
Antimicrobial Impacts of Microbial Metabolites on the Preservation of Fish and Fishery Products: A Review with Current Knowledge

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
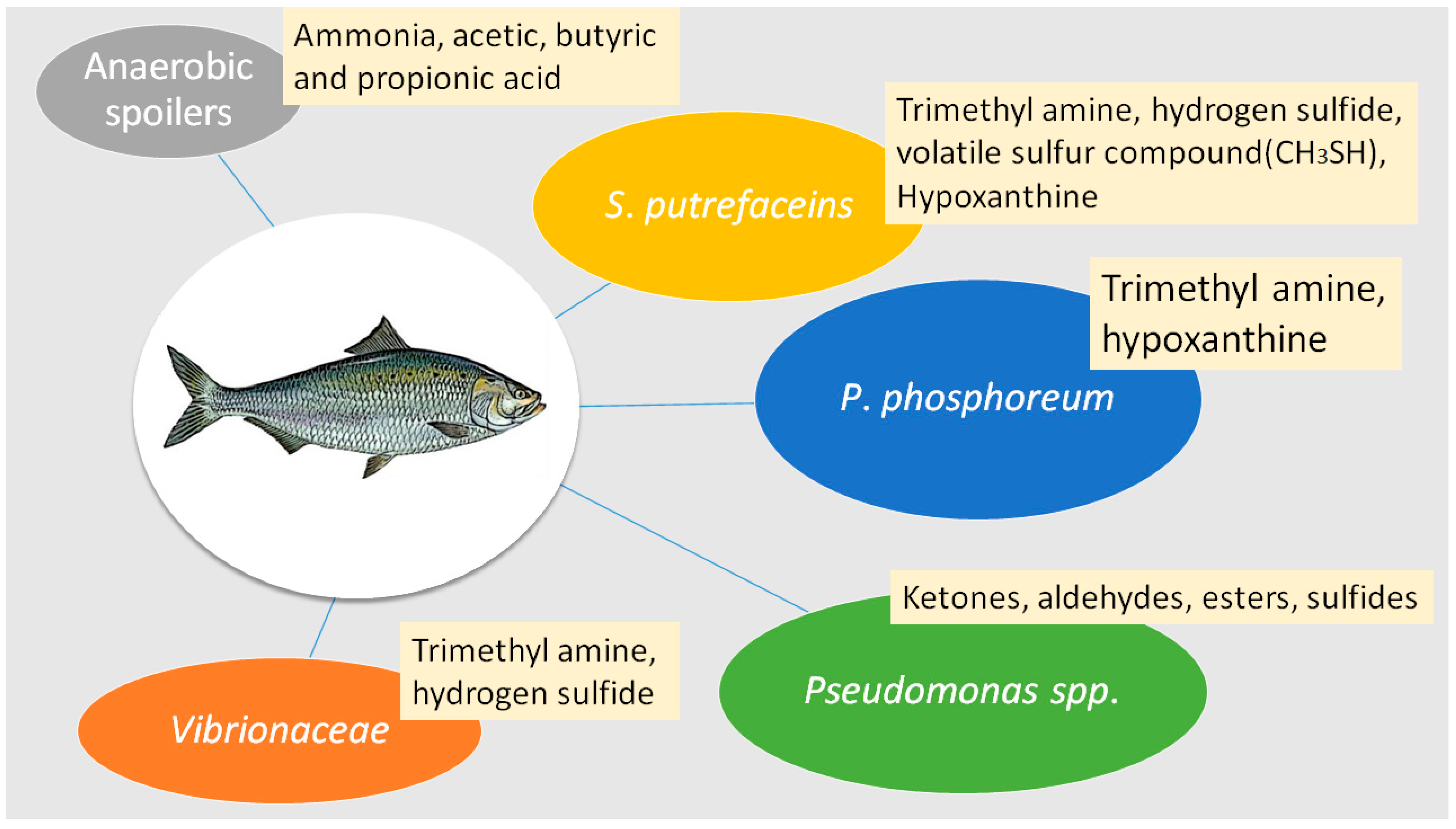
2. Microbial Spoilage of Fish and Fishery Products
3. Antimicrobial Mechanism of Microbial Metabolites
3.1. Bacteriocins

{kind=link}
{kind=link}
| Source/Bacteriocin/Organic Acid | Evaluated Matrix | Preservative Effect | Bibliographic References |
|---|---|---|---|
| Bacteriocins | |||
| Combination of glazing with nisin and irradiations treatment at 2 kGy and 5 kGy | Seer fish (Scomberomorous guttatus) steaks | Both treatments significantly improved the shelf-life of steaks from 7 days to 32 days (2 kGy) and 42 days (5 kGy) by lowering the microbial and oxidative changes in the steaks. | [84] |
| Combination of nisin (10 µg/g) with high-pressure processing (450 and 600 MPa) at low temperature (−30 °C) | Dry-cured cold-smoked salmon | The combination treatments significantly inhibited the Listeria innocua and other spoilage microbiota of the salmon compared to the control. Additionally, combined treatments improved the sensory properties, peelability and consumer preference of salmon. | [85] |
| Nisin enriched osmotic solution and vacuum packaging at chilled temperature | Tuna fillets | The combined effect of nisin, osmotic solution and vacuum packaging significantly improved the shelf-life of tuna fillets from 10 days to 51 days at 5 °C by significantly reducing spoilage microorganisms and chemical changes during storage. | [86] |
| Nisin encapsulated in chitosan microcapsules | Small yellow croaker (Pseudosciaena polyactis) | The prepared nisin loaded chitosan microcapsules significantly reduced microbial growth, lipid oxidation, and protein degradation compared to alone hurdle or control, thereby extending shelf-life of croaker by 6–9 days. | [87] |
| Nisin in combination with tea polyphenols during chilled storage | Tortoise meat (Trachemys scripta elegans) | The combined effect of nisin and tea polyphenol treatment to tortoise meat reduced microbial growth, chemical changes and retarded water loss compared to the control. | [88] |
| Nisin in combination with chitosan. Stored at 4 °C for 8 days | Large yellow croaker (Pseudosciaena crocea) | Chitosan (1%) with nisin (0.6%) showed higher efficiency in controlling water loss and other physicochemical indexes, as well as lowered chemical and microbial changes in the fish. | [89] |
| Nisin-loaded nano-carriers with essential oils (EO) micro-emulsion (EOs used were rosemary, thyme, oregano, and dittany) | Not applied | Enhanced antibacterial activities against Lactococcuslactis, Staphylococcus aureus, Listeria monocytogens, and Bacillus cereus. | [83] |
| Biodegradable packaging material (poly lactic acid/sawdust particles) incorporated with bacteriocin 7293 | Pangasius fish fillets | Bioactive film effectively inhibited Gram-positive (Listeria monocytogens and S. aureus) and Gram-negative (Pseudomonas aeruginosa, Aeromonas hydrphila, Escherichia coli and Salmonella thyphimurium) bacteria. | [90] |
| Reuterin produced by Lactobacillus reuteri INIA P579 | Cold smoked salmon | Reuterin effectively inhibited the three different strains of Listeria monocytogens in tryptic soy broth assay with concentration range of 2–4 AU/mL. When reuterin was tested on cold-smoked salmon at 8 °C for 15 days and 30 °C for 48 h, L. monocytogens counts lowered by 2.0 and 1.0 log (CFU/g) compared to the control. | [102] |
| Coagulin L1208 from Bacillus coagulans L1208 | Yellow croaker (Pseudosciaena crocea) | Bacteriocin Coagulin L1208 inhibited total viable count, Pseudomonadaceae, Shewanella, Photobacterium and Enterobacteriaceae by producing bacteriostatic ingredients. | [103] |
| Enterococcus mundtii STw38 | Fish paste from fresh hake | The bacteriocin inclusion could inhibit native fish spoilage microbiota, especially when packed under vacuum. | [104] |
| Streptococcus infantarius | Trout and tilapia | Inclusion of bacteriocin completely inhibited Escherichia coli, Staphylococcus aureus and Listeria monocytogenes in fish media. While used as wrapping material, it lowered total aerobic count. | [105] |
| Organic acids | |||
| Aromatic vinegar | Salmon fillets | No impacts on total viable counts and Enterobacteriaceae was found. Superior inhibition of Pseudomonas spp. and Psychrotrophic count was observed. Aromatic vinegar had combined effects of phenolics and organic acids in inhibition of microorganisms. | [106] |
| Citric acid and lactic acid | European hake and megrim | Inhibition of aerobic, anaerobic, psychrotrohic count and Enterobacteriaceae population | [107] |
| Acetic and ascorbic acid spray | Silver carp (Hypophthalmichthys molitrix) | Combination of acetic (1%) and ascorbic (2%) acid exhibited higher inhibition of total viable counts than individual treatments. | [108] |
| Acetic and citric acid pre-treatment | Bolti Fish (Tilapia nilotica) | Combination of acetic acid and citric acid (1 and 3%) exhibited highest inhibition of total viable bacterial count, psychrophilic bacteria, coliform and yeast and mould count. However, the difference was non- significant amongst group and significant in comparison to control. | [109] |
| Potassium acetate and potassium lactate | Catfish fillet | Combination of organic acid inhibited psychrotrophic bacterial count and extended shelf-life by additional four days. | [110] |
| Ascorbic, citric and lactic acid based icing | Hake, megrim and angler | Organic acid at 800 mg/kg concentration inhibited mesophilic aerobes in hake, megrim and angler. Additionally, inhibition of psychrophilic and proteolytic bacteria was also found. | [111] |
| Sodium acetate, sodium lactate or sodium citrate | Salmon | Levels of 2.5% exhibited activity against aerobic and psychrotrophic mircroorganisms, Pseudomonas spp., H2-S prodcuing, lactic acid and Entoerobacteriaceae bacteria. | [112] |
3.2. Reuterin
3.3. Bacteriophages and Endolysins
3.4. Organic Acids
3.5. Other Metabolites
| Seafood Products | Lactic Acid Bacteria (LAB) | Effect | Bibliographic References |
|---|---|---|---|
