
Generation and Utilization of a Monoclonal Antibody against Hepatitis B Virus Core Protein for a Comprehensive Interactome Analysis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Construction of Wheat Germ Cell-Free Expression Vector
2.3. Cell-Free Protein Synthesis and Purification
2.4. Immunization and Generation of mAbs
2.5. Cell Culture
2.6. HBV Infection
2.7. Epitope Mapping and Specificity of mAb #7 Using AlphaScreen Assay
2.8. Bioinformatic Analysis
2.9. Immunoblot Analysis
2.10. Immunofluorescence Analysis
2.11. Immunoprecipitation Analysis
2.12. Immunohistochemistry Analysis
2.13. Antibody-Based In Situ Biotinylation
2.14. Sample Preparation for Proteomic Analysis
2.15. Proteomic Analysis and Data Analysis
2.16. Statistical Analysis
3. Results
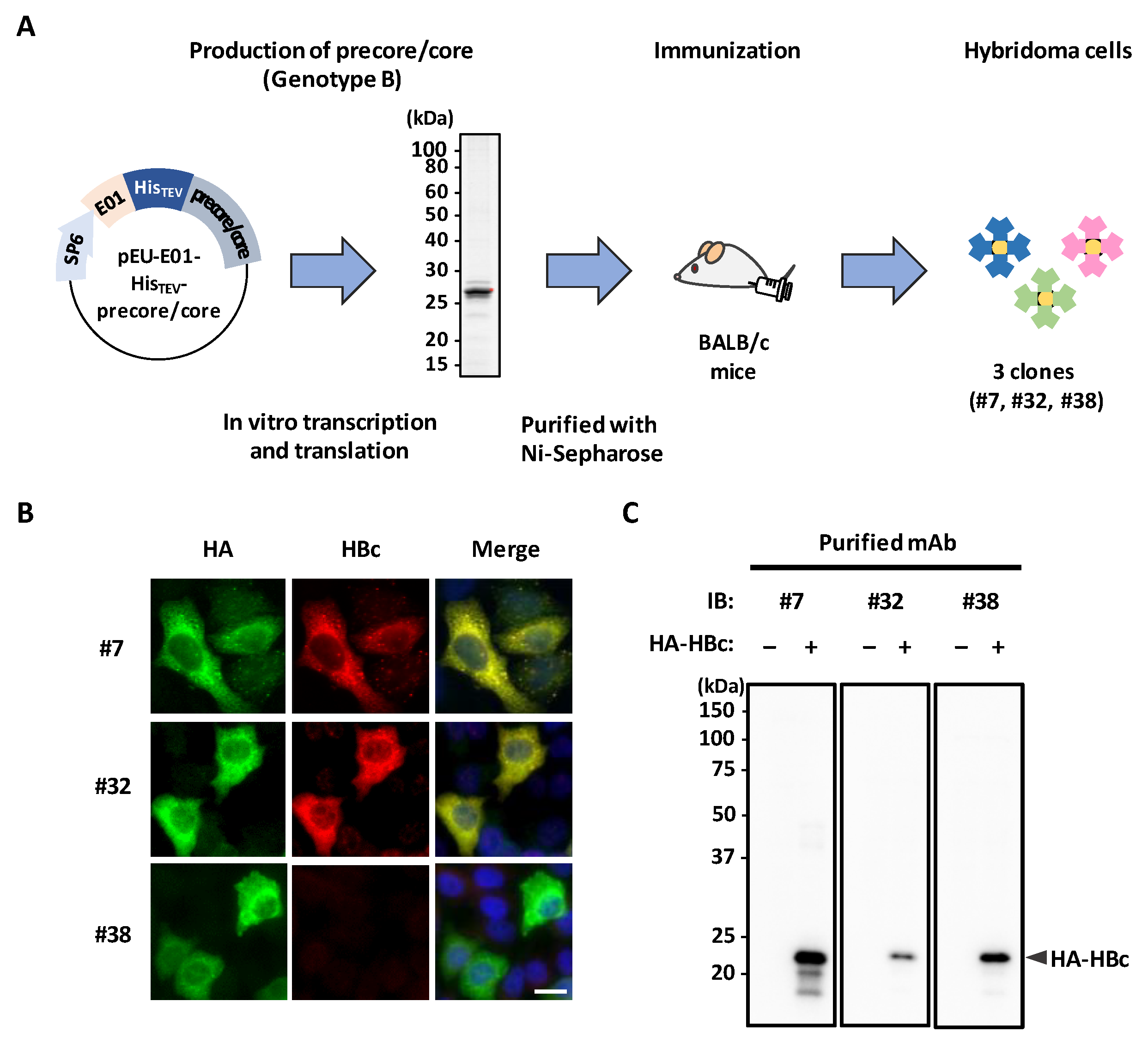
3.1. Generation of mAbs Targeting HBc Antigen
3.2. Detection of HBc Antigen in HBV Infected Cells
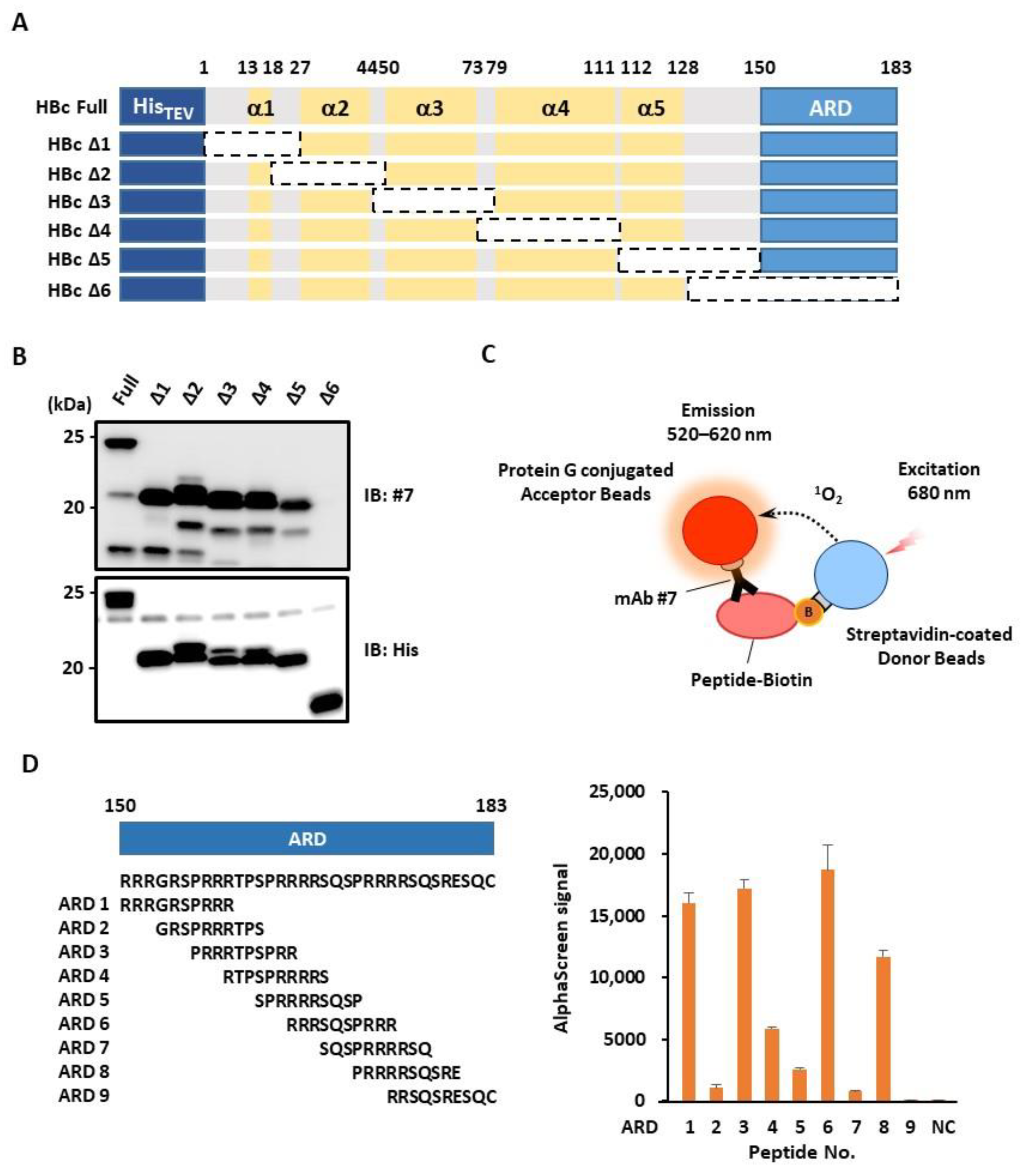
3.3. mAb #7 Recognizes the Arginine-Rich Domain (ARD) of HBc
3.4. mAb #7 Detects HBc Derived from Multiple Genotypes
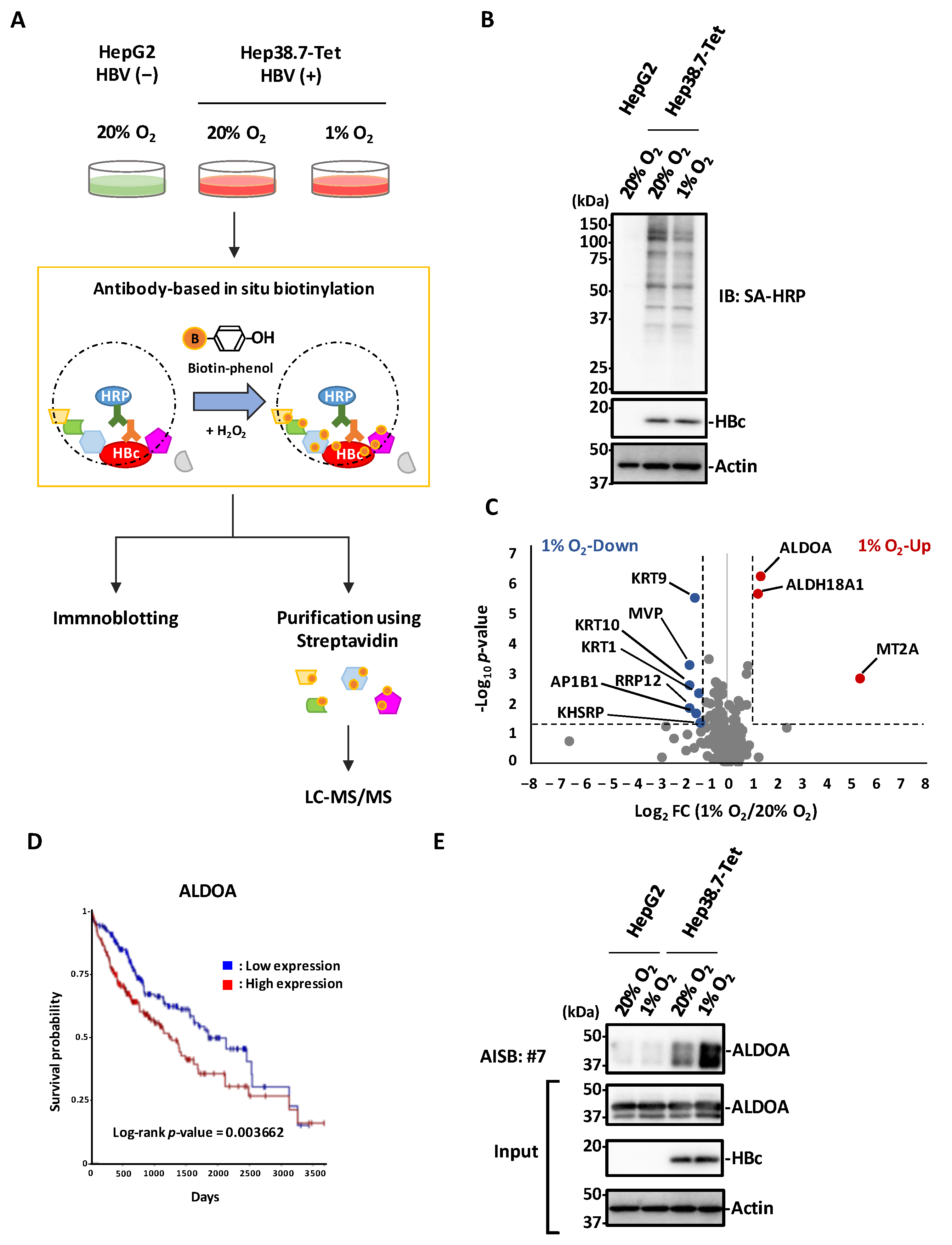
3.5. Antibody-Based In Situ Biotinylation with mAb #7
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Megahed, F.A.K.; Zhou, X.; Sun, P. The interactions between HBV and the innate immunity of hepatocytes. Viruses 2020, 12, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Arienzo, V.; Ferguson, J.; Giraud, G.; Chapus, F.; Harris, J.M.; Wing, P.A.C.; Claydon, A.; Begum, S.; Zhuang, X.; Balfe, P.; et al. The CCCTC-binding factor CTCF represses Hepatitis B virus enhancer I and regulates viral transcription. Cell. Microbiol. 2021, 23, e13274. [Google Scholar] [CrossRef] [PubMed]
- Virzi, A.; Motos, V.G.; Tripon, S.; Baumert, T.F.; Virzi, A.; Motos, V.G.; Tripon, S.; Baumert, T.F.; Profi-, J.L.; Virz, A.; et al. Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development To cite this version: HAL Id: Hal-03603544 Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development. J. Clin. Med. 2021, 10, 977. [Google Scholar] [PubMed]
