinal Microbiota Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eslam, M.; Valenti, L.; Romeo, S. Genetics and Epigenetics of NAFLD and NASH: Clinical Impact. J. Hepatol. 2018, 68, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Rinella, M.; Charlton, M. The Globalization of Nonalcoholic Fatty Liver Disease: Prevalence and Impact on World Health. Hepatology 2016, 64, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, C.D.; Targher, G. NAFLD: A Multisystem Disease. J. Hepatol. 2015, 62, S47–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, M.J.; Adams, L.A.; Canbay, A.; Syn, W.-K. Extrahepatic Complications of Nonalcoholic Fatty Liver Disease. Hepatology 2014, 59, 1174–1197. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Kawada, N. Inflammation and Fibrogenesis in Steatohepatitis. J. Gastroenterol. 2012, 47, 215–225. [Google Scholar] [CrossRef]
- Kitade, H.; Chen, G.; Ni, Y.; Ota, T. Nonalcoholic Fatty Liver Disease and Insulin Resistance: New Insights and Potential New Treatments. Nutrients 2017, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- Adipogenesis—An Overview|ScienceDirect Topics. Available online: https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/adipogenesis (accessed on 4 July 2022).
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding Adipocyte Differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [Green Version]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and Metabolic Health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Lipid-Droplet-Accumulating Microglia Represent a Dysfunctional and Proinflammatory State in the Aging Brain|Nature Neuroscience. Available online: https://www.nature.com/articles/s41593-019-0566-1 (accessed on 4 July 2022).
- Wobser, H.; Dorn, C.; Weiss, T.S.; Amann, T.; Bollheimer, C.; Büttner, R.; Schölmerich, J.; Hellerbrand, C. Lipid Accumulation in Hepatocytes Induces Fibrogenic Activation of Hepatic Stellate Cells. Cell Res. 2009, 19, 996–1005. [Google Scholar] [CrossRef] [Green Version]
- Sittipo, P.; Lobionda, S.; Lee, Y.K.; Maynard, C.L. Intestinal Microbiota and the Immune System in Metabolic Diseases. J. Microbiol. 2018, 56, 154–162. [Google Scholar] [CrossRef]
- Wang, R.; Tang, R.; Li, B.; Ma, X.; Schnabl, B.; Tilg, H. Gut Microbiome, Liver Immunology, and Liver Diseases. Cell Mol. Immunol. 2021, 18, 4–17. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, Their Health Benefits and Applications for Developing Healthier Foods: A Review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Gui, W.; Koo, I.; Smith, P.B.; Allman, E.L.; Nichols, R.G.; Rimal, B.; Cai, J.; Liu, Q.; Patterson, A.D. The Microbiome Modulating Activity of Bile Acids. Gut Microbes 2020, 11, 979–996. [Google Scholar] [CrossRef]
- Giraffa, G.; Chanishvili, N.; Widyastuti, Y. Importance of Lactobacilli in Food and Feed Biotechnology. Res. Microbiol. 2010, 161, 480–487. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in Transition: Evolution and Natural History of the Genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Genes and Molecules of Lactobacilli Supporting Probiotic Action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, C.N.; Rosenfeldt Nielsen, V.; Hayford, A.E.; Møller, P.L.; Michaelsen, K.F.; Paerregaard, A.; Sandström, B.; Tvede, M.; Jakobsen, M. Screening of Probiotic Activities of Forty-Seven Strains of Lactobacillus Spp. by in Vitro Techniques and Evaluation of the Colonization Ability of Five Selected Strains in Humans. Appl. Environ. Microbiol. 1999, 65, 4949–4956. [Google Scholar] [CrossRef] [Green Version]
- Lactobacillus reuteri, Sp. Nov., a New Species of Heterofermentative Lactobacilli—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0172556480800078?via%3Dihub (accessed on 8 June 2022).
