
Hfq Is a Critical Modulator of Pathogenicity of Dickeya oryzae in Rice Seeds and Potato Tubers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Cultural Conditions
2.2. Construction of the HfqEC1 Deletion Mutant and Complementation Strain
2.3. Cell Wall–Degrading Enzyme Activities
2.4. Motility Assay and Biofilm Formation
2.5. Zeamine Production Assay
2.6. RNA Extraction and Quantitative Real-Time Reverse-Transcription PCR (qRT-PCR) Analysis
2.7. Pathogenicity Assay on Potato Tuber Slices
2.8. Rice Seed Germination Assay
2.9. Hypersensitive Response Assay
2.10. Statistical Analysis
3. Results
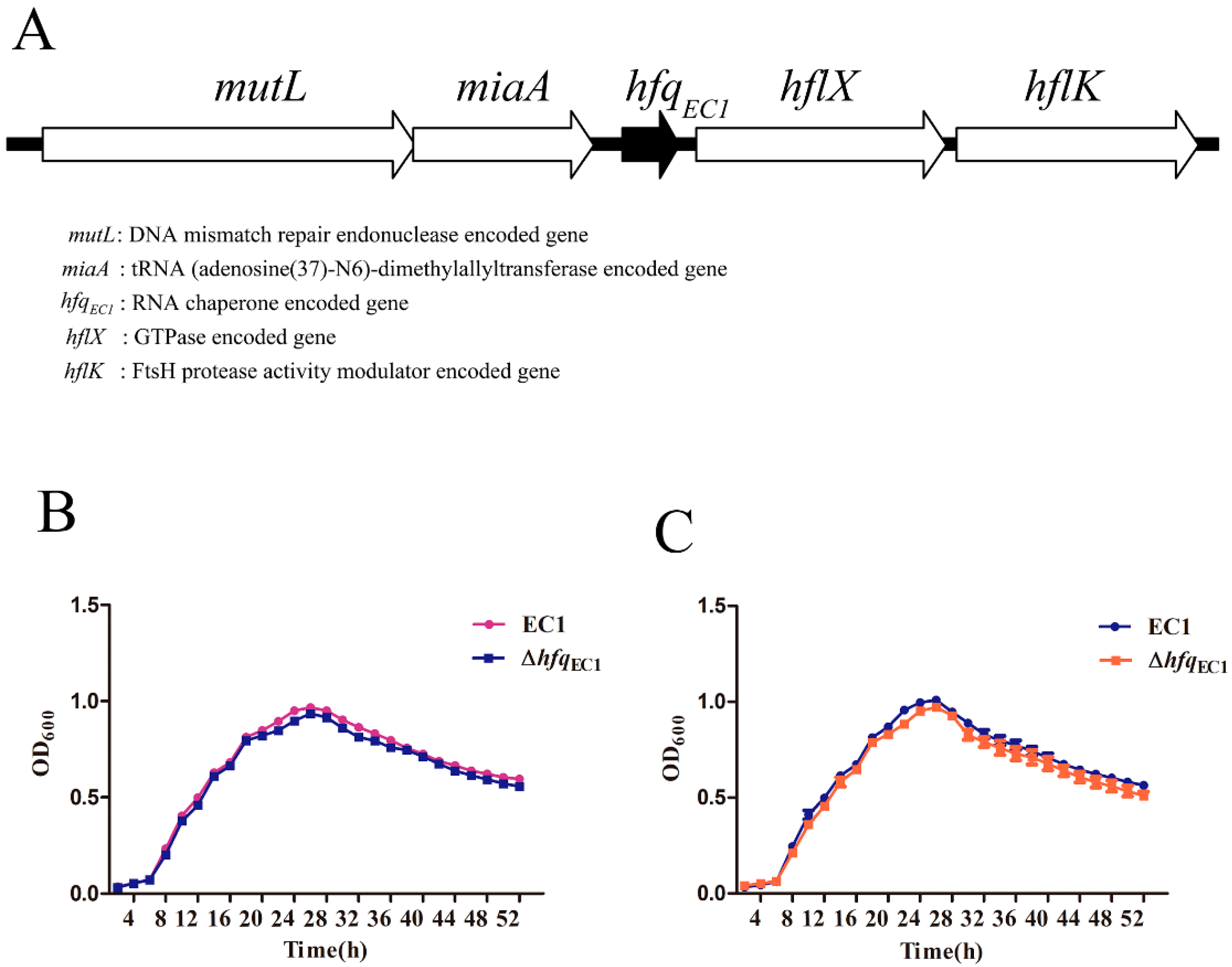
3.1. Identification of Hfq in D. oryzae EC1
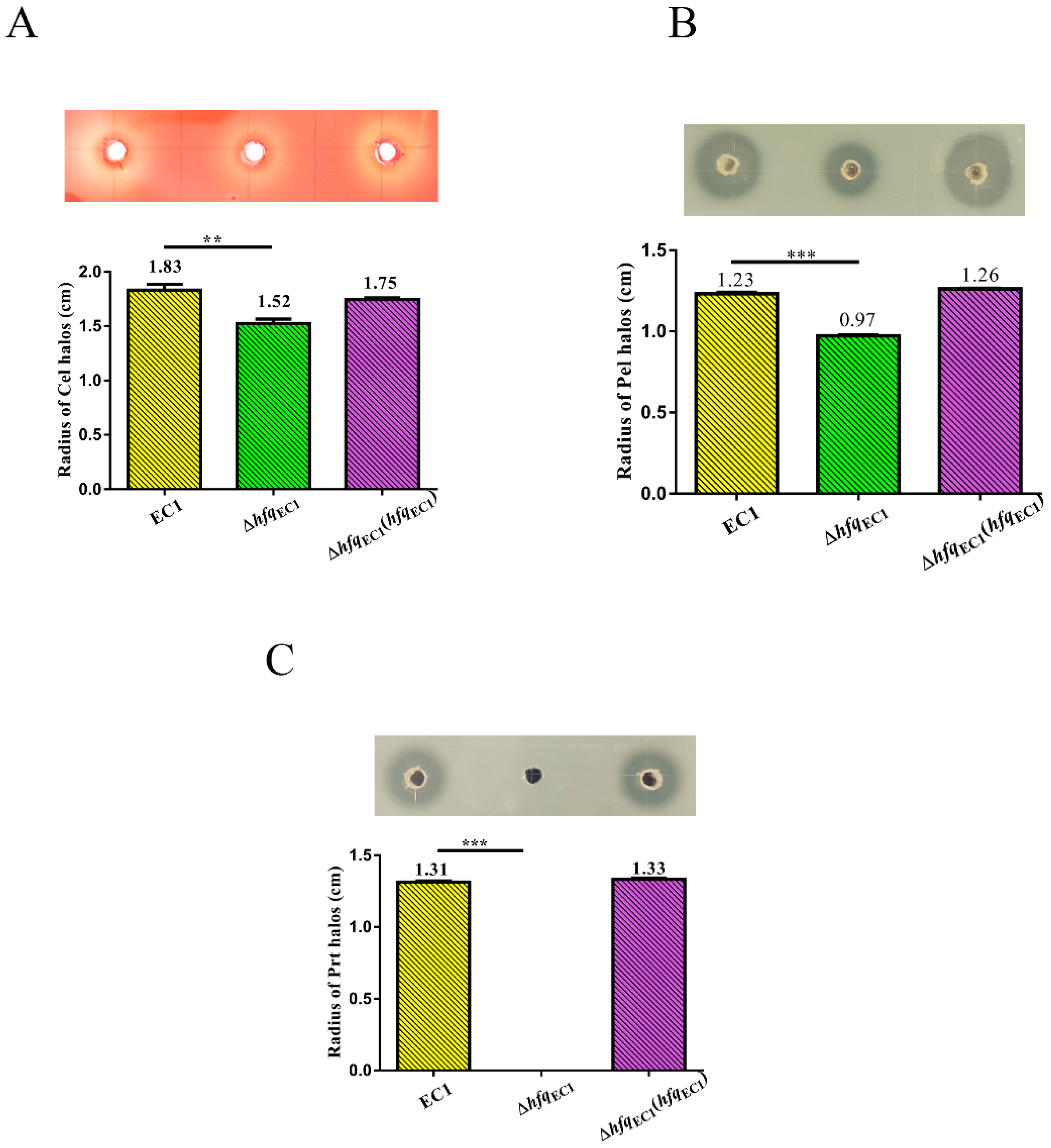
3.2. Deletion of HfqEC1 Decreases the Production of Extracellular Degrading Enzymes in D. oryzae EC1
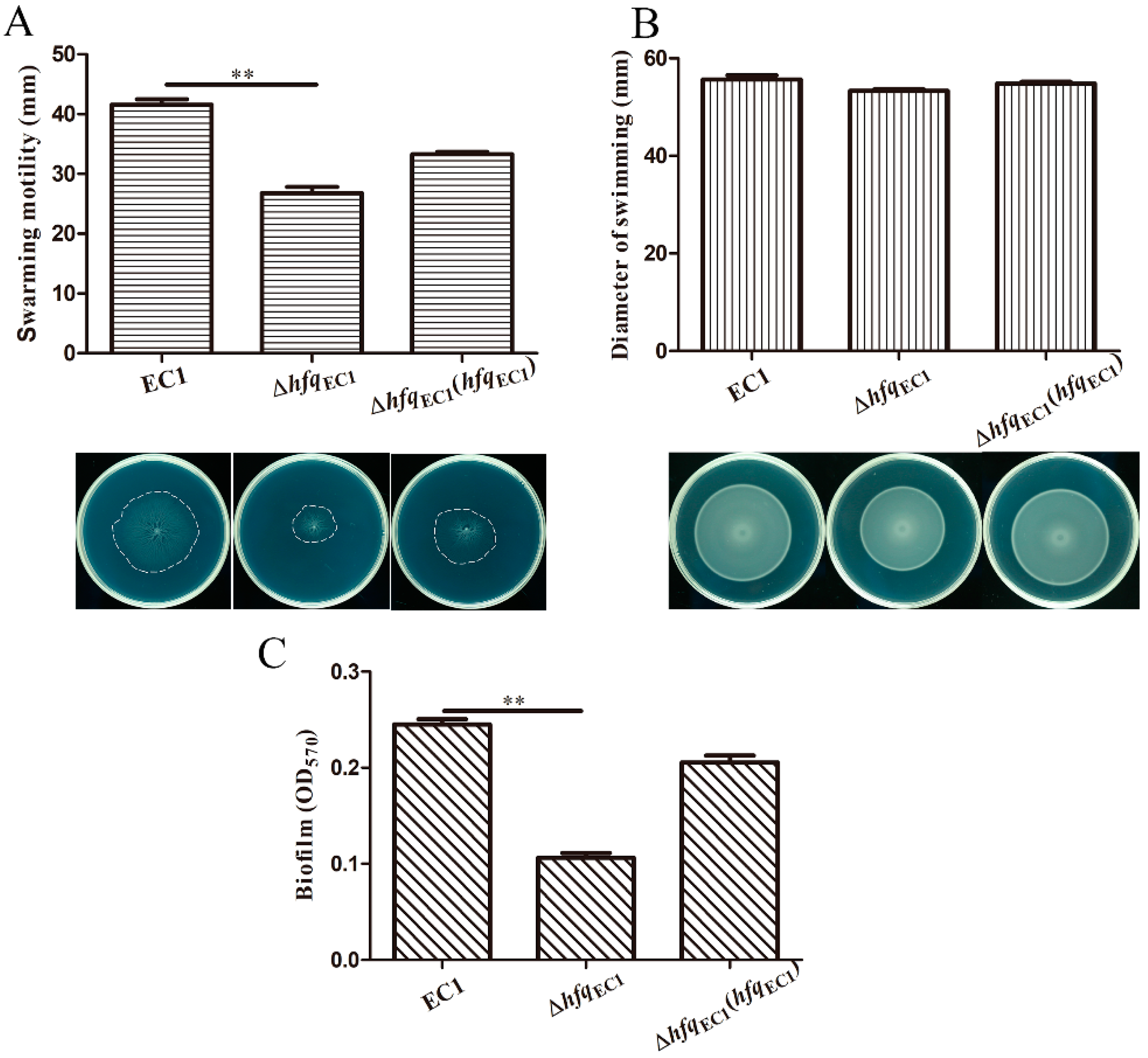
3.3. HfqEC1 Contributes to Swarming Motility and Biofilm Formation in D. oryzae EC1
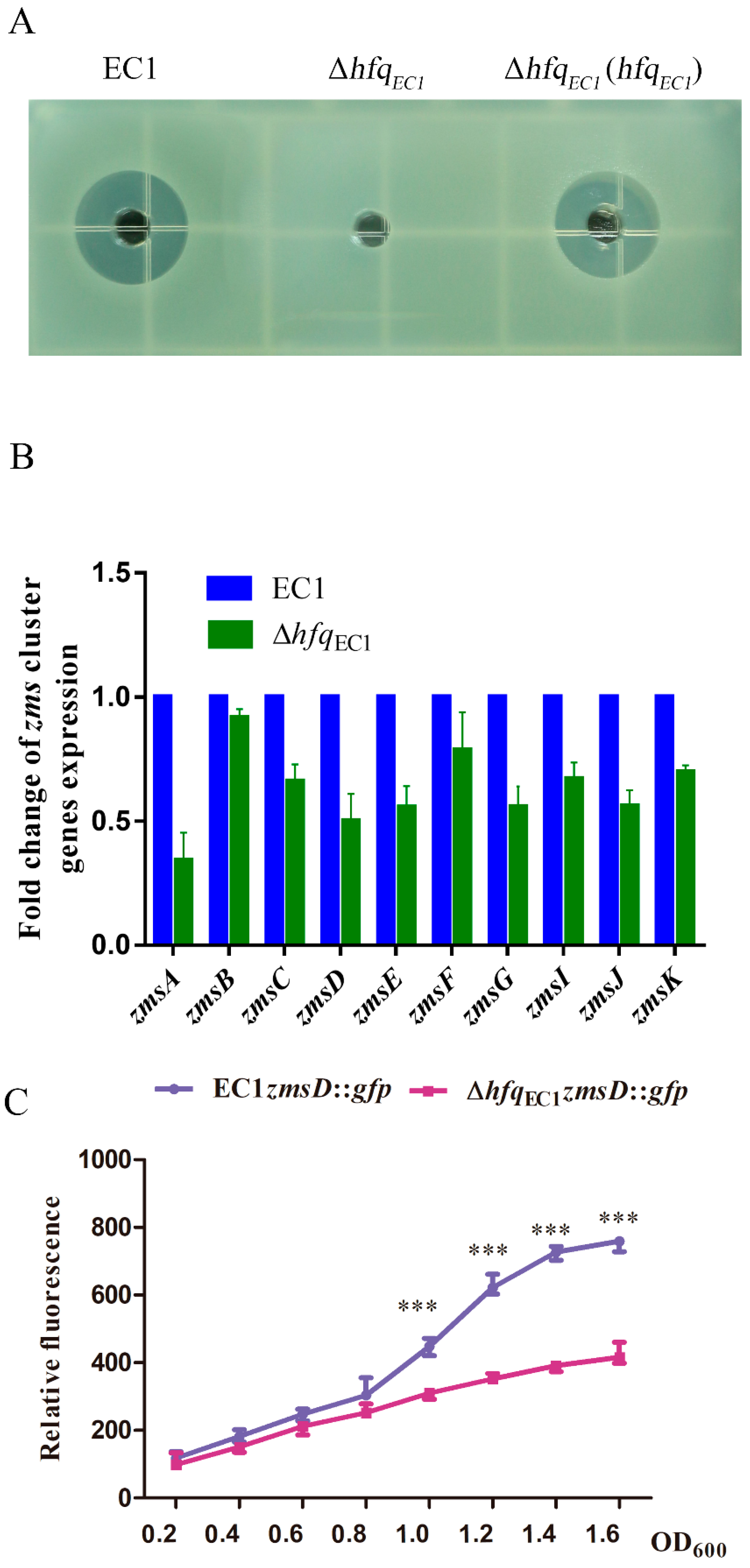
3.4. HfqEC1 Regulates Zeamine Production through Modulating the Expression of Key Zms Cluster Genes
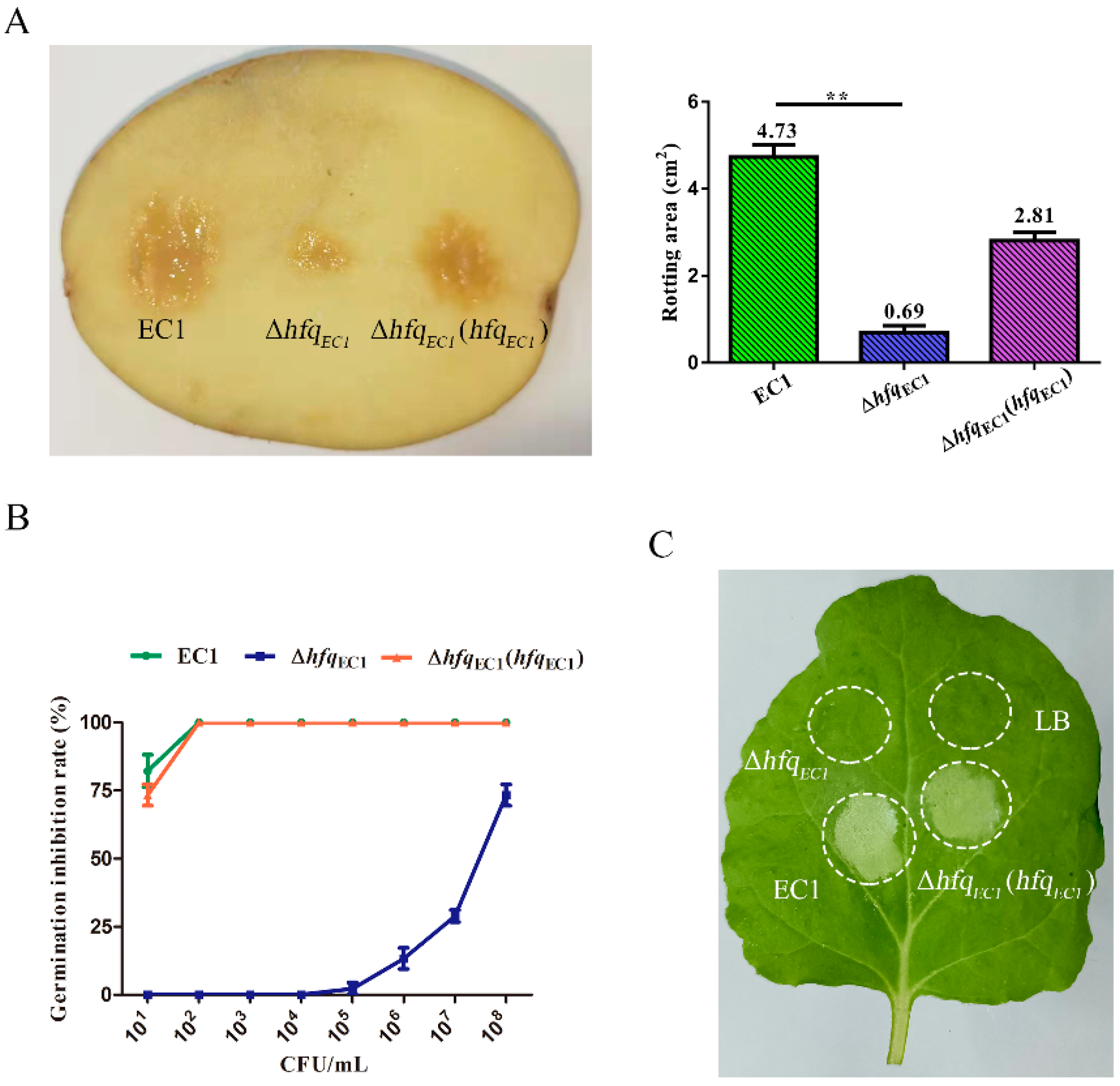
3.5. HfqEC1 Is Required for the Virulence of D. oryzae EC1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reverchon, S.; Nasser, W. Dickeya Ecology, Environment Sensing and Regulation of Virulence Programme. Environ. Microbiol. Rep. 2013, 5, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.B.; Zhang, H.B.; Xu, J.L.; Liu, Q.; Jiang, Z.; Zhang, L.H. The Acyl-Homoserine Lactone-Type Quorum-Sensing System Modulates Cell Motility and Virulence of Erwinia chrysanthemi pv. zeae. J. Bacteriol. 