
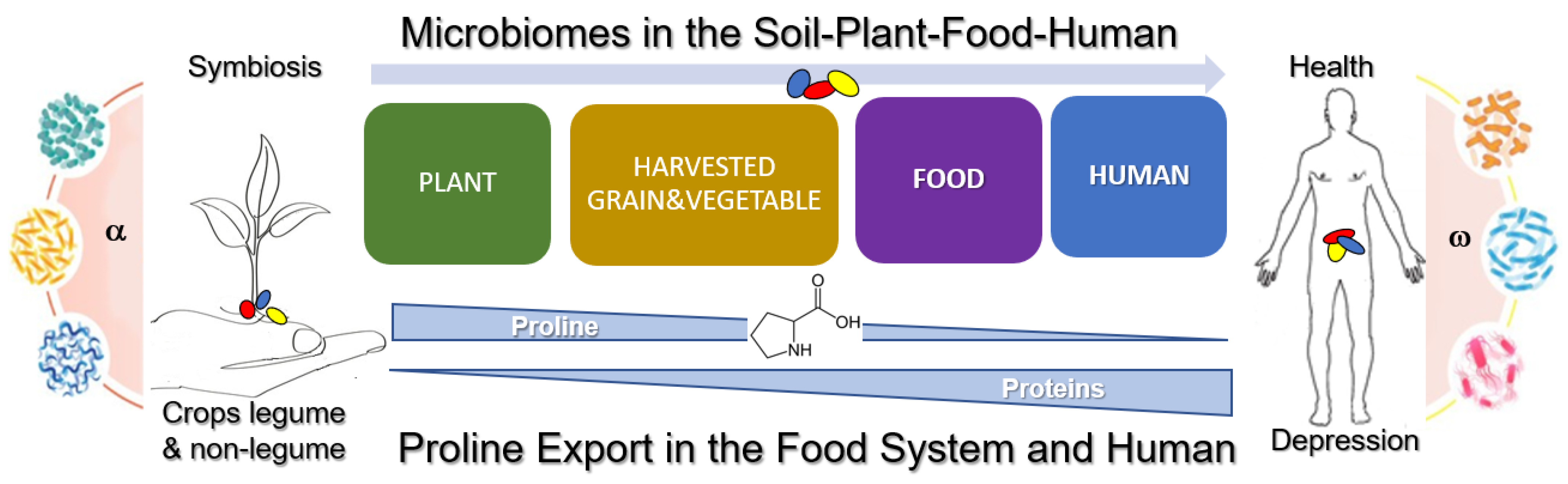
Microbiome-Driven Proline Biogenesis in Plants under Stress: Perspectives for Balanced Diet to Minimize Depression Disorders in Humans

{kind=link}
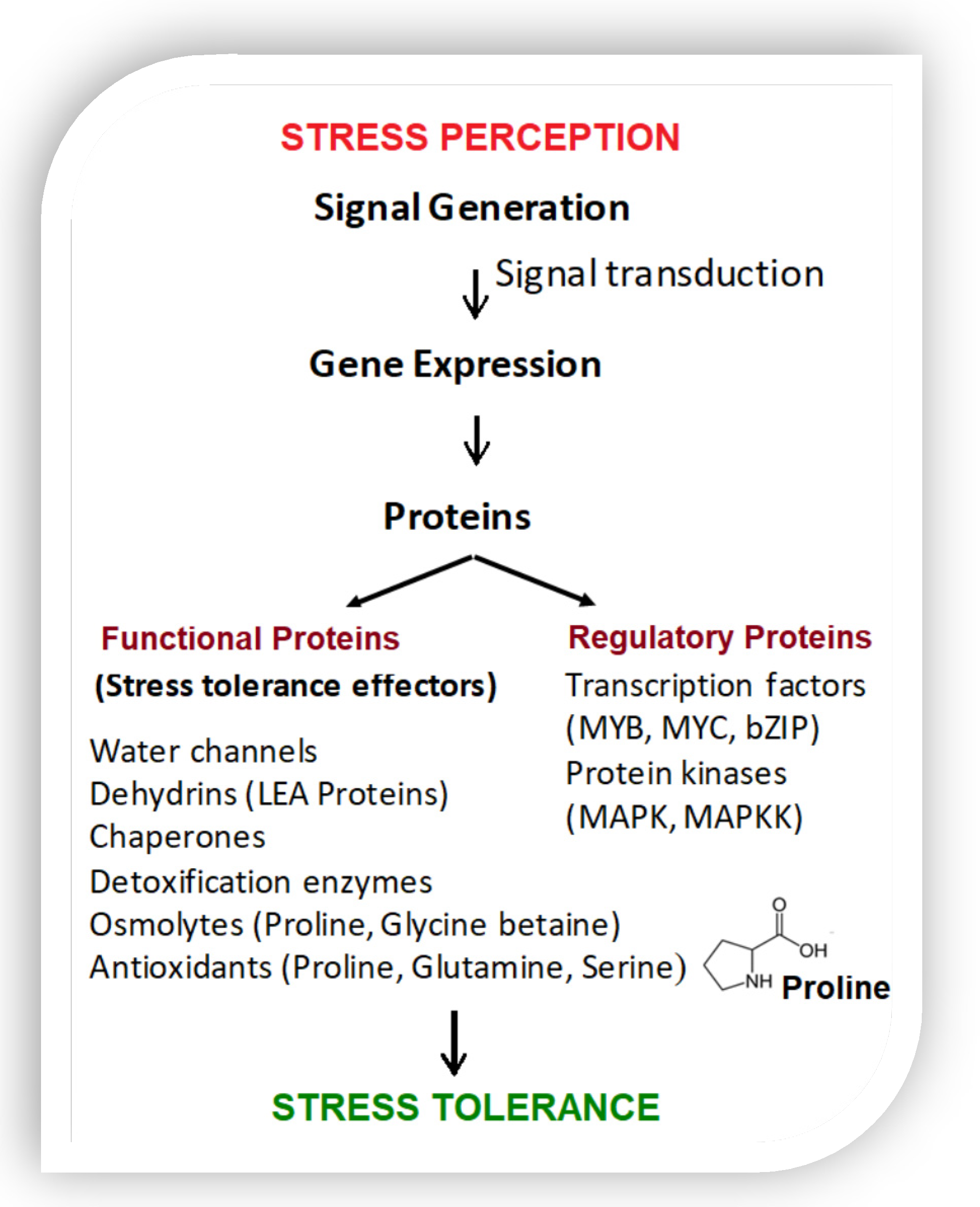{kind=link}
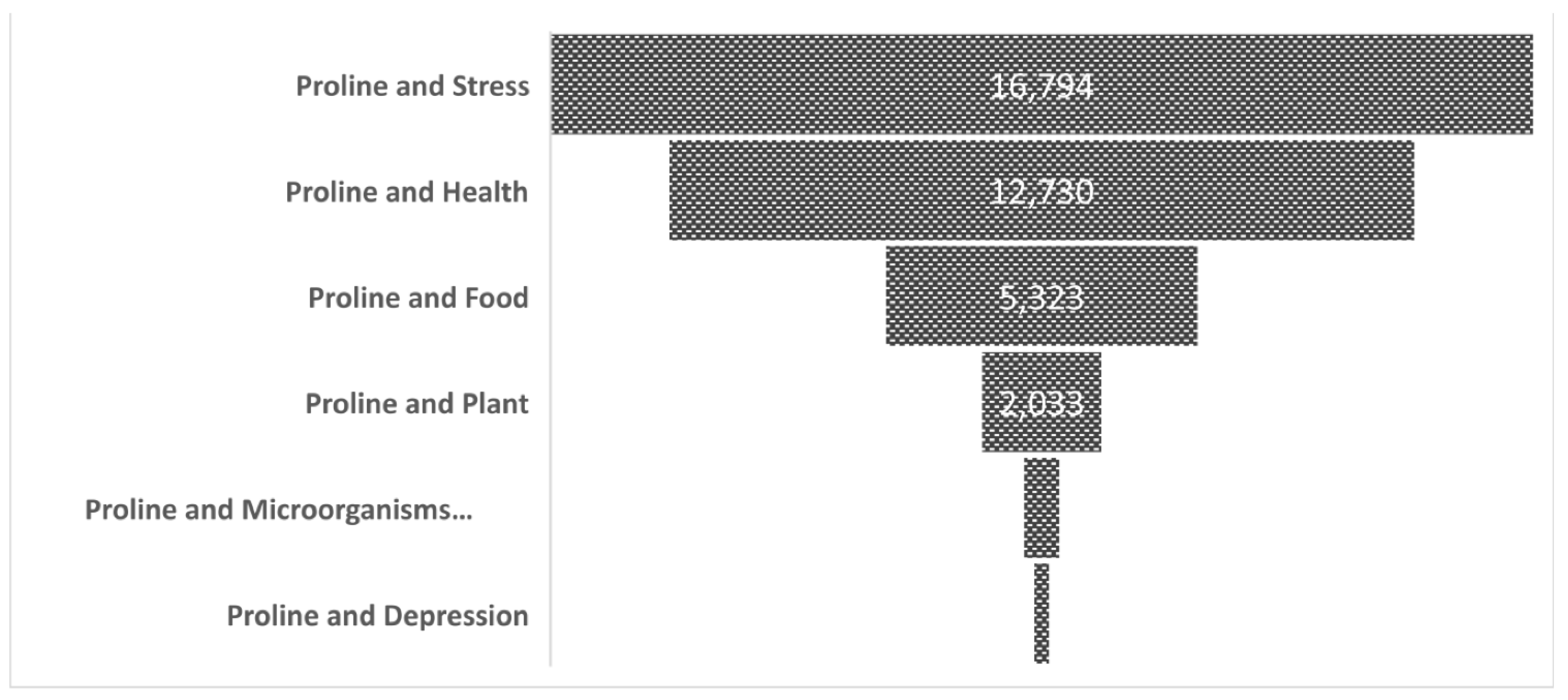{kind=link}
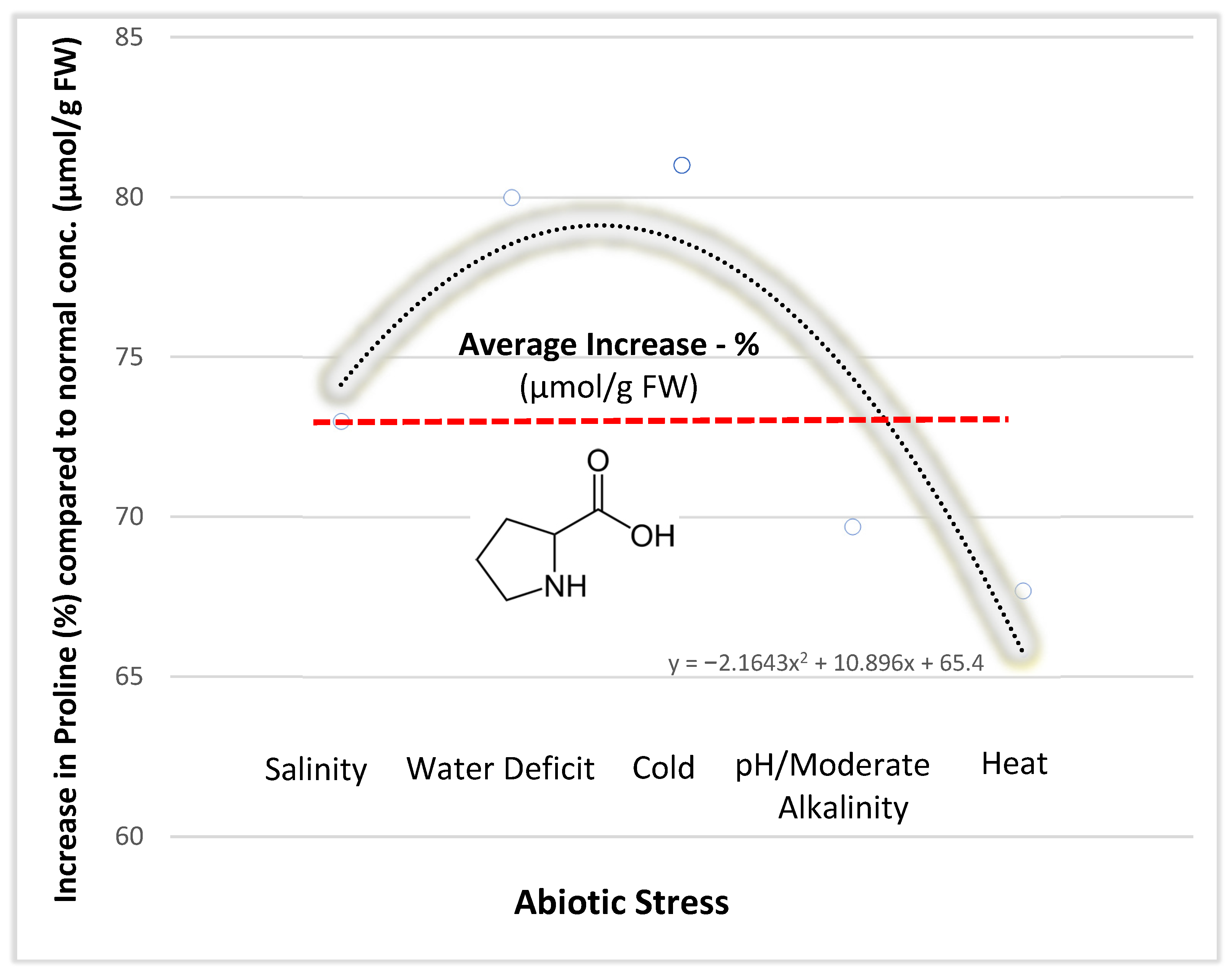{kind=link}
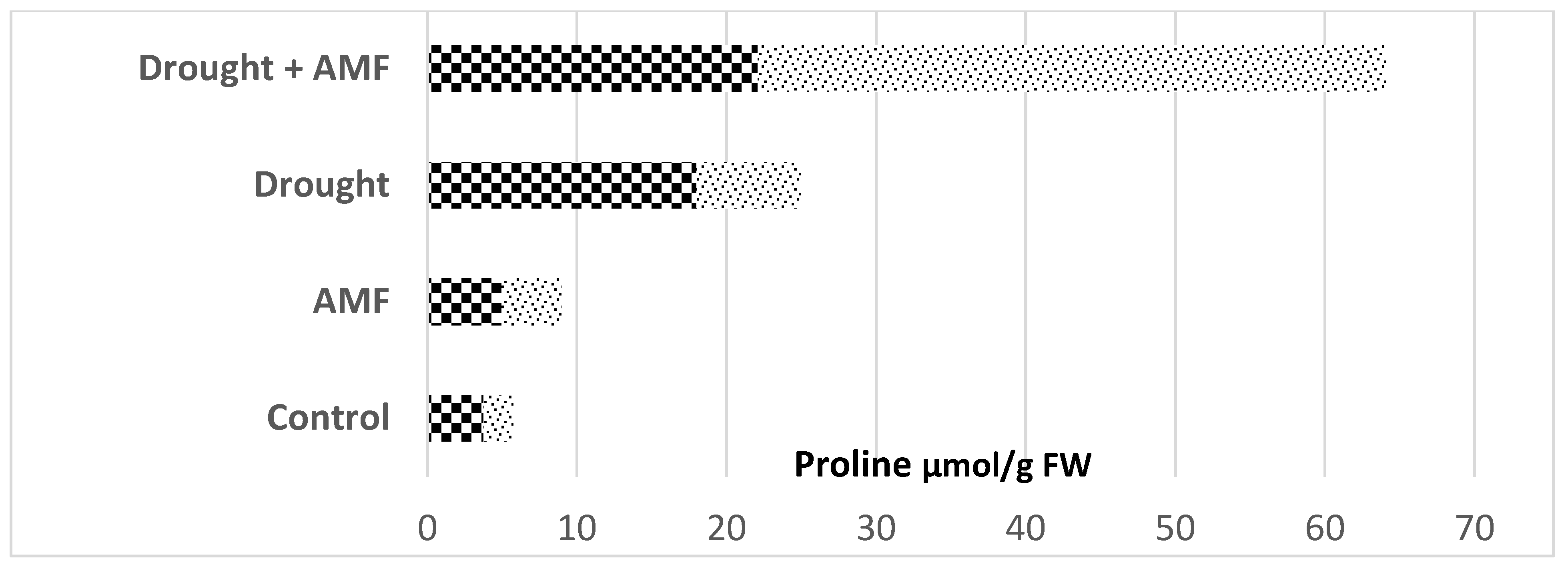{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Proline: Contradictory Roles in Healing and Excitotoxin Functions
3. Where Lie the Opportunities for Proline Science
4. Proline: Microbiome–Plant Axis
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mayneris-Perxachs, J.; Castells-Nobau, A.; Arnoriaga-Rodríguez, M.; Martin, M.; de la Vega-Correa, L.; Zapata, C.; Burokas, A.; Blasco, G.; Coll, C.; Escrichs, A.; et al. Microbiota alterations in proline metabolism impact depression. Cell Metab. 2022, 34, 681–701.e10. [Google Scholar] [CrossRef]
- Di Giulio, M. An extension of the coevolution theory of the origin of the genetic code. Biol. Direct 2008, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Trifanov, E.N. The triplet code from first principles. J. Biomol. Struct. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; 352p. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Beck, E.H.; Fettig, S.; Knake, C.; Hartig, K.; Bhattarai, T. Specific and Unspecific Responses of Plants to Cold and Drought Stress. J. Biosci. 2007, 32, 501–510. [Google Scholar] [CrossRef]
- Inbaraj, M.P. Plant-Microbe Interactions in Alleviating Abiotic Stress—A Mini Review. Front. Agron. 2021, 3, 667903. [Google Scholar] [CrossRef]
- Stec, N.; Saleem, A.; Darveau, C.-A. Proline as a Sparker Metabolite of Oxidative Metabolismduring the Flight of the Bumblebee, Bombus impatiens. Metabolites 2021, 11, 511. [Google Scholar] [CrossRef]
- Teulier, L.; Weber, J.-M.; Crevier, J.; Darveau, C.-A. Proline as a fuel for insect flight: Enhancing carbohydrate oxidation in hymenopterans. Proc. R. Soc. B 2016, 283, 20160333. [Google Scholar] [CrossRef]
- Beykaya, M. Determination of Physiochemical Properties of Raw Honey Samples. Prog. Nutr. 2021, 23, e2021020. Available online: https://www.mattioli1885journals.com/index.php/progressinnutrition/article/view/9510 (accessed on 21 June 2022).
