
Heterologous Expression, Purification, and Immunomodulatory Effects of Recombinant Lipoprotein GUDIV-103 Isolated from Ureaplasma diversum

 , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phylogenetic Analysis
2.1.1. Culture Conditions for U. diversum and DNA Extraction
2.1.2. Primer Designing, PCR, and Electrophoresis
2.1.3. PCR Product Purification, Sequencing, and Phylogenetic Tree Construction
2.2. Purification of rGUDIV-103 and Determination of Its Expression
2.2.1. Cloning, Protein Expression, and Solubility Tests
2.2.2. SDS–PAGE and Western Blotting
2.2.3. Protein Extraction and Purification
2.3. Antigenicity and Immunogenicity Assays
2.3.1. Production of Polyclonal Antibodies
2.3.2. Purification of Serum IgG Fractions
2.3.3. Indirect ELISA
2.3.4. Kinetics of Mono-Specific Polyclonal Antibodies
2.3.5. Dot Blotting
2.3.6. LAMP Extraction and Western Blotting
2.4. Immunomodulation
2.4.1. Bovine Peripheral Blood Collection and PBMC Isolation
2.4.2. rGUDIV-103 Incubation with Bovine Cell Cultures
2.4.3. Proliferation Assay
2.4.4. Gene Expression
2.4.5. H2O2 and NO Quantification
2.5. Statistical Analysis
3. Results
3.1. Cloning GUDIV-103 Inserted into pET-28a(+) Expression Vector in E. coli BL21 Star (DE3)
3.2. Expression and Purification of Soluble rGUDIV-103 from E. coli BL21 (DE3) Star
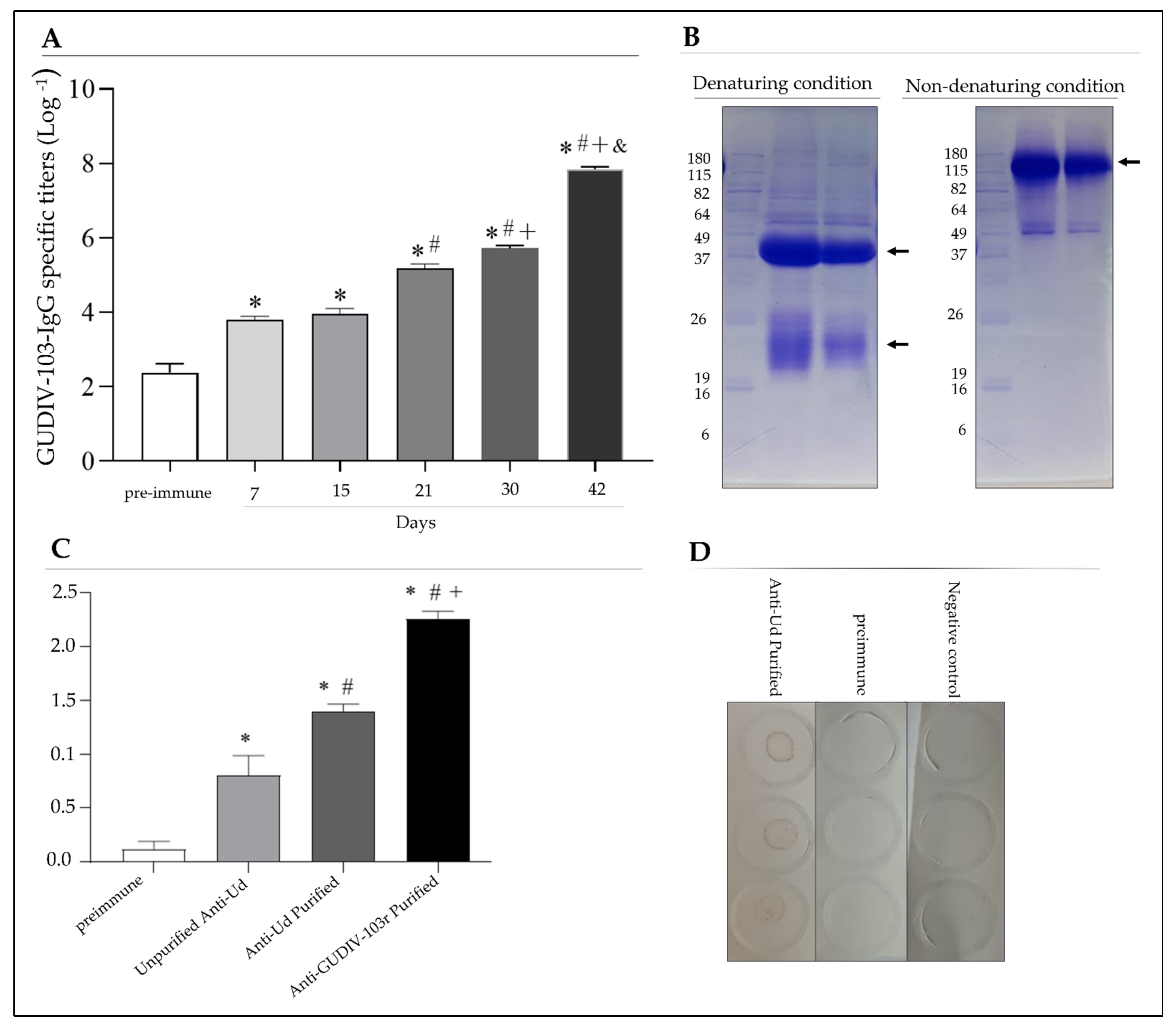
3.3. rGUDIV-103 Is an Immunogenic and Antigenic Protein
3.4. Intraspecific Analysis and Phylogenetic Tree Construction Revealed GUDIV-103 Polymorphism and Genetic Proximity among Strains from the Same Locality
3.5. GUDIV-103 Was Detected in the Total Membrane Protein Extract of U. diversum Strains
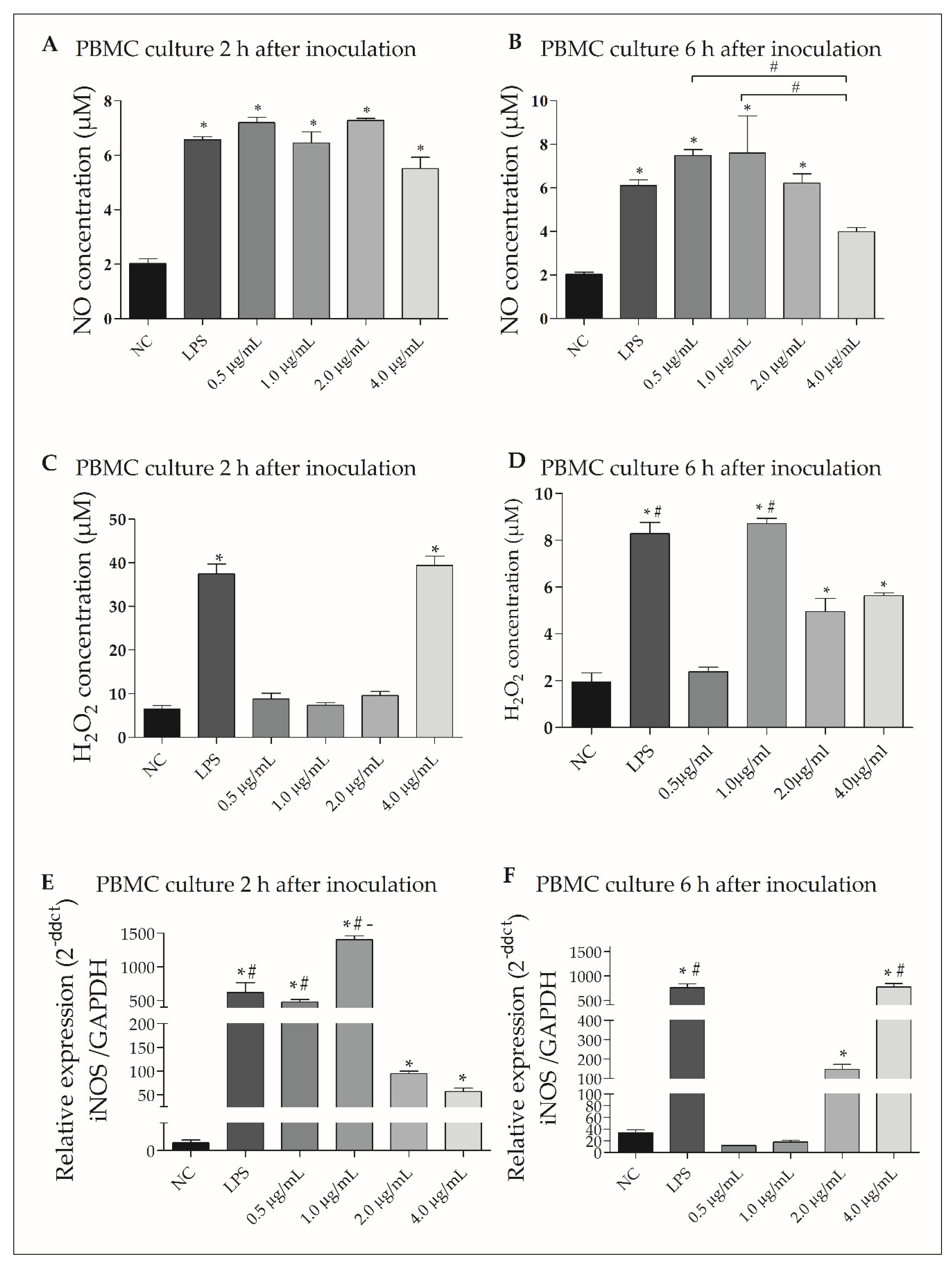
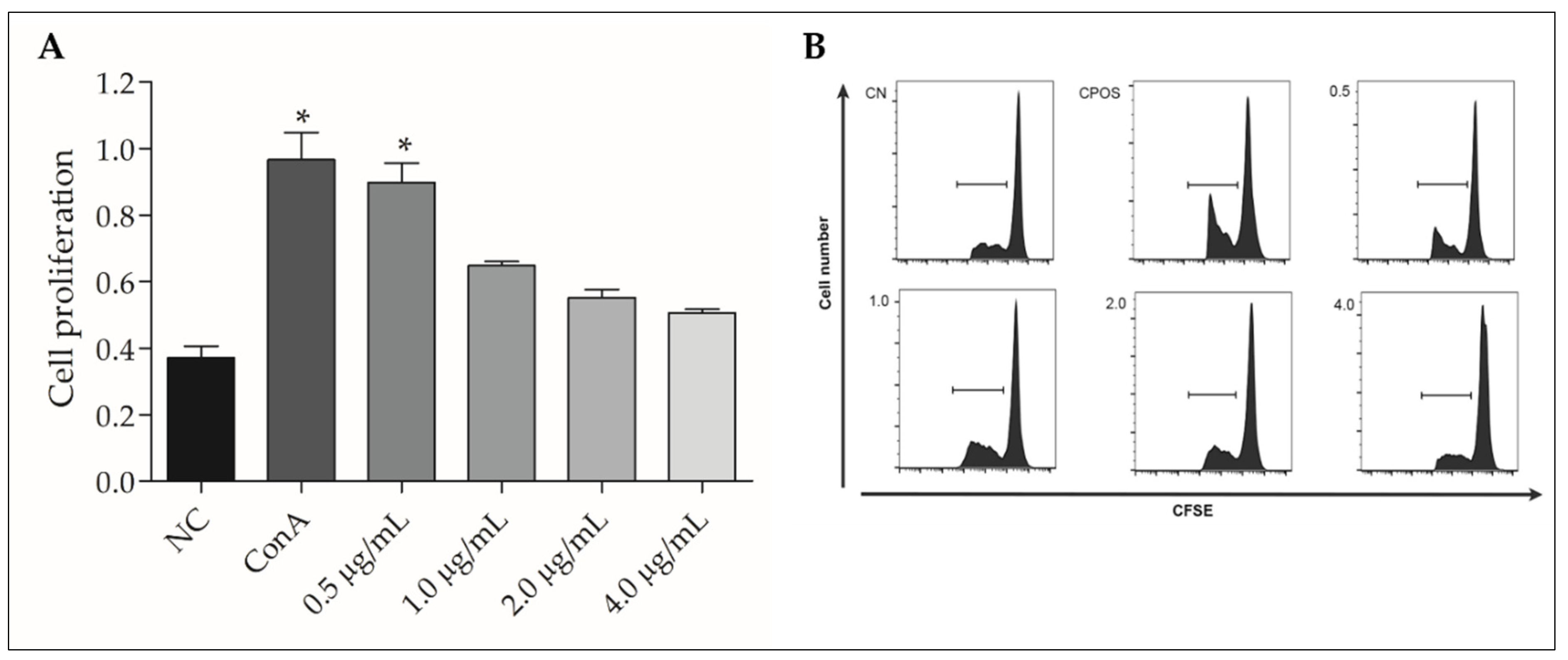
3.6. rGUDIV-103 Induced NO and H2O2 Production in Bovine PBMCs
3.7. rGUDIV-103 Induced Pro-Inflammatory Gene Expression in Bovine PBMCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buzinhani, M.; Yamaguti, M.; Oliveira, R.C.; Cortez, B.A.; Marques, L.M.; Machado-Santelli, G.M.; Assumpção, M.E.; Timenetsky, J. Invasion of Ureaplasma diversum in bovine spermatozoids. BMC Res. Notes 2011, 4, 455. [Google Scholar] [CrossRef]
- Marques, L.; Buzinhani, M.; Neto, R.; Oliveira, R.; Yamaguti, M.; Guimarães, A.; Timenetsky, J. Detection of Ureaplasma diversum in bovine semen straws for artificial insemination. Vet. Rec. 2009, 165, 572. [Google Scholar] [CrossRef]
- Burgher, Y.; Koszegi, M.; St-Sauveur, V.G.; Fournier, D.; Lafond-Lambert, C.; Provost, C.; Gagnon, C.A. Canada: First report of Ureaplasma diversum, a bovine pathogen, in the respiratory tract of swine in Canada. Can. Vet. J. 2018, 59, 1333–1337. [Google Scholar]
- Santos, M.N., Jr.; de Macêdo Neres, N.S.; Campos, G.B.; Bastos, B.L.; Timenetsky, J.; Marques, L.M. A Review of Ureaplasma diversum: A Representative of the Mollicute Class Associated with Reproductive and Respiratory Disorders in Cattle. Front. Vet. Sci. 2021, 8, 572171. [Google Scholar] [CrossRef]
- Marques, L.M.; Guimarães, A.M.; Martins, H.B.; Rezende, I.S.; Barbosa, M.S.; Campos, G.B.; do Nascimento, N.C.; dos Santos, A.P.; Amorim, A.T.; Santos, V.M. Genome sequence of Ureaplasma diversum strain ATCC 49782. Genome Announc. 2015, 3, e00314-15. [Google Scholar] [CrossRef] [Green Version]
