
Benthic Microbial Communities in a Seasonally Ice-Covered Sub-Arctic River (Pasvik River, Norway) Are Shaped by Site-Specific Environmental Conditions

 ,
,  , ,
, ,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
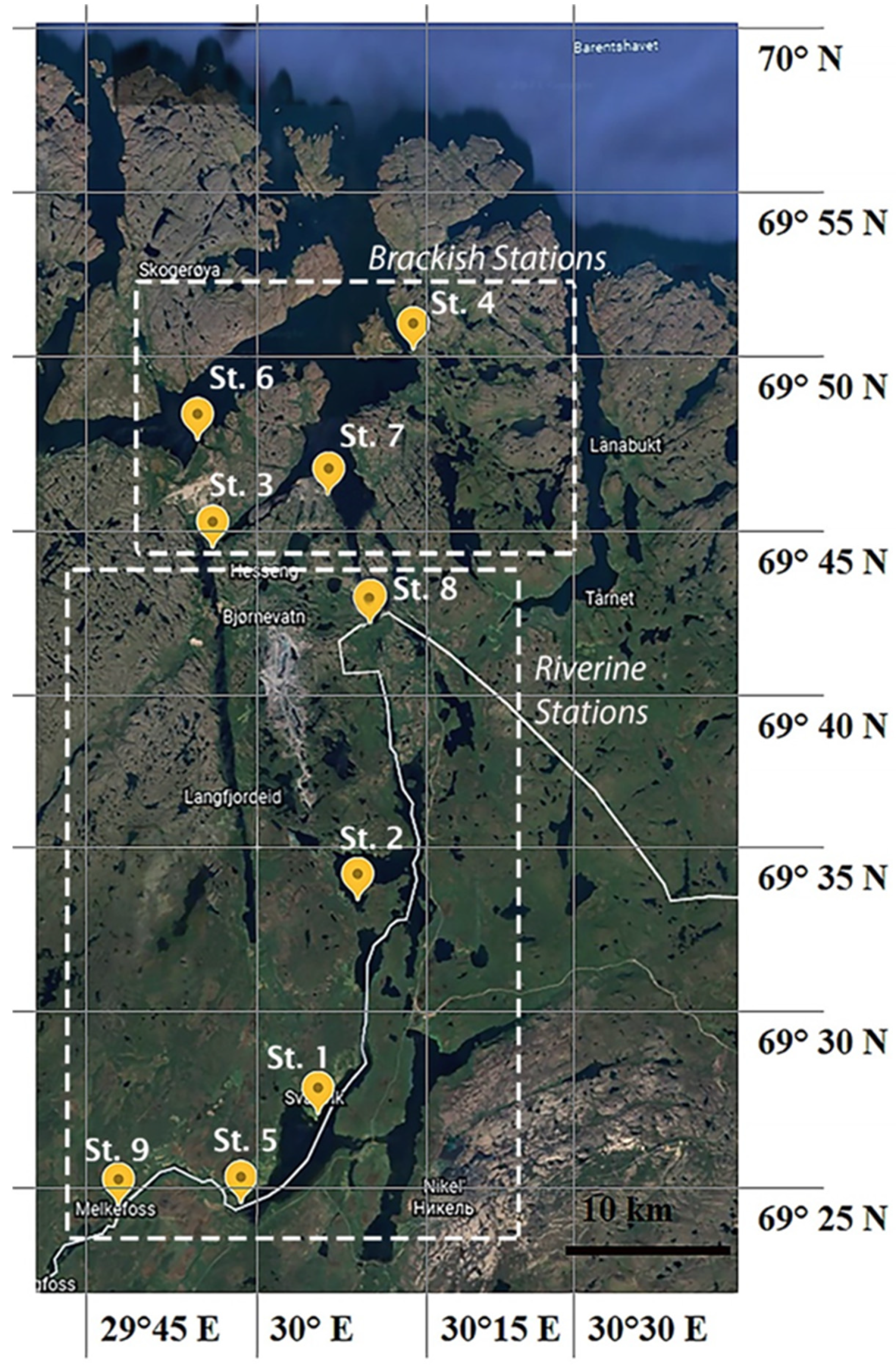
2.1. Sampling Area
2.2. Sample Collection
2.3. Particulate Material in Sediment Samples
2.4. Determination of Persistent Organic Pollutants (POPs)
2.5. Estimation of Microbial Cell Abundance, Biomass, and Enzymatic Activities
2.5.1. Total Cell Count by Flow Cytometry
2.5.2. Total Prokaryotic Biomass, Cell Volume, and Morphotypes by Image Analysis
2.5.3. Enzymatic Activities
2.6. Phylogenetic Composition of the Benthic Bacterial Community
DNA Extraction and PCR Amplification
2.7. Statistical Analyses
3. Results
3.1. Sediment Characteristics
3.2. Evaluation of Contamination Level
- -
- ACY, ACE, FLU, PHE, ANT, FLA, PYR, BaA, CRY, BbF, BkF, BaP, IPY, and BPE at 0.02, 0.05, 0.1, 0.2, 0.5, 1.0, 2.0, 5.0 ng mL−1;
- -
- All PCBs and OCPs at 0.01, 0.02, 0.05, 0.1, 0.2, 0.5, 1, and 2 ng mL−1.
3.3. Estimation of Microbial Abundances, Biomass, and Enzymatic Activities
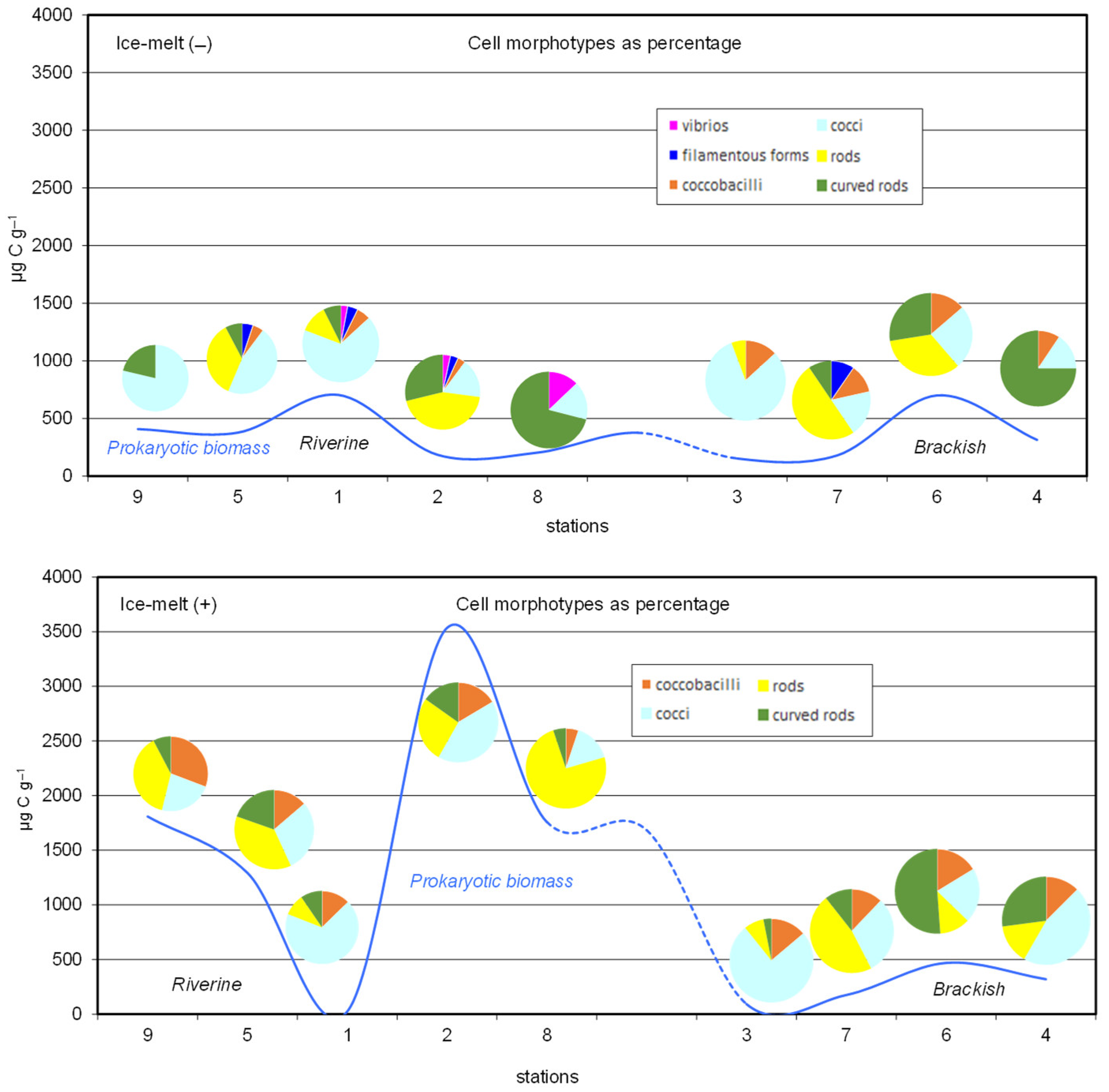
3.3.1. Total Cell Counts, Prokaryotic Biomass, and Cell Morphotypes
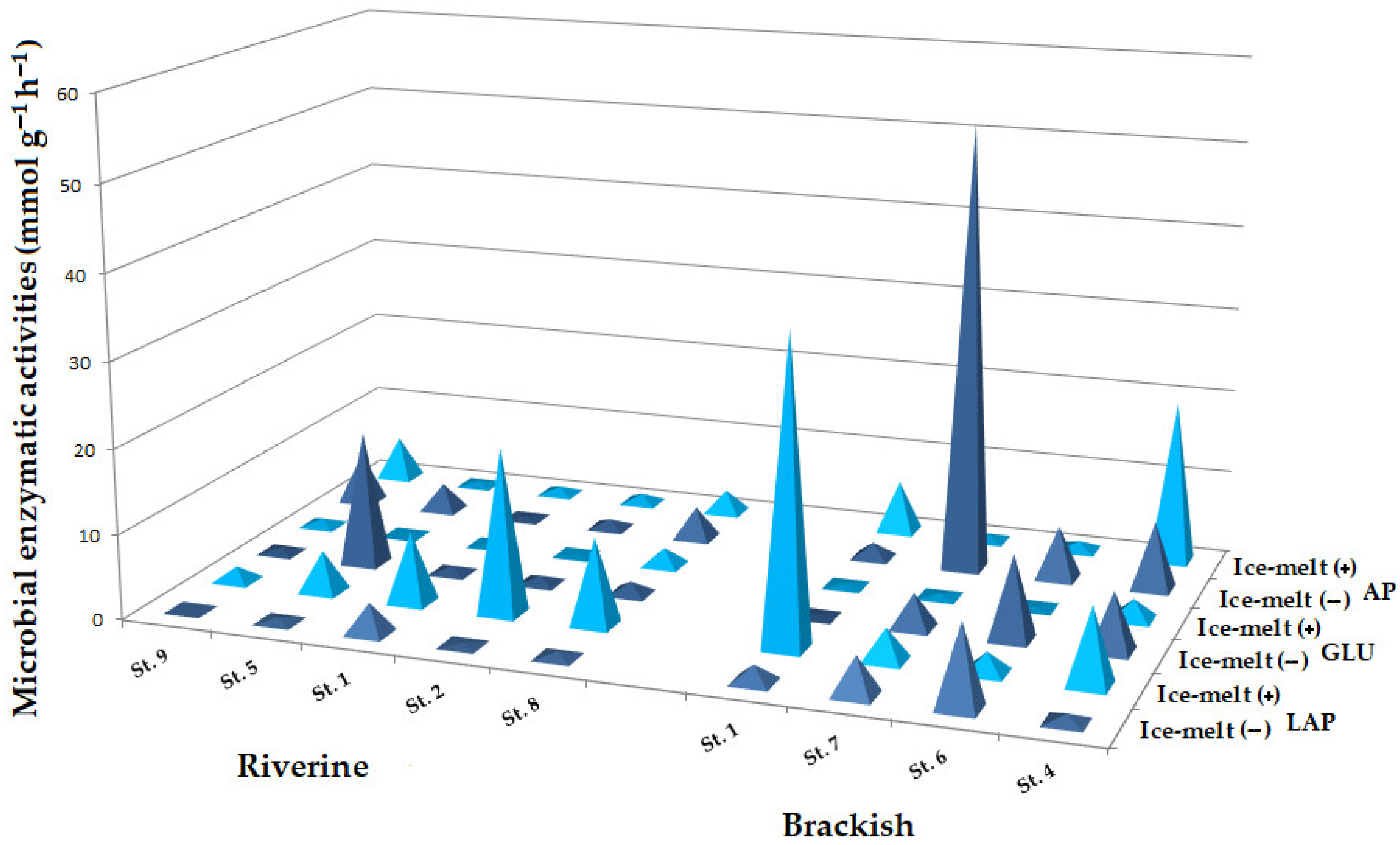
3.3.2. Microbial Enzymatic Activities
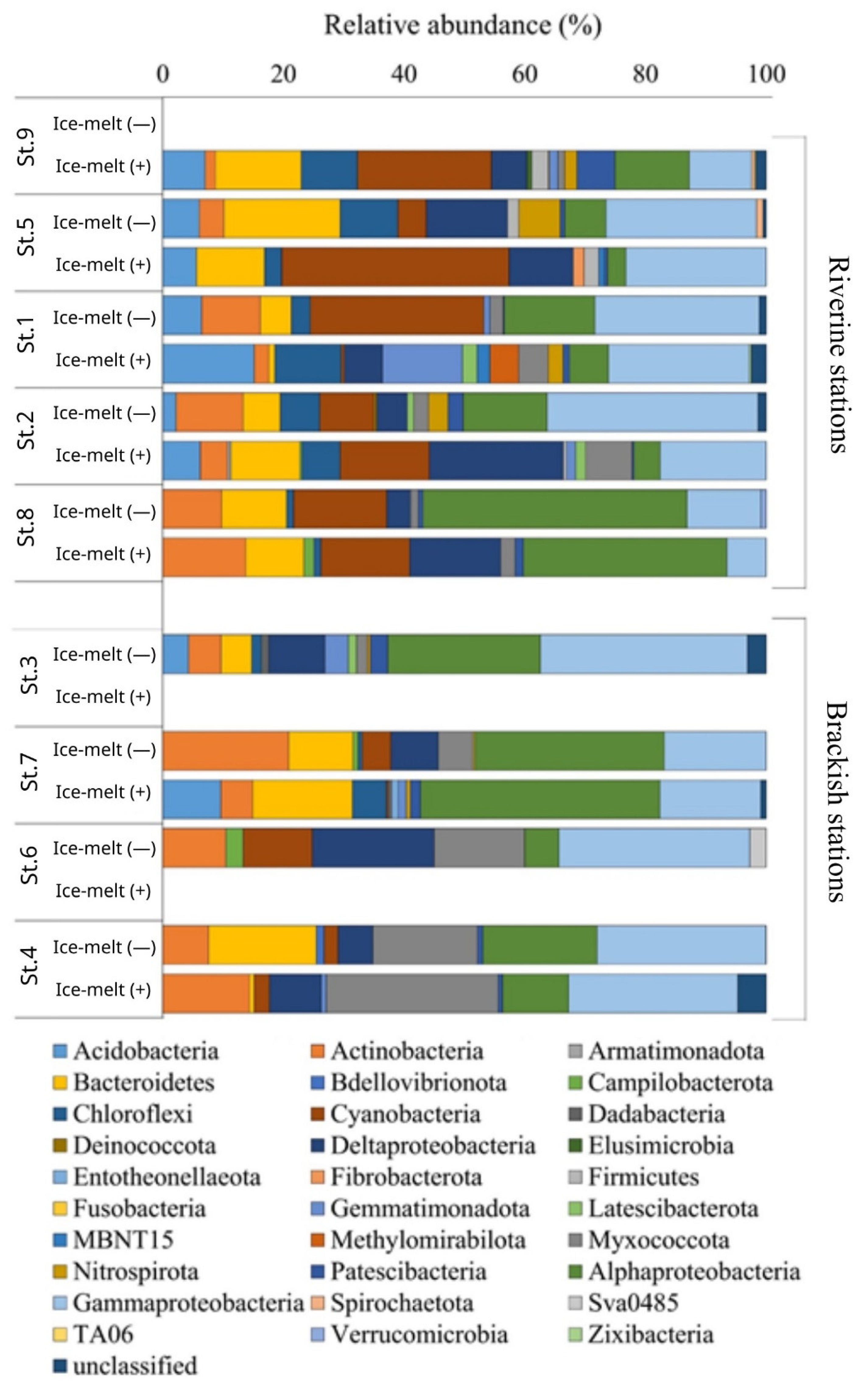
3.4. Phylogenetic Composition of the Bacterial Community
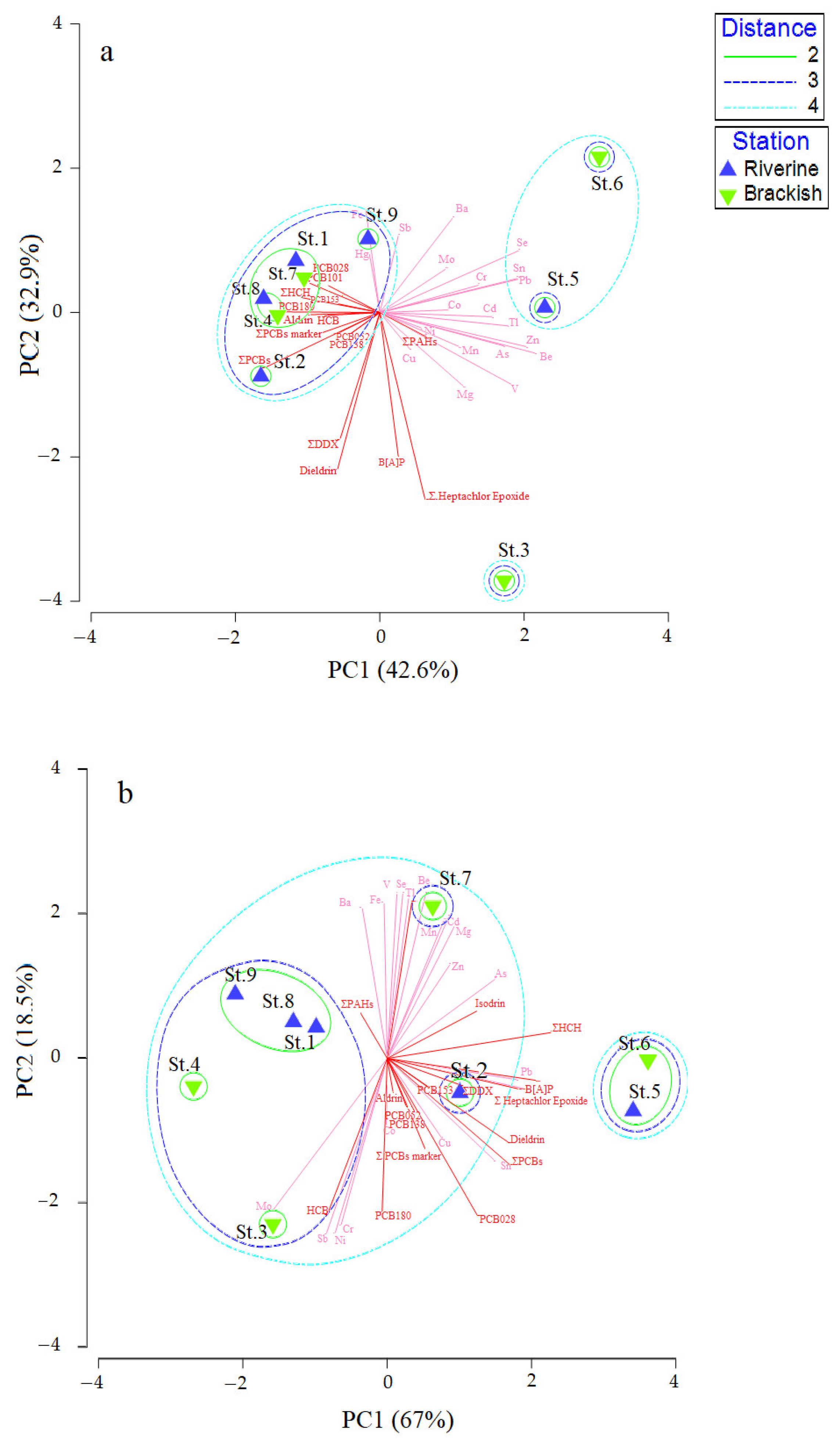
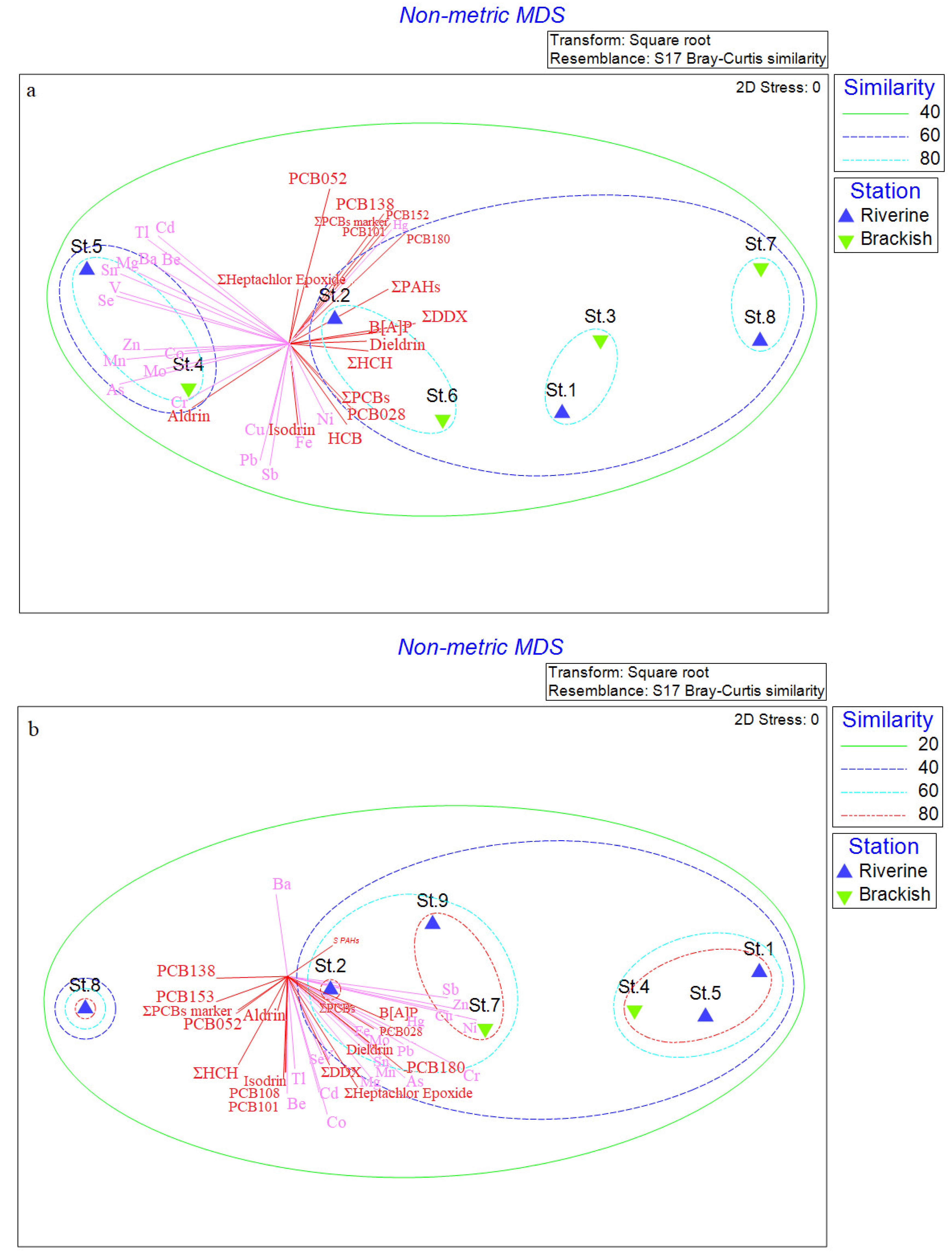
3.5. Statistical Analyses
4. Discussion
4.1. Chemical Contamination in Sediments of the Pasvik River
4.2. Estimation of Microbial Biomass and Enzymatic Activities
4.3. Main Features of the Bacterial Community along the Pasvik River
4.4. Bacteria Involved in the Biodegradation of Organic Pollutants
4.5. Bacteria Involved in Biogeochemical Cycles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jordaan, K.; Comeau, A.M.; Khasa, D.P.; Bezuidenhout, C.C. An integrated insight into the response of bacterial communities to anthropogenic contaminants in a river: A case study of the Wonderfonteinspruit catchment area, South Africa. PLoS ONE 2019, 14, e0216758. [Google Scholar] [CrossRef] [PubMed]
- Rajeev, M.; Sushmitha, T.J.; Aravindraja, C.; Toleti, S.R.; Pandian, S.K. Exploring the impacts of heavy metals on spatial variations of sediment-associated bacterial communities. Ecotoxicol. Environ. 2021, 209, 111808. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Niu, J.; Ren, Y.; Cong, J.; Zhang, X.; Fan, F.; Xiao, Y.; Zhang, X.; Deng, J.; Xie, M.; et al. An integrated insight into the response of sedimentary microbial communities to heavy metal contamination. Sci. Rep. 2015, 5, 14266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauvalter, V.; Kashulin, N.; Sandimirov, S.; Terentjev, P.; Denisov, D.; Amundsen, P.A. Chemical composition of lake sediments along a pollution gradient in a Subarctic watercourse. J. Environ. Sci. Health A 2011, 46, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Dauvalter, V.; Rognerud, S. Heavy metal pollution in sediments of the Pasvik River drainage. Chemosphere 2001, 42, 9–18. [Google Scholar] [CrossRef]
- Mousavi, S.K.; Primicerio, R.; Amundsen, P.-A. Diversity and structure of Chironomidae (Diptera) communities along a gradient of heavy metal contamination in a subarctic watercourse. Sci. Total Environ. 2003, 307, 93–110. [Google Scholar]
- Amundsen, P.A.; Kashulin, N.A.; Terentjev, P.; Gjelland, K.Ø.; Koroleva, I.M.; Dauvalter, V.A.; Sandimirov, S.; Kashulin, A.; Knudsen, R. Heavy metal contents in whitefish (Coregonus lavaretus) along a pollution gradient in a subarctic watercourse. Environ. Monit. Assess. 2011, 182, 301–316. [Google Scholar] [CrossRef]
- Dauvalter, V.; Christensen, G.N.; Andersen, H.J. Toxic substances on the sediments of the Pasvik River. In Environmental Challenges in the Norwegian, Finnish and Russian Border Area; Ylikörkkö, J., Christensen, G.N., Kashulin, N., Denisov, D., Andersen, H.J., Jelkänen, E., Eds.; Report 41; Centre for Economic Development, Transport and the Environment for Lapland: Tampere, Finland, 2015; Chapter 3; pp. 52–59. [Google Scholar]
- Dauvalter, V.A.; Kashulin, N.A. Assessment of the ecological state of the Arctic freshwater system based on concentrations of heavy metals in the bottom sediments. Geochem. Int. 2018, 56, 842–856. [Google Scholar] [CrossRef]
- Caputo, S.; Papale, M.; Rizzo, C.; Giannarelli, S.; Conte, A.; Moscheo, F.; Graziano, M.; Aspholm, P.E.; Onor, M.; De Domenico, E.; et al. Heavy metal resistance in bacteria from contaminated Arctic sediment is driven by heavy metal local inputs. Arch. Environ. Contam. Toxicol. 2019, 77, 291–307. [Google Scholar] [CrossRef]
