
Condensate Formation by Metabolic Enzymes in Saccharomyces cerevisiae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Filaments Formed by a Single Metabolic Enzyme in S. cerevisiae
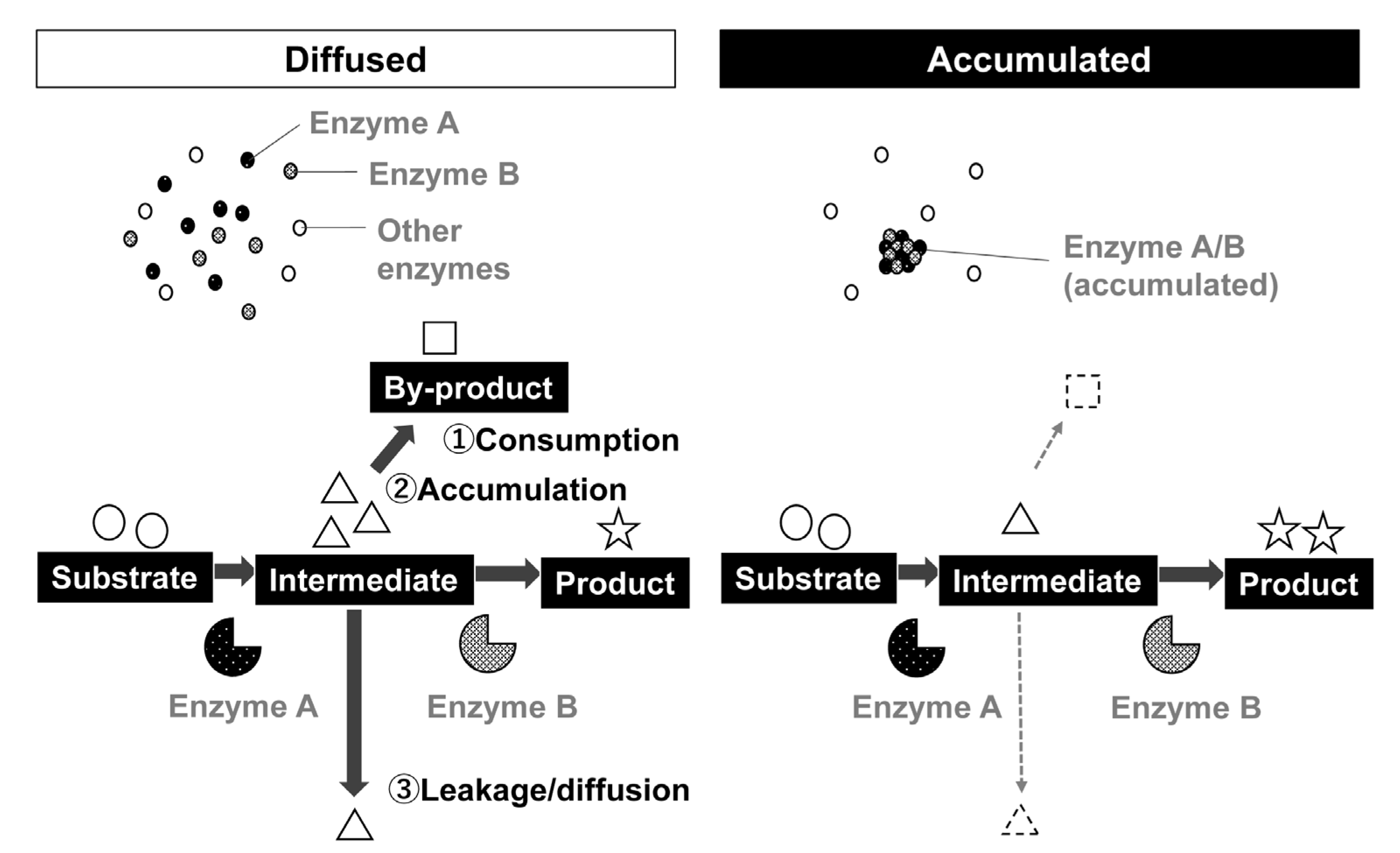
3. Predicted Effect of Condensate Formation by Metabolic Enzymes

4. Condensate Formation by a Group of Enzymes in the Purine Synthesis Pathway
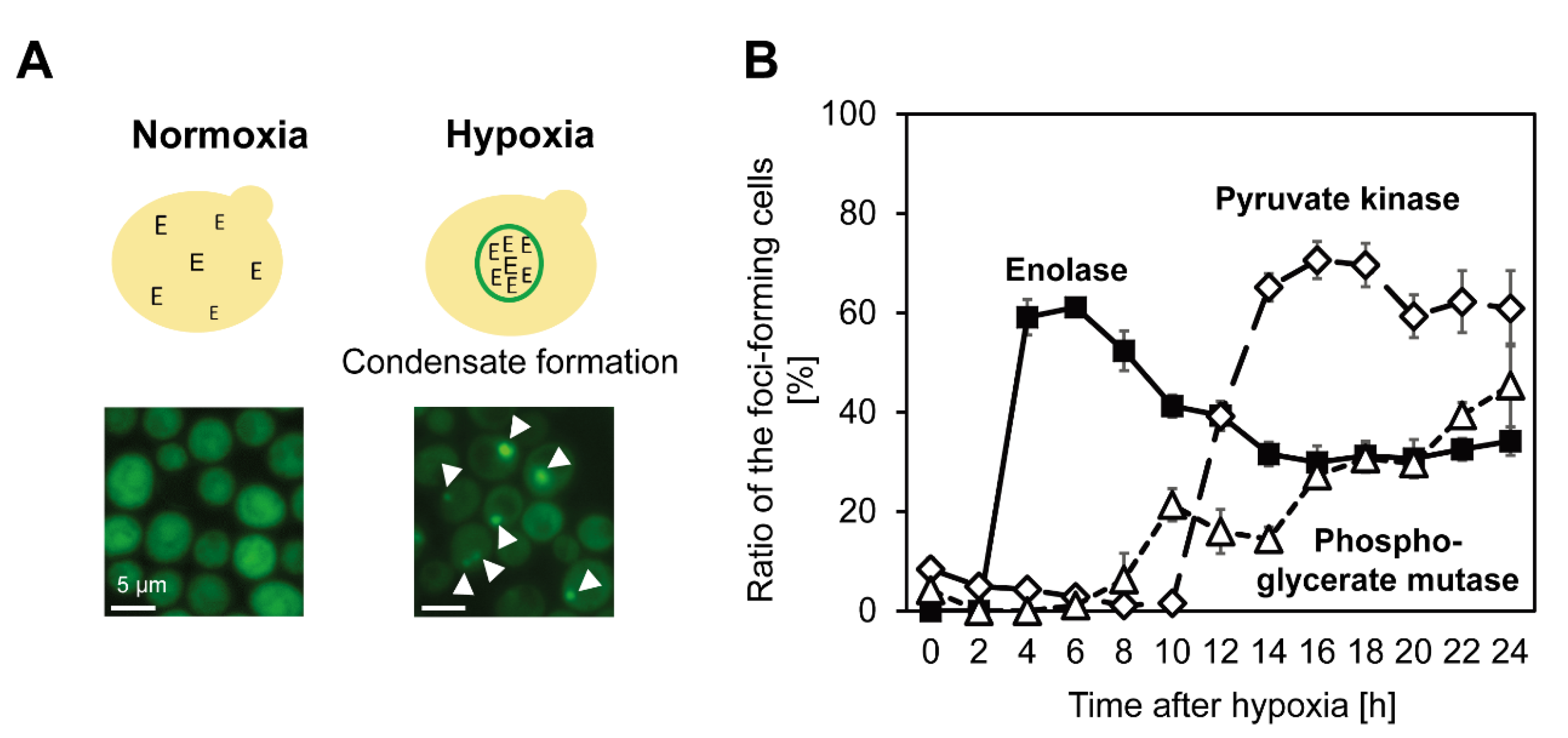
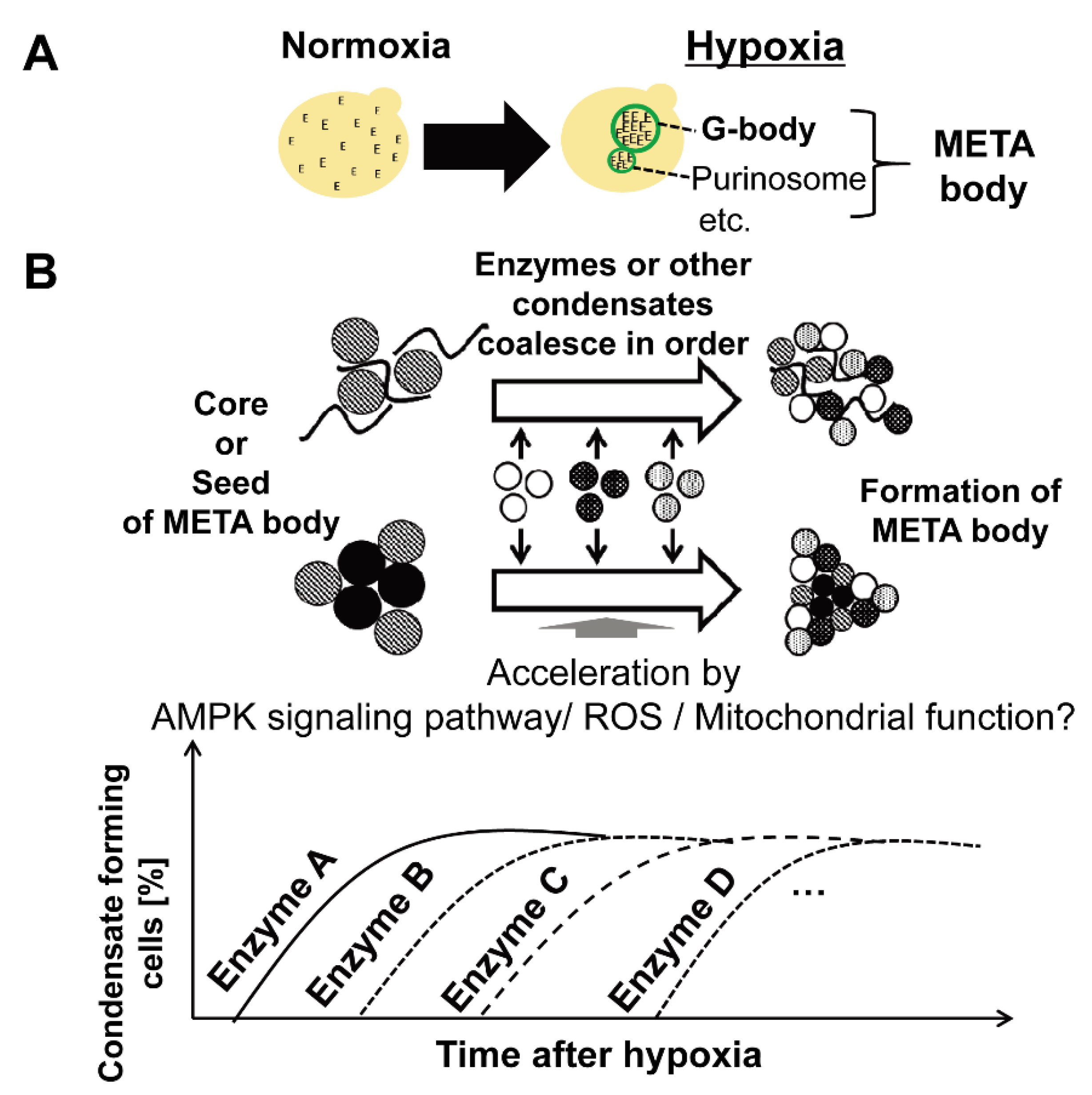
5. Coalesced Metabolic Enzymes under Hypoxia
6. Effect on Cellular Metabolism by Spatial Reorganization of Metabolic Enzymes under Hypoxia
7. Regulatory Machinery of Condensate Formation under Hypoxia
8. Summary and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y. The green fluorescent protein. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, C.J. Moonlighting proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef]
- Curtis, N.J.; Jeffery, C.J. The expanding world of metabolic enzymes moonlighting as rna binding proteins. Biochem. Soc. Trans. 2021, 49, 1099–1108. [Google Scholar] [CrossRef]
- Noree, C.; Begovich, K.; Samilo, D.; Broyer, R.; Monfort, E.; Wilhelm, J.E. A quantitative screen for metabolic enzyme structures reveals patterns of assembly across the yeast metabolic network. Mol. Biol. Cell 2019, 30, 2721–2736. [Google Scholar] [CrossRef]
- Noree, C.; Sato, B.K.; Broyer, R.M.; Wilhelm, J.E. Identification of novel filament-forming proteins in Saccharomyces cerevisiae and Drosophila melanogaster. J. Cell Biol. 2010, 190, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.J.; Kassim, H.; Huang, Y.; Li, H.; Zhang, J.; Li, G.; Wang, P.Y.; Yan, J.; Ye, F.; Liu, J.L. Filamentation of metabolic enzymes in Saccharomyces cerevisiae. J. Genet. Genom. 2016, 43, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Suresh, H.G.; da Silveira Dos Santos, A.X.; Kukulski, W.; Tyedmers, J.; Riezman, H.; Bukau, B.; Mogk, A. Prolonged starvation drives reversible sequestration of lipid biosynthetic enzymes and organelle reorganization in Saccharomyces cerevisiae. Mol. Biol. Cell 2015, 26, 1601–1615. [Google Scholar] [CrossRef]