- Marques, L.M.; Rezende, I.S.; Barbosa, M.S.; Guimarães, A.M.; Martins, H.B.; Campos, G.B.; do Nascimento, N.C.; Dos Santos, A.P.; Amorim, A.T.; Santos, V.M.; et al. Ureaplasma diversum Genome Provides New Insights about the Interaction of the Surface Molecules of This Bacterium with the Host. PLoS ONE 2016, 11, e0161926. [Google Scholar] [CrossRef]
- Howard, C.; Gourlay, R. Proposal for a second species within the genus Ureaplasma, Ureaplasma diversum sp. nov. Int. J. Syst. Evol. Microbiol. 1982, 32, 446–452. [Google Scholar] [CrossRef]
- Dando, S.J.; Sweeney, E.L.; Knox, C.L. Ureaplasma. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; John Wiley & Sons, Inc.: London, UK, 2019. [Google Scholar] [CrossRef]
- Razin, S. Molecular properties of mollicutes: A synopsis. In Molecular and Diagnostic Procedures in Mycoplasmology; Academic Press: London, UK, 1995; Volume 1, pp. 1–25. [Google Scholar]
- Baseman, J.B.; Tully, J.G. Mycoplasmas: Sophisticated, reemerging, and burdened by their notoriety. Emerg. Infect. Dis. 1997, 3, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Argue, B.; Chousalkar, K.K.; Chenoweth, P.J. Presence of Ureaplasma diversum in the Australian cattle population. Aust. Vet. J. 2013, 91, 99–101. [Google Scholar] [CrossRef]
- Marques, L.M.; Amorim, A.T.; Martins, H.B.; Rezende, I.S.; Barbosa, M.S.; Lobão, T.N.; Campos, G.B.; Timenetsky, J. A quantitative TaqMan PCR assay for the detection of Ureaplasma diversum. Vet. Microbiol. 2013, 167, 670–674. [Google Scholar] [CrossRef]
- Szacawa, E.; Jawor, P.; Dudek, K.; Bednarek, D.; Stefaniak, T. Screening for Mollicutes microorganisms in perinatal calf mortality cases in Polish dairy herds. Pol. J. Vet. Sci. 2018, 21, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.V.; Scarcelli, E.; Grasso, L.M.; Teixeira, S.R.; Genovez, M.E. Ureaplasma diversum and reproductive disorder in Brazilian cows and heifers; first report. Anim. Reprod. Sci. 2000, 63, 137–143. [Google Scholar] [CrossRef]
- Vasconcellos Cardoso, M.; Blanchard, A.; Ferris, S.; Verlengia, R.; Timenetsky, J.; Florio Da Cunha, R.A. Detection of Ureaplasma diversum in cattle using a newly developed PCR-based detection assay. Vet. Microbiol. 2000, 72, 241–250. [Google Scholar] [CrossRef]
- Azevedo, J.B.; Silva, G.S.; Rocha, P.S.; Pitchenin, L.C.; Dutra, V.; Nakazato, L.; de Oliveira, A.C.; Pescador, C.A. Presence of Ureaplasma diversum in the genital tracts of female dairy cattle in Mato Grosso State, Brazil. Trop. Anim. Health Prod. 2017, 49, 311–316. [Google Scholar] [CrossRef]
- Lobo, E.; Ramírez, A.; Poveda, J. Detección de Ureaplasma spp. en pulmones de cerdos, con lesiones compatibles a la neumonía enzoótica porcina, procedentes de la región occidental y central de Cuba. Rev. Salud Anim. 2013, 35, 69–71. [Google Scholar]
- Taylor-Robinson, D.; Haig, D.A.; Williams, M.H. Bovine T-strain mycoplasma. Ann. N. Y. Acad. Sci. 1967, 143, 517–518. [Google Scholar] [CrossRef]
- Ball, H.J.; Mackie, D.P. The ovine mammary gland as an experimental model to determine the virulence of animal ureaplasmas. J. Hyg. 1985, 95, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Doig, P.A.; Ruhnke, H.L.; Palmer, N.C. Experimental bovine genital ureaplasmosis. I. Granular vulvitis following vulvar inoculation. Can. J. Comp. Med. 1980, 44, 252–258. [Google Scholar]
- Pôrto, R.; Oliveira, B.; Ferraz, H.; Caixeta, L.; Viu, M.A.; Gambarini, M.L. Histopatology of the reproductive tract of Nellore pubertal heifers with genital ureaplasmosis. Acad. Bras. Cienc. 2017, 89, 2987–2996. [Google Scholar] [CrossRef] [Green Version]