- Averina, M.; Hervig, T.; Huber, S.; Kjær, M.; Kristoffersen, E.K.; Bolann, B. Environmental pollutants in blood donors: The multicentre Norwegian donor study. Transfus. Med. 2020, 30, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Amundsen, P.-A.; Staldvik, F.J.; Reshetnikov, Y.S.; Kashulin, N.; Lukin, A.; Bøhn, T.; Sandlund, O.T.; Popova, O.A. Invasion of vendace Coregonus albula in a subarctic watercourse. Biol. Conserv. 1999, 88, 405–413. [Google Scholar] [CrossRef]
- Papale, M.; Rizzo, C.; Caruso, G.; Amalfitano, S.; Maimone, G.; Miserocchi, S.; La Ferla, R.; Aspholm, P.E.; Decembrini, F.; Azzaro, F.; et al. Ice melt-induced variations of structural and functional traits of the aquatic microbial community along an Arctic river (Pasvik River, Norway). Water 2021, 13, 2297. [Google Scholar] [CrossRef]
- Hedges, J.I.; Stern, J.H. Carbon and nitrogen determination of carbonate-containing solids. Limnol. Oceanogr. 1984, 19, 984–989. [Google Scholar]
- Monaco, A.; Biscaye, P.; Soyer, J.; Pocklington, R.; Heussner, S. Particle fluxes and ecosystem response on a continental margin: The 1985–1988 Mediterranean ECOMARGE experiment. Cont. Shelf Res. 1990, 10, 809–839. [Google Scholar] [CrossRef]
- Tesi, T.; Miserocchi, S.; Goni, M.E.A.; Langone, L.; Boldrin, A.; Turchetto, M. Organic matter origin and distribution in suspended particulate materials and surficial sediments from the western Adriatic Sea (Italy). Estuar. Coast. Shelf Sci. 2007, 73, 431–446. [Google Scholar] [CrossRef]
- Vagi, M.C.; Petsas, A.S.; Kostopoulou, M.N.; Karamanoli, M.K.; Lekkas, T.D. Determination of organochlorine pesticides in marine sediments samples using ultrasonic solvent extraction followed by GC/ECD. Desalination 2007, 210, 146–156. [Google Scholar] [CrossRef]
- Tian, L.; Huang, D.; Shi, Y.; Han, F.; Wang, Y.; Ye, H.; Tang, Y.; Yu, H. Method for the Analysis of 7 Indictor Polychlorinated Biphenyls (PCBs) and 13 Organochlorine Pesticide Residues in Sediment by Gas Chromatography (GC). Earth Environ. Sci. 2019, 237, 022053. [Google Scholar] [CrossRef]
- Jensen, S.; Renberg, L.; Reutergardh, L. Residue analysis of sediment and sewage sludge for organochlorines in the presence of elemental sulfur. Anal. Chem. 1997, 49, 316–318. [Google Scholar] [CrossRef]
- Borghini, F.; Colacevich, A.; Bargagli, R. A study of autotrophic communities in two Victoria Land lakes (Continental Antarctica) using photosynthetic pigments. J. Limnol. 2010, 69, 333–340. [Google Scholar] [CrossRef] [Green Version]
- US EPA 2014. Method 3620C, Florisil Cleanup; SW-846 Update V; US EPA: Santa Monica, CA, USA, 2014. [Google Scholar]
- Amalfitano, S.; Fazi, S.; Puddu, A. Flow cytometric analysis of benthic prokaryotes attached to sediment particles. J. Microbiol. Methods 2009, 79, 246–249. [Google Scholar] [CrossRef]
- Conte, A.; Papale, M.; Amalfitano, S.; Mikkonen, A.; Rizzo, C.; De Domenico, E.; Michaud, L.; Lo Giudice, A. Bacterial community structure along the subtidal sandy sediment belt of a high Arctic fjord (Kongsfjorden, Svalbard Islands). Sci. Total Environ. 2018, 619, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Amalfitano, S.; Fazi, S. Recovery and quantification of bacterial cells associated with streambed sediments. J. Microbiol. Methods 2008, 75, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.G.; Feig, Y.S. The Use of DAPI for Identifying and Counting Aquatic Microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- La Ferla, R.; Maimone, G.; Azzaro, M.; Conversano, F.; Brunet, C.; Cabral, A.S.; Paranhos, R. Vertical distribution of the prokaryotic cell size in the Mediterranean Sea. Helgol. Mar. Res. 2012, 66, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, H.G. Use of fluorogenic model substrates for extracellular enzyme activity (EEA) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publisher: Boca Raton, FL, USA, 1993; pp. 423–432. [Google Scholar]
- Papale, M.; Lo Giudice, A.; Conte, A.; Rizzo, C.; Rappazzo, C.; Maimone, G.; Caruso, G.; La Ferla, R.; Azzaro, M.; Gugliandolo, C.; et al. Microbial assemblages in pressurized Antarctic brine pockets (Tarn Flat, Northern Victoria Land): A hotspot of biodiversity and activity. Microorganisms 2019, 7, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papale, M.; Rappazzo, A.C.; Mikkonen, A.; Rizzo, C.; Moscheo, F.; Conte, A.; Michaud, L.; Lo Giudice, A. Bacterial diversity in a dynamic and extreme sub-Arctic watercourse (Pasvik River, Norwegian Arctic). Water 2020, 12, 3098. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; Da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets 1fthrough Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Rappazzo, A.C.; Papale, M.; Rizzo, C.; Conte, A.; Giannarelli, S.; Onor, M.; Abete, C.; Cefali, P.; De Domenico, E.; Michaud, L.; et al. Heavy metal Tolerance and Polychlorinated biphenyl oxidation in bacterial communities inhabiting the Pasvik River and the Varanger fjord (Arctic Norway). Mar. Poll. Bull. 2019, 141, 535–549. [Google Scholar] [CrossRef]