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 2008, 44, 671–681. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Dickman, M.B. Proline suppresses apoptosis in the fungal pathogen of Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [Green Version]
- Vance, L.A.; Mukherjee, K.; Barbul, A. Proline Precursors and Collagen Synthesis: Biochemical Challenges of Nutrient Supplementation and Wound Healing. J. Nutr. 2017, 147, 2011–2017. [Google Scholar] [CrossRef] [Green Version]
- Henzi, V.; Reichling, D.B.; Helm, S.W.; MacDermott, A.B. L-proline activates glutamate and glycine receptors in cultured rat dorsal horn neurons. Mol. Pharmacol. 1992, 41, 793–801. [Google Scholar]
- Arslan, O.E. Neuroanatomical Basis of Clinical Neurology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2014; p. 309. ISBN 978-1-4398-4833-3. [Google Scholar]
- Shimada, A.; Cairns, B.E.; Vad, N.; Ulriksen, K.; Pedersen, A.M.L.; Svensson, P.; Baad-Hansen, L. Headache and mechanical sensitization of human pericranial muscles after repeated intake of monosodium glutamate (MSG). J. Headache Pain 2013, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Kishor, P.B.K.; Suravajhala, P.; Rathnagiri, P.; Sreenivasulu, N. Intriguing Role of Proline in Redox Potential Conferring High Temperature Stress Tolerance. Front. Plant Sci. 2022, 13, 867531. [Google Scholar] [CrossRef]
- Vujanovic, V.; Korber, D.R.; Vujanovic, S.; Vujanovic, J.; Jabaji, S. Scientific Prospects for Cannabis-Microbiome Research to Ensure Quality and Safety of Products. Microorganisms 2020, 8, 290. [Google Scholar] [CrossRef] [Green Version]
- Christgen, S.L.; Becker, D.F. Role of Proline in Pathogen and Host Interactions. Antioxid. Redox Signal. 2019, 30, 683–709. [Google Scholar] [CrossRef]
- Csonka, L.N.; Hanson, A.D. Prokaryotic osmoregulation: Genetics and physiology. Annu. Rev. Microbiol. 1991, 45, 569–606. [Google Scholar] [CrossRef]
- Roy, D.; Basu, N.; Bhunia, A.; Banerjee, S. Counteraction of exogenous L-proline with NaCl in salt-sensitive cultivar of rice. Biol. Plant. 1993, 35, 69–72. [Google Scholar] [CrossRef]
