
Phylogeographic Patterns of Haemoproteid Assemblages of Selected Avian Hosts: Ecological and Evolutionary Implications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area, Field Methods and Study Species
2.2. DNA Extraction
2.3. PCR Analyses
2.4. Phylogenetic and Haplotype Network Analyses
2.5. Statistical Analyses
3. Results
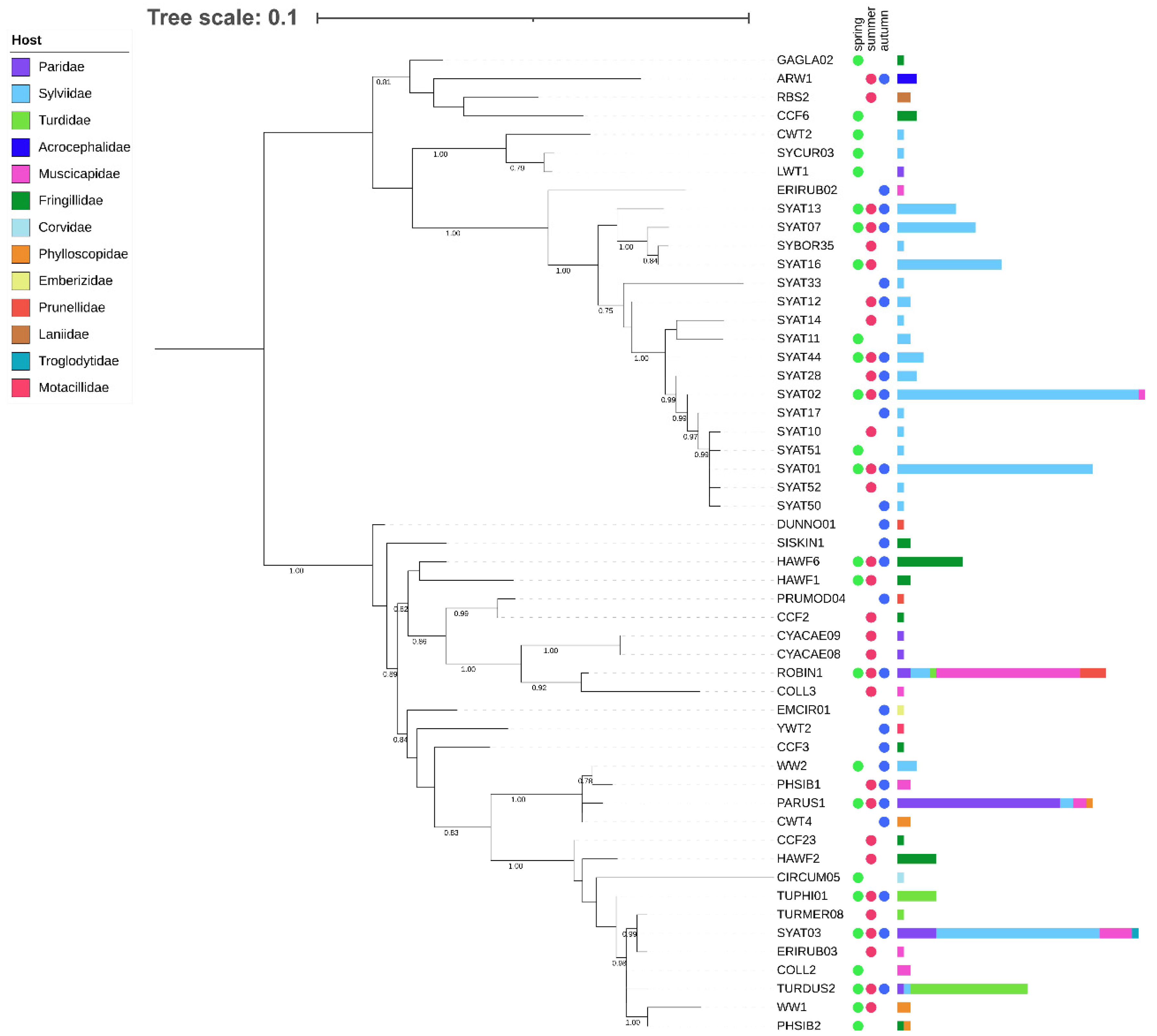
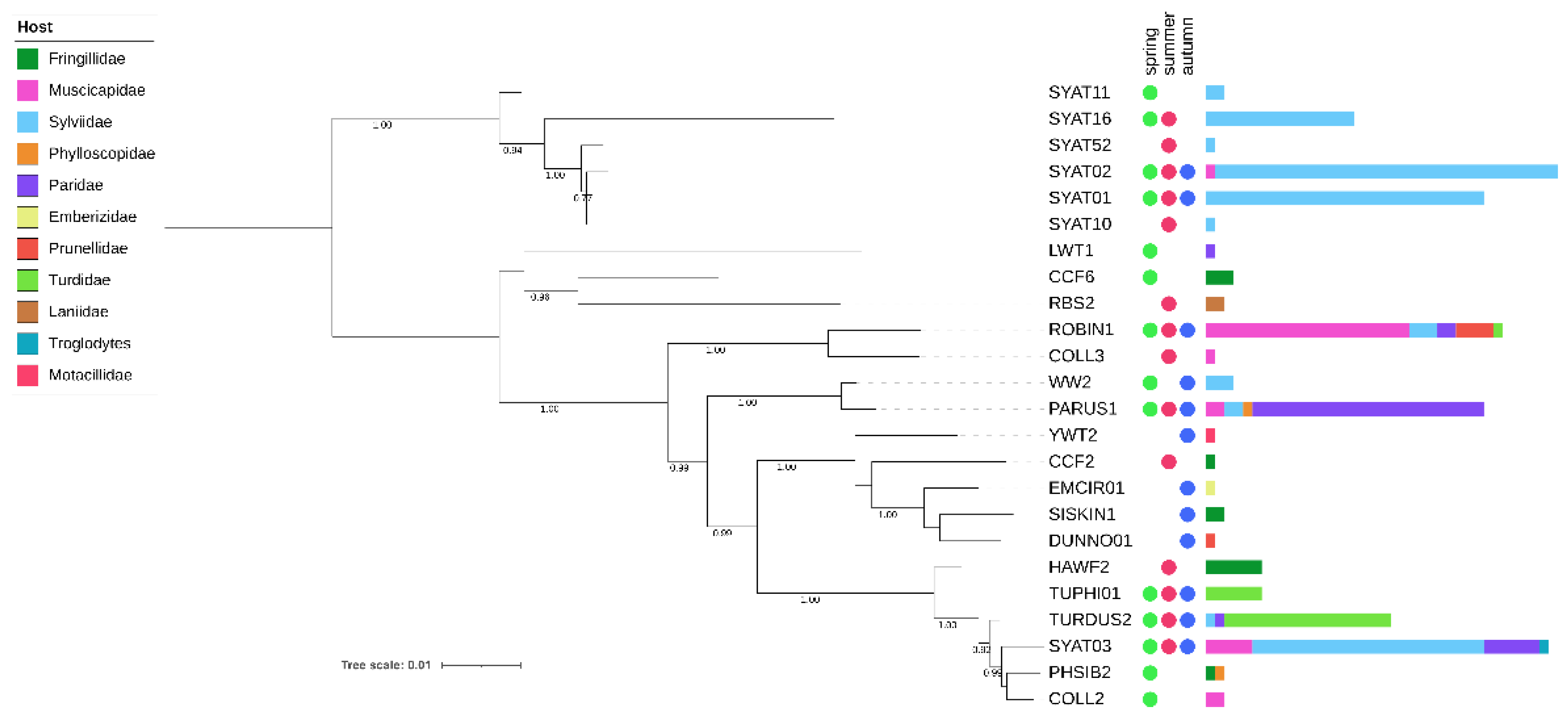
3.1. Phylogenetic Analysis
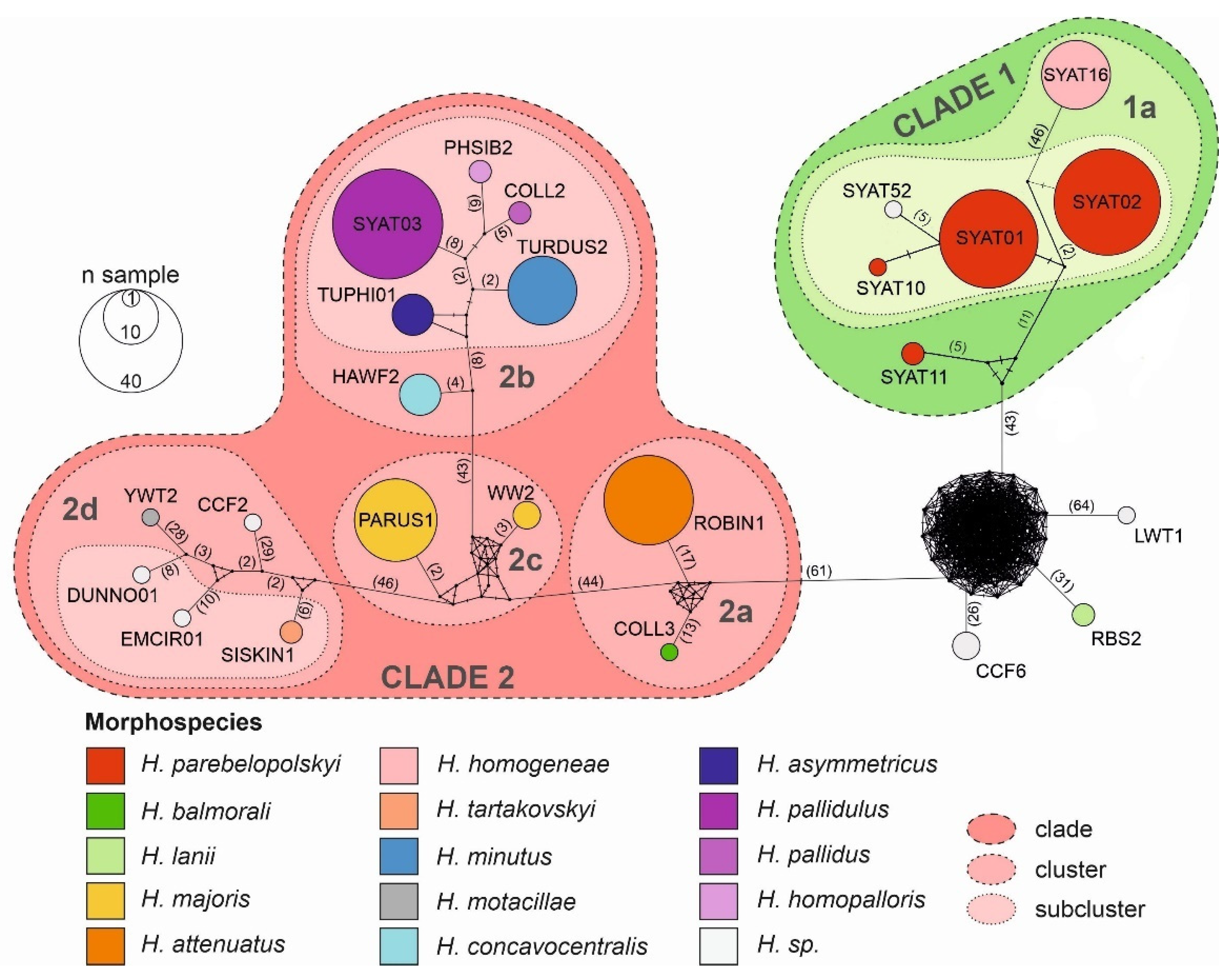
3.2. Haplotype Network Analysis
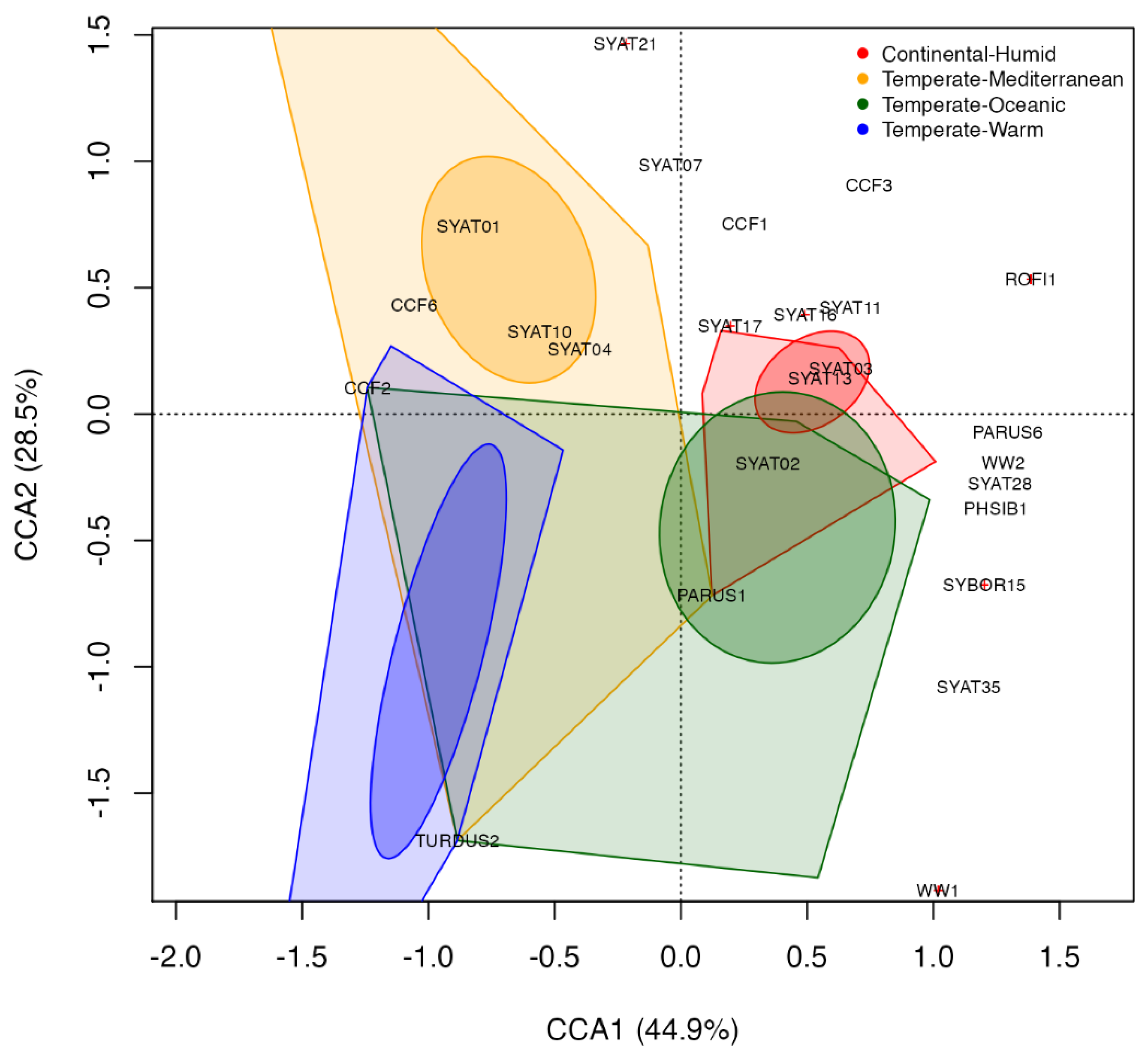
3.3. Ordination Analyses
4. Discussion
Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global Trends in Emerging Infectious Diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.C.; Hawley, D.M.; Huyvaert, K.P. Infectious Disease Ecology of Wild Birds; Oxford University Press: Oxford, NY, USA, 2021; ISBN 978-0-19-874624-9. [Google Scholar]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Ostman, O.; Hansson, B.; Westerdahl, H.; Pinheiro, R.T. Host Specificity in Avian Blood Parasites: A Study of Plasmodium and Haemoproteus Mitochondrial DNA Amplified from Birds. Proc. Biol. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricklefs, R.E.; Fallon, S.M. Diversification and Host Switching in Avian Malaria Parasites. Proc. R. Soc. B Biol. Sci. 2002, 269, 885–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, V.A.; Huang, X.; Westerdahl, H.; Jönsson, J.; Hasselquist, D.; Neto, J.M.; Nilsson, J.-A.; Nilsson, J.; Hegemann, A.; Hellgren, O. Explaining Prevalence, Diversity and Host Specificity in a Community of Avian Haemosporidian Parasites. Oikos 2020, 129, 1314–1329. [Google Scholar] [CrossRef]
- Waldenström, J.; Bensch, S.; Kiboi, S.; Hasselquist, D.; Ottosson, U. Cross-Species Infection of Blood Parasites between Resident and Migratory Songbirds in Africa. Mol. Ecol. 2002, 11, 1545–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Tris, J.; Hellgren, O.; Križanauskienė, A.; Waldenström, J.; Secondi, J.; Bonneaud, C.; Fjeldså, J.; Hasselquist, D.; Bensch, S. Within-Host Speciation of Malaria Parasites. PLoS ONE 2007, 2, e235. [Google Scholar] [CrossRef]
- Beadell, J.S.; Covas, R.; Gebhard, C.; Ishtiaq, F.; Melo, M.; Schmidt, B.K.; Perkins, S.L.; Graves, G.R.; Fleischer, R.C. Host Associations and Evolutionary Relationships of Avian Blood Parasites from West Africa. Int. J. Parasitol. 2009, 39, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.N. Concepts of Coevolution. Trends Ecol. Evol. 1989, 4, 179–183. [Google Scholar] [CrossRef]
- Pulgarín, P.; Gómez, C.; Bayly, N.J.; Bensch, S.; FitzGerald, A.M.; Starkloff, N.; Kirchman, J.J.; González-Prieto, A.M.; Hobson, K.A.; Ungvari-Martin, J.; et al. Migratory Birds as Vehicles for Parasite Dispersal? Infection by Avian Haemosporidians over the Year and throughout the Range of a Long-Distance Migrant. J. Biogeogr. 2019, 46, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Fallon, S.M.; Bermingham, E.; Ricklefs, R.E. Host Specialization and Geographic Localization of Avian Malaria Parasites: A Regional Analysis in the Lesser Antilles. Am. Nat. 2005, 165, 466–480. [Google Scholar] [CrossRef]