2008, 190, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, M. Bacterial Foot Rot of Rice Caused by a Strain of Erwinia chrysanthemi. Phytopathology 1979, 69, 213. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.G.; Zhang, Q.; Wei, C.D. Advances in Research of Rice Bacterial Foot Rot. Sci. Agric. Sin. 2013, 46, 2923–2931. [Google Scholar]
- Pritchard, L.; Humphris, S.; Saddler, G.S.; Elphinstone, J.G.; Pirhonen, M.; Toth, I.K. Draft Genome Sequences of 17 Isolates of the Plant Pathogenic Bacterium Dickeya. Genome Announc. 2013, 1, e00978-13. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Liu, X.; An, S.; Chang, C.; Zou, Y.; Huang, L.; Zhong, J.; Liu, Q.; Jiang, Z.; Zhou, J.; et al. A Nonribosomal Peptide Synthase Containing a Stand-Alone Condensation Domain Is Essential for Phytotoxin Zeamine Biosynthesis. Mol. Plant-Microbe Interact. 2013, 26, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, H.; Wu, J.; Liu, Q.; Xi, P.; Lee, J.; Liao, J.; Jiang, Z.; Zhang, L.H. A Novel Multidomain Polyketide Synthase Is Essential for Zeamine Production and the Virulence of Dickeya zeae. Mol. Plant-Microbe Interact. 2011, 24, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Wang, Q.; Li, Y.; Liang, Z.; Xu, L.; Zhou, J.; Cui, Z.; Zhang, L.H. Putrescine Is an Intraspecies and Interkingdom Cell-Cell Communication Signal Modulating the Virulence of Dickeya zeae. Front. Microbiol. 