- Fisman, D.N.; Agrawal, D.; Leder, K. The effect of age on immunologic response to recombinant hepatitis B vaccine: A meta-analysis. Clin. Infect. Dis. 2002, 35, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Pattyn, J.; Hendrickx, G.; Vorsters, A.; Van Damme, P. Hepatitis B Vaccines. J. Infect. Dis. 2021, 224 (Suppl. 4), S343–S351. [Google Scholar] [CrossRef]
- Lubyova, B.; Hodek, J.; Zabransky, A.; Prouzova, H.; Hubalek, M.; Hirsch, I.; Weber, J. PRMT5: A novel regulator of Hepatitis B virus replication and an arginine methylase of HBV core. PLoS ONE 2017, 12, e0186982. [Google Scholar] [CrossRef]
- Blondot, M.L.; Bruss, V.; Kann, M. Intracellular transport and egress of Hepatitis B virus. J. Hepatol. 2016, 64, S49–S59. [Google Scholar] [CrossRef]
- Basagoudanavar, S.H.; Perlman, D.H.; Hu, J. Regulation of Hepadnavirus Reverse Transcription by Dynamic Nucleocapsid Phosphorylation. J. Virol. 2007, 81, 1641–1649. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.H.; Li, Y.N.; Zhao, J.R.; Zhang, J.; Yan, Z. HBc binds to the CpG islands of HBV cccDNA and promotes an epigenetic permissive state. Epigenetics 2011, 6, 720–726. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Liang, X.; Liu, Y.; Qu, Z.; Gao, L.; Han, L.; Liu, S.; Cui, M.; Shi, Y.; Zhang, Z.; et al. Hepatitis B virus core protein inhibits TRAIL-induced apoptosis of hepatocytes by blocking DR5 expression. Cell Death Differ. 2009, 16, 219–229. [Google Scholar] [CrossRef]
- Xie, Q.; Fan, F.; Wei, W.; Liu, Y.; Xu, Z.; Zhai, L.; Qi, Y.; Ye, B.; Zhang, Y.; Basu, S.; et al. Multi-omics analyses reveal metabolic alterations regulated by Hepatitis B virus core protein in hepatocellular carcinoma cells. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dunham, W.H.; Mullin, M.; Gingras, A.C. Affinity-purification coupled to mass spectrometry: Basic principles and strategies. Proteomics 2012, 12, 1576–1590. [Google Scholar] [CrossRef] [PubMed]
- Sardiu, M.E.; Washburn, M.P. Building protein-protein interaction networks with proteomics and informatics tools. J. Biol. Chem. 2011, 286, 23645–23651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A.; Kuster, B. Affinity purification-mass spectrometry: Powerful tools for the characterization of protein complexes. Eur. J. Biochem. 2003, 270, 570–578. [Google Scholar] [CrossRef]