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-I.; You, S.; Kim, S.; Won, G.; Kang, C.-H.; Kim, G.-H. Weissella Cibaria MG5285 and Lactobacillus reuteri MG5149 Attenuated Fat Accumulation in Adipose and Hepatic Steatosis in High-Fat Diet-Induced C57BL/6J Obese Mice. Food Nutr. Res. 2021, 65, 8087. [Google Scholar] [CrossRef]
- Ting, W.-J.; Kuo, W.-W.; Hsieh, D.J.-Y.; Yeh, Y.-L.; Day, C.-H.; Chen, Y.-H.; Chen, R.-J.; Padma, V.V.; Chen, Y.-H.; Huang, C.-Y. Heat Killed Lactobacillus reuteri GMNL-263 Reduces Fibrosis Effects on the Liver and Heart in High Fat Diet-Hamsters via TGF-β Suppression. Int. J. Mol. Sci. 2015, 16, 25881–25896. [Google Scholar] [CrossRef]
- Zhu, T.; Mao, J.; Zhong, Y.; Huang, C.; Deng, Z.; Cui, Y.; Liu, J.; Wang, H.L. Reuteri ZJ617 Inhibits Inflammatory and Autophagy Signaling Pathways in Gut-Liver Axis in Piglet Induced by Lipopolysaccharide. J. Anim. Sci. Biotechnol. 2021, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Wang, Z.; Stanton, C.; Ross, R.P.; Zhao, J.; Zhang, H.; Yang, B.; Chen, W. Lactobacillus rhamnosus FJSYC4-1 and Lactobacillus reuteri FGSZY33L6 Alleviate Metabolic Syndrome via Gut Microbiota Regulation. Food Funct. 2021, 12, 3919–3930. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Park, S.-B.; Kim, S.; Yang, S.H.; Suh, J.-W.; Lee, C.; Kim, J. Suppressing Activity of Staurosporine from Streptomyces sp. MJM4426 against Rice Bacterial Blight Disease. J. Appl. Microbiol. 2016, 120, 975–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.R.; Park, H.-J.; Kang, D.; Chung, H.; Nam, M.H.; Lee, Y.; Park, J.-H.; Lee, H.-Y. A Protective Mechanism of Probiotic Lactobacillus against Hepatic Steatosis via Reducing Host Intestinal Fatty Acid Absorption. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Wang, G.; Xia, Y.; Xiong, Z.; Ai, L. Bile Salt Hydrolase-Overexpressing Lactobacillus Strains Can Improve Hepatic Lipid Accumulation in Vitro in an NAFLD Cell Model. Food Nutr. Res. 2020, 64, 3751. [Google Scholar] [CrossRef]
- Pohanka, M. D-Lactic Acid as a Metabolite: Toxicology, Diagnosis, and Detection. Biomed Res. Int. 2020, 2020, 3419034. [Google Scholar] [CrossRef]
- Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance|EFSA. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/2740 (accessed on 20 July 2022).
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Palaniyandi, S.A.; Damodharan, K.; Suh, J.-W.; Yang, S.H. In Vitro Characterization of Lactobacillus Plantarum Strains with Inhibitory Activity on Enteropathogens for Use as Potential Animal Probiotics. Indian J. Microbiol. 2017, 57, 201–210. [Google Scholar] [CrossRef]
- Mouries, J.; Brescia, P.; Silvestri, A.; Spadoni, I.; Sorribas, M.; Wiest, R.; Mileti, E.; Galbiati, M.; Invernizzi, P.; Adorini, L.; et al. Microbiota-Driven Gut Vascular Barrier Disruption Is a Prerequisite for Non-Alcoholic Steatohepatitis Development. J. Hepatol. 2019, 71, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, L.A.; Salem, M.B.; Seif el-Din, S.H.; El-Lakkany, N.M.; Ahmed, H.O.; Nasr, S.M.; Hammam, O.A.; Botros, S.S.; Saleh, S. Gut Microbiota Modulation as a Promising Therapy with Metformin in Rats with Non-Alcoholic Steatohepatitis: Role of LPS/TLR4 and Autophagy Pathways. Eur. J. Pharmacol. 2020, 887, 173461. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and Validation of a Histological Scoring System for Nonalcoholic Fatty Liver Disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Lee, N.Y.; Yoon, S.J.; Han, D.H.; Gupta, H.; Youn, G.S.; Shin, M.J.; Ham, Y.L.; Kwak, M.J.; Kim, B.Y.; Yu, J.S.; et al. Lactobacillus and Pediococcus Ameliorate Progression of Non-Alcoholic Fatty Liver Disease through Modulation of the Gut Microbiome. Gut Microbes 2020, 11, 882–899. [Google Scholar] [CrossRef]