2019, 10, 1950. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.N.; Zhang, H.B.; Lv, M.F.; Chen, Y.F.; Liao, L.S.; Cheng, Y.Y.; Liu, S.Y.; Chen, S.H.; He, F.; Cui, Z.N.; et al. SlyA Regulates Phytotoxin Production and Virulence in Dickeya zeae EC1. Mol. Plant Pathol. 2016, 17, 1398–1408. [Google Scholar] [CrossRef]
- Lv, M.; Chen, Y.; Liao, L.; Liang, Z.; Shi, Z.; Tang, Y.; Ye, S.; Zhou, J.; Zhang, L. Fis Is a Global Regulator Critical for Modulation of Virulence Factor Production and Pathogenicity of Dickeya zeae. Sci. Rep. 2018, 8, 341. [Google Scholar] [CrossRef] [Green Version]
- Lv, M.; Hu, M.; Li, P.; Jiang, Z.; Zhang, L.H.; Zhou, J. A Two-Component Regulatory System VfmIH Modulates Multiple Virulence Traits in Dickeya zeae. Mol. Microbiol. 2019, 111, 1493–1509. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Y.; Zhu, M.; Lv, M.; Liu, Z.; Chen, Z.; Huang, Y.; Gu, W.; Liang, Z.; Chang, C.; et al. The GacA-GacS Type Two-Component System Modulates the Pathogenicity of Dickeya oryzae EC1 Mainly by Regulating the Production of Zeamines. Mol. Plant. Microbe. Interact. 2022, 35, 369–379. [Google Scholar] [CrossRef]
- Chen, Y.; Lv, M.; Liao, L.; Gu, Y.; Liang, Z.; Shi, Z.; Liu, S.; Zhou, J.; Zhang, L. Genetic Modulation of c-di-GMP Turnover Affects Multiple Virulence Traits and Bacterial Virulence in Rice Pathogen Dickeya zeae. PLoS ONE 2016, 11, e0165979. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, J.; Lv, M.; Liang, Z.; Parsek, M.R.; Zhang, L.H. Systematic Analysis of c-di-GMP Signaling Mechanisms and Biological Functions in Dickeya zeae EC1. MBio 2020, 11, e02993-20. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.G.H.; Romby, P. Small RNAs in Bacteria and Archaea: Who They Are, What They Do, and How They Do It. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [CrossRef]
- Waters, L.S.; Storz, G. Regulatory RNAs in Bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franze de Fernandez, M.T.; Eoyang, L.; August, J.T. Factor Fraction Required for the Synthesis of Bacteriophage Qbeta-RNA. Nature 1968, 219, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Updegrove, T.B.; Zhang, A.; Storz, G. 2016 Hfq: The Flexible RNA Hitchmaker. Physiol. Behav. 2016, 176, 100–106. [Google Scholar] [CrossRef]
- Feliciano, J.R.; Grilo, A.M.; Guerreiro, S.I.; Sousa, S.A.; Leitão, J.H. Hfq: A Multifaceted RNA Chaperone Involved in Virulence. Future Microbiol. 2016, 11, 137–151. [Google Scholar] [CrossRef]
- Chao, Y.; Vogel, J. The Role of Hfq in Bacterial Pathogens. Curr. Opin. Microbiol. 2010, 13, 24–33. [Google Scholar] [CrossRef]
- Mizrahi, S.P.; Elbaz, N.; Argaman, L.; Altuvia, Y.; Katsowich, N.; Socol, Y.; Bar, A.; Rosenshine, I.; Margalit, H. The Impact of Hfq-Mediated sRNA-mRNA Interactome on the Virulence of Enteropathogenic Escherichia coli. Sci. Adv. 2021, 7, eabi8228. [Google Scholar] [CrossRef] [PubMed]