- Ondrasek, G.; Rathod, S.; Manohara, K.K.; Gireesh, C.; Anantha, M.S.; Sakhare, A.S.; Parmar, B.; Yadav, B.K.; Bandumula, N.; Raihan, F.; et al. Salt Stress in Plants and Mitigation Approaches. Plants 2022, 11, 717. [Google Scholar] [CrossRef]
- Naidu, B.P.; Paleg, L.G.; Aspinall, D.; Jennings, A.C.; Jones, G.P. Amino Acid and Glycine Betaine Accumulation in Cold Stressed Wheat Seedlings. Phytochemistry 1991, 30, 407–409. [Google Scholar] [CrossRef]
- El-Shintinawy, F.; El-Shourbagy, M.N. Alleviation of Changes in Protein Metabolism in NaCl-Stressed Wheat Seedlings by Thiamine. Biol. Plant. 2001, 44, 541–545. [Google Scholar] [CrossRef]
- Misra, N.; Saxen, P. Effect of Salicylic Acid on Proline Metabolism in Lentil Grown under Salinity Stress. Plant Sci. 2009, 177, 181–189. [Google Scholar] [CrossRef]
- Johari-Pireivatlou, M. Effect of soil water stress on yield and proline content of four wheat lines. Afr. J. Biotechnol. 2010, 9, 36–40. Available online: http://www.academicjournals.org/AJB (accessed on 21 October 2022).
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Xia, X.; Li, H.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef] [Green Version]
- Janda1, T.; Khalil, R.; Tajti, J.; Pál, M.; Darkó, E. Responses of young wheat plants to moderate heat stress. Acta Physiol. 2019, 41, 137. [Google Scholar] [CrossRef] [Green Version]
- Kishor, P.B.K.; Kumari, P.H.; Sunita, M.S.L.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Bandurska, H.; Niedziela, J.; Pietrowska-Borek, M.; Nuc, K.; Chadzinikolau, T.; Radzikowska, D. Regulation of proline biosynthesis and resistance to drought stress in two barley (Hordeum vulgare L.) genotypes of different origin. Plant Physiol. Biochem. 2017, 118, 427–437. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, O.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, R.S.M. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, V.H.; Lee, B.R.; Islam, T.; Park, S.H.; Jung, H.I.; Bae, D.W.; Kim, T.-H. Characterization of salicylic acid-mediated modulation of the drought stress responses: Reactive oxygen species, proline, and redox state in Brassica napus. Environ. Exp. Bot. 2020, 157, 512. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline Metabolism and Its Implications for Plant-Environment Interaction. Arabidopsis Book 2010, 8, e0140. [Google Scholar] [CrossRef] [Green Version]