- Cornuault, J.; Bataillard, A.; Warren, B.H.; Lootvoet, A.; Mirleau, P.; Duval, T.; Milá, B.; Thébaud, C.; Heeb, P. The Role of Immigration and In-Situ Radiation in Explaining Blood Parasite Assemblages in an Island Bird Clade. Mol. Ecol. 2012, 21, 1438–1452. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E.; Ellis, V.A.; Medeiros, M.C.; Svensson-Coelho, M. Duration of Embryo Development and the Prevalence of Haematozoan Blood Parasites in Birds. Auk 2018, 135, 276–283. [Google Scholar] [CrossRef]
- Ruhs, E.C.; Martin, L.B.; Downs, C.J. The Impacts of Body Mass on Immune Cell Concentrations in Birds. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200655. [Google Scholar] [CrossRef] [PubMed]
- Lutz, H.L.; Hochachka, W.M.; Engel, J.I.; Bell, J.A.; Tkach, V.V.; Bates, J.M.; Hackett, S.J.; Weckstein, J.D. Parasite Prevalence Corresponds to Host Life History in a Diverse Assemblage of Afrotropical Birds and Haemosporidian Parasites. PLoS ONE 2015, 10, e0128851. [Google Scholar] [CrossRef] [Green Version]
- Minias, P. Evolution of Heterophil/Lymphocyte Ratios in Response to Ecological and Life-history Traits: A Comparative Analysis across the Avian Tree of Life. J. Anim. Ecol. 2019, 88, 554–565. [Google Scholar] [CrossRef]
- Fecchio, A.; Clark, N.J.; Bell, J.A.; Skeen, H.R.; Lutz, H.L.; De La Torre, G.M.; Vaughan, J.A.; Tkach, V.V.; Schunck, F.; Ferreira, F.C. Global Drivers of Avian Haemosporidian Infections Vary across Zoogeographical Regions. Glob. Ecol. Biogeogr. 2021, 30, 2393–2406. [Google Scholar] [CrossRef]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- de Angeli Dutra, D.; Fecchio, A.; Martins Braga, É.; Poulin, R. Migratory Birds Have Higher Prevalence and Richness of Avian Haemosporidian Parasites than Residents. Int. J. Parasitol. 2021, 51, 877–882. [Google Scholar] [CrossRef]
- Kelly, T.R.; MacGillivray, H.L.; Sarquis-Adamson, Y.; Watson, M.J.; Hobson, K.A.; MacDougall-Shackleton, E.A. Seasonal Migration Distance Varies with Natal Dispersal and Predicts Parasitic Infection in Song Sparrows. Behav. Ecol. Sociobiol. 2016, 70, 1857–1866. [Google Scholar] [CrossRef]
- Cosgrove, C.L.; Wood, M.J.; Day, K.P.; Sheldon, B.C. Seasonal Variation in Plasmodium Prevalence in a Population of Blue Tits Cyanistes Caeruleus. J. Anim. Ecol. 2008, 77, 540–548. [Google Scholar] [CrossRef]
- Arizaga, J.; Esparza, X.; Barba, E. Haemosporidians in Migratory Blackcaps (Sylvia Atricapilla), a Comparison between Autumn and Spring Periods of Passage; Universidad de Murcia, Servicio de Publicaciones: Murcia, Spain, 2010. [Google Scholar]
- Križanauskienė, A.; Perez-Tris, J.; Palinauskas, V.; Hellgren, O.; Bensch, S.; Valkiūnas, G. Molecular Phylogenetic and Morphological Analysis of Haemosporidian Parasites (Haemosporida) in a Naturally Infected European Songbird, the Blackcap Sylvia Atricapilla, with Description of Haemoproteus Pallidulus Sp Nov. Parasitology 2010, 137, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Palinauskas, V.; Iezhova, T.A.; Križanauskienė, A.; Markovets, M.Y.; Bensch, S.; Valkiūnas, G. Molecular Characterization and Distribution of Haemoproteus Minutus (Haemosporida, Haemoproteidae): A Pathogenic Avian Parasite. Parasitol. Int. 2013, 62, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Podmokła, E.; Dubiec, A.; Arct, A.; Drobniak, S.M.; Gustafsson, L.; Cichoń, M. Malaria Infection Status Predicts Extra-Pair Paternity in the Blue Tit. J. Avian Biol. 2015, 46, 303–306. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, E.; Taubert, H.; Hellgren, O.; Huang, X.; Palinauskas, V.; Markovets, M.Y.; Valkiūnas, G.; Bensch, S. Multiple Cryptic Species of Sympatric Generalists within the Avian Blood Parasite Haemoproteus Majoris. J. Evolut. Biol. 2016, 29, 1812–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Chagas, C.R.F.; Bernotienė, R.; Himmel, T.; Harl, J.; Weissenbock, H.; Iezhova, T.A. Molecular Characterization of Six Widespread Avian Haemoproteids, with Description of Three New Haemoproteus Species. Acta Trop. 2019, 197, 105051. [Google Scholar] [CrossRef] [PubMed]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Bakonyi, T.; Weissenböck, H. Geographic and Host Distribution of Haemosporidian Parasite Lineages from Birds of the Family Turdidae. Malar. J. 2020, 19, 335. [Google Scholar] [CrossRef]
- Pacheco, M.A.; Matta, N.E.; Valkiūnas, G.; Parker, P.G.; Mello, B.; Stanley, C.E.; Lentino, M.; Garcia-Amado, M.A.; Cranfield, M.; Pond, S.L.K.; et al. Mode and Rate of Evolution of Haemosporidian Mitochondrial Genomes: Timing the Radiation of Avian Parasites. Mol. Biol. Evol. 2018, 35, 383–403. [Google Scholar] [CrossRef] [Green Version]
- Bensch, S.; Hellgren, O. The Use of Molecular Methods in Studies of Avian Haemosporidians. In Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Santiago-Alarcon, D., Marzal, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 113–135. ISBN 978-3-030-51633-8. [Google Scholar]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Duc, M.; Iezhova, T.A. Description of Haemoproteus Asymmetricus n. Sp. (Haemoproteidae), with Remarks on Predictability of the DNA Haplotype Networks in Haemosporidian Parasite Taxonomy Research. Acta Trop. 2021, 218, 105905. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Palinauskas, V.; Schaefer, H.M. Diptera Vectors of Avian Haemosporidian Parasites: Untangling Parasite Life Cycles and Their Taxonomy. Biol. Rev. 2012, 87, 928–964. [Google Scholar] [CrossRef]
- Šujanová, A.; Špitalská, E.; Václav, R. Seasonal Dynamics and Diversity of Haemosporidians in a Natural Woodland Bird Community in Slovakia. Diversity 2021, 13, 439. [Google Scholar] [CrossRef]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A Public Database of Malaria Parasites and Related Haemosporidians in Avian Hosts Based on Mitochondrial Cytochrome b Lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Mtierová, Z.; Derdáková, M.; Chvostáč, M.; Didyk, Y.M.; Mangová, B.; Rusňáková Tarageľová, V.; Selyemová, D.; Šujanová, A.; Václav, R. Local Population Structure and Seasonal Variability of Borrelia Garinii Genotypes in Ixodes Ricinus Ticks, Slovakia. Int. J. Environ. Res. Public Health 2020, 17, 3607. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, O.; Waldenström, J.; Bensch, S. A New PCR Assay for Simultaneous Studies of Leucocytozoon, Plasmodium, and Haemoproteus from Avian Blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, M.A.; Cepeda, A.S.; Bernotienė, R.; Lotta, I.A.; Matta, N.E.; Valkiūnas, G.; Escalante, A.A. Primers Targeting Mitochondrial Genes of Avian Haemosporidians: PCR Detection and Differential DNA Amplification of Parasites Belonging to Different Genera. Int. J. Parasitol. 2018, 48, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE team. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucl. Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast Selection of Best-Fit Models of Protein Evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Maximum Likelihood Phylogenetic Estimation from DNA Sequences with Variable Rates over Sites: Approximate Methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Soubrier, J.; Steel, M.; Lee, M.S.Y.; Der Sarkissian, C.; Guindon, S.; Ho, S.Y.W.; Cooper, A. The Influence of Rate Heterogeneity among Sites on the Time Dependence of Molecular Rates. Mol. Biol. Evol. 2012, 29, 3345–3358. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.-F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-Based Approximation Schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.N.; de Moraes Russo, C.A. An Empirical Test of the Midpoint Rooting Method. Biol. J. Linn. Soc. 2007, 92, 669–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v4: Recent Updates and New Developments. Nucl. Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. Popart: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Clark, N.J. Phylogenetic Uniqueness, Not Latitude, Explains the Diversity of Avian Blood Parasite Communities Worldwide. Glob. Ecol. Biogeogr. 2018, 27, 744–755. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and Future Köppen-Geiger Climate Classification Maps at 1-Km Resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; ISBN 978-1-4419-7975-9. [Google Scholar]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward Selection of Explanatory Variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; The R Foundation: Vienna, Austria, 2020. [Google Scholar]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Križanauskienė, A.; Hellgren, O.; Kosarev, V.; Sokolov, L.; Bensch, S.; Valkiūnas, G. Variation in Host Specificity between Species of Avian Hemosporidian Parasites: Evidence from Parasite Morphology and Cytochrome b Gene Sequences. J. Parasitol. 2006, 92, 1319–1324. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Mettler, R.; Segelbacher, G.; Schaefer, H.M. Haemosporidian Parasitism in the Blackcap Sylvia Atricapilla in Relation to Spring Arrival and Body Condition. J. Avian Biol. 2013, 44, 521–530. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, A.; de la Hera, I.; Bensch, S.; Pérez-Tris, J. Evolution of Seasonal Transmission Patterns in Avian Blood-Borne Parasites. Int. J. Parasitol. 2015, 45, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tris, J.; Bensch, S. Dispersal Increases Local Transmission of Avian Malarial Parasites. Ecol. Lett. 2005, 8, 838–845. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Križanauskienė, A.; Iezhova, T.A.; Hellgren, O.; Bensch, S. Molecular Phylogenetic Analysis of Circumnuclear Hemoproteids (Haemosporida: Haemoproteidae) of Sylviid Birds, with a Description of Haemoproteus Parabelopolskyi Sp. Nov. J. Parasitol. 2007, 93, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Bukauskaitė, D.; Ilgūnas, M.; Iezhova, T.; Valkiūnas, G. A New Blood Parasite of Leaf Warblers: Molecular Characterization, Phylogenetic Relationships, Description and Identification of Vectors. Parasites Vectors 2018, 11, 538. [Google Scholar] [CrossRef]
- Bobeva, A.; Zehtindjiev, P.; Bensch, S.; Radrova, J. A Survey of Biting Midges of the Genus Culicoides Latreille, 1809 (Diptera: Ceratopogonidae) in NE Bulgaria, with Respect to Transmission of Avian Haemosporidians. Acta Parasitol. 2013, 58, 585–591. [Google Scholar] [CrossRef] [Green Version]