- Tsui, H.C.; Leung, H.C.; Winkler, M.E. Characterization of Broadly Pleiotropic Phenotypes Caused by an Hfq Insertion Mutation in Escherichia coli K-12. Mol. Microbiol. 1994, 13, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Davis, B.M.; Waldor, M.K. Hfq Is Essential for Vibrio cholerae Virulence and Downregulates σE Expression. Mol. Microbiol. 2004, 53, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Schiano, C.A.; Bellows, L.E.; Lathem, W.W. The Small RNA Chaperone Hfq Is Required for the Virulence of Yersinia pseudotuberculosis. Infect. Immun. 2010, 78, 2034–2044. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.; Villard, C.; Nasser, W.; Reverchon, S.; Hommais, F. RNA Chaperones Hfq and ProQ Play a Key Role in the Virulence of the Plant Pathogenic Bacterium Dickeya dadantii. Front. Microbiol. 2021, 12, 687484. [Google Scholar] [CrossRef]
- Lai, J.L.; Tang, D.J.; Liang, Y.W.; Zhang, R.; Chen, Q.; Qin, Z.P.; Ming, Z.H.; Tang, J.L. The RNA Chaperone Hfq Is Important for the Virulence, Motility and Stress Tolerance in the Phytopathogen Xanthomonas campestris. Environ. Microbiol. Rep. 2018, 10, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Wilms, I.; Möller, P.; Stock, A.M.; Gurski, R.; Lai, E.M.; Narberhaus, F. Hfq Influences Multiple Transport Systems and Virulence in the Plant Pathogen Agrobacterium tumefaciens. J. Bacteriol. 2012, 194, 5209–5217. [Google Scholar] [CrossRef] [Green Version]