| Not Applied | LAB isolated from intestine of Oreochromis sp. Live LAB cells, cell-free supernatant (CFS), alkaline CFS, and heated CFS | Anti-listeria activity, antagonistic activity. | [145] |
| White leg shrimp | Lactic acid bacterium (Pediococcus pentosaceus LJR1) | Inhibited Staphyloccoccus typhi (MIC-7.81 µg/mL) and Listeria monocytogenes (MIC-15.63 µg/mL) by causing craters and belbs on the microbial surface. Reduction of L. monocytogenes by 1 log on shrimp was also found. | [151] |
| Shrimp (Penaeus vannamei) | Lactobacillus plantarum FGC-12 | Significant reduction of total viable count of Vibrio parahaemolyticus was observed. Bacteriocin disrupted bacterial cell wall causing lysis. | [152] |
| Ready-to-eat fish products (sliced surimi and tuna paste) | Latilactobacillus sakei CTC494 | The microbial inhibition by bacteriocin was dependent upon the product and exhibited antagonistic and mutual interaction on lag phase. | [153] |
| Sea bass | Mixture of lactic acid bacteria | Antagonistic effect was observed for samples treated with bacteriocin and essential oils. Inhibition of mesophilic aerobic plate count and psychrotrophic bacterial count was observed. Complete inhibition of Listeria monocytogenes, coliform, yeast and mold during storage was observed. | [154] |
| Horse Mackerel fillet | Lactobacillus plantarum and Lactobacillus sakei | Inclusion of culture reducing Staphylococcus aureus by 1 log cycle was observed. | [155] |
| Mussels | Lactobacillus plantarum | Inhibition of Vibrio spp. was obtained using bacteriocin. | [156] |
| Salmon dill gravlax | Spraying of selected LAB cultures and vacuum packaging stored at 8 °C | The strain Carnobacteriummaltaromaticum SF1944 had antimicrobial activity against spoilage microbiota and Listeria monocytogens. On the other hand, the strain Vagococcus fluvialis CD264 had mild antimicrobial activity, but extended the sensory quality of salmon by more than 25 days. | [16] |
| Three different types of cold-smoked salmon | Spraying method using bacteriocins producing three different strains: L.curvatus, Carnobacterium maltaromaticum, and Lactobacillus sakei CTC494. Vacuum packaging and storage temperature of 8 °C | Lactobacillus sakei CTC494 inhibited the growth of Listeria monocytogens and other spoilage microbiota even after 21 days of study, thereby increasing shelf-life of all three types of smoked salmon. However, other two strains limited the pathogens’ growth depending on the type of smoked salmon product. Hence, Lactobacillus sakei CTC494 was recommended as a biopreservative for smoked salmon. | [146] |
| Fish paste | Bacteriocins producing LAB strain (Enterococcus mundtii). Vacuum packed and stored at 4 °C | Enterococcus mundtii STw38 showed highest activities against Gram-positive bacteria including Listeria innocua and L. plantarum. Additionally, STw38 strain survived and produce bacteriocins at 4 °C. | [104] |
| Shrimp (Litopenaeus vannamei) | Combined culture of Lactobacillus plantarum AB-1 and Lactobacillus casei applied on the shrimp at refrigerated temperature | The application of combined LAB cultures significantly reduced spoilage microorganisms, mainly Shewanella baltica, total volatile base and pH of the shrimp, thereby increasing the shelf-life of the shrimp. | [148] |
| Peeled and cooked shrimp (Penaeus vannamei) | Combination of Lactococcus piscium CNCM I-4031 and C. divergens V41 applied on the shrimp and packed in modified atmospheric packaging (50% CO2 and 50% N2) at 8 °C temperature | The results indicated that shrimp treated with combined cultures had higher sensorial properties and lowered microbial and chemical changes at the end of storage time (28 days) compared to the treatment with single LAB culture. | [149] |
| Cod and salmon based products | Six LAB strains with no histamine production ability, in combination with other hurdles including chitosan, modified atmosphere packaging (MAP), and super chilling. | Improved sensory properties and reduced microbial and chemical changes in cod and salmon products. | [69] |
| Young hake (Merluccius merluccius) and megrim (Lepidorhombus boscii) | (1) The multi-bacteriocinogenic L. curvatus BCS35 culture; (2) their CFS; (3) Lyophilized bacteriocin powder stored with ice at 0–2 °C for 14 days | The BCS35 culture and their CFS significantly lowered spoilage microorganism as well as Listeria spp. Additionally, the sensory properties of both fish were maintained during storage days. | [150] |
| Cold-smoked salmon | Lactococcus piscium EU2241 strain | Lactococcus piscium EU2241 prevented the spoilage caused by Brochothrix thermosphacta and Serratia proteamaculans by acultural and cultural method in cold-smoked salmon, thereby maintaining the sensory properties of the product. | [147] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Quinto, E.J.; Caro, I.; Villalobos-Delgado, L.H.; Mateo, J.; De-Mateo-Silleras, B.; Redondo-Del-Río, M.P. Food Safety through Natural Antimicrobials. Antibiotics 2019, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, A.; Abraha, B.; Samuel, M.; Mohammedidris, H.; Abraham, W.; Mahmud, E. Fish Preservation: A Multi-Dimensional Approach. MOJ Food Process. Technol 2018, 6, 303–310. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial Metabolites in Nutrition, Healthcare and Agriculture. 3 Biotech 2017, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Banerjee, R.; Dwivedi, H.; Juneja, V. Bacteriocins|Potential in Food Preservation. Encycl. Food Microbiol. 2014, 1, 180–186. [Google Scholar]
- Xu, D.-P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.-J.; Li, H.-B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.M.; Iatropoulos, M.; Whysner, J. Safety Assessment of Butylated Hydroxyanisole and Butylated Hydroxytoluene as Antioxidant Food Additives. Food Chem. Toxicol. 1999, 37, 1027–1038. [Google Scholar] [CrossRef]
- Adolfsson-Erici, M.; Pettersson, M.; Parkkonen, J.; Sturve, J. Triclosan, a Commonly Used Bactericide Found in Human Milk and in the Aquatic Environment in Sweden. Chemosphere 2002, 46, 1485–1489. [Google Scholar] [CrossRef]
