
Low-Temperature Biodegradation of Lignin-Derived Aromatic Model Monomers by the Cold-Adapted Yeast Rhodosporidiobolus colostri Isolated from Alpine Forest Soil


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Chemicals
2.3. Biodegradation of Lignin-Derived Aromatic Monomers
2.4. HPLC Analysis
3. Results
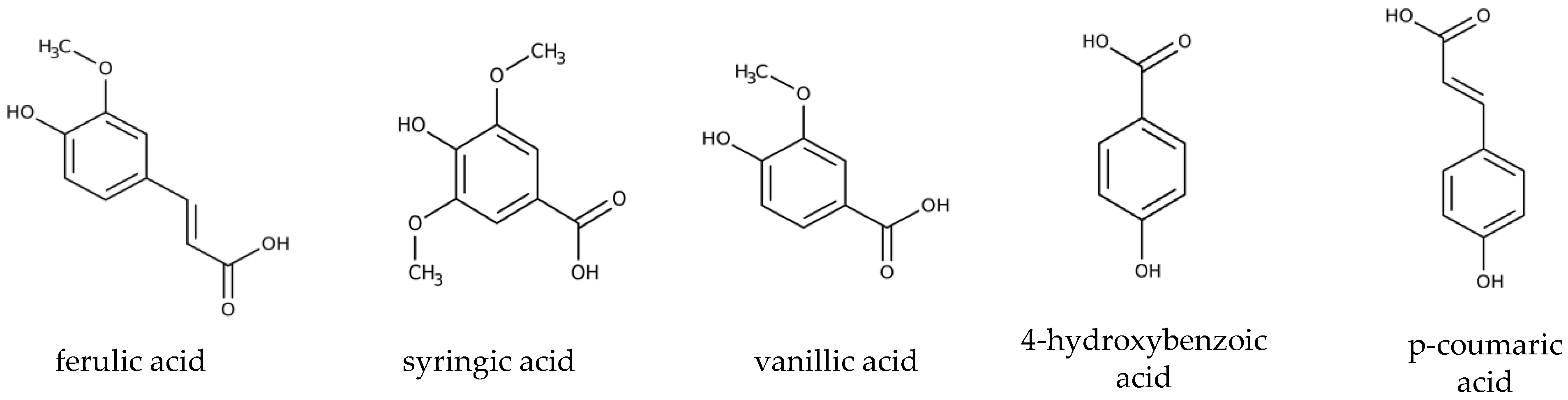
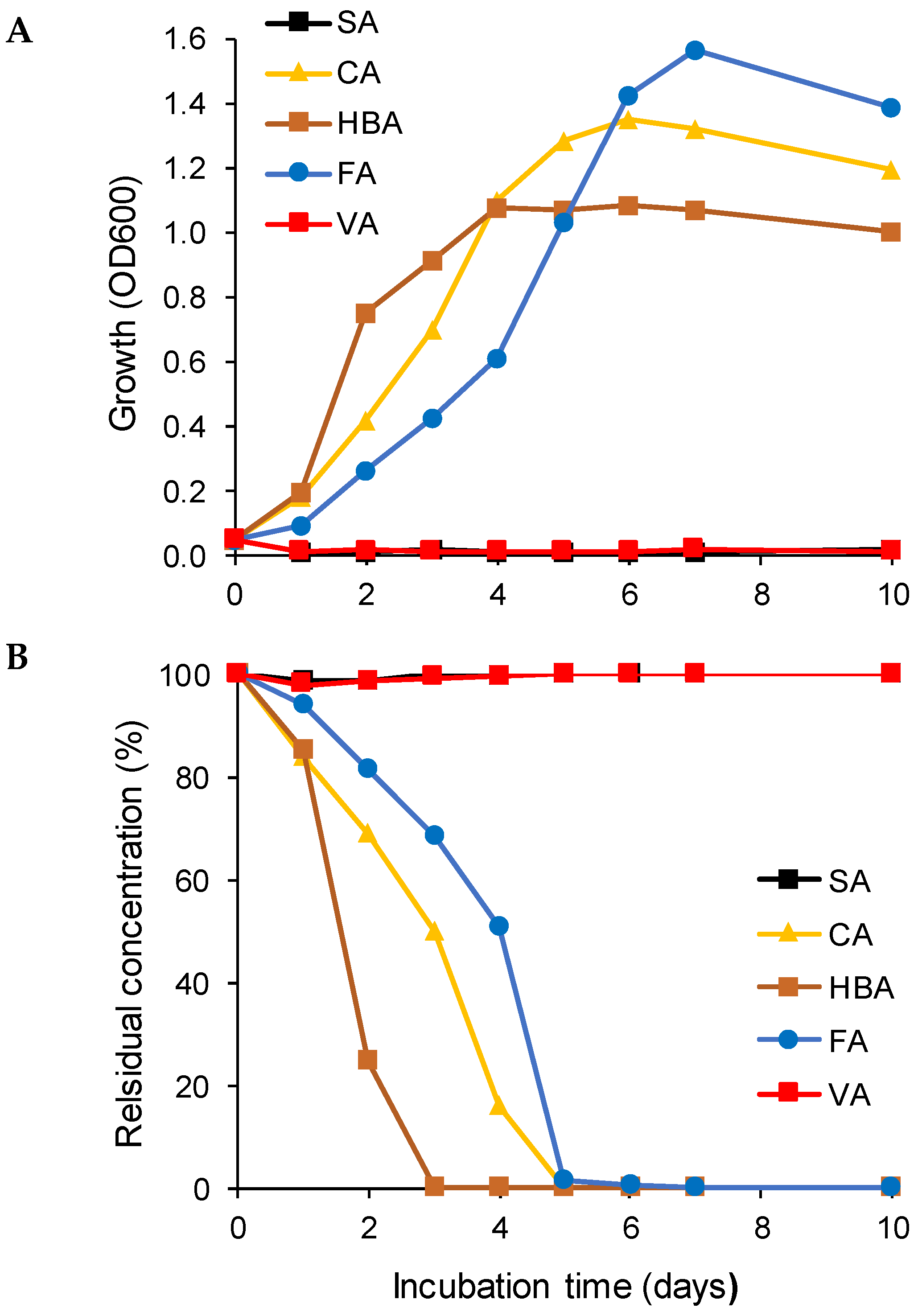
3.1. Biodegradation of Individual Lignin-Derived Aromatic Monomers
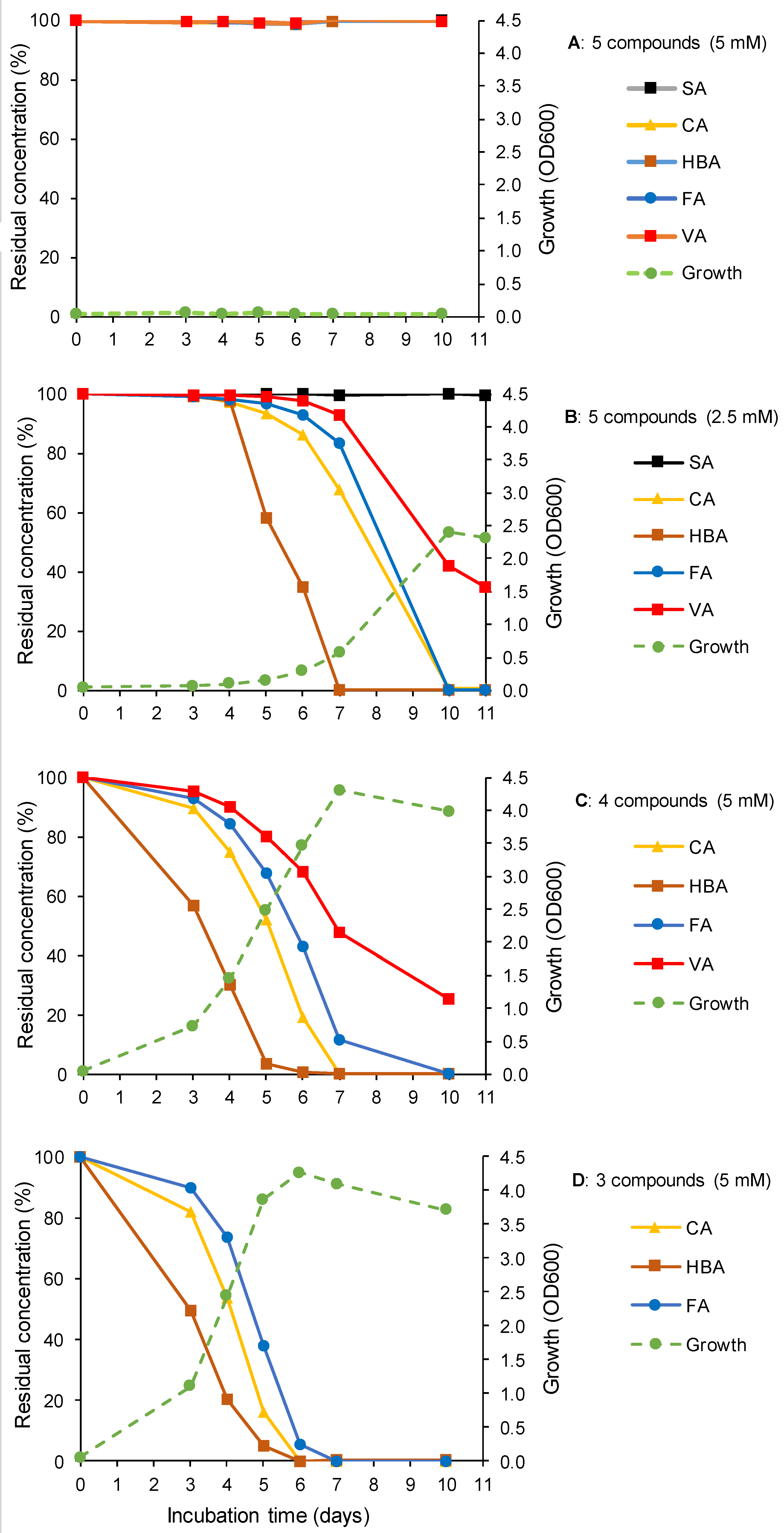
3.2. Biodegradation of LigninDerived Aromatic Monomers in Mixture
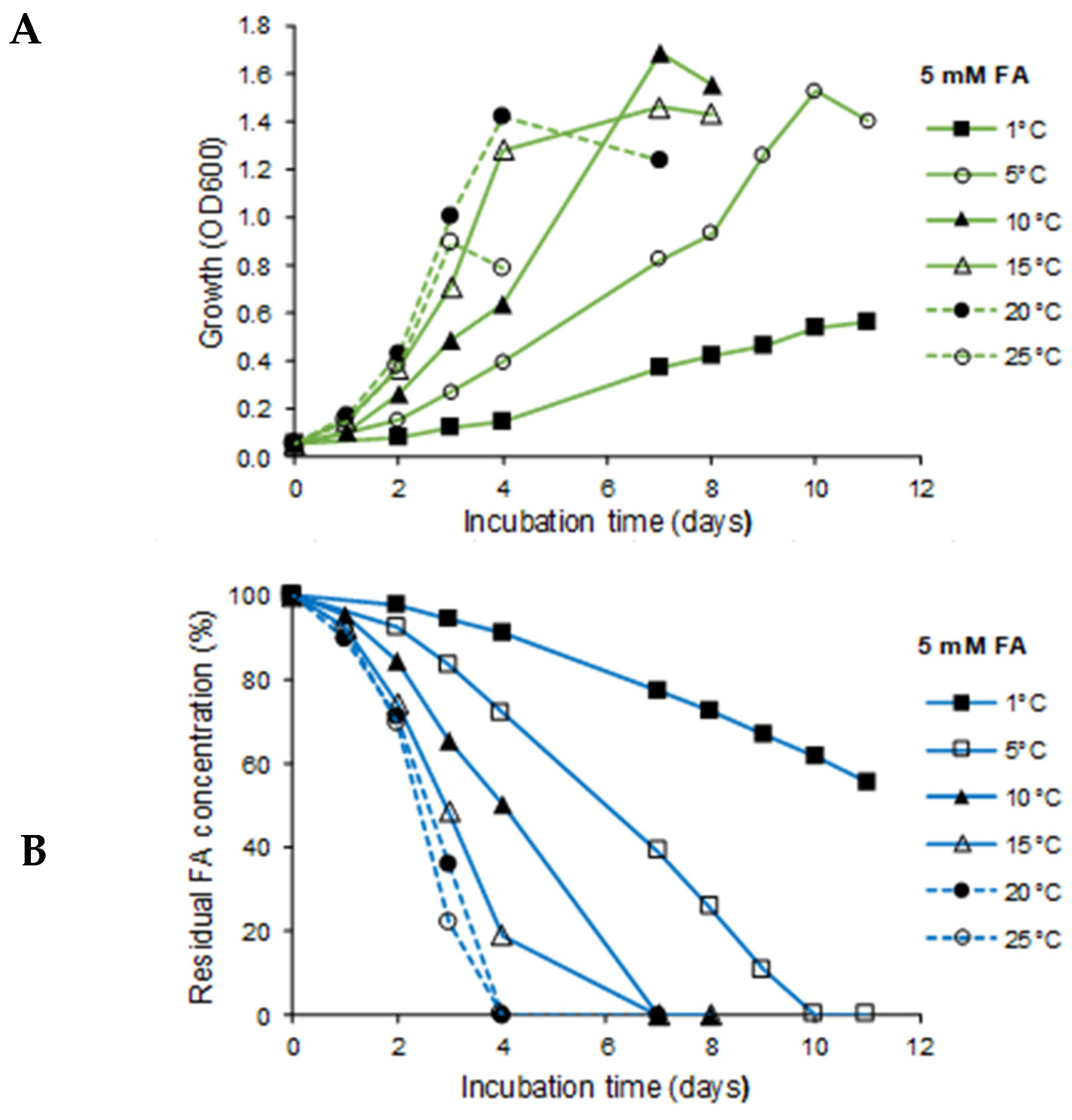
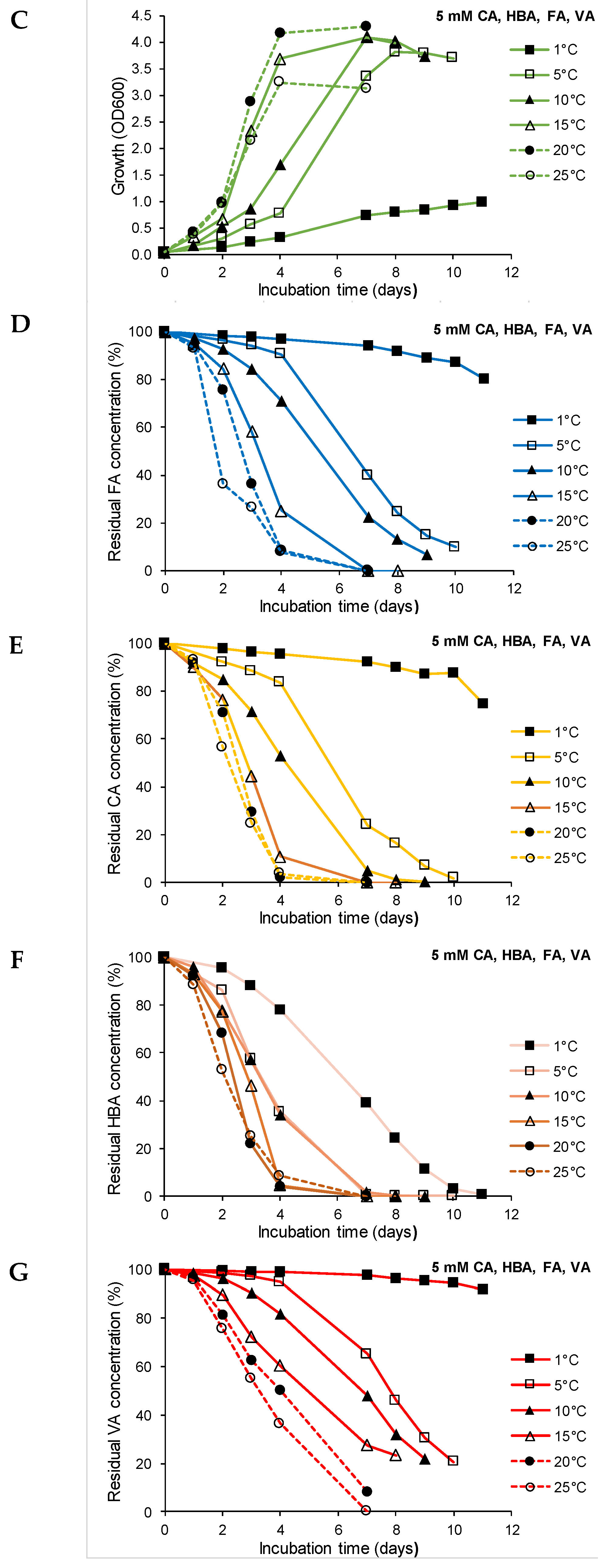
3.3. Effect of Temperature on Growth and Biodegradation
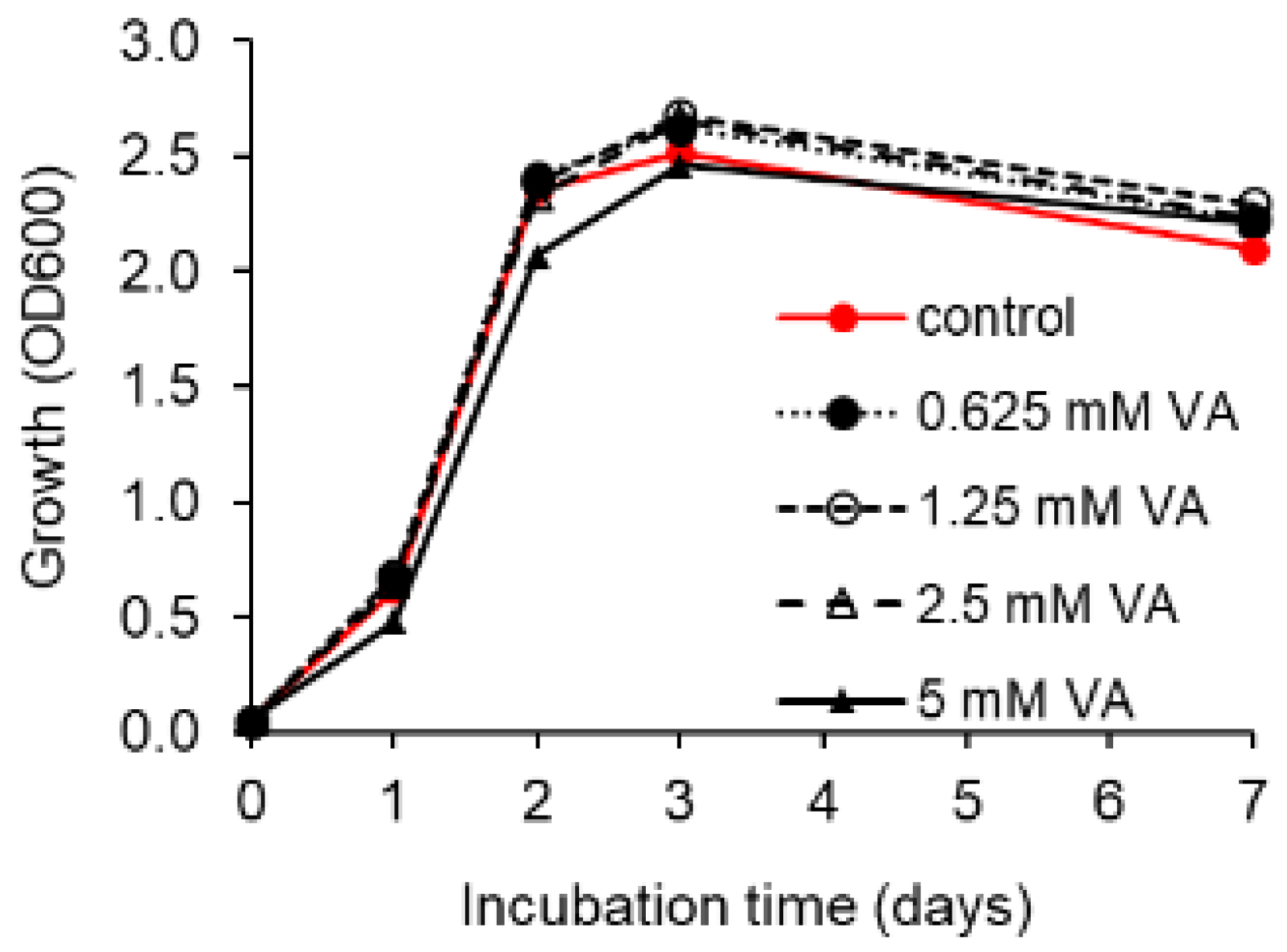
3.4. Toxicity of VA
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brink, D.P.; Ravi, K.; Liden, G.; Gorwa-Grauslund, M.F. Mapping the diversity of microbial lignin catabolism: Experiences from the eLignin database. Appl. Microbiol. Biotechnol. 2019, 103, 3979–4002. [Google Scholar] [CrossRef] [Green Version]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Singh, R. The emerging role for bacteria in lignin degradation and bio-product formation. Curr. Opin. Biotechnol. 2011, 22, 394–400. [Google Scholar] [CrossRef]
- Hainal, A.R.; Capraru, A.M.; Volf, I.; Popa, V.I. Lignin as a carbon source for the cultivation of some Rhodotorula species. Cellul. Chem. Technol. 2012, 46, 87–96. [Google Scholar]
- Ganewatta, M.S.; Lokupitiya, H.N.; Tang, C.B. Lignin biopolymers in the age of controlled polymerization. Polymers 2019, 11, 1176. [Google Scholar] [CrossRef] [Green Version]