- Shin, G.Y.; Schachterle, J.K.; Shyntum, D.Y.; Moleleki, L.N.; Coutinho, T.A.; Sundin, G.W. Functional Characterization of a Global Virulence Regulator Hfq and Identification of Hfq-Dependent sRNAs in the Plant Pathogen Pantoea ananatis. Front. Microbiol. 2019, 10, 2075. [Google Scholar] [CrossRef]
- Wang, C.; Pu, T.; Lou, W.; Wang, Y.; Gao, Z.; Hu, B.; Fan, J. Hfq, a RNA Chaperone, Contributes to Virulence by Regulating Plant Cell Wall-Degrading Enzyme Production, Type VI Secretion System Expression, Bacterial Competition, and Suppressing Host Defense Response in Pectobacterium carotovorum. Mol. Plant-Microbe Interact. 2018, 31, 1166–1178. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; McNally, R.R.; Sundin, G.W. Global Small RNA Chaperone Hfq and Regulatory Small RNAs Are Important Virulence Regulators in Erwinia amylovora. J. Bacteriol. 2013, 195, 1706–1717. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Zhao, Y.T.; Zhang, J.Q.; Wang, X.J.; Fang, R.X.; Jia, Y.T. Identification and Functional Characterization of Small Non-Coding RNAs in Xanthomonas oryzae pathovar oryzae. BMC Genom. 2011, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidtke, C.; Abendroth, U.; Brock, J.; Serrania, J.; Becker, A.; Bonas, U. Small RNA SX13: A Multifaceted Regulator of Virulence in the Plant Pathogen Xanthomonas. PLoS Pathog. 2013, 9, e1003626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhang, H.B.; Xu, J.L.; Cox, R.J.; Simpson, T.J.; Zhang, L.H. 13C Labeling Reveals Multiple Amination Reactions in the Biosynthesis of a Novel Polyketide Polyamine Antibiotic Zeamine from Dickeya zeae. Chem. Commun. 