- Bussey, K.A.; Lau, U.; Schumann, S.; Gallo, A.; Osbelt, L.; Stempel, M.; Arnold, C.; Wissing, J.; Gad, H.H.; Hartmann, R.; et al. The Interferon-Stimulated Gene Product Oligoadenylate Synthetase-Like Protein Enhances Replication of Kaposi’s Sarcoma-Associated Herpesvirus (KSHV) and Interacts with the KSHV ORF20 Protein. PLoS Pathog. 2018, 14, e1006937. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, Q.; Zheng, M.; Wen, J.; Li, Q.; Zhao, G. Viral–Host Interactome Analysis Reveals Chicken STAU2 Interacts with Non-structural Protein 1 and Promotes the Replication of H5N1 Avian Influenza Virus. Front. Immunol. 2021, 12, 590679. [Google Scholar] [CrossRef]
- Shah, P.S.; Link, N.; Jang, G.M.; Sharp, P.P.; Zhu, T.; Swaney, D.L.; Johnson, J.R.; Von Dollen, J.; Ramage, H.R.; Satkamp, L.; et al. Comparative Flavivirus-Host Protein Interaction Mapping Reveals Mechanisms of Dengue and Zika Virus Pathogenesis. Cell 2019, 175, 1931–1945. [Google Scholar] [CrossRef] [Green Version]
- Roux, K.J.; Kim, D.I.; Raida, M.; Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Burke, B.E.; Roux, K.J.; Kim, D.I.; Burke, B.E. BioID: A Screen for Protein-Protein Interactions. Curr. Protoc. Protein Sci. 2018, 91, 19.23.1–19.23.15. [Google Scholar] [CrossRef]
- Bar, D.Z.; Atkatsh, K.; Tavarez, U.; Erdos, M.R.; Gruenbaum, Y.; Collins, F.S. Biotinylation by antibody recognition—A method for proximity labeling. Nat. Methods 2018, 15, 127–133. [Google Scholar] [CrossRef]
- Suzuki, H.; Abe, R.; Shimada, M.; Hirose, T.; Hirose, H.; Noguchi, K.; Ike, Y.; Yasui, N.; Furugori, K.; Yamaguchi, Y.; et al. The 3′ Pol II pausing at replication-dependent histone genes is regulated by Mediator through Cajal bodies’ association with histone locus bodies. Nat. Commun. 2022, 13, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Foglia, B.; Novo, E.; Protopapa, F.; Maggiora, M.; Bocca, C.; Cannito, S.; Parola, M. Hypoxia, hypoxia-inducible factors and liver fibrosis. Cells 2021, 10, 1764. [Google Scholar] [CrossRef] [PubMed]