- Bhaskara, G.B.; Yang, T.-H.; Verslues, P.E. Dynamic proline metabolism: Importance and regulation in water limited environments. Front. Plant Sci. 2015, 6, 484. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef] [Green Version]
- Patriarca, E.J.; Cermola, F.; D’Aniello, C.; Fico, A.; Guardiola, O.; de Cesare, D.; Minchiotti, G. The Multifaceted Roles of Proline in Cell Behavior. Front. Cell Dev. Biol. 2021, 9, 728576. [Google Scholar] [CrossRef]
- Godard, T.; Zühlke, D.; Richter, G.; Wall, M.; Rohde, M.; Riedel, K.; Poblete-Castro, I.; Krull, R.; Biedendieck, R. Metabolic Rearrangements Causing Elevated Proline and Polyhydroxybutyrate Accumulation During the Osmotic Adaptation Response of Bacillus megaterium. Front. Bioeng. Biotechnol. 2020, 8, 47. [Google Scholar] [CrossRef]
- Chua, A.; Sherwood, O.L.; Fitzhenry, L.; Ng, C.K.-Y.; McCabe, P.F.; Daly, C.T. Cyanobacteria-Derived Proline Increases Stress Tolerance in Arabidopsis thaliana Root Hairs by Suppressing Programmed Cell Death. Front. Plant Sci. 2020, 11, 490075. [Google Scholar] [CrossRef]
- Poolman, B.; Glaasker, E. Regulation of compatible solute accumulation in bacteria. Mol. Microbiol. 1998, 29, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Baltrus, D.A. Adaptation, specialization, and coevolution within phytobiomes. Curr. Opin. Plant Biol. 2017, 38, 109–116. [Google Scholar] [CrossRef]
- Vujanovic, V.; Germida, J.J. Seed endosymbiosis: A vital relationship in providing prenatal care to plants. Can. J. Plant Sci. 2017, 97, 972–981. [Google Scholar] [CrossRef]
- Gordon, J.; Youle, M.; Knowlton, N.; Rohwer, F.; Relman, D.A. Superorganisms and Holobionts. Microbe 2013, 8, 152–153. [Google Scholar] [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–microbiome interactions under a changing world: Responses, consequences and perspectives. New Phytol. 2022, 234, 1903–2139. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones, and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in Chickpea (Cicer arietinum L.) Induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef] [Green Version]