- Kahkashan, S.; Wang, X.; Chen, J.; Bai, Y.; Ya, M.; Wu, Y.; Cai, Y.; Wang, S.; Saleem, M.; Aftab, J.; et al. Concentration, distribution and sources of perfluoroalkyl substances and organochlorine pesticides in surface sediments of the northern Bering Sea, Chukchi Sea and adjacent Arctic Ocean. Chemosphere 2019, 235, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Evenset, A.; Christensen, G.N.; Carroll, J.; Zaborska, A.; Berger, U.; Herzke, D.; Gregor, D. Historical trends in persistent organic pollutants and metals recorded in sediment from Lake Ellasjøen, Bjørnøya, Norwegian Arctic. Environ. Pollut. 2007, 146, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Zhang, X.; Chen, X.; Liu, X.; Hu, X.; Yuan, X. Response of soil bacterial community to bioaugmentation with a plant residue-immobilized bacterial consortium for crude oil removal. Chemosphere 2019, 222, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hou, J.; Yuan, J.; Wu, Y.; Liu, W.; Luo, Y.; Christie, P. Evaluation of fatty acid derivatives in the remediation of aged PAH-contaminated soil and microbial community and degradation gene response. Chemosphere 2020, 248, 125983. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sheng, H.-F.; He, Y.; Wu, J.-Y.; Jiang, Y.-X.; Tam, N.F.-Y.; Zhou, H.-W. Comparison of the levels of bacterial diversity in freshwater, intertidal wetland, and marine sediments by using millions of illumina tags. Appl. Environ. Microbiol. 2012, 78, 8264–8271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, K.Y.; Zakaria, N.A.; Foo, K.Y. Geochemistry pollution status and ecotoxicological risk assessment of heavy metals in the Pahang River sediment after the high magnitude of flood event. Hydrol. Res. 2021, 52, 107–124. [Google Scholar] [CrossRef]
- Molvær, J.; Knutzen, J.; Magnusson, J.; Rygg, B.; Skei, J.; Sørensen, J. Klassifisering av Miljøkvalitet i Fjorder og Kystfarvann (Classification of Environmental Quality in Fjords and Coastal Areas); SFT TA-1467/1997; Norwegian Pollution Control Authority: Oslo, Norway, 1997. (In Norwegian) [Google Scholar]
- Rognerud, S.; Skotvold, T.; Fjeld, E.; Norton, S.A.; Hobæk, A. Concentrations of trace elements in recent and preindustrial sediments from Norwegian and Russian Arctic lakes. Can. J. Fish. Aquat. Sci. 1998, 55, 1512–1523. [Google Scholar] [CrossRef]
- Skotvold, T.; Wartena, E.M.M.; Rognerud, S. Heavy Metals and Persistent Organic Pollutants in Sediments and Fish from Lakes in Northern and Arctic Regions of Norway. Statlig Program for Forurensningsovervakning; Norwegian Pollution Control Authority Report 688y97; Norwegian Pollution Control Authority: Oslo, Norway, 1997. [Google Scholar]
- Koo, H.; Hakim, J.A.; Morrow, C.D.; Crowley, M.R.; Andersen, D.T.; Bej, A.K. Metagenomic analysis of microbial community compositions and cold-responsive stress genes in selected Antarctic lacustrine and soil ecosystems. Life 2018, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Portillo, M.C.; Anderson, S.P.; Fierer, N. Temporal variability in the diversity and composition of stream bacterioplankton communities. Environ. Microbiol. 2012, 14, 2417–2428. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [Green Version]
- Cruaud, P.; Vigneron, A.; Fradette, M.-S.; Dorea, C.; Culley, A.; Rodriguez, M.; Charette, S. Annual bacterial community cycle in a seasonally ice-covered river reflects environmental and climatic conditions. Limnol. Oceanogr. 2020, 65, S21–S37. [Google Scholar] [CrossRef]
- Callieri, C.; Cronberg, G.; Stockner, J.G. Freshwater picocyanobacteria: Single cells, microcolonies and colonial forms. In Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: New York, NY, USA, 2012; pp. 229–269. [Google Scholar]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the Cytophaga–Flavobacter cluster consuming low-and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, M.T.; Sommaruga, R. Differential effect of algal- and soil-derived dissolved organic matter on alpine lake bacterial community composition and activity. Limnol. Oceanogr. 2006, 51, 2527–2537. [Google Scholar] [CrossRef] [Green Version]