- Barkallah, M.; Gharbi, Y.; Hassena, A.B.; Slima, A.B.; Mallek, Z.; Gautier, M.; Greub, G.; Gdoura, R.; Fendri, I. Survey of infectious etiologies of bovine abortion during mid- to late gestation in dairy herds. PLoS ONE 2014, 9, e91549. [Google Scholar] [CrossRef] [Green Version]
- Eaglesome, M.; Garcia, M.; Stewart, R. Microbial agents associated with bovine genital tract infections and semen. Part II. Haemophilus somnus, Mycoplasma spp. and Ureaplasma spp., Chlamydia; pathogens and semen contaminants; treatments of bull semen with antimicrobial agents. Vet. Bull. 1992, 62, 887–910. [Google Scholar]
- Miller, R.B.; Ruhnke, H.L.; Doig, P.A.; Poitras, B.J.; Palmer, N.C. The effects of Ureplasma diversum inoculated into the amniotic cavity in cows. Theriogenology 1983, 20, 367–374. [Google Scholar] [CrossRef]
- Howard, C.J.; Gourlay, R.N.; Thomas, L.H.; Stott, E.J. Induction of pneumonia in gnotobiotic calves following inoculation of Mycoplasma dispar and ureaplasmas (T-mycoplasmas). Res. Vet. Sci. 1976, 21, 227–231. [Google Scholar] [CrossRef]
- Ter Laak, E.A.; Noordergraaf, J.H.; Dieltjes, R.P. Prevalence of mycoplasmas in the respiratory tracts of pneumonic calves. J. Vet. Med. Ser. B 1992, 39, 553–562. [Google Scholar] [CrossRef]
- Hobson, N.; Chousalkar, K.K.; Chenoweth, P.J. Ureaplasma diversum in bull semen in Australia: Its detection and potential effects. Aust. Vet. J. 2013, 91, 469–473. [Google Scholar] [CrossRef]
- Crane, M.B.; Hughes, C.A. Can Ureaplasma diversum be transmitted from donor to recipient through the embryo? Two case reports outlining U. diversum losses in bovine embryo pregnancies. Can. Vet. J. 2018, 59, 43. [Google Scholar]
- Santos, M.N., Jr.; Rezende, I.S.; Souza, C.L.S.; Barbosa, M.S.; Campos, G.B.; Brito, L.F.; Queiroz, É.; Barbosa, E.N.; Teixeira, M.M.; Da Silva, L.O.; et al. Its Membrane-Associated Lipoproteins Activate Inflammatory Genes Through the NF-κB Pathway via Toll-Like Receptor 4. Front. Microbiol. 2018, 9, 1538. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.N., Jr.; Santos, R.S.; Neves, W.S.; Fernandes, J.M.; de Brito Guimarães, B.C.; Barbosa, M.S.; Silva, L.S.C.; Gomes, C.P.; Rezende, I.S.; Oliveira, C.N.T.; et al. Immunoinformatics and analysis of antigen distribution of Ureaplasma diversum strains isolated from different Brazilian states. BMC Vet. Res. 2020, 16, 379. [Google Scholar] [CrossRef]
- Silva, J.R.; Ferreira, L.F.; Oliveira, P.V.; Nunes, I.V.; Pereira, Í.S.; Timenetsky, J.; Marques, L.M.; Figueiredo, T.B.; Silva, R.A. Intra-uterine experimental infection by Ureaplasma diversum induces TNF-α mediated womb inflammation in mice. An. Acad. Bras. Cienc. 2016, 88, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Ki, M.-R.; Pack, S.P. Fusion tags to enhance heterologous protein expression. Appl. Microbiol. Biotechnol. 2020, 104, 2411–2425. [Google Scholar] [CrossRef]