- Yooyongwech, S.; Phaukinsang, N.; Cha-Um, S.; Supaibulwatana, K. Arbuscular mycorrhiza improved growth performance in Macadamia tetraphylla L. grown under water deficit stress involves soluble sugar and proline accumulation. Plant Growth Regul. 2013, 69, 285–293. [Google Scholar] [CrossRef]
- Wu, H.-H.; Zou, Y.N.; Rahman, M.M.; Ni, Q.-D.; Wu, Q.-S. Mycorrhizas alter sucrose and proline metabolism in trifoliate orange exposed to drought stress. Sci. Rep. 2017, 7, 42389. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, A.; Yoshikawa, Y.; Ichikawa, K.; Takemoto, T.; Tanahashi, R.; Takagi, H. Longevity Regulation by Proline Oxidation in Yeast. Microorganisms 2021, 9, 1650. [Google Scholar] [CrossRef]
- Chen, C.; Wanduragala, S.; Becker, D.F.; Dickman, M.A. Tomato QM-like protein protects Saccharomyces cerevisiae cells against oxidative stress by regulating intracellular proline levels. Appl. Environ. Microbiol. 2006, 72, 4001–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerswald, L.; Gäde, G. Energy substrates for flight in the blister beetle Decapotoma Lunata (meloidae). J. Exp. Biol. 1995, 198, 1423–1431. [Google Scholar] [CrossRef]
- Ghaffari, H.; Tadayon, M.R.; Bahador, M.; Razmjoo, J. Investigation of the proline role in controlling traits related to sugar and root yield of sugar beet under water deficit conditions. Agric. Water Manag. 2021, 243, 106448. [Google Scholar] [CrossRef]
- Jones, M.M.; Osmond, C.B.; Turner, N.C. Accumulation of solutes in leaves of sorghum and sunflower in response to water deficits. Aust. J. Plant Physiol. 1980, 7, 193–205. [Google Scholar] [CrossRef]
- Czipa, N.; Borbély, M.; Győri, Z. Proline content of different honey types. Acta Aliment. 2012, 41, 26–32. [Google Scholar] [CrossRef]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Lavy, O.; Gophna, U.; Gefen, E.; Ayali, A. Locust Bacterial Symbionts: An Update. Insects 2020, 11, 655. [Google Scholar] [CrossRef]