- Narayanaswamy, R.; Levy, M.; Tsechansky, M.; Stovall, G.M.; O’Connell, J.D.; Mirrielees, J.; Ellington, A.D.; Marcotte, E.M. Widespread reorganization of metabolic enzymes into reversible assemblies upon nutrient starvation. Proc. Natl. Acad. Sci. USA 2009, 106, 10147–10152. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Fuller, G.G.; Han, T.; Yao, Y.; Alessi, A.F.; Freeberg, M.A.; Roach, N.P.; Moresco, J.J.; Karnovsky, A.; Baba, M.; et al. Glycolytic enzymes coalesce in g bodies under hypoxic stress. Cell Rep. 2017, 20, 895–908. [Google Scholar] [CrossRef] [Green Version]
- Miura, N.; Shinohara, M.; Tatsukami, Y.; Sato, Y.; Morisaka, H.; Kuroda, K.; Ueda, M. Spatial reorganization of Saccharomyces cerevisiae enolase to alter carbon metabolism under hypoxia. Eukaryot. Cell 2013, 12, 1106–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, Y.; Hirayama, R.; Miura, N.; Utsumi, R.; Kuroda, K.; Ueda, M.; Kataoka, M. Small-scale hypoxic cultures for monitoring the spatial reorganization of glycolytic enzymes in Saccharomyces cerevisiae. Cell Biol. Int. 2021, 45, 1776–1783. [Google Scholar] [CrossRef]
- Park, C.K.; Horton, N.C. Structures, functions, and mechanisms of filament forming enzymes: A renaissance of enzyme filamentation. Biophys. Rev. 2019, 11, 927–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinschmidt, A.K.; Moss, J.; Lane, D.M. Acetyl coenzyme a carboxylase: Filamentous nature of the animal enzymes. Science 1969, 166, 1276–1278. [Google Scholar] [CrossRef] [PubMed]
- Hunkeler, M.; Hagmann, A.; Stuttfeld, E.; Chami, M.; Guri, Y.; Stahlberg, H.; Maier, T. Structural basis for regulation of human acetyl-coa carboxylase. Nature 2018, 558, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Meredith, M.J.; Lane, M.D. Acetyl-coa carboxylase. Evidence for polymeric filament to protomer transition in the intact avian liver cell. J. Biol. Chem. 1978, 253, 3381–3383. [Google Scholar] [CrossRef]
- Costello, L.C.; Franklin, R.B. A review of the important central role of altered citrate metabolism during the process of stem cell differentiation. J. Regen Med. Tissue Eng. 2013, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Bennett, B.D.; Kimball, E.H.; Gao, M.; Osterhout, R.; Van Dien, S.J.; Rabinowitz, J.D. Absolute metabolite concentrations and implied enzyme active site occupancy in Escherichia coli. Nat. Chem. Biol. 2009, 5, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.; Yang, D.; Hou, Y.; Liu, S.; Zhao, M.; Qin, Y.; Chen, R.; Teng, Y.; Liu, M. Intracellular citrate accumulation by oxidized atm-mediated metabolism reprogramming via pfkp and cs enhances hypoxic breast cancer cell invasion and metastasis. Cell Death Dis. 2019, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- Ingerson-Mahar, M.; Briegel, A.; Werner, J.N.; Jensen, G.J.; Gitai, Z. The metabolic enzyme CTP synthase forms cytoskeletal filaments. Nat. Cell Biol. 2010, 12, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L. Intracellular compartmentation of CTP synthase in Drosophila. J. Genet. Genom. 2010, 37, 281–296. [Google Scholar] [CrossRef]
- Carcamo, W.C.; Satoh, M.; Kasahara, H.; Terada, N.; Hamazaki, T.; Chan, J.Y.; Yao, B.; Tamayo, S.; Covini, G.; von Muhlen, C.A.; et al. Induction of cytoplasmic rods and rings structures by inhibition of the CTP and GTP synthetic pathway in mammalian cells. PLoS ONE 2011, 6, e29690. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.M.; Horowitz, A.; Lynch, E.M.; Farrell, D.P.; Quispe, J.; DiMaio, F.; Kollman, J.M. Cryo-em structures of CTP synthase filaments reveal mechanism of ph-sensitive assembly during budding yeast starvation. Elife 2021, 10I, e73368. [Google Scholar] [CrossRef]
- Aughey, G.N.; Grice, S.J.; Shen, Q.J.; Xu, Y.; Chang, C.C.; Azzam, G.; Wang, P.Y.; Freeman-Mills, L.; Pai, L.M.; Sung, L.Y.; et al. Nucleotide synthesis is regulated by cytoophidium formation during neurodevelopment and adaptive metabolism. Biol. Open 2014, 3, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Barry, R.M.; Bitbol, A.F.; Lorestani, A.; Charles, E.J.; Habrian, C.H.; Hansen, J.M.; Li, H.J.; Baldwin, E.P.; Wingreen, N.S.; Kollman, J.M.; et al. Large-scale filament formation inhibits the activity of CTP synthetase. Elife 2014, 3, e03638. [Google Scholar] [CrossRef]
- Noree, C.; Monfort, E.; Shiau, A.K.; Wilhelm, J.E. Common regulatory control of CTP synthase enzyme activity and filament formation. Mol. Biol. Cell 2014, 25, 2282–2290. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, J.L. Forming cytoophidia prolongs the half-life of CTP synthase. Cell Discov. 