- Podar, M.; May, A.L.; Bai, W.; Peyton, K.; Klingeman, W.M.; Swift, C.M.; Linson, D.A.F.; Mathieu, J.; Siljeström, D.; Beneyto, I.; et al. Microbial diversity analysis of two full-scale seawater desalination treatment trains provides insights into detrimental biofilm formation. J. Membr. Sci. Lett. 2021, 1, 100001. [Google Scholar] [CrossRef]
- Uchino, Y.; Hamada, T.; Yokota, A. Proposal of Pseudorhodobacter ferrugineus gen. nov., comb. nov., for a non-photosynthetic marine bacterium, Agrobacterium ferrugineum, related to the genus Rhodobacter. J. Gen. Appl. Microbiol. 2002, 48, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.; Zhao, Y.; Keskar, J.; Ji, M.; Wang, Z.; Li, X. Simultaneous removal of COD, nitrogen and phosphorus and the tridimensional microbial response in a sequencing batch biofilm reactor: With varying C/N/P ratios. Biochem. Eng. J. 2020, 154, 107215. [Google Scholar] [CrossRef]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, Q.; Dong, D.; Hu, H.; Wu, B.; Ren, H. AHLs-mediated quorum sensing threshold and its response towards initial adhesion of wastewater biofilms. Water Res. 2021, 194, 116925. [Google Scholar] [CrossRef]
- Matar, G.K.; Bagchi, S.; Zhang, K.; Oerther, D.B.; Saikaly, P.E. Membrane biofilm communities in full-scale membrane bioreactors are not randomly assembled and consist of a core microbiome. Water Res. 2017, 123, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Hamdan, H.Z.; Salam, D.A.; Saikaly, P.E. Characterization of the microbial community diversity and composition of the coast of Lebanon: Potential for petroleum oil biodegradation. Mar. Poll. Bull. 2019, 149, 110508. [Google Scholar] [CrossRef]
- Papadatou, M.; Robson, S.C.; Dobretsov, S.; Watts, J.E.M.; Longyear, J.; Salta, M. Marine biofilms on different fouling control coating types reveal differences in microbial community composition and abundance. Microbiol. Open 2021, 10, e1231. [Google Scholar] [CrossRef] [PubMed]
- Laganà, P.; Votano, L.; Caruso, G.; Azzaro, M.; Lo Giudice, A.; Delia, S. Bacterial isolates from the Arctic region (Pasvik River, Norway): Assessment of biofilm production and antibiotic susceptibility profiles. Environ. Sci. Pollut. Res. 2018, 25, 1089–1102. [Google Scholar] [CrossRef] [PubMed]
- Sattley, W.M.; Madigan, M.T. Cold-active acetogenic bacteria from surficial sediments of perennially ice-covered Lake Fryxell, Antarctica. FEMS Microbiol. Lett. 2007, 272, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oug, E. Relating species patterns and environmental variables by canonical ordination: An analysis of soft-bottom macrofauna in the region of Tromsø northern Norway. Mar. Environ. Res. 1998, 45, 29–45. [Google Scholar] [CrossRef]
- Schubert, C.J.; Calvert, S.E. Nitrogen and carbon isotopic composition of marine and terrestrial organic matter in Arctic Ocean sediments: Implications for nutrient utilization and organic matter composition. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2001, 48, 789–810. [Google Scholar] [CrossRef]
- Rodríguez, J.; Gallampois, C.M.J.; Timonen, S.; Andersson, A.; Sinkko, H.; Haglund, P.; Berglund, Å.M.M.; Ripszam, M.; Figueroa, D.; Tysklind, M.; et al. Effects of Organic Pollutants on Bacterial Communities Under Future Climate Change Scenarios. Front. Microbiol. 2018, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Luo, C.; Jiang, L.; Zhang, D.; Wang, Y.; Zhang, G. Identification of Benzo[a]pyrene-Metabolizing Bacteria in Forest Soils by Using DNA-Based Stable-Isotope Probing. Appl. Environ. Microbiol. 2015, 81, 7368–7376. [Google Scholar] [CrossRef] [Green Version]
- Lambo, A.J.; Patel, T.R. Cometabolic Degradation of Polychlorinated Biphenyls at Low Temperature by Psychrotolerant Bacterium Hydrogenophaga sp. IA3-A. Curr. Microbiol. 2006, 53, 48–52. [Google Scholar] [CrossRef]
- Molina, M.; Araujo, R.; Hodson, R.E. Cross-induction of pyrene and phenanthrene in a Mycobacterium sp. isolated from polycyclic aromatic hydrocarbon contaminated river sediments. Can. J. Microbiol. 1999, 45, 520–529. [Google Scholar] [CrossRef]