- Cammack, R.; Joannou, C.; Cui, X.-Y.; Martinez, C.T.; Maraj, S.R.; Hughes, M.N. Nitrite and Nitrosyl Compounds in Food Preservation. Biochim. Biophys. Acta (BBA)-Bioenerg. 1999, 1411, 475–488. [Google Scholar] [CrossRef] [Green Version]
- Tavares, R.S.; Martins, F.C.; Oliveira, P.J.; Ramalho-Santos, J.; Peixoto, F.P. Parabens in Male Infertility—Is There a Mitochondrial Connection? Reprod. Toxicol. 2009, 27, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vally, H.; Misso, N.L.; Madan, V. Clinical Effects of Sulphite Additives. Clin. Exp. Allergy 2009, 39, 1643–1651. [Google Scholar] [CrossRef]
- Gultekin, F.; Doguc, D.; Husamettin, V.; Taysi, E. The Effects of Food and Food Additives on Behaviors. Int. J. Health Nutr. 2013, 4, 21–32. [Google Scholar]
- Martyn, D.M.; McNulty, B.A.; Nugent, A.P.; Gibney, M.J. Food Additives and Preschool Children. Proc. Nutr. Soc. 2013, 72, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devcich, D.A.; Pedersen, I.K.; Petrie, K.J. You Eat What You Are: Modern Health Worries and the Acceptance of Natural and Synthetic Additives in Functional Foods. Appetite 2007, 48, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.; Roopesh, M. Synergistically Enhanced Salmonella Typhimurium Reduction by Sequential Treatment of Organic Acids and Atmospheric Cold Plasma and the Mechanism Study. Food Microbiol. 2021, 104, 103976. [Google Scholar] [CrossRef]
- Rathod, N.B.; Ranveer, R.C.; Benjakul, S.; Kim, S.-K.; Pagarkar, A.U.; Patange, S.; Ozogul, F. Recent Developments of Natural Antimicrobials and Antioxidants on Fish and Fishery Food Products. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4182–4210. [Google Scholar] [CrossRef]
- Wiernasz, N.; Leroi, F.; Chevalier, F.; Cornet, J.; Cardinal, M.; Rohloff, J.; Passerini, D.; Skırnisdóttir, S.; Pilet, M.-F. Salmon Gravlax Biopreservation with Lactic Acid Bacteria: A Polyphasic Approach to Assessing the Impact on Organoleptic Properties, Microbial Ecosystem and Volatilome Composition. Front. Microbiol. 2020, 10, 3103. [Google Scholar] [CrossRef] [PubMed]
- Rathod, N.B.; Phadke, G.G.; Tabanelli, G.; Mane, A.; Ranveer, R.C.; Pagarkar, A.; Ozogul, F. Recent Advances in Bio-Preservatives Impacts of Lactic Acid Bacteria and Their Metabolites on Aquatic Food Products. Food Biosci. 2021, 44, 101440. [Google Scholar] [CrossRef]
- Ağagündüz, D.; Yılmaz, B.; Şahin, T.Ö.; Güneşliol, B.E.; Ayten, Ş.; Russo, P.; Spano, G.; Rocha, J.M.; Bartkiene, E.; Özogul, F. Dairy Lactic Acid Bacteria and Their Potential Function in Dietetics: The Food–Gut-Health Axis. Foods 2021, 10, 3099. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and Their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2020, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trakselyte-Rupsiene, K.; Juodeikiene, G.; Alzbergaite, G.; Zadeike, D.; Bartkiene, E.; Ozogul, F.; Ruller, L.; Robert, J.; Rocha, J.M.F. Bio-Refinery of Plant Drinks Press Cake Permeate Using Ultrafiltration and Lactobacillus fermentation into Antimicrobials and Its Effect on the Growth of Wheatgrass in Vivo. Food Biosci. 2021, 46, 101427. [Google Scholar] [CrossRef]
- Kuley, E.; Özyurt, G.; Özogul, I.; Boga, M.; Akyol, I.; Rocha, J.M.; Özogul, F. The Role of Selected Lactic Acid Bacteria on Organic Acid Accumulation during Wet and Spray-Dried Fish-Based Silages. Contributions to the Winning Combination of Microbial Food Safety and Environmental Sustainability. Microorganisms 2020, 8, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novotni, D.; Gänzle, M.; Rocha, J.M. Composition and Activity of Microbiota in Sourdough and Their Effect on Bread Quality and Safety. Trends Wheat Bread Mak. 2021, 129–172. [Google Scholar] [CrossRef]
- Păcularu-Burada, B.; Georgescu, L.A.; Vasile, M.A.; Rocha, J.M.; Bahrim, G.-E. Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs. Microorganisms 2020, 8, 643. [Google Scholar] [CrossRef] [PubMed]
- Păcularu-Burada, B.; Turturică, M.; Rocha, J.M.; Bahrim, G.-E. Statistical Approach to Potentially Enhance the Postbiotication of Gluten-Free Sourdough. Appl. Sci. 2021, 11, 5306. [Google Scholar] [CrossRef]
- Sharma, N.; Kaur, G.; Khatkar, S.K. Optimization of Emulsification Conditions for Designing Ultrasound Assisted Curcumin Loaded Nanoemulsion: Characterization, Antioxidant Assay and Release Kinetics. LWT 2021, 141, 110962. [Google Scholar] [CrossRef]
- Zokaityte, E.; Cernauskas, D.; Klupsaite, D.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Ruzauskas, M.; Gruzauskas, R.; Juodeikiene, G.; Rocha, J.M.; et al. Bioconversion of Milk Permeate with Selected Lactic Acid Bacteria Strains and Apple By-Products into Beverages with Antimicrobial Properties and Enriched with Galactooligosaccharides. Microorganisms 2020, 8, 1182. [Google Scholar] [CrossRef]
- Gökoğlu, N.; Cengız, E.; Yerlıkaya, P. Determination of the Shelf Life of Marinated Sardine (Sardina pilchardus) Stored at 4 C. Food Control. 2004, 15, 1–4. [Google Scholar] [CrossRef]
- Nirmal, N.P.; Benjakul, S. Effect of Green Tea Extract in Combination with Ascorbic Acid on the Retardation of Melanosis and Quality Changes of Pacific White Shrimp during Iced Storage. Food Bioprocess Technol. 2012, 5, 2941–2951. [Google Scholar] [CrossRef]
- Amaral, A.B.; Silva, M.V.D.; Lannes, S.C.D.S. Lipid Oxidation in Meat: Mechanisms and Protective Factors–A Review. Food Sci. Technol. 2018, 38, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kontominas, M.G.; Badeka, A.V.; Kosma, I.S.; Nathanailides, C.I. Innovative Seafood Preservation Technologies: Recent Developments. Animals 2021, 11, 92. [Google Scholar] [CrossRef]
- Mei, J.; Ma, X.; Xie, J. Review on Natural Preservatives for Extending Fish Shelf Life. Foods 2019, 8, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez, J.A.; González, M.; Murado, M. Effects of Lactic Acid Bacteria Cultures on Pathogenic Microbiota from Fish. Aquaculture 2005, 245, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Taylor, T.M.; Doores, S.X. Organic Acids. In Antimicrobials in Food; CRC Press: Boca Raton, FL, USA, 2020; pp. 133–190. [Google Scholar]
- USDA-FSIS. FSIS Directive 7120.1, Revision 56. Safe and Suitable Ingredients Used in the Production of Meat and Poultry, and Egg Products; USDA-FSIS: Washington, DC, USA, 2019.