- Lubbers, R.J.M.; Dilokpimol, A.; Visser, J.; Makela, M.R.; Hilden, K.S.; de Vries, R.P. A comparison between the homocyclic aromatic metabolic pathways from plant-derived compounds by bacteria and fungi. Biotechnol. Adv. 2019, 37, 107396. [Google Scholar] [CrossRef]
- Tian, J.H.; Pourcher, A.M.; Bouchez, T.; Gelhaye, E.; Peu, P. Occurrence of lignin degradation genotypes and phenotypes among prokaryotes. Appl. Microbiol. Biotechnol. 2014, 98, 9527–9544. [Google Scholar] [CrossRef]
- Palazzolo, M.A.; Kurina-Sanz, M. Microbial utilization of lignin: Available biotechnologies for its degradation and valorization. World J. Microbiol. Biotechnol. 2016, 32, 173. [Google Scholar] [CrossRef]
- Becker, J.; Wittmann, C. A field of dreams: Lignin valorization into chemicals, materials, fuels, and health-care products. Biotechnol. Adv. 2019, 37, 107360. [Google Scholar] [CrossRef]
- Sampaio, J.P. Utilization of low-molecular weight lignin-related aromatic compounds for the selective isolation of yeasts: Rhodotorula vanillica, a new basidiomycetous yeast species. Syst. Appl. Microbiol. 1995, 17, 613–619. [Google Scholar] [CrossRef]
- Ravi, K.; Garcia-Hidalgo, J.; Nobel, M.; Gorwa-Grauslund, M.F.; Liden, G. Biological conversion of aromatic monolignol compounds by a Pseudomonas isolate from sediments of the Baltic Sea. AMB Express 2018, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Margesin, R.; Volgger, G.; Wagner, A.O.; Zhang, D.C.; Poyntner, C. Biodegradation of lignin monomers and bioconversion of ferulic acid to vanillic acid by Paraburkholderia aromaticivorans AR20-38 isolated from Alpine forest soil. Appl. Microbiol. Biotechnol. 2021, 105, 2967–2977. [Google Scholar] [CrossRef]
- Botha, A. The importance and ecology of yeasts in soil. Soil Biol. Biochem. 2011, 43, 1–8. [Google Scholar] [CrossRef]