- Pickup, R.W.; Rhodes, G.; Bull, T.J.; Arnott, S.; Sidi-Boumedine, K.; Hurley, M.; Hermon-Taylor, J. Mycobacterium avium subsp. paratuberculosis in lake catchments, in river water abstracted for domestic use, and in effluent from domestic sewage treatment works: Diverse opportunities for environmental cycling and human exposure. Appl. Environ. Microbiol. 2006, 72, 4067–4077. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.P.; Lou, J.; Wang, H.Z.; Yang, Y.; Wu, L.S.; Wu, J.J.; Xu, J.M. Biodegradation, biosorption of phenanthrene and its trans-membrane transport by Massilia sp. WF1 and Phanerochaete chrysosporium. Front. Microbiol. 2016, 7, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhu, S.; Lin, D.; Wang, X.; Yang, J.; Chen, J. Devosia naphthalenivorans sp. nov., isolated from East Pacific Ocean sediment. Int. J. Syst. Evol. Microbiol. 2019, 69, 1974–1979. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ge, W.; Zhang, X.; Chai, C.; Wu, J.; Xiang, D.; Chen, X. Biodegradation of aged polycyclic aromatic hydrocarbons in agricultural soil by Paracoccus sp. LXC combined with humic acid and spent mushroom substrate. J. Hazard. Mater. 2019, 379, 120820. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Yonemitsu, Y.; Matsushita, M.; Shin, Y.K.; Kuraishi, H.; Kawahara, K. Characterization of Porphyrobacter sanguineus sp. nov., an aerobic bacteriochlorophyll-containing bacterium capable of degrading biphenyl and dibenzofuran. Arch. Microbiol. 2002, 178, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Gray, N.D.; Sherry, A.; Grant, R.J.; Rowan, A.K.; Hubert, C.R.J.; Callbeck, C.M.; Aitken, C.M.; Jones, D.M.; Adams, J.J.; Larter, S.R.; et al. The quantitative significance of Syntrophaceae and syntrophic partnerships in methanogenic degradation of crude oil alkanes. Environ. Microbiol. 2011, 13, 2957–2975. [Google Scholar] [CrossRef] [Green Version]
- Junghare, M.; Spiteller, D.; Schink, B. Anaerobic degradation of xenobiotic isophthalate by the fermenting bacterium Syntrophorhabdus aromaticivorans. ISME J. 2019, 13, 1252–1268. [Google Scholar] [CrossRef]
- Ji, J.H.; Zhou, L.; Mbadinga, S.M.; Irfan, M.; Liu, Y.-F.; Pan, P.; Qi, Z.-Z.; Chen, J.; Liu, J.-F.; Yang, S.-Z.; et al. Methanogenic biodegradation of C9 to C12n-alkanes initiated by Smithella via fumarate addition mechanism. AMB Express 2020, 10, 23. [Google Scholar] [CrossRef]
- Hedges, J.I.; Clark, W.A.; Quay, P.D.; Richey, J.E.; Devol, A.H.; Santos, U.M. Composition and fluxes of particulate organic material in the Amazon River. Limnol. Oceanogr. 1986, 31, 717–738. [Google Scholar] [CrossRef] [Green Version]
- Yong, Z.; Lin, M.; Xiong, W.; Wang, M.; Zhang, J.; Wang, M.; Sun, Y. Metagenomic insights into the effect of oxytetracycline on microbial structures, functions and functional genes in sediment denitrification. Ecotoxicol. Environ. Saf. 2018, 161, 85–91. [Google Scholar]
- Yin, S.; Li, J.; Dong, H.; Qiang, Z. Enhanced nitrogen removal through marine anammox bacteria (MAB) treating nitrogen-rich saline wastewater with Fe(III) addition: Nitrogen shock loading and community structure. Bioresour. Technol. 2019, 287, 1214052. [Google Scholar] [CrossRef]
- Ahmad, H.A.; Guo, B.; Zhuang, X.; Zhao, Y.; Ahmad, S.; Lee, T.; Zhu, J.; Dong, Y.; Nie, S.-O. A twilight for the complete nitrogen removal via synergistic partial-denitrification, anammox, and DNRA process. NPJ Clean Water 2021, 4, 31. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Lücker, S.; Vejmelkova, D.; Kostrikina, N.A.; Kleerebezem, R.; Rijpstra, W.I.; Damsté, J.S.; Le Paslier, D.; Muyzer, G.; Wagner, M.; et al. Nitrification expanded: Discovery, physiology and genomics of a nitrite-oxidizing bacterium from the phylum Chloroflexi. ISME J. 2012, 6, 2245–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaichi, S.; Ito, N.; Kamikawa, R.; Sugawara, T.; Yoshida, T.; Sako, Y. Ardenticatena maritima gen. nov., sp. nov., a ferric iron- and nitrate reducing bacterium of the phylum ‘Chloroflexi’ isolated from an iron-rich coastal hydrothermal field, and description of Ardenticatenia classis nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 2992–3002. [Google Scholar] [CrossRef] [PubMed]
- Ettwig, K.F.; Butler, M.K.; Le Paslier, D.; Pelletier, E.; Mangenot, S.; Kuypers, M.M.M.; Schreiber, F.; Dutilh, B.E.; Zedelius, J.; De Beer, D.; et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 2010, 464, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, K.; Izallalen, M.; Mouser, P.; Richter, H.; Risso, C.; Mahadevan, R.; Lovely, D.R. Genome-scale dynamic modeling of the competition between Rhodoferax and Geobacter in anoxic subsurface environments. ISME J. 2011, 5, 305–316. [Google Scholar] [CrossRef]
- Masuda, Y.; Shiratori, Y.; Ohba, H.; Ishida, T.; Takano, R.; Satoh, S.; Shen, W.; Gao, N.; Itoh, H.; Senoo, K. Enhancement of the nitrogen-fixing activity of paddy soils owing to iron application. Soil Sci. Plant Nutr. 2021, 67, 243–247. [Google Scholar] [CrossRef]