2019, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Aughey, G.N.; Liu, J.L. Metabolic regulation via enzyme filamentation. Crit. Rev. Biochem. Mol. Biol. 2015, 51, 282–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, E.M.; Hicks, D.R.; Shepherd, M.; Endrizzi, J.A.; Maker, A.; Hansen, J.M.; Barry, R.M.; Gitai, Z.; Baldwin, E.P.; Kollman, J.M. Human CTP synthase filament structure reveals the active enzyme conformation. Nat. Struct. Mol. Biol. 2017, 24, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Simonet, J.C.; Foster, M.J.; Lynch, E.M.; Kollman, J.M.; Nicholas, E.; O’Reilly, A.M.; Peterson, J.R. CTP synthase polymerization in germline cells of the developing Drosophila egg supports egg production. Biol. Open 2020, 9, bio050328. [Google Scholar] [CrossRef]
- Stoddard, P.R.; Lynch, E.M.; Farrell, D.P.; Dosey, A.M.; Di Maio, F.; Williams, T.A.; Kollman, J.M.; Murray, A.W.; Garner, E.C. Polymerization in the actin atpase clan regulates hexokinase activity in yeast. Science 2020, 367, 1039–1042. [Google Scholar] [CrossRef] [PubMed]
- Cereghetti, G.; Saad, S.; Dechant, R.; Peter, M. Reversible, functional amyloids: Towards an understanding of their regulation in yeast and humans. Cell Cycle 2018, 17, 1545–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cereghetti, G.; Wilson-Zbinden, C.; Kissling, V.M.; Diether, M.; Arm, A.; Yoo, H.; Piazza, I.; Saad, S.; Picotti, P.; Drummond, D.A.; et al. Reversible amyloids of pyruvate kinase couple cell metabolism and stress granule disassembly. Nat. Cell Biol. 2021, 23, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Saad, S.; Cereghetti, G.; Feng, Y.; Picotti, P.; Peter, M.; Dechant, R. Reversible protein aggregation is a protective mechanism to ensure cell cycle restart after stress. Nat. Cell Biol. 2017, 19, 1202–1213. [Google Scholar] [CrossRef]
- Grignaschi, E.; Cereghetti, G.; Grigolato, F.; Kopp, M.R.G.; Caimi, S.; Faltova, L.; Saad, S.; Peter, M.; Arosio, P. A hydrophobic low-complexity region regulates aggregation of the yeast pyruvate kinase Cdc19 into amyloid-like aggregates In vitro. J. Biol. Chem. 2018, 293, 11424–11432. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.A.; Dosey, A.M.; Wittmann, T.; Kollman, J.M.; Barber, D.L. The glycolytic enzyme phosphofructokinase-1 assembles into filaments. J. Cell Biol. 2017, 216, 2305–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Nelson, J.C.; Bend, E.G.; Rodriguez-Laureano, L.; Tueros, F.G.; Cartagenova, L.; Underwood, K.; Jorgensen, E.M.; Colon-Ramos, D.A. Glycolytic enzymes localize to synapses under energy stress to support synaptic function. Neuron 2016, 90, 278–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, E.; Shorter, J. The molecular language of membraneless organelles. J. Biol. Chem. 2019, 294, 7115–7127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivey, H.O.; Merz, J.M. Metabolic compartmentation. Bioessays 1989, 10, 127–130. [Google Scholar] [CrossRef]
- Schmid-Dannert, C.; Lopez-Gallego, F. Advances and opportunities for the design of self-sufficient and spatially organized cell-free biocatalytic systems. Curr. Opin. Chem. Biol. 2019, 49, 97–104. [Google Scholar] [CrossRef]
- Bae, J.; Kuroda, K.; Ueda, M. Proximity effect among cellulose-degrading enzymes displayed on the Saccharomyces cerevisiae cell surface. Appl. Environ. Microbiol. 2015, 81, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Ito, J.; Ueda, M.; Fukuda, H.; Kondo, A. Synergistic saccharification, and direct fermentation to ethanol, of amorphous cellulose by use of an engineered yeast strain codisplaying three types of cellulolytic enzyme. Appl. Environ. Microbiol. 2004, 70, 1207–1212. [Google Scholar] [CrossRef] [Green Version]
- Peeples, W.B.; Rosen, M.K. Phase separation can increase enzyme activity by concentration and molecular organization. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Ura, T.; Kagawa, A.; Yagi, H.; Tochio, N.; Kigawa, T.; Mikawa, T.; Shiraki, K. Hyperactivation of l-lactate oxidase by liquid–liquid phase separation. bioRxiv 2020. [Google Scholar] [CrossRef]
- An, S.; Kumar, R.; Sheets, E.D.; Benkovic, S.J. Reversible compartmentalization of de novo purine biosynthetic complexes in living cells. Science 2008, 320, 103–106. [Google Scholar] [CrossRef]