- Ghanbari, M.; Jami, M.; Domig, K.J.; Kneifel, W. Seafood Biopreservation by Lactic Acid Bacteria–A Review. LWT-Food Sci. Technol. 2013, 54, 315–324. [Google Scholar] [CrossRef]
- EU Commission. Commission Regulation (EU) No 1129/2011 of 11 November 2011 Amending Annex II to Regulation (EC) No 1333/2008 of the European Parliament and of the Council by Establishing a Union List of Food Additives. Off. J. Eur. Union L 2011, 295, 1–177. Available online: https://eur-lex.europa.eu/eli/reg/2011/1129/oj (accessed on 14 March 2022).
- Food Additives Permitted for Direct Addition to Food for Human Consumption; Bacteriophage Preparation. Available online: https://www.federalregister.gov/documents/2006/08/18/E6-13621/food-additives-permitted-for-direct-addition-to-food-for-human-consumption-bacteriophage-preparation (accessed on 14 March 2022).
- Davidson, P.M.; Taylor, T.M.; David, J.R. Antimicrobials in Food; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Bou, R.; Claret, A.; Stamatakis, A.; Martínez, B.; Guerrero, L. Quality Changes and Shelf-Life Extension of Ready-to-Eat Fish Patties by Adding Encapsulated Citric Acid. J. Sci. Food Agric. 2017, 97, 5352–5360. [Google Scholar] [CrossRef] [PubMed]
- USDA; HHS. Dietary Guidelines for Americans, 2020–2025, 9th ed.; USDA: Washington, DC, USA, 2020.
- FAO. The State of World Fisheries and Aquaculture 2016. In Contributing to Food Security and Nutrition for All; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; p. 200. [Google Scholar]
- Yearbook, F. Fishery and Aquaculture Statistics 2016; FAO: Rome, Italy, 2019. [Google Scholar]
- Zhuang, S.; Hong, H.; Zhang, L.; Luo, Y. Spoilage-Related Microbiota in Fish and Crustaceans during Storage: Research Progress and Future Trends. Compr. Rev. Food Sci. Food Saf. 2021, 20, 252–288. [Google Scholar] [CrossRef] [PubMed]
- Khalili Tilami, S.; Sampels, S. Nutritional Value of Fish: Lipids, Proteins, Vitamins, and Minerals. Rev. Fish. Sci. Aquac. 2018, 26, 243–253. [Google Scholar] [CrossRef]
- Balami, S.; Sharma, A.; Karn, R. Significance of Nutritional Value of Fish for Human Health. Malays. J. Halal Res. 2019, 2, 32–34. [Google Scholar] [CrossRef] [Green Version]
- Erkmen, O.; Bozoglu, T.F. Food Microbiology, 2 Volume Set: Principles into Practice; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Cold Plasma for the Preservation of Aquatic Food Products: An Overview-Rathod-2021-Comprehensive Reviews in Food Science and Food Safety-Wiley. Available online: https://ift.onlinelibrary.wiley.com/doi/abs/10.1111/1541-4337.12815 (accessed on 14 March 2022).
- Rathod, N.B.; Kulawik, P.; Ozogul, Y.; Ozogul, F.; Bekhit, A.E.-D.A. Recent Developments in Non-Thermal Processing for Seafood and Seafood Products: Cold Plasma, Pulsed Electric Field and High Hydrostatic Pressure. Int. J. Food Sci. Technol. 2022, 57, 774–790. [Google Scholar] [CrossRef]
- Bocker, H. JC Ayres, JO Mundt and WE Sandine, Microbiology of Foods. XII+ 708 S., 134 Abb., 66 Table 20 Tafeln. San Francisco 1980; Wiley Online Library: Hoboken, NJ, USA, 1981. [Google Scholar]
- Shewan, J. The Bacteriology of Fresh and Spoiling Fish and Some Related Chemical Changes. Recent Adv. Food Sci. 1962, 1, 167–193. [Google Scholar]
- LC, N.; Ajani, E.; Bamidele, S. Use of Lactic Acid Bacteria from Nile Tilapia Oreochromis Niloticus as Probiotics for Sustainable Production and Improvement in Fish Welfare. Isr. J. Aquac.-Bamidgeh 2014, 66, 1–10. [Google Scholar]
- Boziaris, I.S.; Parlapani, F.F. Specific Spoilage Organisms (SSOs) in Fish. In The Microbiological Quality of Food; Elsevier: Amsterdam, The Netherlands, 2017; pp. 61–98. [Google Scholar]
- Chen, X.-L.; Wang, Y.; Wang, P.; Zhang, Y.-Z. Proteases from the Marine Bacteria in the Genus Pseudoalteromonas: Diversity, Characteristics, Ecological Roles, and Application Potentials. Mar. Life Sci. Technol. 2020, 2, 309–323. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Alegbeleye, O.O.; Strateva, M.; Stratev, D. Understanding Spoilage Microbial Community and Spoilage Mechanisms in Foods of Animal Origin. Compr. Rev. Food Sci. Food Saf. 2020, 19, 311–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewan, J.M. The Bacteriology of Fresh and Spoiling Fish and the Biochemical Changes Induced by Bacterial Sections. Sci. Inf. Database 1977, 36, 328–334. [Google Scholar]
- Al Bulushi, I.M.; Poole, S.E.; Barlow, R.; Deeth, H.C.; Dykes, G.A. Speciation of Gram-Positive Bacteria in Fresh and Ambient-Stored Sub-Tropical Marine Fish. Int. J. Food Microbiol. 2010, 138, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Surendran, P.; Joseph, J.; Shenoy, A.; Perigreen, P.; Iyer, K.M.; Gopakumar, K. Studies of Spoilage of Commercially Important Tropical Fishes under Iced Storage. Fish. Res. 1989, 7, 1–9. [Google Scholar] [CrossRef]
- Houicher, A.; Bensid, A.; Regenstein, J.M.; Özogul, F. Control of Biogenic Amine Production and Bacterial Growth in Fish and Seafood Products Using Phytochemicals as Biopreservatives: A Review. Food Biosci. 2021, 39, 100807. [Google Scholar] [CrossRef]
- Fujioka, R.S.; Tenno, K.; Kansako, S. Naturally Occurring Fecal Coliforms and Fecal Streptococci in Hawaii’s Freshwater Streams. Toxic. Assess. 1988, 3, 613–630. [Google Scholar] [CrossRef]
- Dalgaard, P.; Madsen, H.; Samieian, N.; Emborg, J. Biogenic Amine Formation and Microbial Spoilage in Chilled Garfish (Belone belone belone)–Effect of Modified Atmosphere Packaging and Previous Frozen Storage. J. Appl. Microbiol. 2006, 101, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Emborg, J.; Laursen, B.G.; Dalgaard, P. Significant Histamine Formation in Tuna (Thunnus albacares) at 2 C—Effect of Vacuum-and Modified Atmosphere-Packaging on Psychrotolerant Bacteria. Int. J. Food Microbiol. 2005, 101, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Dalgaard, P. Fish Spoilage Bacteria–Problems and Solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef]