- Finneran, K.T.; Johnsen, C.V.; Lovely, D.R. Rhodoferax ferrireducens sp. nov., a psychrotolerant, facultatively anaerobic bacterium that oxidizes acetate with the reduction of Fe(III). Int. J. Syst. Evol. Microbiol. 2003, 53, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.H.; Baek, S.K.; Lee, S.T. Ferruginibacter alkalilentus gen. nov., sp. nov. and Ferruginibacter lapsinanis sp. nov., novel members of the family Chitinophagaceae in the phylum Bacteroidetes, isolated from freshwater sediment. Int. J. Syst. Evol. Microbiol. 2009, 59, 2394–2399. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Sun, X.; Han, F.; Li, B.; Xiao, E.; Xiao, T.; Yang, Z.; Sun, W. Impacts of antimony and arsenic co-contamination on the river sedimentary microbial community in an antimony-contaminated river. Sci. Total Environ. 2020, 713, 136451. [Google Scholar] [CrossRef]
- Tahon, G.; Willems, A. Isolation and characterization of aerobic anoxygenic phototrophs from exposed soils from the Sør Rondane Mountains, East Antarctica. Syst. Appl. Microbiol. 2017, 40, 357–369. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter * | Acronym | Riverine Stations | Brackish Stations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | |||
| Grain size (%) | MUD ** | Ice-melt(−) | 13.4 | 39.0 | 5.3 | 5.6 | 6.4 | 44.5 | 6.2 | 20.4 | 10.3 |
| Ice-melt(+)ple | 0.9 | 60.2 | 16.6 | 49 | 12.8 | 45.4 | 1.4 | 51.3 | 2.5 | ||
| SAND ** | Ice-melt(−) | 86.6 | 61.0 | 94.7 | 94.4 | 93.6 | 55.5 | 93.8 | 79.6 | 89.7 | |
| Ice-melt(+) | 99.1 | 39.8 | 83.4 | 51 | 87.2 | 54.6 | 98.6 | 48.7 | 97.5 | ||
| Organic C content (%) | C-ORG ** | Ice-melt(−) | 0.67 | 0.49 | 0.54 | 0.12 | 0.19 | 1.90 | 0.54 | 0.95 | 0.12 |
| Ice-melt(+) | 0.08 | 1.19 | 0.22 | 0.64 | 0.75 | nd | 0.75 | 1.34 | nd | ||
| δ13C ‰ | D13C | Ice-melt(−) | −28.60 | −27.20 | −24.35 | −24.96 | −24.60 | −22.65 | −22.07 | −22.82 | −23.08 |
| Ice-melt(+) | −26.31 | −27.07 | −20.13 | −25.14 | −25.09 | nd | −24.92 | −23.44 | nd | ||
| Nitrogen content (%) | N-TOT | Ice-melt(−) | 0.047 | 0.085 | 0.040 | nd | 0.018 | nd | 0.044 | 0.118 | 0.011 |
| Ice-melt(+) | nd | 0.14 | 0.02 | 0.05 | nd | nd | 0.04 | 0.1 | nd | ||
| Persistant Organic Pollutants * | Riverine Stations | Brackish Stations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | |||
| Polycyclic aromatic hydrocarbons (ng g−1) | Benzo[A]Pyrene | Ice-melt(−) | 3.64 | 0.55 | 0.25 | 0.12 | 11.19 | 19.55 | 0.11 | <LOD | 0.04 |
| Ice-melt(+) | 0.03 | 50.76 | 29.45 | <LOD | 14.24 | 0.28 | 1.21 | 22.32 | 0.22 | ||
| ∑ PAHs | Ice-melt(−) | 142.02 | 192.80 | 27.74 | 26.33 | 681.17 | 141.90 | 125.54 | 38.01 | 1.80 | |
| Ice-melt(+) | 12.99 | 183.30 | 2969.23 | 24.01 | 1267.19 | 41.81 | 87.73 | 575.02 | 60.96 | ||
| Polychlorinated biphenyls (ng g−1) | ∑ PCB marker | Ice-melt(−) | 0.26 | 1.12 | 1.43 | 0.88 | 0.37 | 0.99 | 13.23 | <LOD | 0.21 |
| Ice-melt(+) | 0.14 | 0.82 | 0.41 | 3.85 | 0.33 | 2.02 | 1.16 | 0.41 | 0.41 | ||
| ∑ PCBs | Ice-melt(−) | 12.95 | 12.13 | 76.42 | 6.63 | 8.48 | 11.55 | 20.15 | <LOD | 5.23 | |
| Ice-melt(+) | 5.78 | 22.94 | 3.72 | 19.64 | 1.32 | 10.30 | 4.52 | 3.56 | 2.10 | ||
| PCB028 | Ice-melt(−) | <LOD | 0.02 | 0.65 | 0.01 | <LOD | 0.05 | 0.21 | <LOD | 0.03 | |
| Ice-melt(+) | 0.01 | 0.15 | <LOD | <LOD | <LOD | 0.39 | <LOD | 0.34 | <LOD | ||
| PCB052 | Ice-melt(−) | <LOD | 0.02 | 0.65 | 0.01 | <LOD | 0.10 | 0.33 | <LOD | 0.02 | |
| Ice-melt(+) | <LOD | 0.08 | 0.16 | 0.98 | 0.05 | 0.54 | 0.46 | 0.07 | 0.01 | ||
| PCB101 | Ice-melt(−) | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | 1.64 | <LOD | <LOD | |
| Ice-melt(+) | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | 0.08 | <LOD | <LOD | ||
| PCB138 | Ice-melt(−) | 0.13 | 0.52 | 0.22 | 0.34 | 0.03 | 0.51 | 4.71 | <LOD | 0.05 | |
| Ice-melt(+) | 0.05 | <LOD | 0.17 | 2.09 | 0.15 | 0.51 | 0.34 | <LOD | 0.03 | ||
| PCB153 | Ice-melt(−) | 0.04 | 0.29 | 0.12 | 0.15 | 0.02 | 0.17 | 4.40 | <LOD | 0.03 | |
| Ice-melt(+) | 0.03 | 0.13 | 0.08 | 0.78 | 0.12 | 0.16 | 0.21 | <LOD | 0.02 | ||
| PCB180 | Ice-melt(−) | 0.07 | <LOD | 0.24 | 0.27 | 0.20 | 0.16 | 1.93 | <LOD | 0.08 | |
| Ice-melt(+) | 0.04 | 0.45 | <LOD | <LOD | 0.01 | 0.41 | 0.08 | <LOD | 0.34 | ||
| Pesticides (ng g−1) | HCB | Ice-melt(−) | <LOD | <LOD | 0.25 | 0.01 | <LOD | 0.05 | 0.04 | <LOD | <LOD |
| Ice-melt(+) | <LOD | <LOD | <LOD | <LOD | <LOD | 0.19 | 0.02 | <LOD | <LOD | ||
| Aldrin | Ice-melt(−) | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | 0.01 | |
| Ice-melt(+) | 0.01 | <LOD | <LOD | <LOD | <LOD | 0.04 | 0.03 | 0.02 | <LOD | ||
| Isodrin | Ice-melt(−) | 0.28 | 53.17 | 287.03 | 11.85 | 6.56 | 2.63 | 0.16 | <LOD | 2.56 | |
| Ice-melt(+) | 0.15 | 2.27 | 0.16 | 0.98 | 1.49 | 24.41 | 43.58 | 34.77 | 0.25 | ||
| Dieldrin | Ice-melt(−) | 3.24 | <LOD | 10.18 | 1.27 | 2.65 | 42.80 | 7.23 | <LOD | 1.01 | |
| Ice-melt(+) | <LOD | 175.30 | 0.04 | <LOD | 6.70 | 31.25 | <LOD | <LOD | <LOD | ||
| ∑ HCH | Ice-melt(−) | 2.17 | 22.26 | 39.76 | 1.83 | 58.82 | 1.72 | 6.46 | <LOD | 0.86 | |