- Sampaio, J.P.; Vanuden, N. Rhodotorula ferulica sp. nov, a yeast that degrades ferulic acid and other phenolic compounds. Syst. Appl. Microbiol. 1991, 14, 146–149. [Google Scholar] [CrossRef]
- Huang, Z.X.; Dostal, L.; Rosazza, J.P.N. Mechanisms of ferulic acid conversions to vanillic acid and guaiacol by Rhodotorula rubra. J. Biol. Chem. 1993, 268, 23954–23958. [Google Scholar] [CrossRef]
- Middelhoven, W.J. Catabolism of benzene compounds by ascomycetous and basidiomycetous yeasts and yeast-like fungi—A literature review and an experimental approach. Antonie Leeuwenhoek 1993, 63, 125–144. [Google Scholar] [CrossRef]
- Rahouti, M.; Steiman, R.; Seigle-Murandi, F.; Christov, L.P. Growth of 1044 strains and species of fungi on 7 phenolic lignin model compounds. Chemosphere 1999, 38, 2549–2559. [Google Scholar] [CrossRef]
- Sampaio, J.P. Utilization of low molecular weight aromatic compounds by heterobasidiomycetous yeasts: Taxonomic implications. Can. J. Microbiol. 1999, 45, 491–512. [Google Scholar] [CrossRef]
- Alvarez-Rodriguez, M.L.; Belloch, C.; Villa, M.; Uruburu, F.; Larriba, G.; Coque, J.J.R. Degradation of vanillic acid and production of guaiacol by microorganisms isolated from cork samples. FEMS Microbiol. Lett. 2003, 220, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Jarboui, R.; Baati, H.; Fetoui, F.; Gargouri, A.; Gharsallah, N.; Ammar, E. Yeast performance in wastewater treatment: Case study of Rhodotorula mucilaginosa. Environ. Technol. 2012, 33, 951–960. [Google Scholar] [CrossRef]
- Max, B.; Tugores, F.; Cortes-Dieguez, S.; Dominguez, J.M. Bioprocess design for the microbial production of natural phenolic compounds by Debaryomyces hansenii. Appl. Biochem. Biotechnol. 2012, 168, 2268–2284. [Google Scholar] [CrossRef]