- Synek, P.; Munclinger, P.; Albrecht, T.; Votýpka, J. Avian Haemosporidians in Haematophagous Insects in the Czech Republic. Parasitol. Res. 2013, 112, 839–845. [Google Scholar] [CrossRef]
- Žiegytė, R.; Platonova, E.; Kinderis, E.; Mukhin, A.; Palinauskas, V.; Bernotienė, R. Culicoides Biting Midges Involved in Transmission of Haemoproteids. Parasites Vectors 2021, 14, 27. [Google Scholar] [CrossRef]
- Ilgūnas, M.; Romeiro Fernandes Chagas, C.; Bukauskaitė, D.; Bernotienė, R.; Iezhova, T.; Valkiūnas, G. The Life-Cycle of the Avian Haemosporidian Parasite Haemoproteus Majoris, with Emphasis on the Exoerythrocytic and Sporogonic Development. Parasites Vectors 2019, 12, 516. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Jönsson, J.; Bensch, S. Persistence of Avian Haemosporidians in the Wild: A Case Study to Illustrate Seasonal Infection Patterns in Relation to Host Life Stages. Int. J. Parasitol. 2020, 50, 611–619. [Google Scholar] [CrossRef]
- Dobzhansky, T. Evolution in the Tropics. Am. Sci. 1950, 38, 209–221. [Google Scholar]
- Fecchio, A.; Wells, K.; Bell, J.A.; Tkach, V.V.; Lutz, H.L.; Weckstein, J.D.; Clegg, S.M.; Clark, N.J. Climate Variation Influences Host Specificity in Avian Malaria Parasites. Ecol. Lett. 2019, 22, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Alarcon, D.; Bloch, R.; Rolshausen, G.; Schaefer, H.M.; Segelbacher, G. Prevalence, Diversity, and Interaction Patterns of Avian Haemosporidians in a Four-Year Study of Blackcaps in a Migratory Divide. Parasitology 2011, 138, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Drovetski, S.V.; Aghayan, S.A.; Mata, V.A.; Lopes, R.J.; Mode, N.A.; Harvey, J.A.; Voelker, G. Does the Niche Breadth or Trade-off Hypothesis Explain the Abundance–Occupancy Relationship in Avian Haemosporidia? Mol. Ecol. 2014, 23, 3322–3329. [Google Scholar] [CrossRef] [PubMed]
- Van Doren, B.M.; Conway, G.J.; Phillips, R.J.; Evans, G.C.; Roberts, G.C.M.; Liedvogel, M.; Sheldon, B.C. Human Activity Shapes the Wintering Ecology of a Migratory Bird. Glob. Change Biol. 2021, 27, 2715–2727. [Google Scholar] [CrossRef]
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A. The Transmission of Haemoproteus Belopolskyi (Haemosporida: Haemoproteidae) of Blackcap by Culicoides Impunctatus (Diptera: Ceratopogonidae). J. Parasitol. 2004, 90, 196–198. [Google Scholar] [CrossRef]
- Žiegytė, R.; Markovets, M.Y.; Bernotienė, R.; Mukhin, A.; Iezhova, T.A.; Valkiūnas, G.; Palinauskas, V. The Widespread Biting Midge Culicoides Impunctatus (Ceratopogonidae) Is Susceptible to Infection with Numerous Haemoproteus (Haemoproteidae) Species. Parasites Vectors 2017, 10, 397. [Google Scholar] [CrossRef]
- Bukauskaitė, D.; Iezhova, T.A.; Ilgūnas, M.; Valkiūnas, G. High Susceptibility of the Laboratory-Reared Biting Midges Culicoides Nubeculosus to Haemoproteus Infections, with Review on Culicoides Species That Transmit Avian Haemoproteids. Parasitology 2019, 146, 333–341. [Google Scholar] [CrossRef]
- Bernotienė, R.; Valkiūnas, G. PCR Detection of Malaria Parasites and Related Haemosporidians: The Sensitive Methodology in Determining Bird-Biting Insects. Malar. J. 2016, 15, 283. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Lara, C.; Duc, M.; Ilgūnas, M.; Valkiūnas, G. Massive Infection of Lungs with Exo-Erythrocytic Meronts in European Robin Erithacus Rubecula during Natural Haemoproteus Attenuatus Haemoproteosis. Animals 2021, 11, 3273. [Google Scholar] [CrossRef]