- Cramer, T.; Vaupel, P. Severe hypoxia is a typical characteristic of human hepatocellular carcinoma: Scientific fact or fallacy? J. Hepatol. 2022, 76, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.J.; Harris, J.M.; Marchi, E.; D’Arienzo, V.; Michler, T.; Wing, P.A.C.; Magri, A.; Ortega-Prieto, A.M.; van de Klundert, M.; Wettengel, J.; et al. Hypoxic gene expression in chronic Hepatitis B virus infected patients is not observed in state-of-the-art in vitro and mouse infection models. Sci. Rep. 2020, 10, 14101. [Google Scholar] [CrossRef]
- Riedl, T.; Faure-dupuy, S.; Rolland, M.; Schuehle, S.; Hizir, Z.; Calderazzo, S.; Zhuang, X.; Wettengel, J.; Lopez, M.; Barnault, R.; et al. Europe PMC Funders Group HIF1 α-mediated RelB / APOBEC3B downregulation allows Hepatitis B Virus persistence. Hepatology 2021, 74, 1766–1781. [Google Scholar] [CrossRef]
- Wing, P.A.C.; Liu, P.J.; Harris, J.M.; Magri, A.; Michler, T.; Zhuang, X.; Borrmann, H.; Minisini, R.; Frampton, N.R.; Wettengel, J.M.; et al. Hypoxia inducible factors regulate Hepatitis B virus replication by activating the basal core promoter. J. Hepatol. 2021, 75, 64–73. [Google Scholar] [CrossRef]
- Sugiyama, M.; Tanaka, Y.; Kato, T.; Orito, E.; Ito, K.; Acharya, S.K.; Gish, R.G.; Kramvis, A.; Shimada, T.; Izumi, N.; et al. Influence of Hepatitis B virus genotypes on the intra- and extracellular expression of viral DNA and antigens. Hepatology 2006, 44, 915–924. [Google Scholar] [CrossRef]
- Matsunaga, S.; Kawakami, S.; Matsuo, I.; Okayama, A.; Tsukagoshi, H.; Kudoh, A.; Matsushima, Y.; Shimizu, H.; Okabe, N.; Hirano, H.; et al. Wheat germ cell-free system-based production of hemagglutinin-neuraminidase glycoprotein of human parainfluenza virus type 3 for generation and characterization of monoclonal antibody. Front. Microbiol. 2014, 5, 208. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, Y.; Matsuyama, S.; Fukushi, S.; Matsunaga, S.; Matsushima, Y.; Kuroyama, H.; Kimura, H.; Takeda, M.; Chimuro, T.; Ryo, A. Development of monoclonal antibody and diagnostic test for Middle East respiratory syndrome coronavirus using cell-free synthesized nucleocapsid antigen. Front. Microbiol. 2016, 7, 509. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, M.; Cai, D.; Sugiyama, M.; Suzuki, R.; Aizaki, H.; Ryo, A.; Ohtani, N.; Tanaka, Y.; Mizokami, M.; Wakita, T.; et al. Functional association of cellular microtubules with viral capsid assembly supports efficient Hepatitis B virus replication. Sci. Rep. 2017, 7, 10620. [Google Scholar] [CrossRef]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and identification of Hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, N.; Watashi, K.; Noguchi, T.; Wakita, T. Formation of covalently closed circular DNA in Hep38.7-Tet cells, a tetracycline inducible Hepatitis B virus expression cell line. Biochem. Biophys. Res. Commun. 2014, 452, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Hayer, J.; Jadeau, F.; Deléage, G.; Kay, A.; Zoulim, F.; Combet, C. HBVdb: A knowledge database for Hepatitis B Virus. Nucleic Acids Res. 2013, 41, 566–570. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, O.O. Candidate vaccine sequences to represent intra- and inter-clade HIV-1 variation. PLoS ONE 2009, 4, e7388. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Yamai, T.; Hikita, H.; Fukuoka, M.; Fukutomi, K.; Murai, K.; Nakabori, T.; Yamada, R.; Miyakawa, K.; Watashi, K.; Ryo, A.; et al. SIRT1 enhances hepatitis virus B transcription independent of hepatic autophagy. Biochem. Biophys. Res. Commun. 2020, 527, 64–70. [Google Scholar] [CrossRef]
- Sunbul, M. Hepatitis B virus genotypes: Global distribution and clinical importance. World J. Gastroenterol. 2014, 20, 5427–5434. [Google Scholar] [CrossRef]
- Lin, C.L.; Kao, J.H. Hepatitis B virus genotypes and variants. Cold Spring Harb. Perspect. Med. 2015, 5, 1–19. [Google Scholar] [CrossRef]
- Van Damme, E.; Vanhove, J.; Severyn, B.; Verschueren, L.; Pauwels, F. The Hepatitis B Virus Interactome: A Comprehensive Overview. Front. Microbiol. 2021, 12, 2581. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.O.; Sui, J.; Young, A.B.; Whittal, R.M. Interferences and contaminants encountered in modern mass spectrometry. Anal. Chim. Acta 2008, 627, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, C.; Guillou-Guillemette, H.L.; Ducancelle, A. A pleiotropic role of the Hepatitis b virus core protein in hepatocarcinogenesis. Int. J. Mol. Sci. 2021, 22, 13651. [Google Scholar] [CrossRef] [PubMed]