- Huang, X.; Qiao, Y.; Zhou, Y.; Ruan, Z.; Kong, Y.; Li, G.; Xie, X.; Zhang, J. Ureaplasma spp. lipid-associated membrane proteins induce human monocyte U937 cell cycle arrest through p53-independent p21 pathway. Int. J. Med. Microbiol. 2018, 308, 819–828. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Li, Y.; Wang, Q.; Shao, J.; Chen, Y.; Xin, J. Mycoplasma bovis-derived lipid-associated membrane proteins activate IL-1β production through the NF-κB pathway via toll-like receptor 2 and MyD88. Dev. Comp. Immunol. 2016, 55, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Chen, Y.; Wang, Y.; Li, Y.; Zhang, L.; Xin, J. TLR2/MyD88/NF-κB signaling pathway regulates IL-1β production in DF-1 cells exposed to Mycoplasma gallisepticum LAMPs. Microb. Pathog. 2018, 117, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Futo, S.; Seto, Y.; Okada, M.; Sato, S.; Suzuki, T.; Kawai, K.; Imada, Y.; Mori, Y. Recombinant 46-kilodalton surface antigen (P46) of Mycoplasma hyopneumoniae expressed in Escherichia coli can be used for early specific diagnosis of mycoplasmal pneumonia of swine by enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1995, 33, 680–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, M.S.; Alves, R.P.D.S.; Rezende, I.S.; Pereira, S.S.; Campos, G.B.; Freitas, L.M.; Chopra-Dewasthaly, R.; Ferreira, L.C.S.; Guimarães, A.M.S.; Marques, L.M.; et al. Novel antigenic proteins of Mycoplasma agalactiae as potential vaccine and serodiagnostic candidates. Vet. Microbiol. 2020, 251, 108866. [Google Scholar] [CrossRef] [PubMed]
- Wawegama, N.K.; Browning, G.F.; Kanci, A.; Marenda, M.S.; Markham, P.F. Development of a recombinant protein-based enzyme-linked immunosorbent assay for diagnosis of Mycoplasma bovis infection in cattle. Clin. Vaccine Immunol. 2014, 21, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhnke, H.; Doig, P.; MacKay, A.; Gagnon, A.; Kierstead, M. Isolation of Ureaplasma from bovine granular vulvitis. Can. J. Comp. Med. 1978, 42, 151. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Lasaro, M.A.; Mathias-Santos, C.; Rodrigues, J.F.; Ferreira, L.C. Functional and immunological characterization of a natural polymorphic variant of a heat-labile toxin (LT-I) produced by enterotoxigenic Escherichia coli (ETEC). FEMS Immunol. Med. Microbiol. 2009, 55, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Rawadi, G.; Roman-Roman, S. Mycoplasma membrane lipoproteins induced proinflammatory cytokines by a mechanism distinct from that of lipopolysaccharide. Infect. Immun. 1996, 64, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Reichelt, P.; Schwarz, C.; Donzeau, M. Single step protocol to purify recombinant proteins with low endotoxin contents. Protein Expr. Purif. 2006, 46, 483–488. [Google Scholar] [CrossRef]
- Garcia, M.; Elsasser, T.H.; Biswas, D.; Moyes, K.M. The effect of citrus-derived oil on bovine blood neutrophil function and gene expression in vitro. J. Dairy Sci. 2015, 98, 918–926. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, D.; Kadanga, A.; Rings, F.; Bauch, K.; Jennen, D.; Nganvongpanit, K.; Hölker, M.; Tholen, E.; Ponsuksili, S.; Wimmers, K.; et al. The effect of nitric oxide inhibition and temporal expression patterns of the mRNA and protein products of nitric oxide synthase genes during in vitro development of bovine pre-implantation embryos. Reprod. Domest. Anim. 2006, 41, 501–509. [Google Scholar] [CrossRef]
- Meça, K.; Martins, A.; Del Puerto, H.; Vasconcelos, A. Apoptose e expressão de Bcl-2 e das caspases 3 e 8 em placenta bovina, em diferentes estádios de gestação. Arq. Bras. Med. Vet. E Zootec. 2010, 62, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tsikas, D. Analysis of nitrite and nitrate in biological fluids by assays based on the Griess reaction: Appraisal of the Griess reaction in the L-arginine/nitric oxide area of research. J. Chromatogr. B 2007, 851, 51–70. [Google Scholar] [CrossRef]