- Gram, L.; Huss, H.H. Fresh and Processed Fish and Shellfish. In The Microbiological Safety and Quality of Food; Aspen Publishers: New York, NY, USA, 2000; pp. 472–506. [Google Scholar]
- Pagarkar, A.; Basu, S.; Mitra, A. storage characteristics of fish soya based ready to cook extruded product. Asian J. Microbiol. Biotechnol. Environ. Sci. 2006, 8, 609. [Google Scholar]
- Venugopal, V. Enzymes from Seafood Processing Waste and Their Applications in Seafood Processing. Adv. Food Nutr. Res. 2016, 78, 47–69. [Google Scholar] [PubMed]
- Shewan, J. The Microbiology of Sea-Water Fish. Fish Food 1961, 1, 487–560. [Google Scholar]
- Lerke, P.; Farber, L.; Adams, R. Bacteriology of Spoilage of Fish Muscle: IV. Role of Protein. Appl. Microbiol. 1967, 15, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Storroe, I.; Dyrset, N.; Larsen, H. Bacterial Decomposition Offish Stored in Bulk. 2. Enumer. Charact. Anaerob. Ammon.-Prod. Bact. 1975. [Google Scholar]
- Wiernasz, N.; Cornet, J.; Cardinal, M.; Pilet, M.-F.; Passerini, D.; Leroi, F. Lactic Acid Bacteria Selection for Biopreservation as a Part of Hurdle Technology Approach Applied on Seafood. Front. Mar. Sci. 2017, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Ng, Z.J.; Zarin, M.A.; Lee, C.K.; Tan, J.S. Application of Bacteriocins in Food Preservation and Infectious Disease Treatment for Humans and Livestock: A Review. RSC Adv. 2020, 10, 38937–38964. [Google Scholar] [CrossRef]
- Darbandi, A.; Asadi, A.; Mahdizade Ari, M.; Ohadi, E.; Talebi, M.; Halaj Zadeh, M.; Darb Emamie, A.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and Potential Use as Antimicrobials. J. Clin. Lab. Anal. 2022, 36, e24093. [Google Scholar] [CrossRef]
- Kaya, H.İ.; Özel, B.; Şimşek, Ö. A Natural Way of Food Preservation: Bacteriocins and Their Applications. In Health and Safety Aspects of Food Processing Technologies; Springer: Berlin/Heidelberg, Germany, 2019; pp. 633–659. [Google Scholar]
- Sharma, G.; Dang, S.; Gupta, S.; Gabrani, R. Antibacterial Activity, Cytotoxicity, and the Mechanism of Action of Bacteriocin from Bacillus subtilis GAS101. Med. Princ. Pract. 2018, 27, 186–192. [Google Scholar] [CrossRef]
- YALÇIN, H.; Üstündağ, H. Bacteriocins and Their Use in Food Products. Mehmet Akif Ersoy Univ. J. Health Sci. Inst. 2017, 5, 53–65. [Google Scholar]
- Punyauppa-path, S.; Phumkhachorn, P.; Rattanachaikunsopon, P. Nisin: Production and Mechanism of Antimicrobial Action. Int. J. Curr. Res. Rev. 2015, 7, 47. [Google Scholar]
- Maryam, H.; Maqsood, S.; Farooq, U. Bacteriocins: A Novel Weapon against Emerging Resistance. Saudi J. Pathol. Microbiol. 2017, 2, 220–227. [Google Scholar]
- Bachmann, D.E.C.; Leroy, F. Use of Bioprotective Cultures in Fish Products. Curr. Opin. Food Sci. 2015, 6, 19–23. [Google Scholar] [CrossRef]
- Anandharaj, M.; Rani, R.P.; Swain, M.R. Probioactives: Bacteriocin and Exopolysaccharides. In Advances in Probiotics; Elsevier: Amsterdam, The Netherlands, 2021; pp. 293–308. [Google Scholar]
- O’Connor, P.M.; Kuniyoshi, T.M.; Oliveira, R.P.; Hill, C.; Ross, R.P.; Cotter, P.D. Antimicrobials for Food and Feed; a Bacteriocin Perspective. Curr. Opin. Biotechnol. 2020, 61, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, N.P.; Santivarangkna, C. Bio-Preservation of Dairy Products: A Non-Thermal Processing and Preservation Approach for Shelf-Life Extension of Dairy Products. In Non-Thermal Processing Technologies for the Dairy Industry; CRC Press: Boca Raton, FL, USA, 2021; pp. 111–126. [Google Scholar]
- Stupar, J.; Holøymoen, I.G.; Hoel, S.; Lerfall, J.; Rustad, T.; Jakobsen, A.N. Diversity and Antimicrobial Activity towards Listeria spp. and Escherichia coli among Lactic Acid Bacteria Isolated from Ready-to-Eat Seafood. Foods 2021, 10, 271. [Google Scholar] [CrossRef]
- Gharsallaoui, A.; Oulahal, N.; Joly, C.; Degraeve, P. Nisin as a Food Preservative: Part 1: Physicochemical Properties, Antimicrobial Activity, and Main Uses. Crit. Rev. Food Sci. Nutr. 2016, 56, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- Chatzidaki, M.D.; Balkiza, F.; Gad, E.; Alexandraki, V.; Avramiotis, S.; Georgalaki, M.; Papadimitriou, V.; Tsakalidou, E.; Papadimitriou, K.; Xenakis, A. Reverse Micelles as Nano-Carriers of Nisin against Foodborne Pathogens. Part II: The Case of Essential Oils. Food Chem. 2019, 278, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Kakatkar, A.S.; Gautam, R.K.; Shashidhar, R. Combination of Glazing, Nisin Treatment and Radiation Processing for Shelf-Life Extension of Seer Fish (Scomberomorous guttatus) Steaks. Radiat. Phys. Chem. 2017, 130, 303–305. [Google Scholar] [CrossRef]
- Lebow, N.K.; DesRocher, L.D.; Younce, F.L.; Zhu, M.-J.; Ross, C.F.; Smith, D.M. Influence of High-Pressure Processing at Low Temperature and Nisin on Listeria Innocua Survival and Sensory Preference of Dry-Cured Cold-Smoked Salmon. J. Food Sci. 2017, 82, 2977–2986. [Google Scholar] [CrossRef] [PubMed]
- Sofra, C.; Tsironi, T.; Taoukis, P.S. Modeling the Effect of Pre-Treatment with Nisin Enriched Osmotic Solution on the Shelf Life of Chilled Vacuum Packed Tuna. J. Food Eng. 2018, 216, 125–131. [Google Scholar] [CrossRef]
- Wu, T.; Wu, C.; Fang, Z.; Ma, X.; Chen, S.; Hu, Y. Effect of Chitosan Microcapsules Loaded with Nisin on the Preservation of Small Yellow Croaker. Food Control 2017, 79, 317–324. [Google Scholar] [CrossRef]
- Yang, A.; Cheng, F.; Tong, P.; Chen, H. Effect of Tea Polyphenol and Nisin on the Quality of Tortoise (Trachemys scripta elegans) Meat during Chilled Storage. J. Food Process. Preserv. 2017, 41, e13308. [Google Scholar] [CrossRef]