- Olias, P.; Wegelin, M.; Zenker, W.; Freter, S.; Gruber, A.D.; Klopfleisch, R. Avian Malaria Deaths in Parrots, Europe. Emerg. Infect. Dis. 2011, 17, 950–952. [Google Scholar] [CrossRef]
- Himmel, T.; Harl, J.; Kübber-Heiss, A.; Konicek, C.; Fernández, N.; Juan-Sallés, C.; Ilgūnas, M.; Valkiūnas, G.; Weissenböck, H. Molecular Probes for the Identification of Avian Haemoproteus and Leucocytozoon Parasites in Tissue Sections by Chromogenic in Situ Hybridization. Parasites Vectors 2019, 12, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; et al. Haemoproteus Minutus Is Highly Virulent for Australasian and South American Parrots. Parasites Vectors 2019, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Brommer, J.E.; Lehikoinen, A.; Valkama, J. The Breeding Ranges of Central European and Arctic Bird Species Move Poleward. PLoS ONE 2012, 7, e43648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakanen, V.-M.; Ahonen, E.; Hohtola, E.; Rytkönen, S. Northward Expanding Resident Species Benefit from Warming Winters through Increased Foraging Rates and Predator Vigilance. Oecologia 2018, 188, 991–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, K.L.; Hatchwell, B.J.; Parnell, M.; Gaston, K.J. A Conceptual Framework for the Colonisation of Urban Areas: The Blackbird Turdus Merula as a Case Study. Biol. Rev. 2010, 85, 643–667. [Google Scholar] [CrossRef]
- Ciloglu, A.; Ergen, A.G.; Inci, A.; Dik, B.; Duzlu, O.; Onder, Z.; Yetismis, G.; Bensch, S.; Valkiūnas, G.; Yildirim, A. Prevalence and Genetic Diversity of Avian Haemosporidian Parasites at an Intersection Point of Bird Migration Routes: Sultan Marshes National Park, Turkey. Acta Trop. 2020, 210, 105465. [Google Scholar] [CrossRef]
- Pellegrino, I.; Ilahiane, L.; Boano, G.; Cucco, M.; Pavia, M.; Prestridge, H.L.; Voelker, G. Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity 2021, 13, 75. [Google Scholar] [CrossRef]
- Helbig, A.J. Inheritance of Migratory Direction in a Bird Species: A Cross-Breeding Experiment with SE- and SW-Migrating Blackcaps (Sylvia Atricapilla). Behav. Ecol. Sociobiol. 1991, 28, 9–12. [Google Scholar] [CrossRef]
- Cepák, J.J.; Klvaňa, P.; Škopek, J.; Schröpfer, L.; Jelínek, M.; Hořák, D.; Formánek, J.; Zárybnický, J. Czech and Slovak Bird Migration Atlas; Aventinum: Prague, Czech Republic, 2008. [Google Scholar]
- Aymí, R.; Gargallo, G.; Christie, D. Eurasian Blackcap (Sylvia Atricapilla), Version 1.0. In Birds of the World; The Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Bearhop, S.; Fiedler, W.; Furness, R.W.; Votier, S.C.; Waldron, S.; Newton, J.; Bowen, G.J.; Berthold, P.; Farnsworth, K. Assortative Mating as a Mechanism for Rapid Evolution of a Migratory Divide. Science 2005, 310, 502–504. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, T.; Thomas, G.H.; Hellgren, O.; Owens, I.P.F. Migratory Behavior of Birds Affects Their Coevolutionary Relationship with Blood Parasites. Evolution 2012, 66, 740–751. [Google Scholar] [CrossRef]
- Poulin, R.; de Angeli Dutra, D. Animal Migrations and Parasitism: Reciprocal Effects within a Unified Framework. Biol. Rev. 2021, 96, 1331–1348. [Google Scholar] [CrossRef] [PubMed]