- Diab, A.; Foca, A.; Zoulim, F.; Durantel, D.; Andrisani, O. The diverse functions of the hepatitis B core/capsid protein (HBc) in the viral life cycle: Implications for the development of HBc-targeting antivirals. Antivir. Res. 2018, 149, 211–220. [Google Scholar] [CrossRef]
- Liu, W.; Guo, T.F.; Jing, Z.T.; Yang, Z.; Liu, L.; Yang, Y.P.; Lin, X.; Tong, Q.Y. Hepatitis B virus core protein promotes hepatocarcinogenesis by enhancing Src expression and activating the Src/PI3K/Akt pathway. FASEB J. 2018, 32, 3033–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabrolles, H.; Auclair, H.; Vegna, S.; Lahlali, T.; Pons, C.; Michelet, M.; Couté, Y.; Belmudes, L.; Chadeuf, G.; Kim, Y.; et al. Hepatitis B virus Core protein nuclear interactome identifies SRSF10 as a host RNA-binding protein restricting HBV RNA production. PLoS Pathog. 2020, 16, e1008593. [Google Scholar] [CrossRef] [PubMed]
- Genera, M.; Quioc-Salomon, B.; Nourisson, A.; Colcombet-Cazenave, B.; Haouz, A.; Mechaly, A.; Matondo, M.; Duchateau, M.; König, A.; Windisch, M.P.; et al. Molecular basis of the interaction of the human tyrosine phosphatase PTPN3 with the Hepatitis B virus core protein. Sci. Rep. 2021, 11, 944. [Google Scholar] [CrossRef]
- de Rocquigny, H.; Rat, V.; Pastor, F.; Darlix, J.L.; Hourioux, C.; Roingeard, P. Phosphorylation of the arginine-rich C-terminal domains of the Hepatitis B virus (HBV) core protein as a fine regulator of the interaction between HBc and nucleic acid. Viruses 2020, 12, 738. [Google Scholar] [CrossRef]
- Li, H.C.; Huang, E.Y.; Su, P.Y.; Wu, S.Y.; Yang, C.C.; Lin, Y.S.; Chang, W.C.; Shih, C. Nuclear export and import of human Hepatitis B virus capsid protein and particles. PLoS Pathog. 2010, 6, e1001162. [Google Scholar] [CrossRef] [Green Version]
- Guidotti, L.G.; Matzke, B.; Schaller, H.; Chisari, F. V High-level Hepatitis B virus replication in transgenic mice. J. Virol. 1995, 69, 6158–6169. [Google Scholar] [CrossRef]