- Licholai, J.A.; Nguyen, K.P.; Fobbs, W.C.; Schuster, C.J.; Ali, M.A.; Kravitz, A.V. Why Do Mice Overeat High-Fat Diets? How High-Fat Diet Alters the Regulation of Daily Caloric Intake in Mice. Obesity 2018, 26, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- De Moura e Dias, M.; dos Reis, S.A.; da Conceição, L.L.; Sediyama, C.M.N.; de Pereira, O.S.S.; de Oliveira, L.L.; do Gouveia Peluzio, M.C.; Martinez, J.A.; Milagro, F.I. Diet-Induced Obesity in Animal Models: Points to Consider and Influence on Metabolic Markers. Diabetol. Metab. Syndr. 2021, 13, 32. [Google Scholar] [CrossRef]
- Engin, A. Non-Alcoholic Fatty Liver Disease. In Obesity and Lipotoxicity; Engin, A.B., Engin, A., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 443–467. ISBN 978-3-319-48382-5. [Google Scholar]
- Sanyal, D.; Mukherjee, P.; Raychaudhuri, M.; Ghosh, S.; Mukherjee, S.; Chowdhury, S. Profile of Liver Enzymes in Non-Alcoholic Fatty Liver Disease in Patients with Impaired Glucose Tolerance and Newly Detected Untreated Type 2 Diabetes. Indian J. Endocrinol. Metab. 2015, 19, 597–601. [Google Scholar] [CrossRef]
- Huang, X.-J.; Choi, Y.-K.; Im, H.-S.; Yarimaga, O.; Yoon, E.; Kim, H.-S. Aspartate Aminotransferase (AST/GOT) and Alanine Aminotransferase (ALT/GPT) Detection Techniques. Sensors 2006, 6, 756–782. [Google Scholar] [CrossRef] [Green Version]
- Fraulob, J.C.; Ogg-Diamantino, R.; Fernandes-Santos, C.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. A Mouse Model of Metabolic Syndrome: Insulin Resistance, Fatty Liver and Non-Alcoholic Fatty Pancreas Disease (NAFPD) in C57BL/6 Mice Fed a High Fat Diet. J. Clin. Biochem. Nutr. 2010, 46, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Omagari, K.; Kadokawa, Y.; Masuda, J.-I.; Egawa, I.; Sawa, T.; Hazama, H.; Ohba, K.; Isomoto, H.; Mizuta, Y.; Hayashida, K.; et al. Fatty Liver in Non-Alcoholic Non-Overweight Japanese Adults: Incidence and Clinical Characteristics. J. Gastroenterol. Hepatol. 2002, 17, 1098–1105. [Google Scholar] [CrossRef]
- Germani, G.; Laryea, M.; Rubbia-Brandt, L.; Egawa, H.; Burra, P.; O’Grady, J.; Watt, K.D. Management of Recurrent and De Novo NAFLD/NASH after Liver Transplantation. Transplantation 2019, 103, 57–67. [Google Scholar] [CrossRef]
- Caldwell, S.; Ikura, Y.; Dias, D.; Isomoto, K.; Yabu, A.; Moskaluk, C.; Pramoonjago, P.; Simmons, W.; Scruggs, H.; Rosenbaum, N.; et al. Hepatocellular Ballooning in NASH. J. Hepatol. 2010, 53, 719–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, H.; Huang, B.; Miao, G.; Yan, X.; Gao, G.; Luo, Y.; Chen, H.; Chen, W.; Yang, L. Mesenchymal Stem Cells Reverse High-fat Diet-induced Non-alcoholic Fatty Liver Disease through Suppression of CD4+ T Lymphocytes in Mice. Mol. Med. Rep. 2018, 17, 3769–3774. [Google Scholar] [CrossRef] [Green Version]
- Arjmand, A.; Angelis, C.T.; Christou, V.; Tzallas, A.T.; Tsipouras, M.G.; Glavas, E.; Forlano, R.; Manousou, P.; Giannakeas, N. Training of Deep Convolutional Neural Networks to Identify Critical Liver Alterations in Histopathology Image Samples. Appl. Sci. 2020, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Foretz, M.; Ancellin, N.; Andreelli, F.; Saintillan, Y.; Grondin, P.; Kahn, A.; Thorens, B.; Vaulont, S.; Viollet, B. Short-Term Overexpression of a Constitutively Active Form of AMP-Activated Protein Kinase in the Liver Leads to Mild Hypoglycemia and Fatty Liver. Diabetes 2005, 54, 1331–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Item, F.; Wueest, S.; Lemos, V.; Stein, S.; Lucchini, F.C.; Denzler, R.; Fisser, M.C.; Challa, T.D.; Pirinen, E.; Kim, Y.; et al. Fas Cell Surface Death Receptor Controls Hepatic Lipid Metabolism by Regulating Mitochondrial Function. Nat. Commun. 2017, 8, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Leon, S.; Tuvblad, C.; Forero, D. Sports Genetics: The PPARA Gene and Athletes’ High Ability in Endurance Sports. A Systematic Review and Meta-Analysis. Biol. Sport 2016, 33, 3–6. [Google Scholar] [CrossRef]