- Cleaveland, S.; Haydon, D.T.; Taylor, L. Overviews of Pathogen Emergence: Which Pathogens Emerge, When and Why? In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission; Childs, J.E., Mackenzie, J.S., Richt, J.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 85–111. ISBN 978-3-540-70962-6. [Google Scholar]
- Hellgren, O.; Križanauskienė, A.; Valkiūnas, G.; Bensch, S. Diversity and phylogeny of mitochondrial cytochrome B lineages from six morphospecies of avian Haemoproteus (Haemosporida: Haemoproteidae). J. Parasitol. 2007, 93, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S. Genetic diversity of avian blood parasites in SE Europe: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Bulgaria. Acta Parasitol. 2010, 55, 201–209. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; MacGregor-Fors, I.; Kühnert, K.; Segelbacher, G.; Schaefer, H.M. Avian haemosporidian parasites in an urban forest and their relationship to bird size and abundance. Urban Ecosyst. 2016, 19, 331–346. [Google Scholar] [CrossRef]
- Bodawatta, K.H.; Synek, P.; Bos, N.; Garcia-del-Rey, E.; Koane, B.; Marki, P.Z.; Albrecht, T.; Lifjeld, J.; Poulsen, M.; Munclinger, P. Spatiotemporal patterns of avian host–parasite interactions in the face of biogeographical range expansions. Mol. Ecol. 2020, 29, 2431–2448. [Google Scholar] [CrossRef] [PubMed]
- Stanković, D.; Jönsson, J.; Raković, M. Diversity of avian blood parasites in wild passerines in Serbia with special reference to two new lineages. J. Ornithol. 2019, 160, 545–555. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Križanauskienė, A.; Palinauskas, V.; Sehgal, R.N.M.; Bensch, S. A comparative analysis of microscopy and PCR-based detection methods for blood parasites. J. Parasitol. 2008, 94, 1395–1401. [Google Scholar] [CrossRef]
- Nourani, L.; Aliabadian, M.; Dinparast Djadid, N.; Mirshamsi, O. Occurrence of Haemoproteus spp. (Haemosporida: Haemoproteidae) in New Host Records of Passerine Birds from the East of Iran. Iran. J. Parasitol. 2018, 13, 267–274. [Google Scholar]
- Knowles, S.C.L.; Wood, M.J.; Alves, R.; Wilkin, T.A.; Bensch, S.; Sheldon, B.C. Molecular epidemiology of malaria prevalence and parasitaemia in a wild bird population. Mol. Ecol. 2011, 20, 1062–1076. [Google Scholar] [CrossRef]
- Stjernman, M.; Råberg, L.; Nilsson, J.Å. Long-term effects of nestling condition on blood parasite resistance in blue tits (Cyanistes caeruleus). Can. J. Zool. 2008, 86, 937–946. [Google Scholar] [CrossRef]
- Dubiec, A.; Podmokła, E.; Zagalska-Neubauer, M.; Drobniak, S.M.; Arct, A.; Gustafsson, L.; Cichoń, M. Differential prevalence and diversity of haemosporidian parasites in two sympatric closely related non-migratory passerines. Parasitology 2016, 143, 1320–1329. [Google Scholar] [CrossRef] [PubMed]
- Schumm, Y.R.; Wecker, C.; Marek, C.; Wassmuth, M.; Bentele, A.; Willems, H.; Reiner, G.; Quillfeldt, P. Blood parasites in Passeriformes in central Germany: Prevalence and lineage diversity of Haemosporida (Haemoproteus, Plasmodium and Leucocytozoon) in six common songbirds. Peerj 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynton-Jenkins, J.G.; Bründl, A.C.; Cauchoix, M.; Lejeune, L.A.; Sallé, L.; Thiney, A.C.; Russell, A.F.; Chaine, A.S.; Bonneaud, C. Contrasting the seasonal and elevational prevalence of generalist avian haemosporidia in co-occurring host species. Ecol. Evol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.J.; Cosgrove, C.L.; Wilkin, T.A.; Knowles, S.C.L.; Day, K.P.; Sheldon, B.C. Within-population variation in prevalence and lineage distribution of avian malaria in blue tits, Cyanistes caeruleus. Mol. Ecol. 2007, 16, 3263–3273. [Google Scholar] [CrossRef]