- Bettio, G.; Zardo, L.C.; Rosa, C.A.; Ayub, M.A.Z. Bioconversion of ferulic acid into aroma compounds by newly isolated yeast strains of the Latin American biodiversity. Biotechnol. Prog. 2021, 37, e3067. [Google Scholar] [CrossRef] [PubMed]
- Guiraud, P.; Steiman, R.; Seiglemurandi, F.; Benoitguyod, J.L. Metabolism of vanillic acid by micromycetes. World J. Microbiol. Biotechnol. 1992, 8, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.; Poyntner, C.; Margesin, R. Culturable bacteria from an Alpine coniferous forest site: Biodegradation potential of organic polymers and pollutants. Folia Microbiologica 2021, 66, 87–98. [Google Scholar] [CrossRef]
- Poyntner, C.; Kutzner, A.; Margesin, R. Biodegradation potential and putative catabolic genes of culturable bacteria from an Alpine deciduous forest site. Microorganisms 2021, 9, 1920. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Margesin, R. (Eds.) Cold-Adapted Yeasts. Biodiversity, Adaptation Strategies and Biotechnological Significance; Springer: Berlin/Heidelberg, Germany, 2014; p. 549. [Google Scholar]
- Margesin, R.; Collins, T. Microbial ecology of the cryosphere (glacial and permafrost habitats): Current knowledge. Appl. Microbiol. Biotechnol. 2019, 103, 2537–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, T.; Margesin, R. Psychrophilic lifestyles: Mechanisms of adaptation and biotechnological tools. Appl. Microbiol. Biotechnol. 2019, 103, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Franca, L.; Sannino, C.; Turchetti, B.; Buzzini, P.; Margesin, R. Seasonal and altitudinal changes of culturable bacterial and yeast diversity in Alpine forest soils. Extremophiles 2016, 20, 855–873. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.M.; Yurkov, A.M.; Goker, M.; Lumbsch, H.T.; Leavitt, S.D.; Groenewald, M.; Theelen, B.; Liu, X.Z.; Boekhout, T.; Bai, F.Y. Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud. Mycol. 2015, 81, 149–189. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.O.; Markt, R.; Puempel, T.; Illmer, P.; Insam, H.; Ebner, C. Sample preparation, preservation, and storage for volatile fatty acid quantification in biogas plants. Eng. Life Sci. 2017, 17, 132–139. [Google Scholar] [CrossRef]
- Turchetti, B.; Selbmann, L.; Gunde-Cimerman, N.; Buzzini, P.; Sampaio, J.P.; Zalar, P. Cystobasidium alpinum sp. nov. and Rhodosporidiobolus oreadorum sp. nov. from European cold environments and Arctic region. Life 2018, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Daskaya-Dikmen, C.; Karbancioglu-Guler, F.; Ozcelik, B. Cold active pectinase, amylase and protease production by yeast isolates obtained from environmental samples. Extremophiles 2018, 22, 599–606. [Google Scholar] [CrossRef]
- Glushakova, A.M.; Kachalkin, A.V. Endophytic yeasts in Malus domestica and Pyrus communis fruits under anthropogenic impact. Microbiology 2017, 86, 128–135. [Google Scholar] [CrossRef]
- Ramirez-Castrillon, M.; Usman, L.M.; Silva-Bedoya, L.M.; Osorio-Cadavid, E. Dominant yeasts associated to mango (Mangifera indica) and rose apple (Syzygium malaccense) fruit pulps investigated by culture-based methods. An. Acad. Bras. Cienc. 2019, 91, e20190052. [Google Scholar] [CrossRef]