- van Raalte, D.H.; Li, M.; Pritchard, P.H.; Wasan, K.M. Peroxisome Proliferator-Activated Receptor (PPAR)-Alpha: A Pharmacological Target with a Promising Future. Pharm. Res. 2004, 21, 1531–1538. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, X.; Yu, S.; Zheng, R. The Leptin Resistance. Adv. Exp. Med. Biol. 2018, 1090, 145–163. [Google Scholar] [CrossRef]
- Tan, Q.; Hu, J.; Yu, X.; Guan, W.; Lu, H.; Yu, Y.; Yu, Y.; Zang, G.; Tang, Z. The Role of IL-1 Family Members and Kupffer Cells in Liver Regeneration. Biomed Res. Int. 2016, 2016, 6495793. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia Muciniphila Is a Promising Probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [Green Version]
- Ding, R.-X.; Goh, W.-R.; Wu, R.-N.; Yue, X.-Q.; Luo, X.; Khine, W.W.T.; Wu, J.-R.; Lee, Y.-K. Revisit Gut Microbiota and Its Impact on Human Health and Disease. J. Food Drug Anal. 2019, 27, 623–631. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut Microbiota and Human NAFLD: Disentangling Microbial Signatures from Metabolic Disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef]
- Islam, S.U. Clinical Uses of Probiotics. Medicine 2016, 95, e2658. [Google Scholar] [CrossRef]
- Steer, T.; Carpenter, H.; Tuohy, K.; Gibson, G.R. Perspectives on the Role of the Human Gut Microbiota and Its Modulation by Pro- and Prebiotics. Nutr. Res. Rev. 2000, 13, 229–254. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Yang, Z.; Chen, X.; Huang, Y.; Yin, B.; Guo, F.; Zhao, H.; Zhao, T.; Qu, H.; Huang, J.; et al. The Effect of Lactobacillus Rhamnosus Hsryfm 1301 on the Intestinal Microbiota of a Hyperlipidemic Rat Model. BMC Complement. Altern. Med. 2014, 14, 386. [Google Scholar] [CrossRef] [Green Version]
- Bagarolli, R.A.; Tobar, N.; Oliveira, A.G.; Araújo, T.G.; Carvalho, B.M.; Rocha, G.Z.; Vecina, J.F.; Calisto, K.; Guadagnini, D.; Prada, P.O.; et al. Probiotics Modulate Gut Microbiota and Improve Insulin Sensitivity in DIO Mice. J. Nutr. Biochem. 2017, 50, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Zeng, X.; Yang, F.; Liu, H.; Qiao, S. Study and Use of the Probiotic Lactobacillus reuteri in Pigs: A Review. J. Anim. Sci. Biotechnol. 2015, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Dore, M.P.; Cuccu, M.; Pes, G.M.; Manca, A.; Graham, D.Y. Lactobacillus reuteri in the Treatment of Helicobacter Pylori Infection. Intern. Emerg. Med. 2014, 9, 649–654. [Google Scholar] [CrossRef]
- Dore, M.P.; Bibbò, S.; Loria, M.; Salis, R.; Manca, A.; Pes, G.M.; Graham, D.Y. Twice-a-Day PPI, Tetracycline, Metronidazole Quadruple Therapy with Pylera® or Lactobacillus reuteri for Treatment Naïve or for Retreatment of Helicobacter Pylori. Two Randomized Pilot Studies. Helicobacter 2019, 24, e12659. [Google Scholar] [CrossRef]
- Hsieh, F.-C.; Lee, C.-L.; Chai, C.-Y.; Chen, W.-T.; Lu, Y.-C.; Wu, C.-S. Oral Administration of Lactobacillus Reuteri GMNL-263 Improves Insulin Resistance and Ameliorates Hepatic Steatosis in High Fructose-Fed Rats. Nutr. Metab. 2013, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasani, A.; Ebrahimzadeh, S.; Hemmati, F.; Khabbaz, A.; Hasani, A.; Gholizadeh, P. The Role of Akkermansia Muciniphila in Obesity, Diabetes and Atherosclerosis. J. Med. Microbiol. 2021, 70, 001435. [Google Scholar] [CrossRef] [PubMed]
- A Potential Probiotic- Lachnospiraceae NK4A136 Group: Evidence from the Restoration of the Dietary Pattern from a High-Fat Diet. Available online: https://www.researchsquare.com (accessed on 18 October 2022).