2010, 46, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Hellberg, J.E.E.U.; Matilla, M.A.; Salmond, G.P.C. The Broad-Spectrum Antibiotic, Zeamine, Kills the Nematode Worm Caenorhabditis elegans. Front. Microbiol. 2015, 6, 137. [Google Scholar] [CrossRef] [Green Version]
- Liao, L.; Zhou, J.; Wang, H.; He, F.; Liu, S.; Jiang, Z.; Chen, S.; Zhang, L.H. Control of Litchi Downy Blight by Zeamines Produced by Dickeya zeae. Sci. Rep. 2015, 5, 15719. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, Y.; Lv, M.; Liao, L.; Chen, Y.; Gu, Y.; Liu, S.; Jiang, Z.; Xiong, Y.; Zhang, L. The Complete Genome Sequence of Dickeya zeae EC1 Reveals Substantial Divergence from Other Dickeya Strains and Species. BMC Genom. 2015, 16, 571. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Huang, L.; He, F.; Zhou, X.; Shi, Z.; Zhou, J.; Chen, Y.; Lv, M.; Chen, Y.; Zhang, L.-H. A Substrate-Activated Efflux Pump, DesABC, Confers Zeamine Resistance to Dickeya zeae. MBio 2019, 10, e00713-19. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, M.G.; Thomason, M.K.; Havelund, J.; Valentin-Hansen, P.; Storz, G. Dual Function of the McaS Small RNA in Controlling Biofilm Formation. Genes Dev. 2013, 27, 1132–1145. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Zeng, Q.; Khokhani, D.; Tian, F.; Severin, G.B.; Waters, C.M.; Xu, J.; Zhou, X.; Sundin, G.W.; Ibekwe, A.M.; et al. A Feed-Forward Signalling Circuit Controls Bacterial Virulence through Linking cyclic di-GMP and Two Mechanistically Distinct sRNAs, ArcZ and RsmB. Environ. Microbiol. 2019, 21, 2755–2771. [Google Scholar] [CrossRef]