- Howard, C.; Gourlay, R.; COLLINS, J. Serological comparison between twenty-five bovine Ureaplasma (T-mycoplasma) strains by immunofluorescence. Int. J. Syst. Evol. Microbiol. 1975, 25, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Doig, P.A.; Ruhnke, H.L.; Palmer, N.C. Experimental bovine genital ureaplasmosis. II. Granular vulvitis, endometritis and salpingitis following uterine inoculation. Can. J. Comp. Med. 1980, 44, 259–266. [Google Scholar]
- Ruhnke, H.L.; Palmer, N.C.; Doig, P.A.; Miller, R.B. Bovine abortion and neonatal death associated with Ureaplasma diversum. Theriogenology 1984, 21, 295–301. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [Green Version]
- Adrio, J.-L.; Demain, A.L. Recombinant organisms for production of industrial products. Bioeng. Bugs 2010, 1, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.H.; Song, J.; Tey, B.T.; Ramanan, R.N. Bioinformatics approaches for improved recombinant protein production in Escherichia coli: Protein solubility prediction. Brief. Bioinform. 2014, 15, 953–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, M.K.; Amin, A.A.; Fulton, K.F.; Fernando, T.; Kamau, L.; Batty, C.; Louca, M.; Ho, S.; Whisstock, J.C.; Bottomley, S.P. The REFOLD database: A tool for the optimization of protein expression and refolding. Nucleic. Acids Res. 2006, 34, D207–D212. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.M.; Panda, A.K. Solubilization and refolding of bacterial inclusion body proteins. J. Biosci. Bioeng. 2005, 99, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.D.; Elisee, C.; Newham, D.M.; Harrison, R.G. New fusion protein systems designed to give soluble expression in Escherichia coli. Biotechnol. Bioeng. 1999, 65, 382–388. [Google Scholar] [CrossRef]
- Ogata, M.; Kotani, H.; Yamamoto, K. Serological comparison of bovine ureaplasmas. Nihon Juigaku Zasshi 1979, 41, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Howard, C.; Gourlay, R.; Collins, J. Serological studies with bovine ureaplasmas (T-mycoplasmas). Int. J. Syst. Evol. Microbiol. 1978, 28, 473–477. [Google Scholar] [CrossRef]
- Koshimizu, K.; Kotani, H.; Yamamoto, K.; Magaribuchi, T.; Harasawa, R.; Ito, M.; Ogata, M. Serological analysis of Ureaplasmas isolated from various animals. Isr. J. Med. Sci. 1984, 20, 950–953. [Google Scholar]
- Li, P.; Zhang, Y.; Li, X.; Zhou, W.; Li, X.; Jiang, F.; Wu, W. Mycoplasma hyopneumoniae Mhp597 is a cytotoxicity, inflammation and immunosuppression associated nuclease. Vet. Microbiol. 2019, 235, 53–62. [Google Scholar] [CrossRef]
- Zhao, G.; Zhu, X.; Zhang, H.; Chen, Y.; Schieck, E.; Hu, C.; Chen, H.; Guo, A. Novel secreted protein of Mycoplasma bovis MbovP280 induces macrophage apoptosis through CRYAB. Front. Immunol. 2021, 12, 309. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Guo, F.; Lei, A.; Xiang, J.; Liu, P.; Ten, W.; Dai, G.; Li, R. GrpE Immunization Protects Against Ureaplasma urealyticum Infection in BALB/C Mice. Front. Immunol. 2020, 11, 1495. [Google Scholar] [CrossRef] [PubMed]