- Lorenzini, M.; Zapparoli, G. Yeast-like fungi and yeasts in withered grape carposphere: Characterization of Aureobasidium pullulans population and species diversity. Int. J. Food Microbiol. 2019, 289, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Kaewkrajay, C.; Putchakarn, S.; Limtong, S. Cultivable yeasts associated with marine sponges in the Gulf of Thailand, South China Sea. Antonie Leeuwenhoek 2021, 114, 253–274. [Google Scholar] [CrossRef]
- Kaewkrajay, C.; Chanmethakul, T.; Limtong, S. Assessment of diversity of culturable marine yeasts associated with corals and zoanthids in the Gulf of Thailand, South China Sea. Microorganisms 2020, 8, 474. [Google Scholar] [CrossRef] [Green Version]
- Masiulionis, V.E.; Pagnocca, F.C. Rhodosporidiobolus geoffroeae sp. nov., a basidiomycetous yeast isolated from the waste deposit of the attine ant Acromyrmex lundii. Int. J. Syst. Evol. Microbiol. 2017, 67, 1028–1032. [Google Scholar] [CrossRef]
- Ravi, K.; Garcia-Hidalgo, J.; Gorwa-Grauslund, M.F.; Liden, G. Conversion of lignin model compounds by Pseudomonas putida KT2440 and isolates from compost. Appl. Microbiol. Biotechnol. 2017, 101, 5059–5070. [Google Scholar] [CrossRef] [Green Version]
- Lubbers, R.J.M.; Dilokpimol, A.; Nousiainen, P.A.; Visser, J.; Bruijnincx, P.C.A.; de Vries, R.P.; Cioc, R.C. Vanillic acid and methoxyhydroquinone production from guaiacyl units and related aromatic compounds using Aspergillus niger cell factories. Microb. Cell Fact. 2021, 20, 151. [Google Scholar] [CrossRef]
- Shen, Y.; Li, H.X.; Wang, X.N.; Zhang, X.R.; Hou, J.; Wang, L.F.; Gao, N.; Bao, X.M. High vanillin tolerance of an evolved Saccharomyces cerevisiae strain owing to its enhanced vanillin reduction and antioxidative capacity. J. Ind. Microbiol. Biotechnol. 2014, 41, 1637–1645. [Google Scholar] [CrossRef]