- Mota de Sá, P.; Richard, A.J.; Hang, H.; Stephens, J.M. Transcriptional Regulation of Adipogenesis. Compr. Physiol. 2017, 7, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.-M.; Han, S.-K.; Kim, J.-K.; Oh, S.-J.; Jang, H.-B.; Kim, D.-H. Lactobacillus Sakei Alleviates High-Fat-Diet-Induced Obesity and Anxiety in Mice by Inducing AMPK Activation and SIRT1 Expression and Inhibiting Gut Microbiota-Mediated NF-ΚB Activation. Mol. Nutr. Food Res. 2019, 63, e1800978. [Google Scholar] [CrossRef]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut Microbial Metabolites in Obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Rotman, Y.; Sanyal, A.J. Current and Upcoming Pharmacotherapy for Non-Alcoholic Fatty Liver Disease. Gut 2017, 66, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Delzenne, N.M. The Role of the Gut Microbiota in Energy Metabolism and Metabolic Disease. Curr. Pharm. Des. 2009, 15, 1546–1558. [Google Scholar] [CrossRef] [Green Version]
- Overview of Steroidogenic Enzymes in the Pathway from Cholesterol to Active Steroid Hormones|Endocrine Reviews|Oxford Academic. Available online: https://academic.oup.com/edrv/article/25/6/947/2195014 (accessed on 15 June 2022).
- Van der Wulp, M.Y.M.; Verkade, H.J.; Groen, A.K. Regulation of Cholesterol Homeostasis. Mol. Cell. Endocrinol. 2013, 368, 1–16. [Google Scholar] [CrossRef]
- Amir, M.; Czaja, M.J. Autophagy in Nonalcoholic Steatohepatitis. Expert Rev. Gastroenterol. Hepatol. 2011, 5, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yu, X.-H.; Ou, X.; Ouyang, X.-P.; Tang, C.-K. Hepatic Cholesterol Transport and Its Role in Non-Alcoholic Fatty Liver Disease and Atherosclerosis. Prog. Lipid Res. 2021, 83, 101109. [Google Scholar] [CrossRef]
- Hayek, T.; Ito, Y.; Azrolan, N.; Verdery, R.B.; Aalto-Setälä, K.; Walsh, A.; Breslow, J.L. Dietary Fat Increases High Density Lipoprotein (HDL) Levels Both by Increasing the Transport Rates and Decreasing the Fractional Catabolic Rates of HDL Cholesterol Ester and Apolipoprotein (Apo) A-I. Presentation of a New Animal Model and Mechanistic Studies in Human Apo A-I Transgenic and Control Mice. J. Clin. Investig. 1993, 91, 1665–1671. [Google Scholar] [CrossRef] [Green Version]
- Palhinha, L.; Liechocki, S.; Hottz, E.D.; da Silva Pereira, J.A.; de Almeida, C.J.; Moraes-Vieira, P.M.M.; Bozza, P.T.; Maya-Monteiro, C.M. Leptin Induces Proadipogenic and Proinflammatory Signaling in Adipocytes. Front. Endocrinol. 2019, 10, 841. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Cortegana, C.; García-Galey, A.; Tami, M.; Del Pino, P.; Carmona, I.; López, S.; Alba, G.; Sánchez-Margalet, V. Role of Leptin in Non-Alcoholic Fatty Liver Disease. Biomedicines 2021, 9, 762. [Google Scholar] [CrossRef]