| Ice-melt(+) | 0.48 | 3.33 | 2.00 | 15.47 | 2.73 | 7.58 | 18.54 | 38.78 | 0.02 | ||
| ∑ DDX | Ice-melt(−) | 0.15 | 1.68 | 2.27 | 0.76 | 16.85 | 8.11 | 1.84 | <LOD | 0.28 | |
| Ice-melt(+) | 0.71 | 10.71 | 8.50 | 10.84 | 3.76 | 7.98 | 9.70 | <LOD | 0.28 | ||
| ∑ Heptachlor Epoxide | Ice-melt(−) | 0.15 | 30.15 | 5.28 | 2.93 | 10.27 | 71.71 | 3.52 | <LOD | 1.72 | |
| Ice-melt(+) | 0.17 | 71.70 | 0.58 | 15.56 | 0.76 | 21.88 | 37.63 | <LOD | 0.18 | ||
| Morphometric Traits | Riverine Stations | Brackish Stations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | ||
| Mean length (µm) | Ice-melt(−) | 0.615 | 0.940 | 0.905 | 1.294 | 1.358 | 0.492 | 1.573 | 1.002 | 1.135 |
| Ice-melt(+) | 0.615 | 1.065 | 0.563 | 0.727 | 0.929 | 0.518 | 0.905 | 1.812 | 1.076 | |
| Mean width (µm) | Ice-melt(−) | 0.387 | 0.365 | 0.403 | 0.338 | 0.282 | 0.421 | 0.438 | 0.380 | 0.308 |
| Ice-melt(+) | 0.387 | 0.369 | 0.375 | 0.372 | 0.331 | 0.429 | 0.368 | 0.393 | 0.420 | |
| Mean volume (µm3) | Ice-melt(−) | 0.050 | 0.072 | 0.076 | 0.098 | 0.075 | 0.059 | 0.186 | 0.091 | 0.068 |
| Ice-melt(+) | 0.072 | 0.102 | 0.049 | 0.059 | 0.071 | 0.063 | 0.079 | 0.170 | 0.117 | |
| CCC (fg C cell−1) | Ice-melt(−) | 16 | 22 | 22 | 29 | 23 | 18 | 50 | 27 | 21 |
| Ice-melt(+) | 22 | 29 | 16 | 19 | 22 | 19 | 24 | 46 | 33 | |
| Cocci (%) | Ice-melt(−) | 78.7 | 46.2 | 67.6 | 16.9 | 16.1 | 81.1 | 19.0 | 25.0 | 15.6 |
| Ice-melt(+) | 23.1 | 29.4 | 68.3 | 41.8 | 15.4 | 75.4 | 30.3 | 20.9 | 45.8 | |
| Rods (%) | Ice-melt(−) | 0.0 | 35.9 | 11.8 | 44.1 | 0.0 | 5.7 | 50.0 | 33.8 | 0.0 |
| Ice-melt(+) | 38.5 | 37.3 | 9.5 | 26.6 | 74.4 | 7.7 | 47.0 | 11.6 | 14.6 | |
| Vibrios (%) | Ice-melt(−) | 0.0 | 0.0 | 2.9 | 3.4 | 12.9 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ice-melt(+) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Coccobacilli (%) | Ice-melt(−) | 0.0 | 5.1 | 5.9 | 3.4 | 0.0 | 13.2 | 11.9 | 13.7 | 9.4 |
| Ice-melt(+) | 30.8 | 13.7 | 12.7 | 16.5 | 5.1 | 13.8 | 12.1 | 16.3 | 12.5 | |
| Curved rods (%) | Ice-melt(−) | 21.3 | 7.7 | 7.4 | 28.8 | 71.0 | 0.0 | 9.5 | 27.5 | 75.0 |
| Ice-melt(+) | 7.7 | 19.6 | 9.5 | 15.2 | 5.1 | 3.1 | 10.6 | 51.2 | 27.1 | |
| Filamentous forms (%) | Ice-melt(−) | 0.0 | 5.1 | 4.4 | 3.4 | 0.0 | 0.0 | 9.6 | 0.0 | 0.0 |
| Ice-melt(+) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Riverine Stations | Brackish Stations | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| St. 9 | St. 5 | St. 1 | St. 2 | St. 8 | St. 3 | St. 7 | St. 6 | St. 4 | |||||||||||
| Ice-Melt Period | Ice-Melt Period | ||||||||||||||||||
| Phylum | Genus | [−] | [+] | [−] | [+] | [−] | [+] | [−] | [+] | [−] | [+] | [−] | [+] | [−] | [+] | [−] | [+] | [−] | [+] |
| Acidobacteriota | Bryobacter | nd | 0.9 | 2.1 | 2.9 | nd | nd | ||||||||||||
| Solibacter | nd | 1.4 | nd | nd | |||||||||||||||
| Paludibaculum | nd | 0.8 | 3.7 | 2.4 | 2.6 | nd | nd | ||||||||||||
| Blastocatella | nd | nd | 1.9 | nd | |||||||||||||||
| Ilumatobacter | nd | 2.7 | 4.8 | 8.1 | nd | 8.0 | 3.0 | 3.9 | nd | 4.1 | 8.4 | ||||||||
| Actinobacteriota | Kineococcus | nd | 2.8 | nd | nd | ||||||||||||||
| Mycobacterium | nd | 5.8 | 1.6 | 2.4 | nd | nd | |||||||||||||
| Nakamurella | nd | nd | 1.5 | nd | |||||||||||||||
| Oryzihumus | nd | 4.1 | 1.7 | nd | nd | ||||||||||||||
| Gaiella | nd | 0.7 | 3.0 | nd | nd | ||||||||||||||
| Bacteroidetes | Ferruginibacter | nd | 3.4 | 4.4 | 3.4 | nd | 5.6 | nd | |||||||||||
| Winogradskyella | nd | nd | nd | 8.1 | |||||||||||||||
| Lentimicrobium | nd | 2.4 | 4.8 | 6.8 | nd | nd | |||||||||||||
| Lutimonas | nd | 1.3 | nd | 4.2 | nd | ||||||||||||||
| Maribacter | nd | nd | 5.4 | nd | 8.6 | ||||||||||||||
| Subsaxibacter | nd | nd | 1.5 | nd | |||||||||||||||
| Terrimonas | nd | 1.1 | 3.0 | 2.0 | 0.2 | nd | 0.5 | nd | |||||||||||
| Ignavibacterium | nd | 1.4 | 0.2 | nd | nd | ||||||||||||||
| Campylobacterota | Sulfurovum | nd | 0.4 | 1.7 | nd | 0.8 | 2.8 | nd | |||||||||||
| Chloroflexi | Kouleothrix | nd | 1.6 | nd | nd | ||||||||||||||
| Cyanobacteria | Aphanizomenon | nd | 2.8 | 3.9 | 1.6 | 0.2 | 0.4 | nd | nd | ||||||||||
| Cyanobium | nd | 6.4 | 7.5 | 6.0 | 2.8 | nd | nd | ||||||||||||
| Tolypothrix | nd | 0.6 | 2.9 | 1.8 | nd | nd | |||||||||||||
| Desulfobacterota | Desulfatirhabdium | nd | 2.2 | nd | nd | ||||||||||||||
| Desulfosarcina | nd | 1.8 | 5.8 | nd | 1.6 | nd | 1.7 | 5.9 | |||||||||||
| Desulfobulbus | nd | 1.1 | 2.6 | 2.3 | nd | nd | |||||||||||||
| Desulfoprunum | nd | 1.3 | nd | nd | |||||||||||||||
| Citrifermentans | nd | 2.1 | 6.6 | nd | nd | ||||||||||||||
| Desulfuromusa | nd | 0.7 | 1.5 | 4.5 | nd | 2.2 | nd | ||||||||||||