- Taraskina, A.; Ignatyeva, O.; Lisovaya, D.; Ivanov, M.; Ivanova, L.; Golovicheva, V.; Baydakova, G.; Silachev, D.; Popkov, V.; Ivanets, T.; et al. Effects of Traumatic Brain Injury on the Gut Microbiota Composition and Serum Amino Acid Profile in Rats. Cells 2022, 11, 1409. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [Green Version]
- Grondin, J.A.; Kwon, Y.H.; Far, P.M.; Haq, S.; Khan, W.I. Mucins in Intestinal Mucosal Defense and Inflammation: Learning from Clinical and Experimental Studies. Front. Immunol. 2020, 11, 2054. [Google Scholar] [CrossRef]
- Skotnicka, M.; Karwowska, K.; Kłobukowski, F.; Borkowska, A.; Pieszko, M. Possibilities of the Development of Edible Insect-Based Foods in Europe. Foods 2021, 10, 766. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.-N.; Wu, Q.-S.; Huang, Y.-M.; Ni, Q.-D.; He, X.-H. Mycorrhizal-Mediated Lower Proline Accumulation in Poncirus trifoliata under Water Deficit Derives from the Integration of Inhibition of Proline Synthesis with Increase of Proline Degradation. PLoS ONE 2013, 8, e80568. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.S.; Xia, R.X. Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthsis of citrus under well-watered and water stress conditions. J. Plant Physiol. 2006, 163, 417–425. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Collados, C.; Barea, J.M.; Azcón, R. Arbuscular mycorrhizal symbiosis can alleviate drought-induced nodule senescence in soybean plants. New Phytol. 2001, 151, 493–502. [Google Scholar] [CrossRef]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline Accumulation Influenced by Osmotic Stress in Arbuscular Mycorrhizal Symbiotic Plants. Front. Microbiol. 2018, 9, 2525. [Google Scholar] [CrossRef] [Green Version]
- Ellouze, W.; Hamel, C.; DePauw, R.M.; Knox, R.E.; Cuthbert, R.D.; Singh, A.K. Potential to breed for mycorrhizal association in durum wheat. Can. J. Microbiol. 2016, 62, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Vujanovic, V.; Germida, J. Endophytic Microbial Symbionts in Plant Prenatal Care. U.S. Patent PCT/CA2013/000091/US11076573B2, 3 August 2013. Available online: https://patents.google.com/patent/US11076573B2/en (accessed on 21 October 2022).