- Howard, C.J.; Gourlay, R.N. Identification of Ureaplasmas from Cattle Using Antisera Prepared in Gnotobiotic Calves. Microbiology 1981, 126, 365–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulira, G.L.; Saunders, J.R.; Barth, A.D. Isolation of Ureaplasma diversum and mycoplasmas from genital tracts of beef and dairy cattle in Saskatchewan. Can. Vet. J. 1992, 33, 46–49. [Google Scholar]
- Christodoulides, A.; Gupta, N.; Yacoubian, V.; Maithel, N.; Parker, J.; Kelesidis, T. The Role of Lipoproteins in Mycoplasma-Mediated Immunomodulation. Front. Microbiol. 2018, 9, 1682. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.; Zhang, B.; Wang, J.; Ni, B.; Ji, Y.; Wei, Y.; Xiao, S.; Feng, Z.; Liu, M.; Shao, G. Characterization of the role in adherence of Mycoplasma hyorhinis variable lipoproteins containing different repeat unit copy numbers. Vet. Microbiol. 2016, 197, 39–46. [Google Scholar] [CrossRef]
- Benčina, D.; Drobnič-Valič, M.; Horvat, S.; Narat, M.; Kleven, S.H.; Dovč, P. Molecular basis of the length variation in the N-terminal part of Mycoplasma synoviae hemagglutinin. FEMS Microbiol. Lett. 2001, 203, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.T.; Thorsen, P.; MØsller, B.; Birkelund, S.; Christiansen, G. Antigenic and genomic homogeneity of successive Mycoplasma hominis isolates. J. Med. Microbiol. 1998, 47, 659–666. [Google Scholar] [CrossRef]
- Marques, L.M.; Buzinhani, M.; Guimaraes, A.M.; Marques, R.C.; Farias, S.T.; Neto, R.L.; Yamaguti, M.; Oliveira, R.C.; Timenetsky, J. Intraspecific sequence variation in 16S rRNA gene of Ureaplasma diversum isolates. Vet. Microbiol. 2011, 152, 205–211. [Google Scholar] [CrossRef]
- Sweeney, E.L.; Kallapur, S.G.; Meawad, S.; Gisslen, T.; Stephenson, S.-A.; Jobe, A.H.; Knox, C.L. Ureaplasma Species Multiple Banded Antigen (MBA) Variation Is Associated with the Severity of Inflammation In Vivo and In Vitro in Human Placentae. Front. Cell. Infect. Microbiol. 2017, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Monara, Y.A.; Santos, M.N., Jr.; Campos, G.; Silva, I.B.; Barbosa, M.; Rezende, I.; Queiroz, E.; Timenetsky, J.; Bastos, B.; Amorim, A.; et al. Multilocus sequence typing characterizes diversity of Ureaplasma diversum strains and intra-species variability induces different profiles of immune response profiles. BMC Vet. Res. 2019, 16, 163. [Google Scholar]
- Hwang, M.-H.; Damte, D.; Lee, J.-S.; Gebru, E.; Chang, Z.-Q.; Cheng, H.; Jung, B.-Y.; Rhee, M.-H.; Park, S.-C. Mycoplasma hyopneumoniae induces pro-inflammatory cytokine and nitric oxide production through NFκB and MAPK pathways in RAW264. 7 cells. Vet. Res. Commun. 2011, 35, 21–34. [Google Scholar] [CrossRef]
- Pôrto, R.N.G.; Junqueira-Kipni, A.P.; Gambarini, M.L. Evaluation of in vitro activation of bovine endometrial and vaginal epithelial and blood mononuclear cells to produce nitric oxide in response to Mycoplasma bovis, Mycoplasma bovigenitalium and Ureaplasma diversum. Acta Vet.-Beogr. 2021, 71, 137–146. [Google Scholar] [CrossRef]
- Chelmonska-Soyta, A.; Miller, R.B.; Ruhnke, L.; Rosendal, S. Activation of murine macrophages and lymphocytes by Ureaplasma diversum. Can. J. Vet. Res. 1994, 58, 275–280. [Google Scholar]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef]
- Bai, F.; Ni, B.; Liu, M.; Feng, Z.; Xiong, Q.; Xiao, S.; Shao, G. Mycoplasma hyopneumoniae-derived lipid-associated membrane proteins induce apoptosis in porcine alveolar macrophage via increasing nitric oxide production, oxidative stress, and caspase-3 activation. Vet. Immunol. Immunopathol. 2013, 155, 155–161. [Google Scholar] [CrossRef]