- Régnier, M.; Polizzi, A.; Smati, S.; Lukowicz, C.; Fougerat, A.; Lippi, Y.; Fouché, E.; Lasserre, F.; Naylies, C.; Bétoulières, C.; et al. Hepatocyte-Specific Deletion of Pparα Promotes NAFLD in the Context of Obesity. Sci. Rep. 2020, 10, 6489. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Hong, F.; Wang, J.; Wang, J.; Zhao, X.; Wang, S.; Xue, T.; Xu, J.; Zheng, X.; Zhai, Y. DBZ Is a Putative PPARγ Agonist That Prevents High Fat Diet-Induced Obesity, Insulin Resistance and Gut Dysbiosis. Biochim. Biophys Acta Gen. Subj. 2017, 1861, 2690–2701. [Google Scholar] [CrossRef]
- Schlaepfer, I.R.; Joshi, M. CPT1A-Mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef]
- Lundsgaard, A.-M.; Fritzen, A.M.; Kiens, B. Molecular Regulation of Fatty Acid Oxidation in Skeletal Muscle during Aerobic Exercise. Trends Endocrinol. Metab. 2018, 29, 18–30. [Google Scholar] [CrossRef]
- Brownsey, R.W.; Boone, A.N.; Elliott, J.E.; Kulpa, J.E.; Lee, W.M. Regulation of Acetyl-CoA Carboxylase. Biochem. Soc. Trans. 2006, 34, 223–227. [Google Scholar] [CrossRef]
- Savari, F.; Badavi, M.; Rezaie, A.; Gharib-Naseri, M.K.; Mard, S.A. Evaluation of the Therapeutic Potential Effect of Fas Receptor Gene Knockdown in Experimental Model of Non-Alcoholic Steatohepatitis. Free Radic. Res. 2019, 53, 486–496. [Google Scholar] [CrossRef]
- Kohjima, M.; Higuchi, N.; Kato, M.; Kotoh, K.; Yoshimoto, T.; Fujino, T.; Yada, M.; Yada, R.; Harada, N.; Enjoji, M.; et al. SREBP-1c, Regulated by the Insulin and AMPK Signaling Pathways, Plays a Role in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Med. 2008, 21, 507–511. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the Complete Program of Cholesterol and Fatty Acid Synthesis in the Liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Mirea, A.-M.; Tack, C.J.; Chavakis, T.; Joosten, L.A.B.; Toonen, E.J.M. IL-1 Family Cytokine Pathways Underlying NAFLD: Towards New Treatment Strategies. Trends Mol. Med. 2018, 24, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Cahill, C.M.; Rogers, J.T. Interleukin (IL) 1β Induction of IL-6 Is Mediated by a Novel Phosphatidylinositol 3-Kinase-Dependent AKT/IκB Kinase α Pathway Targeting Activator Protein-1. J. Biol. Chem. 2008, 283, 25900–25912. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer | Sequence (5′ to 3′) |
|---|---|---|
| FAS | Forward | AGGGGTCGACCTGGTCCTCA |
| Reverse | GCCATGCCCAGAGGGTGGTT | |
| ACC | Forward | AACATCCCGCACCTTCTTCTAC |
| Reverse | CTTCCACAAACCAGCGTCTC | |
| PPARα | Forward | AGAGCCCCATCTGTCCTCTC |
| Reverse | ACTGGTAGTCTGCAAAACCAAA | |
| CPT1A | Forward | TGGCATCATCACTGGTGTGTT |
| Reverse | GTCTAGGGTCCGATTGATCTTTG | |
| IL-6 | Forward | ACAACCACGGCCTTCCCTACTT |
| Reverse | CACGATTTCCCAGAGAACATGTG | |
| β-actin | Forward | ACAACCACGGCCTTCCCTACTT |
| Reverse | CACGATTTCCCAGAGAACATGTG |
| Safety Test | L. reuteri MJM60668 (MJM60668) | LGG |
|---|---|---|