| Geobacter | nd | 5.3 | 5.7 | 5.1 | 5.3 | nd | nd | ||||||||||||
| Smithella | nd | 5.9 | 3.5 | nd | nd | ||||||||||||||
| Syntrophus | nd | 1.8 | 3.3 | nd | nd | ||||||||||||||
| Syntrophorhabdus | nd | 0.5 | 1.4 | nd | nd | ||||||||||||||
| Firmicutes | Acetobacterium | nd | 1.9 | 0.6 | nd | nd | |||||||||||||
| Anaerovorax | nd | 1.1 | nd | nd | |||||||||||||||
| Clostridium | nd | 1.2 | nd | nd | |||||||||||||||
| Latescibacterota | Latescibacterota | nd | 2.5 | 1.1 | 1.8 | 1.4 | nd | nd | |||||||||||
| Methylomirabilota | Methylomirabilis | nd | 4.9 | nd | nd | ||||||||||||||
| Myxococcota | Anaeromyxobacter | nd | 1.2 | 2.2 | nd | nd | |||||||||||||
| Phaselicystis | nd | 1.7 | nd | nd | |||||||||||||||
| Nitrospirota | Nitrospira | nd | 2.5 | 3.2 | 0.5 | nd | 0.3 | 0.7 | nd | ||||||||||
| Patescibacteria | Kaiserbacteria | nd | 1.7 | 1.0 | 0.7 | 0.3 | 1.4 | nd | 1.7 | nd | 0.5 | 0.7 | |||||||
| Moranbacteria | nd | 1.1 | 0.7 | 1.0 | nd | nd | 0.2 | ||||||||||||
| Staskawiczbacteria | nd | 1.6 | nd | nd | |||||||||||||||
| Alphaproteobacteria | Afifella | nd | 2.7 | nd | nd | ||||||||||||||
| Bradyrhizobium | nd | 4.9 | nd | nd | |||||||||||||||
| Devosia | nd | nd | 2.6 | nd | |||||||||||||||
| Erythrobacter | nd | 1.3 | nd | nd | |||||||||||||||
| Limibaculum | nd | nd | 8.0 | nd | |||||||||||||||
| Methylocystis | nd | 6.7 | nd | nd | |||||||||||||||
| Paracoccus | nd | nd | 5.3 | nd | |||||||||||||||
| Polymorphobacter | nd | 1.5 | 3.0 | 2.6 | nd | nd | |||||||||||||
| Porphyrobacter | nd | 1.7 | nd | nd | |||||||||||||||
| Pseudahrensia | nd | nd | 1.5 | nd | |||||||||||||||
| Pseudolabrys | nd | 3.6 | nd | nd | |||||||||||||||
| Pseudorhodobacter | nd | 0.2 | 11.6 | 7.5 | nd | nd | |||||||||||||
| Robiginitomaculum | nd | nd | nd | 1.4 | 3.0 | ||||||||||||||
| Roseomonas | nd | 1.5 | nd | nd | |||||||||||||||
| Sphingopyxis | nd | nd | 1.5 | nd | |||||||||||||||
| Sphingorhabdus | nd | 2.7 | 1.7 | nd | 1.2 | 1.3 | nd | ||||||||||||
| Yoonia-Loktanella | nd | nd | 16.5 | nd | |||||||||||||||
| Gammaproteobacteria | Acidibacter | nd | 1.1 | 1.2 | nd | nd | |||||||||||||
| Arenimonas | nd | 1.2 | 0.6 | nd | nd | ||||||||||||||
| Crenothrix | nd | 3.7 | nd | nd | |||||||||||||||
| Dechloromonas | nd | 1.9 | nd | nd | |||||||||||||||
| Gallionella | nd | 1.7 | 4.8 | nd | nd | ||||||||||||||
| Granulosicoccus | nd | nd | 3.3 | nd | |||||||||||||||
| Halioglobus | nd | 1.8 | nd | nd | |||||||||||||||
| Hydrogenophaga | nd | 1.7 | 3.3 | nd | 2.4 | nd | |||||||||||||
| Marinicella | nd | nd | nd | 5.6 | |||||||||||||||
| Massilia | nd | 5.0 | nd | nd | |||||||||||||||
| Nitrosomonas | nd | 2.3 | nd | nd | 1.3 | ||||||||||||||
| Porticoccus | nd | 1.5 | nd | nd | |||||||||||||||
| Pseudomonas | nd | nd | 2.5 | nd | |||||||||||||||
| Psychromonas | nd | 1.2 | nd | nd | 4.2 | ||||||||||||||
| Rhizobacter | nd | 3.5 | 2.3 | nd | nd | ||||||||||||||
| Rhodoferax | nd | 6.6 | nd | nd | |||||||||||||||
| Woeseia | nd | 22.7 | nd | 12.6 | nd | ||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papale, M.; Rizzo, C.; Giannarelli, S.; Caruso, G.; Amalfitano, S.; Aspholm, P.E.; Maimone, G.; Miserocchi, S.; Rappazzo, A.C.; Lo Giudice, A.; et al. Benthic Microbial Communities in a Seasonally Ice-Covered Sub-Arctic River (Pasvik River, Norway) Are Shaped by Site-Specific Environmental Conditions. Microorganisms 2022, 10, 1022. https://doi.org/10.3390/microorganisms10051022
Papale M, Rizzo C, Giannarelli S, Caruso G, Amalfitano S, Aspholm PE, Maimone G, Miserocchi S, Rappazzo AC, Lo Giudice A, et al. Benthic Microbial Communities in a Seasonally Ice-Covered Sub-Arctic River (Pasvik River, Norway) Are Shaped by Site-Specific Environmental Conditions. Microorganisms. 2022; 10(5):1022. https://doi.org/10.3390/microorganisms10051022
Chicago/Turabian StylePapale, Maria, Carmen Rizzo, Stefania Giannarelli, Gabriella Caruso, Stefano Amalfitano, Paul Eric Aspholm, Giovanna Maimone, Stefano Miserocchi, Alessandro Ciro Rappazzo, Angelina Lo Giudice, and et al. 2022. "Benthic Microbial Communities in a Seasonally Ice-Covered Sub-Arctic River (Pasvik River, Norway) Are Shaped by Site-Specific Environmental Conditions" Microorganisms 10, no. 5: 1022. https://doi.org/10.3390/microorganisms10051022
APA StylePapale, M., Rizzo, C., Giannarelli, S., Caruso, G., Amalfitano, S., Aspholm, P. E., Maimone, G., Miserocchi, S., Rappazzo, A. C., Lo Giudice, A., & Azzaro, M. (2022). Benthic Microbial Communities in a Seasonally Ice-Covered Sub-Arctic River (Pasvik River, Norway) Are Shaped by Site-Specific Environmental Conditions. Microorganisms, 10(5), 1022. https://doi.org/10.3390/microorganisms10051022