- Dusanic, D.; Bencina, D.; Oven, I.; Cizelj, I.; Bencina, M.; Narat, M. Mycoplasma synoviae induces upregulation of apoptotic genes, secretion of nitric oxide and appearance of an apoptotic phenotype in infected chicken chondrocytes. Vet. Res. 2012, 43, 7. [Google Scholar] [CrossRef] [Green Version]
- Beere, H.M.; Green, D.R. Stress management–heat shock protein-70 and the regulation of apoptosis. Trends Cell Biol. 2001, 11, 6–10. [Google Scholar] [CrossRef]
- Choi, S.Y.; Lim, J.W.; Shimizu, T.; Kuwano, K.; Kim, J.M.; Kim, H. Reactive oxygen species mediate Jak2/Stat3 activation and IL-8 expression in pulmonary epithelial cells stimulated with lipid-associated membrane proteins from Mycoplasma pneumoniae. Inflamm. Res. 2012, 61, 493–501. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, Z.; Xue, D.; Deng, G.; Li, M.; Liu, X.; Wang, Y. Mycoplasma ovipneumoniae induces sheep airway epithelial cell apoptosis through an ERK signalling-mediated mitochondria pathway. BMC Microbiol. 2016, 16, 222. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.; Li, Y.; Jiang, Z.; Deng, G.; Li, M.; Liu, X.; Wang, Y. A ROS-dependent and Caspase-3-mediated apoptosis in sheep bronchial epithelial cells in response to Mycoplasma ovipneumoniae infections. Vet. Immunol. Immunopathol. 2017, 187, 55–63. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, D.; Yuan, F.; Liu, Z.; Duan, Z.; Yang, K.; Guo, R.; Li, M.; Li, S.; Fang, L.; et al. Surface proteins mhp390 (P68) contributes to cilium adherence and mediates inflammation and apoptosis in Mycoplasma hyopneumoniae. Microb. Pathog. 2019, 126, 92–100. [Google Scholar] [CrossRef]
- Lu, T.-H.; Su, C.-C.; Chen, Y.-W.; Yang, C.-Y.; Wu, C.-C.; Hung, D.-Z.; Chen, C.-H.; Cheng, P.-W.; Liu, S.-H.; Huang, C.-F. Arsenic induces pancreatic β-cell apoptosis via the oxidative stress-regulated mitochondria-dependent and endoplasmic reticulum stress-triggered signaling pathways. Toxicol. Lett. 2011, 201, 15–26. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, M.; Xu, S.; Khan, M.A.; Shi, Y.; Qu, W.; Gao, J.; Liu, G.; Kastelic, J.P.; Han, B. Mycoplasma bovis-generated reactive oxygen species and induced apoptosis in bovine mammary epithelial cell cultures. J. Dairy Sci. 2020, 103, 10429–10445. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, S.; Long, C.; An, Z.; Xing, X.; Wen, F.; Bao, S. Mycoplasmas bovis P48 induces apoptosis in EBL cells via an endoplasmic reticulum stress-dependent signaling pathway. Vet. Microbiol. 2021, 255, 109013. [Google Scholar] [CrossRef]
- Amorim, A.T.; Marques, L.M.; Santos, A.M.O.G.; Martins, H.B.; Barbosa, M.S.; Rezende, I.S.; Andrade, E.F.; Campos, G.B.; Lobão, T.N.; Cortez, B.A. Apoptosis in HEp-2 cells infected with Ureaplasma diversum. Biol. Res. 2014, 47, 38. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, W.; Ajuwape, A.T.P.; Rosenbusch, R.F. Global suppression of mitogen-activated ovine peripheral blood mononuclear cells by surface protein activity from Mycoplasma ovipneumoniae. Vet. Immunol. Immunopathol. 2010, 136, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Mulongo, M.; Prysliak, T.; Scruten, E.; Napper, S.; Perez-Casal, J. In vitro infection of bovine monocytes with Mycoplasma bovis delays apoptosis and suppresses production of gamma interferon and tumor necrosis factor alpha but not interleukin-10. Infect. Immun. 