| Antibiotics * | ||
| Ampicilin | 1 | 1 |
| Vancomycin | 512 (NR) | 512 (NR) |
| Gentamicin | 8 (R) | 32 (R) |
| Kanamycin | R | R |
| Streptomycin | 128 (R) | 32 (R) |
| Tetracycline | 16 | 1 |
| Clindamycin | 1 | 1 |
| Erythromycin | 1 | 1 |
| Chloramphenicol | 4 | 4 |
| D-lactate production | - | - |
| Bile salt deconjugation | - | - |
| Bioamin production | - | - |
| L-Histidine | - | - |
| L-Tyrosine | - | - |
| L-phenylalanine | - | - |
| Arginine | - | - |
| Tryptophan | - | - |
| L-ornithine | - | - |
| Mucin degradation | - | - |
| Hemolytic activity Adhesion activity | - 5.1 ± 0.43% | - 3.1 ± 0.29% |
| Strains | Diameter of Zone Inhibition (mm) | |
|---|---|---|
| L.ruteri MJM60668 | LGG | |
| Salmonella gallirarum KCTC 2931 | 10 | 10 |
| Escherichia coli K99 | 8 | 8 |
| Escherichia coli O1 KCTC 2441 | 10 | 8 |
| Escherichia coli 0138 | 6 | 6 |
| Escherichia coli ATCC25922 | 8 | 8 |
| Salmonella chloreraesuis KCTC 2932 | 10 | 8 |
| Salmonella typhi KCTC 2514 | 8 | 8 |
| Pseudomonas aeruginosa KCCM 11802 | 10 | 10 |
| OGI Transit | (Log10 CFU/mL) | ||
|---|---|---|---|
| MJM60668 | LGG | ||
| Initinal | 9.20 ± 0.048 | 9.12 ± 0.165 | |
| Oral stress | - | 9.19 ± 0.04 | 9.09 ± 0.039 |
| + | 9.09 ± 0.1 | 9.00 ± 0.006 | |
| Gastric stress (pH3) | - | 9.16 ± 0.06 | 9.07 ± 0.026 |
| + | 9.06 ± 0.042 | 8.84 ± 0.052 | |
| Gastric stress (pH2) | - | 9.18 ± 0.053 | 9.05 ± 0.007 |
| + | 8.97 ± 0.027 | 8.74 ± 0.061 | |
| Intestinal stress | - | 9.15 ± 0.009 | 9.08 ± 0.092 |
| + | 8.68± 0.006 | 7.79 ± 0.095 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werlinger, P.; Nguyen, H.T.; Gu, M.; Cho, J.-H.; Cheng, J.; Suh, J.-W. Lactobacillus reuteri MJM60668 Prevent Progression of Non-Alcoholic Fatty Liver Disease through Anti-Adipogenesis and Anti-Inflammatory Pathway. Microorganisms 2022, 10, 2203. https://doi.org/10.3390/microorganisms10112203
Werlinger P, Nguyen HT, Gu M, Cho J-H, Cheng J, Suh J-W. Lactobacillus reuteri MJM60668 Prevent Progression of Non-Alcoholic Fatty Liver Disease through Anti-Adipogenesis and Anti-Inflammatory Pathway. Microorganisms. 2022; 10(11):2203. https://doi.org/10.3390/microorganisms10112203
Chicago/Turabian StyleWerlinger, Pia, Huong Thi Nguyen, Mingkun Gu, Joo-Hyung Cho, Jinhua Cheng, and Joo-Won Suh. 2022. "Lactobacillus reuteri MJM60668 Prevent Progression of Non-Alcoholic Fatty Liver Disease through Anti-Adipogenesis and Anti-Inflammatory Pathway" Microorganisms 10, no. 11: 2203. https://doi.org/10.3390/microorganisms10112203
APA StyleWerlinger, P., Nguyen, H. T., Gu, M., Cho, J. -H., Cheng, J., & Suh, J. -W. (2022). Lactobacillus reuteri MJM60668 Prevent Progression of Non-Alcoholic Fatty Liver Disease through Anti-Adipogenesis and Anti-Inflammatory Pathway. Microorganisms, 10(11), 2203. https://doi.org/10.3390/microorganisms10112203