- An, S.; Deng, Y.; Tomsho, J.W.; Kyoung, M.; Benkovic, S.J. Microtubule-assisted mechanism for functional metabolic macromolecular complex formation. Proc. Natl. Acad. Sci. USA 2010, 107, 12872–12876. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.Y.; Zhao, H.; Pugh, R.J.; Pedley, A.M.; French, J.; Jones, S.A.; Zhuang, X.; Jinnah, H.; Huang, T.J.; Benkovic, S.J. Purinosome formation as a function of the cell cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 1368–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedley, A.M.; Benkovic, S.J. A new view into the regulation of purine metabolism: The purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohnhorst, C.L.; Kyoung, M.; Jeon, M.; Schmitt, D.L.; Kennedy, E.L.; Ramirez, J.; Bracey, S.M.; Luu, B.T.; Russell, S.J.; An, S. Identification of a multienzyme complex for glucose metabolism in living cells. J. Biol. Chem. 2017, 292, 9191–9203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doigneaux, C.; Pedley, A.M.; Mistry, I.N.; Papayova, M.; Benkovic, S.J.; Tavassoli, A. Hypoxia drives the assembly of the multienzyme purinosome complex. J. Biol. Chem 2020, 295, 9551–9566. [Google Scholar] [CrossRef]
- Chinopoulos, C. Which way does the citric acid cycle turn during hypoxia? The critical role of alpha-ketoglutarate dehydrogenase complex. J. Neurosci. Res. 2013, 91, 1030–1043. [Google Scholar] [CrossRef] [Green Version]
- Wiebe, M.G.; Rintala, E.; Tamminen, A.; Simolin, H.; Salusjarvi, L.; Toivari, M.; Kokkonen, J.T.; Kiuru, J.; Ketola, R.A.; Jouhten, P.; et al. Central carbon metabolism of Saccharomyces cerevisiae in anaerobic, oxygen-limited and fully aerobic steady-state conditions and following a shift to anaerobic conditions. FEMS Yeast Res. 2008, 8, 140–154. [Google Scholar] [CrossRef]
- Fuller, G.G.; Han, T.; Freeberg, M.A.; Moresco, J.J.; Ghanbari Niaki, A.; Roach, N.P.; Yates, J.R., 3rd; Myong, S.; Kim, J.K. RNA promotes phase separation of glycolysis enzymes into yeast g bodies in hypoxia. Elife 2020, 9, e48480. [Google Scholar] [CrossRef] [PubMed]
- Morales-Polanco, F.; Bates, C.; Lui, J.; Casson, J.; Solari, C.A.; Pizzinga, M.; Forte, G.; Griffin, C.; Garner, K.E.L.; Burt, H.E.; et al. Core fermentation (CoFe) granules focus coordinated glycolytic mRNA localization and translation to fuel glucose fermentation. iScience 2021, 24, 102069. [Google Scholar] [CrossRef]