- Falconnier, B.; Lapierre, C.; Lesagemeessen, L.; Yonnet, G.; Brunerie, P.; Colonnaceccaldi, B.; Corrieu, G.; Asther, M. Vanillin as a product of ferulic acid biotransformation by the white-rot fungus Pycnoporus cinnabarinus I-937: Identification of metabolic pathways. J. Biotechnol. 1994, 37, 123–132. [Google Scholar] [CrossRef]
- Ronnander, J.; Ljunggren, J.; Hedenstrom, E.; Wright, S.A.I. Biotransformation of vanillin into vanillyl alcohol by a novel strain of Cystobasidium laryngis isolated from decaying wood. AMB Express 2018, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, P.; Singh, N.K.; Tupe, R.; Odenath, A.; Lali, A. Biotransformation of corn bran derived ferulic acid to vanillic acid using engineered Pseudomonas putida KT2440. Prep. Biochem. Biotechnol. 2020, 50, 341–348. [Google Scholar] [CrossRef]
- Wang, J.X.; Liang, J.D.; Gao, S. Biodegradation of lignin monomers vanillic, p-coumaric, and syringic acid by the bacterial strain, Sphingobacterium sp. HY-H. Curr. Microbiol. 2018, 75, 1156–1164. [Google Scholar] [CrossRef]
- Gu, H.Q.; Zhu, Y.Y.; Peng, Y.F.; Liang, X.J.; Liu, X.G.; Shao, L.Z.; Xu, Y.Y.; Xu, Z.H.; Liu, R.; Li, J. Physiological mechanism of improved tolerance of Saccharomyces cerevisiae to lignin-derived phenolic acids in lignocellulosic ethanol fermentation by short-term adaptation. Biotechnol. Biofuels 2019, 12, 268. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.J.; Fels, M.; Dragone, G.; Mussatto, S.I. Effects of inhibitory compounds derived from lignocellulosic biomass on the growth of the wild-type and evolved oleaginous yeast Rhodosporidium toruloides. Ind. Crop. Prod. 2021, 170, 113799. [Google Scholar] [CrossRef]
- Margesin, R.; Gander, S.; Zacke, G.; Gounot, A.M.; Schinner, F. Hydrocarbon degradation and enzyme activities of cold-adapted bacteria and yeasts. Extremophiles 2003, 7, 451–458. [Google Scholar] [CrossRef]