- Hui, G.; Liu, W.; Feng, H.; Li, J.; Gao, Y. Effects of Chitosan Combined with Nisin Treatment on Storage Quality of Large Yellow Croaker (Pseudosciaena crocea). Food Chem. 2016, 203, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Woraprayote, W.; Pumpuang, L.; Tosukhowong, A.; Zendo, T.; Sonomoto, K.; Benjakul, S.; Visessanguan, W. Antimicrobial Biodegradable Food Packaging Impregnated with Bacteriocin 7293 for Control of Pathogenic Bacteria in Pangasius Fish Fillets. LWT 2018, 89, 427–433. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lee, N.-K.; Cho, S.-M.; Kim, K.-T.; Paik, H.-D. Inhibition of Spoilage and Pathogenic Bacteria by Lacticin NK24, a Bacteriocin Produced by Lactococcus lactis NK24 from Fermented Fish Food. Korean J. Food Sci. Technol. 1999, 31, 1035–1043. [Google Scholar]
- Kim, Y.-M.; Paik, H.-D.; Lee, D.-S. Shelf-Life Characteristics of Fresh Oysters and Ground Beef as Affected by Bacteriocin-Coated Plastic Packaging Film. J. Sci. Food Agric. 2002, 82, 998–1002. [Google Scholar] [CrossRef]
- Komora, N.; Maciel, C.; Pinto, C.A.; Ferreira, V.; Brandão, T.R.; Saraiva, J.M.; Castro, S.M.; Teixeira, P. Non-Thermal Approach to Listeria Monocytogenes Inactivation in Milk: The Combined Effect of High Pressure, Pediocin PA-1 and Bacteriophage P100. Food Microbiol. 2020, 86, 103315. [Google Scholar] [CrossRef]
- Hwanhlem, N.; Aran, H. Biopreservation of Seafood by Using Bacteriocins and Bacteriocinogenic Lactic Acid Bacteria as Potential Bio-Control Agents. Benef. Microorg. Agric. Aquac. Other Areas 2015, 29, 183–213. [Google Scholar]
- Zhu, L.; Zeng, J.; Wang, C.; Wang, J. Structural Basis of Pore Formation in the Mannose Phosphotransferase System (Man-PTS) by Pediocin PA-1. Appl. Environ. Microbiol. 2021, 88, AEM-01992. [Google Scholar]
- Pinto, A.L.; Fernandes, M.; Pinto, C.; Albano, H.; Castilho, F.; Teixeira, P.; Gibbs, P.A. Characterization of Anti-Listeria Bacteriocins Isolated from Shellfish: Potential Antimicrobials to Control Non-Fermented Seafood. Int. J. Food Microbiol. 2009, 129, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.-J.; Wu, C.-W.; Jiang, S.-T. Biopreservative Effect of Pediocin ACCEL on Refrigerated Seafood. Fish. Sci. 2007, 73, 907–912. [Google Scholar] [CrossRef]
- Sarika, A.; Lipton, A.P.; Aishwarya, M. Biopreservative Efficacy of Bacteriocin GP1 of Lactobacillus rhamnosus GP1 on Stored Fish Filets. Front. Nutr. 2019, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Behnam, S.; Anvari, M.; Rezaei, M.; Soltanian, S.; Safari, R. Effect of Nisin as a Biopreservative Agent on Quality and Shelf Life of Vacuum Packaged Rainbow Trout (Oncorhynchus mykiss) Stored at 4 °C. J. Food Sci. Technol. 2015, 52, 2184–2192. [Google Scholar] [CrossRef] [Green Version]
- Mei, J.; Shen, Y.; Liu, W.; Lan, W.; Li, N.; Xie, J. Effectiveness of Sodium Alginate Active Coatings Containing Bacteriocin EFL4 for the Quality Improvement of Ready-to-Eat Fresh Salmon Fillets during Cold Storage. Coatings 2020, 10, 506. [Google Scholar] [CrossRef]
- Fahim, H.A.; Khairalla, A.S.; El-Gendy, A.O. Nanotechnology: A Valuable Strategy to Improve Bacteriocin Formulations. Front. Microbiol. 2016, 7, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel, R.; Martín-Cabrejas, I.; Langa, S.; El Aouad, N.; Arqués, J.; Reyes, F.; Medina, M. Antimicrobial Activity of Reuterin Produced by Lactobacillus Reuteri on Listeria Monocytogenes in Cold-Smoked Salmon. Food Microbiol. 2014, 44, 1–5. [Google Scholar] [CrossRef]
- Fu, L.; Wang, C.; Ruan, X.; Li, G.; Zhao, Y.; Wang, Y. Preservation of Large Yellow Croaker (Pseudosciaena crocea) by Coagulin L1208, a Novel Bacteriocin Produced by Bacillus coagulans L1208. Int. J. Food Microbiol. 2018, 266, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Delcarlo, S.B.; Parada, R.; Schelegueda, L.I.; Vallejo, M.; Marguet, E.R.; Campos, C.A. From the Isolation of Bacteriocinogenic LAB Strains to the Application for Fish Paste Biopreservation. LWT 2019, 110, 239–246. [Google Scholar] [CrossRef]
- Pérez-Arauz, Á.-O.; Rodríguez-Hernández, A.-I.; del Rocío López-Cuellar, M.; Martínez-Juárez, V.-M.; Chavarría-Hernández, N. Films Based on Pectin, Gellan, EDTA, and Bacteriocin-like Compounds Produced by Streptococcus infantarius for the Bacterial Control in Fish Packaging. J. Food Process. Preserv. 2021, 45, e15006. [Google Scholar] [CrossRef]
- Di Toro, J.; Branciari, R.; Roila, R.; Altissimi, S.; Jang, H.; Zhou, K.; Perucci, S.; Codini, M.; Ranucci, D. Eficacy of an Aromatic Vinegar in Reducing Psychrotrophic Bacteria and Biogenic Amines in Salmon Fillets (Salmo salar) Stored in Modified Atmosphere Packaging. Pol. J. Food Nutr. Sci. 2019, 69, 397–405. [Google Scholar] [CrossRef]
- García-Soto, B.; Fernández-No, I.C.; Barros-Velázquez, J.; Aubourg, S.P. Use of Citric and Lactic Acids in Ice to Enhance Quality of Two Fish Species during On-Board Chilled Storage. Int. J. Refrig. 2014, 40, 390–397. [Google Scholar] [CrossRef]
- Monirul, I.; Yang, F.; Niaz, M.; Qixing, J.; Wenshui, X. Effectiveness of Combined Acetic Acid and Ascorbic Acid Spray on Fresh Silver Carp (Hypophthalmichthys molitrix) Fish to Increase Shelf-Life at Refrigerated Temperature. Curr. Res. Nutr. Food Sci. J. 2019, 7, 415–426. [Google Scholar] [CrossRef] [Green Version]
- El-Shemy, M.; Yasin, N.M.; Gadallah, M.; Hanafi, E. Microbiological Quality and Enzymes Activity of Refrigerated Bolti Fish (Tilapia nilotica) Pretreated with Organic Acids. J. Agric. Vet. Sci. 2016, 267, 1–14. [Google Scholar] [CrossRef]
- Kin, S.; Schilling, M.W.; Smith, B.S.; Silva, J.L.; Kim, T.; Pham, A.J.; Campano, S.G. Potassium Acetate and Potassium Lactate Enhance the Microbiological and Physical Properties of Marinated Catfish Fillets. J. Food Sci. 2011, 76, S242–S250. [Google Scholar] [CrossRef] [PubMed]
- Rey, M.S.; García-Soto, B.; Fuertes-Gamundi, J.R.; Aubourg, S.; Barros-Velázquez, J. Effect of a Natural Organic Acid-Icing System on the Microbiological Quality of Commercially Relevant Chilled Fish Species. LWT-Food Sci. Technol. 2012, 46, 217–223. [Google Scholar] [CrossRef]
- Antimicrobial and Antioxidant Effects of Sodium Acetate, Sodium Lactate, and Sodium Citrate in Refrigerated Sliced Salmon-ScienceDirect. Available online: https://www.sciencedirect.com/science/article/pii/S0956713506000247 (accessed on 14 March 2022).