2014, 82, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Arachchige, S.N.K.; Condello, A.K.; Zhu, L.; Shil, P.K.; Tivendale, K.A.; Underwood, G.J.; Noormohammadi, A.H.; Browning, G.F.; Wawegama, N.K. Effects of immunosuppression on the efficacy of vaccination against Mycoplasma gallisepticum infection in chickens. Vet. Microbiol. 2021, 260, 109182. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brazilian State | U. diversum Strain | PCR | Western Blotting | |
|---|---|---|---|---|
| Hydrophobic Phase (UdLAMPs) | Hydrophilic Phase (Cytosolic Proteins) | |||
| BA | BA78 | + | + | − |
| 34 | + | + | − | |
| 35 | + | + | − | |
| 37 | + | + | − | |
| 47 | + | + | − | |
| 51 | + | + | − | |
| 52 | − | + | − | |
| 54 | + | + | − | |
| 55 | − | + | − | |
| 56 | − | + | − | |
| 78 | − | + | − | |
| 84.2 | − | + | − | |
| 89 | − | + | − | |
| 111 | + | + | − | |
| 133 | + | + | − | |
| 148 | − | + | − | |
| 174 | − | + | − | |
| 198 | − | + | − | |
| 234 | + | + | − | |
| 249 | + | + | − | |
| MS | 805 | + | + | − |
| 9653 | + | + | − | |
| SP | A203 | − | + | − |
| GOTA | + | + | − | |
| S6 | − | + | − | |
| S8 | − | + | − | |
| 5T | − | + | − | |
| 7T | + | + | − | |
| 10T | + | + | − | |
| 13T | + | + | − | |
| 16T | + | + | − | |
| 72.2 | + | + | − | |
| 73 | + | + | − | |
| 84 | + | + | − | |
| 93 | + | + | − | |
| 94 | + | + | − | |
| 95 | + | + | − | |
| 98 | + | + | − | |
| 100 | + | + | − | |
| 59 | + | + | − | |
| # | A523 | − | + | − |
| ATCC49782 | + | + | − | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Junior, M.N.; Neves, W.S.; Santos, R.S.; Almeida, P.P.; Fernandes, J.M.; Guimarães, B.C.d.B.; Barbosa, M.S.; da Silva, L.S.C.; Gomes, C.P.; Sampaio, B.A.; et al. Heterologous Expression, Purification, and Immunomodulatory Effects of Recombinant Lipoprotein GUDIV-103 Isolated from Ureaplasma diversum. Microorganisms 2022, 10, 1032. https://doi.org/10.3390/microorganisms10051032
Santos-Junior MN, Neves WS, Santos RS, Almeida PP, Fernandes JM, Guimarães BCdB, Barbosa MS, da Silva LSC, Gomes CP, Sampaio BA, et al. Heterologous Expression, Purification, and Immunomodulatory Effects of Recombinant Lipoprotein GUDIV-103 Isolated from Ureaplasma diversum. Microorganisms. 2022; 10(5):1032. https://doi.org/10.3390/microorganisms10051032
Chicago/Turabian StyleSantos-Junior, Manoel Neres, Wanderson Souza Neves, Ronaldo Silva Santos, Palloma Porto Almeida, Janaina Marinho Fernandes, Bruna Carolina de Brito Guimarães, Maysa Santos Barbosa, Lucas Santana Coelho da Silva, Camila Pacheco Gomes, Beatriz Almeida Sampaio, and et al. 2022. "Heterologous Expression, Purification, and Immunomodulatory Effects of Recombinant Lipoprotein GUDIV-103 Isolated from Ureaplasma diversum" Microorganisms 10, no. 5: 1032. https://doi.org/10.3390/microorganisms10051032
APA StyleSantos-Junior, M. N., Neves, W. S., Santos, R. S., Almeida, P. P., Fernandes, J. M., Guimarães, B. C. d. B., Barbosa, M. S., da Silva, L. S. C., Gomes, C. P., Sampaio, B. A., Rezende, I. d. S., Correia, T. M. L., Neres, N. S. d. M., Campos, G. B., Bastos, B. L., Timenetsky, J., & Marques, L. M. (2022). Heterologous Expression, Purification, and Immunomodulatory Effects of Recombinant Lipoprotein GUDIV-103 Isolated from Ureaplasma diversum. Microorganisms, 10(5), 1032. https://doi.org/10.3390/microorganisms10051032