- Kumari, V.; Vujanovic, V. Transgenerational benefits of endophytes on resilience and antioxidant genes expressions in pea (Pisum sativum L.) under osmotic stress. Acta Physiol. Plant. 2020, 42, 49. [Google Scholar] [CrossRef]
- Kumari, V.; Germida, J.; Vujanovic, V. Legume endosymbionts: Drought stress tolerance in second-generation chickpea (Cicer arietinum) seeds. J. Agron. Crop Sci. 2018, 204, 529–540. [Google Scholar] [CrossRef]
- Vujanovic, V.; Islam, M.N.; Daida, P. Transgenerational role of seed mycobiome—An endosymbiotic fungal composition as a prerequisite to stress resilience and adaptive phenotypes in Triticum. Sci. Rep. 2019, 9, 18483. [Google Scholar] [CrossRef] [Green Version]
- Vujanovic, V.; Yuan, X.; Daida, P.; Milunovic, B.; Germida, J. Manipulation of cold stratification and endophytic effects on expression patterns of RSG and KAO genes in coleorhiza of wheat seeds. Plant Growth Regul. 2016, 79, 219–227. [Google Scholar] [CrossRef]
- Vujanovic, V.; Kim, S.H.; Lahlali, R.; Karunakaran, C. Spectroscopy and SEM imaging reveal endosymbiont-dependent components changes in germinating kernel through direct and indirect coleorhiza-fungus interactions under stress. Sci. Rep. 2019, 9, 1665. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lahlali, R.; Karunakaran, S.; Vujanovic, V. Specific mycoparasite-Fusarium graminearum molecular signatures in germinating seeds disabled Fusarium Head Blight pathogen’s infection. Int. J. Mol. Sci. 2021, 22, 2461. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, L.; Salamon, S.; Mikołajczak, K. Fungi Inhabiting the Wheat Endosphere. Pathogens 2021, 10, 1288. [Google Scholar] [CrossRef] [PubMed]
- Fried, E.I. Studying mental health problems as systems, not syndromes. PsyArXiv, 2022; preprint. [Google Scholar] [CrossRef]
- Kraft, S.; Buchenauer, L.; Polte, T. Mold, Mycotoxins and a Dysregulated Immune System: A combination of concern? Int. J. Mol. Sci. 2021, 22, 12269. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Zhang, W.; Jiang, H.; Liu, D.; Liu, L.; Li, L.; Li, C.; Xiao, X.; Tang, S.; Li, D. Food-Origin Mycotoxin-Induced Neurotoxicity: Intend to Break the Rules of Neuroglia Cells. Oxidative Med. Cell. Longev. 2021, 2021, 9967334. [Google Scholar] [CrossRef]
- Ratnaseelan, A.M.; Tsilioni, I.; Theoharides, T.C. Effects of Mycotoxins on Neuropsychiatric Symptoms and Immune Processes. Clin. Ther. 2018, 40, 903–917. Available online: https://pubmed.ncbi.nlm.nih.gov/29880330/ (accessed on 21 October 2022).
- Grenier, G.; Applegate, T.J. Modulation of Intestinal Functions Following Mycotoxin Ingestion: Meta-Analysis of Published Experiments in Animals toxins. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [Green Version]
- Omotayo, O.P.; Omotayo, A.O.; Mwanza, M.; Babalol, O.O. Prevalence of Mycotoxins and Their Consequences on Human Health. Toxicol. Res. 2019, 35, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Inamura, K.; Hamada, T.; Bullman, S.; Ugai, T.; Yachida, S.; Ogino, S. Cancer as microenvironmental, systemic and environmental diseases: Opportunity for transdisciplinary microbiomics science. Gut 2022, 71, 10. [Google Scholar] [CrossRef]
- Clark, D.P.; Schwartz, F.R.; Marin, D.; Ramirez-Giraldo, J.C.; Badea, C.T. Deep learning based spectral extrapolation for dual-source, dual-energy X-ray computed tomography. Med. Phys. 2020, 47, 4150–4163. [Google Scholar] [CrossRef]
- Odedra, D.; Narayanasamy, S.; Sabongui, S.; Priya, S.; Krishna, S.; Sheikh, A. Dual Energy CT Physics—A Primer for the Emergency Radiologist. Front. Radiol. 2022, 2, 820430. [Google Scholar] [CrossRef]
- Deng, K.; Sun, C.; Liu, C.; Ma, R. Initial experience with visualizing hand and foot tendons by dual-energy computed tomography. Clin. Imaging 2009, 33, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Zhang, C.; Liu, F.; Wang, W.; Lu, Y.; Chen, R.; He, Y. High-Throughput Screening of Free Proline Content in Rice Leaf under Cadmium Stress Using Hyperspectral Imaging with Chemometrics. Sensors 2020, 20, 3229. [Google Scholar] [CrossRef] [PubMed]
- Agriopoulou, S.; Stamatelopoulou, S.; Varzakas, T. Advances in Analysis and Detection of Major Mycotoxins in Foods. Foods 2020, 9, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, J.; Maye, D. What Are the Implications of Digitalisation for Agricultural Knowledge? Front. Sustain. Food Syst. 2020, 4, 66. [Google Scholar] [CrossRef]
- Gandhi, S.; Mosleh, W.; Shen, J.; Chow, C.M. Automation, machine learning, and artificial intelligence in echocardiography: A brave new world. Endocardiography 2018, 35, 1402–1418. [Google Scholar] [CrossRef]
- Parhi, R.; Nowak, R.D. What Kinds of Functions do Deep Neural Networks Learn? Insights from Variational Spline Theory. SIAM J. Math. Data Sci. 2022, 4, 464–489. [Google Scholar] [CrossRef]




 macadamia nut trees (Macadamia tetraphylla L.) and
macadamia nut trees (Macadamia tetraphylla L.) and  cucumber (Cuccumis sativus) plants (data from Hushem et al. [48] and Yooyongwech et al. [50]).
cucumber (Cuccumis sativus) plants (data from Hushem et al. [48] and Yooyongwech et al. [50]).