- Wiersch, S.C.; Lubjuhn, T.; Maier, W.A.; Kampen, H. Haemosporidian infection in passerine birds from Lower Saxony. J. Ornithol. 2007, 148, 17–24. [Google Scholar] [CrossRef]
- Garcia-Longoria, L.; Marzal, A.; De Lope, F.; Garamszegi, L. Host-parasite interaction explains variation in the prevalence of avian haemosporidians at the community level. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Glaizot, O.; Fumagalli, L.; Iritano, K.; Lalubin, F.; van Rooyen, J.; Christe, P. High prevalence and lineage diversity of avian malaria in wild populations of great tits (parus major) and mosquitoes (culex pipiens). PLoS ONE 2012, 7, e34964. [Google Scholar] [CrossRef]
- van Rooyen, J.; Lalubin, F.; Glaizot, O.; Christe, P. Avian haemosporidian persistence and co-infection in great tits at the individual level. Malaria J. 2013, 12. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, A.; Ramírez, A.; Richardson, D.S.; Pérez-Tris, J. Evolution of parasite island syndromes without long-term host population isolation: Parasite dynamics in Macaronesian blackcaps Sylvia atricapilla. Glob. Ecol. Biogeogr. 2013, 22, 1272–1281. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Evans, E.; Carlson, J.S.; Martínez-Gómez, J.E.; Sehgal, R.N.M. Two new Haemoproteus species (Haemosporida: Haemoproteidae) from columbiform birds. J. Parasitol. 2013, 99, 513–521. [Google Scholar] [CrossRef]
- Martínez, J.; Vásquez, R.A.; Venegas, C.; Merino, S. Molecular characterisation of haemoparasites in forest birds from Robinson Crusoe Island: Is the Austral Thrush a potential threat to endemic birds? Bird Conserv. Int. 2015, 25, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Mata, V.A.; da Silva, L.P.; Lopes, R.J.; Drovetski, S.V. The Strait of Gibraltar Poses an Effective Barrier to Host-Specialised but Not to Host-Generalised Lineages of Avian Haemosporidia. Int. J. Parasitol. 2015, 45, 711–719. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šujanová, A.; Václav, R. Phylogeographic Patterns of Haemoproteid Assemblages of Selected Avian Hosts: Ecological and Evolutionary Implications. Microorganisms 2022, 10, 1019. https://doi.org/10.3390/microorganisms10051019
Šujanová A, Václav R. Phylogeographic Patterns of Haemoproteid Assemblages of Selected Avian Hosts: Ecological and Evolutionary Implications. Microorganisms. 2022; 10(5):1019. https://doi.org/10.3390/microorganisms10051019
Chicago/Turabian StyleŠujanová, Alžbeta, and Radovan Václav. 2022. "Phylogeographic Patterns of Haemoproteid Assemblages of Selected Avian Hosts: Ecological and Evolutionary Implications" Microorganisms 10, no. 5: 1019. https://doi.org/10.3390/microorganisms10051019
APA StyleŠujanová, A., & Václav, R. (2022). Phylogeographic Patterns of Haemoproteid Assemblages of Selected Avian Hosts: Ecological and Evolutionary Implications. Microorganisms, 10(5), 1019. https://doi.org/10.3390/microorganisms10051019