- Ling, X.B.; Wei, H.W.; Wang, J.; Kong, Y.Q.; Wu, Y.Y.; Guo, J.L.; Li, T.F.; Li, J.K. Mammalian metallothionein-2A and oxidative stress. Int. J. Mol. Sci. 2016, 17, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Shi, F.; Huang, W.; Kang, Y.J. Metallothionein gene transfection reverses the phenotype of activated human hepatic stellate cells. J. Pharmacol. Exp. Ther. 2013, 346, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Zeng, J.; Geng, P.; Fang, C.; Wang, Y.; Sun, M.; Wang, C.; Wang, J.; Yin, P.; Hu, C.; et al. Global metabolic profiling identifies a pivotal role of proline and hydroxyproline metabolism in supporting hypoxic response in hepatocellular carcinoma. Clin. Cancer Res. 2018, 24, 474–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.Y.; Liu, J.Y.; Zhu, T.; Liu, C.; Gao, Y.; Dai, W.T.; Zhuo, W.; Mao, X.Y.; He, B.M.; Liu, Z.Q. Effects of Glycolysis-Related Genes on Prognosis and the Tumor Microenvironment of Hepatocellular Carcinoma. Front. Pharmacol. 2022, 13, 895608. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakai, Y.; Miyakawa, K.; Yamaoka, Y.; Hatayama, Y.; Nishi, M.; Suzuki, H.; Kimura, H.; Takahashi, H.; Kimura, Y.; Ryo, A. Generation and Utilization of a Monoclonal Antibody against Hepatitis B Virus Core Protein for a Comprehensive Interactome Analysis. Microorganisms 2022, 10, 2381. https://doi.org/10.3390/microorganisms10122381
Nakai Y, Miyakawa K, Yamaoka Y, Hatayama Y, Nishi M, Suzuki H, Kimura H, Takahashi H, Kimura Y, Ryo A. Generation and Utilization of a Monoclonal Antibody against Hepatitis B Virus Core Protein for a Comprehensive Interactome Analysis. Microorganisms. 2022; 10(12):2381. https://doi.org/10.3390/microorganisms10122381
Chicago/Turabian StyleNakai, Yusuke, Kei Miyakawa, Yutaro Yamaoka, Yasuyoshi Hatayama, Mayuko Nishi, Hidefumi Suzuki, Hirokazu Kimura, Hidehisa Takahashi, Yayoi Kimura, and Akihide Ryo. 2022. "Generation and Utilization of a Monoclonal Antibody against Hepatitis B Virus Core Protein for a Comprehensive Interactome Analysis" Microorganisms 10, no. 12: 2381. https://doi.org/10.3390/microorganisms10122381
APA StyleNakai, Y., Miyakawa, K., Yamaoka, Y., Hatayama, Y., Nishi, M., Suzuki, H., Kimura, H., Takahashi, H., Kimura, Y., & Ryo, A. (2022). Generation and Utilization of a Monoclonal Antibody against Hepatitis B Virus Core Protein for a Comprehensive Interactome Analysis. Microorganisms, 10(12), 2381. https://doi.org/10.3390/microorganisms10122381