- Kiefhaber, T.; Rudolph, R.; Kohler, H.H.; Buchner, J. Protein aggregation In vitro and In vivo: A quantitative model of the kinetic competition between folding and aggregation. Biotechnology 1991, 9, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Folding proteins in fatal ways. Nature 2003, 426, 900–904. [Google Scholar] [CrossRef]
- Molliex, A.; Temirov, J.; Lee, J.; Coughlin, M.; Kanagaraj, A.P.; Kim, H.J.; Mittag, T.; Taylor, J.P. Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell 2015, 163, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, Y.; Alam, J.M.; Noshiro, D.; Mouri, K.; Ando, T.; Okada, Y.; May, A.I.; Knorr, R.L.; Suzuki, K.; Ohsumi, Y.; et al. Phase separation organizes the site of autophagosome formation. Nature 2020, 578, 301–305. [Google Scholar] [CrossRef]
- Noda, N.N.; Wang, Z.; Zhang, H. Liquid–liquid phase separation in autophagy. J. Cell Biol. 2020, 219, e202004062. [Google Scholar] [CrossRef]
- Yasuda, S.; Tsuchiya, H.; Kaiho, A.; Guo, Q.; Ikeuchi, K.; Endo, A.; Arai, N.; Ohtake, F.; Murata, S.; Inada, T.; et al. Stress- and ubiquitylation-dependent phase separation of the proteasome. Nature 2020, 578, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Menard, L.; Maughan, D.; Vigoreaux, J. The structural and functional coordination of glycolytic enzymes in muscle: Evidence of a metabolon? Biology 2014, 3, 623–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agbor, T.A.; Cheong, A.; Comerford, K.M.; Scholz, C.C.; Bruning, U.; Clarke, A.; Cummins, E.P.; Cagney, G.; Taylor, C.T. Small ubiquitin-related modifier (SUMO)-1 promotes glycolysis in hypoxia. J. Biol. Chem. 2011, 286, 4718–4726. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miura, N. Condensate Formation by Metabolic Enzymes in Saccharomyces cerevisiae. Microorganisms 2022, 10, 232. https://doi.org/10.3390/microorganisms10020232
Miura N. Condensate Formation by Metabolic Enzymes in Saccharomyces cerevisiae. Microorganisms. 2022; 10(2):232. https://doi.org/10.3390/microorganisms10020232
Chicago/Turabian StyleMiura, Natsuko. 2022. "Condensate Formation by Metabolic Enzymes in Saccharomyces cerevisiae" Microorganisms 10, no. 2: 232. https://doi.org/10.3390/microorganisms10020232
APA StyleMiura, N. (2022). Condensate Formation by Metabolic Enzymes in Saccharomyces cerevisiae. Microorganisms, 10(2), 232. https://doi.org/10.3390/microorganisms10020232