- Prem, E.M.; Mutschlechner, M.; Stres, B.; Illmer, P.; Wagner, A.O. Lignin intermediates lead to phenyl acid formation and microbial community shifts in meso- and thermophilic batch reactors. Biotechnol. Biofuels 2021, 14, 27. [Google Scholar] [CrossRef]
- Buzzini, P.; Turchetti, B.; Yurkov, A. Extremophilic yeasts: The toughest yeasts around? Yeast 2018, 35, 487–497. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margesin, R.; Ludwikowski, T.M.; Kutzner, A.; Wagner, A.O. Low-Temperature Biodegradation of Lignin-Derived Aromatic Model Monomers by the Cold-Adapted Yeast Rhodosporidiobolus colostri Isolated from Alpine Forest Soil. Microorganisms 2022, 10, 515. https://doi.org/10.3390/microorganisms10030515
Margesin R, Ludwikowski TM, Kutzner A, Wagner AO. Low-Temperature Biodegradation of Lignin-Derived Aromatic Model Monomers by the Cold-Adapted Yeast Rhodosporidiobolus colostri Isolated from Alpine Forest Soil. Microorganisms. 2022; 10(3):515. https://doi.org/10.3390/microorganisms10030515
Chicago/Turabian StyleMargesin, Rosa, Thomas Marek Ludwikowski, Andrea Kutzner, and Andreas Otto Wagner. 2022. "Low-Temperature Biodegradation of Lignin-Derived Aromatic Model Monomers by the Cold-Adapted Yeast Rhodosporidiobolus colostri Isolated from Alpine Forest Soil" Microorganisms 10, no. 3: 515. https://doi.org/10.3390/microorganisms10030515
APA StyleMargesin, R., Ludwikowski, T. M., Kutzner, A., & Wagner, A. O. (2022). Low-Temperature Biodegradation of Lignin-Derived Aromatic Model Monomers by the Cold-Adapted Yeast Rhodosporidiobolus colostri Isolated from Alpine Forest Soil. Microorganisms, 10(3), 515. https://doi.org/10.3390/microorganisms10030515