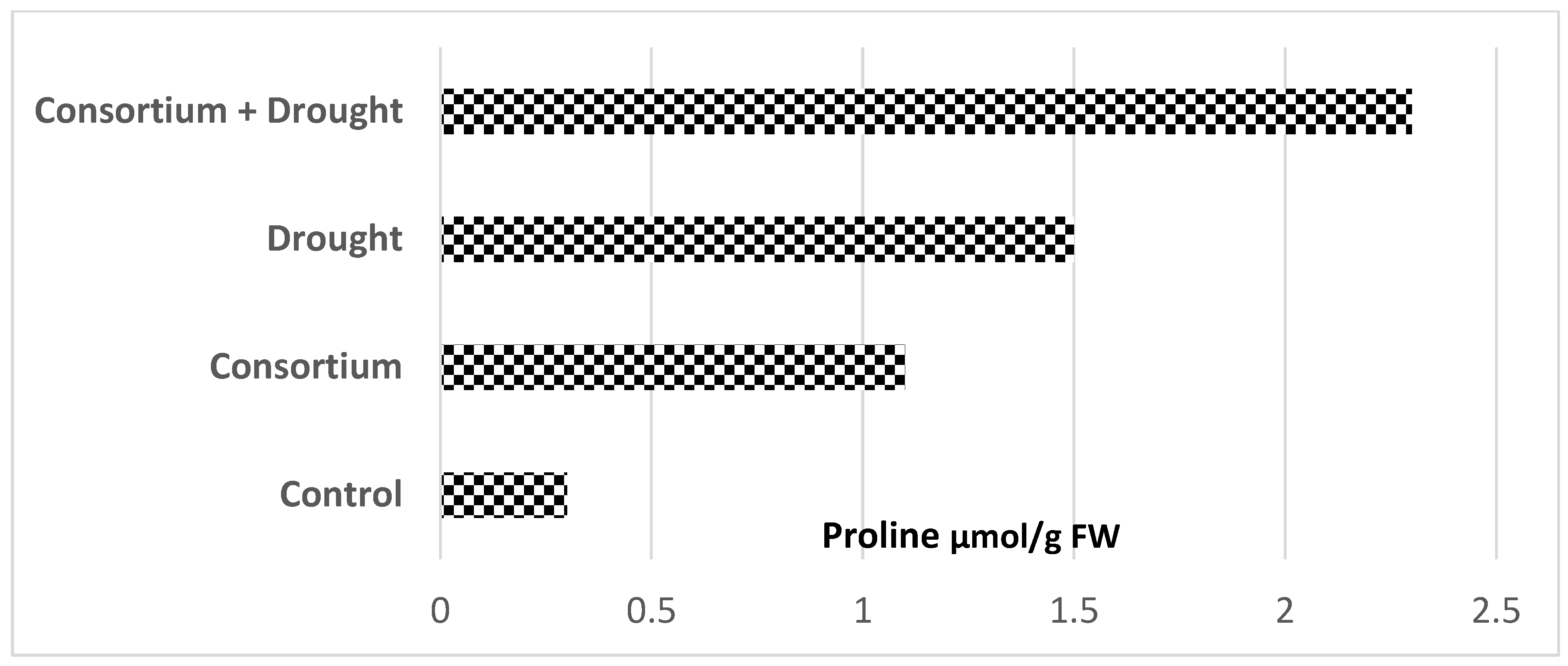
 chickpea (Cicer arietinum L.) cultivars (data from Khan et al. [49]).
chickpea (Cicer arietinum L.) cultivars (data from Khan et al. [49]).
chickpea (Cicer arietinum L.) cultivars (data from Khan et al. [49]).
chickpea (Cicer arietinum L.) cultivars (data from Khan et al. [49]).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujanovic, S.; Vujanovic, J.; Vujanovic, V. Microbiome-Driven Proline Biogenesis in Plants under Stress: Perspectives for Balanced Diet to Minimize Depression Disorders in Humans. Microorganisms 2022, 10, 2264. https://doi.org/10.3390/microorganisms10112264
Vujanovic S, Vujanovic J, Vujanovic V. Microbiome-Driven Proline Biogenesis in Plants under Stress: Perspectives for Balanced Diet to Minimize Depression Disorders in Humans. Microorganisms. 2022; 10(11):2264. https://doi.org/10.3390/microorganisms10112264
Chicago/Turabian StyleVujanovic, Silva, Josko Vujanovic, and Vladimir Vujanovic. 2022. "Microbiome-Driven Proline Biogenesis in Plants under Stress: Perspectives for Balanced Diet to Minimize Depression Disorders in Humans" Microorganisms 10, no. 11: 2264. https://doi.org/10.3390/microorganisms10112264
APA StyleVujanovic, S., Vujanovic, J., & Vujanovic, V. (2022). Microbiome-Driven Proline Biogenesis in Plants under Stress: Perspectives for Balanced Diet to Minimize Depression Disorders in Humans. Microorganisms, 10(11), 2264. https://doi.org/10.3390/microorganisms10112264