- Napolean, J.; Leny, C. A Study of Antimicrobial Effects of Reuterin by Lactobacillus reuteri DSM 17938 on Escherichia coli. 1970. Available online: http://lup.lub.lu.se/student-papers/record/8996582 (accessed on 14 March 2022).
- Garde, S.; Gómez-Torres, N.; Hernández, M.; Ávila, M. Susceptibility of Clostridium Perfringens to Antimicrobials Produced by Lactic Acid Bacteria: Reuterin and Nisin. Food Control 2014, 44, 22–25. [Google Scholar] [CrossRef]
- Vollenweider, S.; Lacroix, C. 3-Hydroxypropionaldehyde: Applications and Perspectives of Biotechnological Production. Appl. Microbiol. Biotechnol. 2004, 64, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Couture, F.; Boutin, Y.; Said, L.B.; Cashman-Kadri, S.; Subirade, M.; Biron, E.; Fliss, I. In Vitro Investigation of Gastrointestinal Stability and Toxicity of 3-Hyrdoxypropionaldehyde (Reuterin) Produced by Lactobacillus Reuteri. Toxicol. Rep. 2021, 8, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Montiel, R.; Martín-Cabrejas, I.; Medina, M. Reuterin, Lactoperoxidase, Lactoferrin and High Hydrostatic Pressure on the Inactivation of Food-Borne Pathogens in Cooked Ham. Food Control 2015, 51, 122–128. [Google Scholar] [CrossRef]
- Angiolillo, L.; Conte, A.; Del Nobile, M. A New Method to Bio-Preserve Sea Bass Fillets. Int. J. Food Microbiol. 2018, 271, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Montiel, R.; Martín-Cabrejas, I.; Gaya, P.; Medina, M. Reuterin and High Hydrostatic Pressure Treatments on the Inactivation of Listeria Monocytogenes and Effect on the Characteristics of Cold-Smoked Salmon. Food Bioprocess Technol. 2014, 7, 2319–2329. [Google Scholar] [CrossRef]
- Angiolillo, L.; Conte, A.; Del Nobile, M.A. Microencapsulated Lactobacillus Reuteri Combined with Modified Atmosphere as a Way to Improve Tuna Burger Shelf Life. Int. J. Food Sci. Technol. 2017, 52, 1576–1584. [Google Scholar] [CrossRef]
- Martínez, B.; García, P.; Rodríguez, A. Swapping the Roles of Bacteriocins and Bacteriophages in Food Biotechnology. Curr. Opin. Biotechnol. 2019, 56, 1–6. [Google Scholar] [CrossRef]
- Carvalho, C.; Costa, A.R.; Silva, F.; Oliveira, A. Bacteriophages and Their Derivatives for the Treatment and Control of Food-Producing Animal Infections. Crit. Rev. Microbiol. 2017, 43, 583–601. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Kim, Y.-T.; Kim, H.B.; Choi, S.H.; Lee, J.-H. Characterization of Bacteriophage VVP001 and Its Application for the Inhibition of Vibrio vulnificus Causing Seafood-Borne Diseases. Food Microbiol. 2021, 94, 103630. [Google Scholar] [CrossRef]
- Chang, Y. Bacteriophage-Derived Endolysins Applied as Potent Biocontrol Agents to Enhance Food Safety. Microorganisms 2020, 8, 724. [Google Scholar] [CrossRef] [PubMed]
- Lasagabaster, A.; Jiménez, E.; Lehnherr, T.; Miranda-Cadena, K.; Lehnherr, H. Bacteriophage Biocontrol to Fight Listeria Outbreaks in Seafood. Food Chem. Toxicol. 2020, 145, 111682. [Google Scholar] [CrossRef] [PubMed]
- Loessner, M.J. Bacteriophage Endolysins—Current State of Research and Applications. Curr. Opin. Microbiol. 2005, 8, 480–487. [Google Scholar] [CrossRef]
- Ahmadi, H.; Anany, H.; Walkling-Ribeiro, M.; Griffiths, M. Biocontrol of Shigella Flexneri in Ground Beef and Vibrio Cholerae in Seafood with Bacteriophage-Assisted High Hydrostatic Pressure (HHP) Treatment. Food Bioprocess Technol. 2015, 8, 1160–1167. [Google Scholar] [CrossRef]
- Zulkarneev, E.; Aleshkin, A.; Kiseleva, I.; Rubalsky, E.; Rubalsky, O. Bacteriophage Cocktail Effectively Prolonging the Shelf-Life of Chilled Fish. Bull. Exp. Biol. Med. 2019, 167, 818–822. [Google Scholar] [CrossRef]
- Galarce, N.E.; Bravo, J.L.; Robeson, J.P.; Borie, C.F. Bacteriophage Cocktail Reduces Salmonella Enterica Serovar Enteritidis Counts in Raw and Smoked Salmon Tissues. Rev. Argent. Microbiol. 2014, 46, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Dewanggana, M.N.; Evangeline, C.; Ketty, M.D.; Waturangi, D.E.; Magdalena, S. Isolation, Characterization, Molecular Analysis and Application of Bacteriophage DW-EC to Control Enterotoxigenic Escherichia coli on Various Foods. Sci. Rep. 2022, 12, 495. [Google Scholar] [CrossRef] [PubMed]
- Miguéis, S.; Saraiva, C.; Esteves, A. Efficacy of LISTEX P100 at Different Concentrations for Reduction of Listeria monocytogenes Inoculated in Sashimi. J. Food Prot. 2017, 80, 2094–2098. [Google Scholar] [CrossRef] [Green Version]
- Soni, K.A.; Nannapaneni, R.; Hagens, S. Reduction of Listeria monocytogenes on the Surface of Fresh Channel Catfish Fillets by Bacteriophage Listex P100. Foodborne Pathog. Dis. 2010, 7, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.; Bolocan, A.S.; Draper, L.A.; Ross, R.P.; Hill, C. The Effect of a Commercially Available Bacteriophage and Bacteriocin on Listeria monocytogenes in Coleslaw. Viruses 2019, 11, 977. [Google Scholar] [CrossRef] [Green Version]
- Kazanavičiūtė, V.; Misiūnas, A.; Gleba, Y.; Giritch, A.; Ražanskienė, A. Plant-Expressed Bacteriophage Lysins Control Pathogenic Strains of Clostridium Perfringens. Sci. Rep. 2018, 8, 10589. [Google Scholar] [CrossRef] [Green Version]
- Misiou, O.; van Nassau, T.J.; Lenz, C.A.; Vogel, R.F. The Preservation of Listeria-Critical Foods by a Combination of Endolysin and High Hydrostatic Pressure. Int. J. Food Microbiol. 2018, 266, 355–362. [Google Scholar] [CrossRef]
- Taylor, T.M.; Ravishankar, S.; Bhargava, K.; Juneja, V.K. Chemical Preservatives and Natural Food Antimicrobials. In Food Microbiology: Fundamentals and Frontiers; Wiley: Hoboken, NJ, USA, 2019; pp. 705–731. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1128/9781555819972.ch27 (accessed on 14 March 2022).