- Balint-Kurti, P. The Plant Hypersensitive Response: Concepts, Control and Consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Xiao, Y.; Zhou, J.M. Regulation of the Type III Secretion System in Phytopathogenic Bacteria. Mol. Plant-Microbe Interact. 2006, 19, 1159–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Wang, Q.; Wang, S.; Wang, C.; Zhang, L.-H.; Liang, Z. Hfq Is a Critical Modulator of Pathogenicity of Dickeya oryzae in Rice Seeds and Potato Tubers. Microorganisms 2022, 10, 1031. https://doi.org/10.3390/microorganisms10051031
Shi Z, Wang Q, Wang S, Wang C, Zhang L-H, Liang Z. Hfq Is a Critical Modulator of Pathogenicity of Dickeya oryzae in Rice Seeds and Potato Tubers. Microorganisms. 2022; 10(5):1031. https://doi.org/10.3390/microorganisms10051031
Chicago/Turabian StyleShi, Zurong, Qingwei Wang, Shunchang Wang, Chengrun Wang, Lian-Hui Zhang, and Zhibin Liang. 2022. "Hfq Is a Critical Modulator of Pathogenicity of Dickeya oryzae in Rice Seeds and Potato Tubers" Microorganisms 10, no. 5: 1031. https://doi.org/10.3390/microorganisms10051031
APA StyleShi, Z., Wang, Q., Wang, S., Wang, C., Zhang, L. -H., & Liang, Z. (2022). Hfq Is a Critical Modulator of Pathogenicity of Dickeya oryzae in Rice Seeds and Potato Tubers. Microorganisms, 10(5), 1031. https://doi.org/10.3390/microorganisms10051031