- Geng, P.; Zhang, L.; Shi, G.Y. Omics Analysis of Acetic Acid Tolerance in Saccharomyces Cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 94. [Google Scholar] [CrossRef]
- Nemes, D.; Kovács, R.; Nagy, F.; Tóth, Z.; Herczegh, P.; Borbás, A.; Kelemen, V.; Pfliegler, W.P.; Rebenku, I.; Hajdu, P.B.; et al. Comparative Biocompatibility and Antimicrobial Studies of Sorbic Acid Derivates. Eur. J. Pharm. Sci. 2020, 143, 105162. [Google Scholar] [CrossRef]
- Baptista, R.C.; Horita, C.N.; Sant’Ana, A.S. Natural Products with Preservative Properties for Enhancing the Microbiological Safety and Extending the Shelf-Life of Seafood: A Review. Food Res. Int. 2020, 127, 108762. [Google Scholar] [CrossRef]
- Smyth, C.; Brunton, N.P.; Fogarty, C.; Bolton, D.J. The Effect of Organic Acid, Trisodium Phosphate and Essential Oil Component Immersion Treatments on the Microbiology of Cod (Gadus morhua) during Chilled Storage. Foods 2018, 7, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, C.; Ravishankar, C.; Kumar, K.A.; Gopal, T.S. Quality and Shelf Life of Sodium-Acetate-Treated Seer Fish (Scomberomorus commerson) Steaks Packed in EVOH Pouches during Chilled Storage. J. Packag. Technol. Res. 2019, 3, 109–116. [Google Scholar] [CrossRef]
- Saeid Asr, E.; Naghibi, S.; Mojaddar Langroodi, A.; Moghaddas Kia, E.; Meshkini, S.; Ehsani, A. Impact of Carboxymethyl Cellulose Coating Incorporated with Rosemary Essential Oil And Sodium Acetate on the Quality and Shelf Life of Rainbow Trout Fillet. J. Aquat. Food Prod. Technol. 2021, 30, 16–30. [Google Scholar] [CrossRef]
- Ahmed, N.S.; Dora, K.; Chowdhury, S.; Sarkar, S.; Mishra, R. Effect of Chitosan and Acetic Acid on the Shelf Life of Sea Bass Fillets Stored at Refrigerated Temperature. J. Appl. Nat. Sci. 2017, 9, 2175–2181. [Google Scholar] [CrossRef] [Green Version]
- Gardner, W.H. Acidulants in Food Processing. Handb. Food Addit. 1972, 2, 225pp. [Google Scholar]
- Sanyal, S.; Chatterjee, A. Antimicrobial Action and Biopreservation by Autochthonous Lactic Acid Bacteria of Fish. AZB 2021, 9, 37–44. [Google Scholar] [CrossRef]
- Aymerich, T.; Rodríguez, M.; Garriga, M.; Bover-Cid, S. Assessment of the Bioprotective Potential of Lactic Acid Bacteria against Listeria Monocytogenes on Vacuum-Packed Cold-Smoked Salmon Stored at 8 °C. Food Microbiol. 2019, 83, 64–70. [Google Scholar] [CrossRef]
- Leroi, F.; Cornet, J.; Chevalier, F.; Cardinal, M.; Coeuret, G.; Chaillou, S.; Joffraud, J.-J. Selection of Bioprotective Cultures for Preventing Cold-Smoked Salmon Spoilage. Int. J. Food Microbiol. 2015, 213, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yang, X.; Shi, G.; Chang, J.; Liu, Z.; Zeng, M. Cooperation of Lactic Acid Bacteria Regulated by the AI-2/LuxS System Involve in the Biopreservation of Refrigerated Shrimp. Food Res. Int. 2019, 120, 679–687. [Google Scholar] [CrossRef]
- Saraoui, T.; Cornet, J.; Guillouet, E.; Pilet, M.F.; Chevalier, F.; Joffraud, J.-J.; Leroi, F. Improving Simultaneously the Quality and Safety of Cooked and Peeled Shrimp Using a Cocktail of Bioprotective Lactic Acid Bacteria. Int. J. Food Microbiol. 2017, 241, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Sala, B.; Herranz, C.; Díaz-Freitas, B.; Hernández, P.E.; Sala, A.; Cintas, L.M. Strategies to Increase the Hygienic and Economic Value of Fresh Fish: Biopreservation Using Lactic Acid Bacteria of Marine Origin. Int. J. Food Microbiol. 2016, 223, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ladha, G.; Jeevaratnam, K. Characterization of Purified Antimicrobial Peptide Produced by Pediococcus pentosaceus LJR1, and Its Application in Preservation of White Leg Shrimp. World J. Microbiol. Biotechnol. 2020, 36, 72. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Du, J.; Jie, Y.; Zhang, B.; Bai, F.; Zhao, H.; Li, J. Purification and Antibacterial Mechanism of Fish-Borne Bacteriocin and Its Application in Shrimp (Penaeus vannamei) for Inhibiting Vibrio parahaemolyticus. World J. Microbiol. Biotechnol. 2017, 33, 156. [Google Scholar] [CrossRef] [PubMed]
- Bolívar, A.; Tarlak, F.; Costa, J.C.C.P.; Cejudo-Gómez, M.; Bover-Cid, S.; Zurera, G.; Pérez-Rodríguez, F. A New Expanded Modelling Approach for Investigating the Bioprotective Capacity of Latilactobacillus sakei CTC494 against Listeria monocytogenes in Ready-to-Eat Fish Products. Food Res. Int. 2021, 147, 110545. [Google Scholar] [CrossRef] [PubMed]
- Boulares, M.; Ben Moussa, O.; Mankai, M.; Sadok, S.; Hassouna, M. Effects of Lactic Acid Bacteria and Citrus Essential Oil on the Quality of Vacuum-Packed Sea Bass (Dicentrarchus labrax) Fillets during Refrigerated Storage. J. Aquat. Food Prod. Technol. 2018, 27, 698–711. [Google Scholar] [CrossRef]
- Nath, S.; Chowdhury, S.; Sarkar, S.; Dora, K. Biopreservation of Horse Mackerel Fillet Using Lactobacillus plantarum (ATCC 8014) and Lactobacillus sakei (ATCC 15521). Environ. Ecol. 2017, 35, 3126–3130. [Google Scholar]
- Vieira, G.R.A.D.S.; Soares, M.; Ramírez, N.C.B.; Schleder, D.D.; da Silva, B.C.; Mouriño, J.L.P.; Andreatta, E.R.; do Nascimento Vieira, F. Lactic Acid Bacteria Used as Preservative in Fresh Feed for Marine Shrimp Maturation. Pesqui. Agropecuária Bras. 2016, 51, 1799–1805. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rathod, N.B.; Nirmal, N.P.; Pagarkar, A.; Özogul, F.; Rocha, J.M. Antimicrobial Impacts of Microbial Metabolites on the Preservation of Fish and Fishery Products: A Review with Current Knowledge. Microorganisms 2022, 10, 773. https://doi.org/10.3390/microorganisms10040773
Rathod NB, Nirmal NP, Pagarkar A, Özogul F, Rocha JM. Antimicrobial Impacts of Microbial Metabolites on the Preservation of Fish and Fishery Products: A Review with Current Knowledge. Microorganisms. 2022; 10(4):773. https://doi.org/10.3390/microorganisms10040773
Chicago/Turabian StyleRathod, Nikheel Bhojraj, Nilesh Prakash Nirmal, Asif Pagarkar, Fatih Özogul, and João Miguel Rocha. 2022. "Antimicrobial Impacts of Microbial Metabolites on the Preservation of Fish and Fishery Products: A Review with Current Knowledge" Microorganisms 10, no. 4: 773. https://doi.org/10.3390/microorganisms10040773
APA StyleRathod, N. B., Nirmal, N. P., Pagarkar, A., Özogul, F., & Rocha, J. M. (2022). Antimicrobial Impacts of Microbial Metabolites on the Preservation of Fish and Fishery Products: A Review with Current Knowledge. Microorganisms, 10(4), 773. https://doi.org/10.3390/microorganisms10040773