
An Unprecedented Number of Cytochrome P450s Are Involved in Secondary Metabolism in Salinispora Species

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species and Database Information
2.2. Genome Data Mining and Identification of P450s
2.3. Assigning Family and Subfamily to P450s
2.4. Phylogenetic Analysis of P450s
2.5. Salinispora Species P450s Profile Heat-Maps
2.6. Identification of P450s Part of smBGCs
2.7. Data Analysis
2.8. Comparative Analysis of P450s and smBGCs Data
3. Results and Discussion
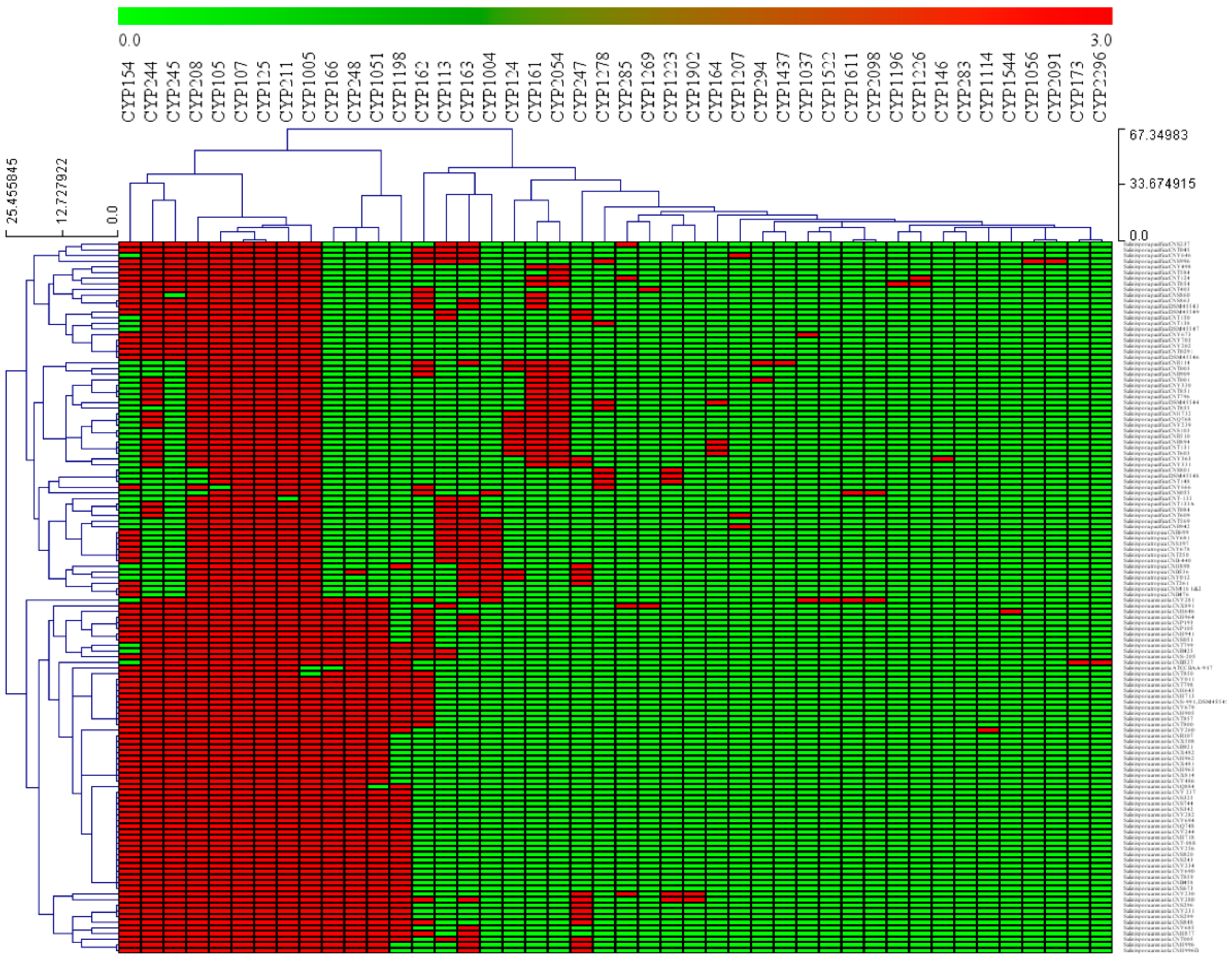
3.1. Salinispora Species P450 Profiles
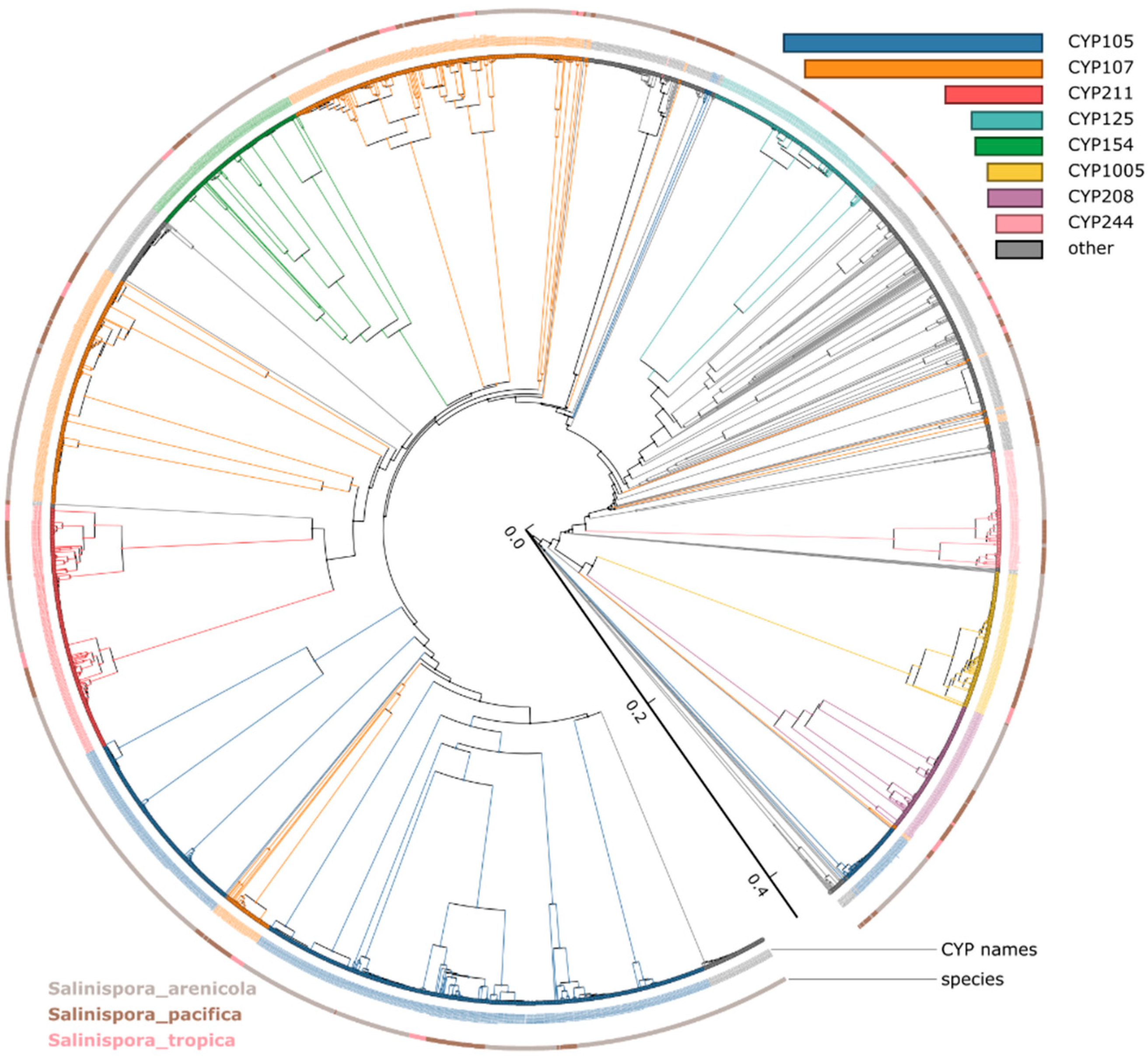
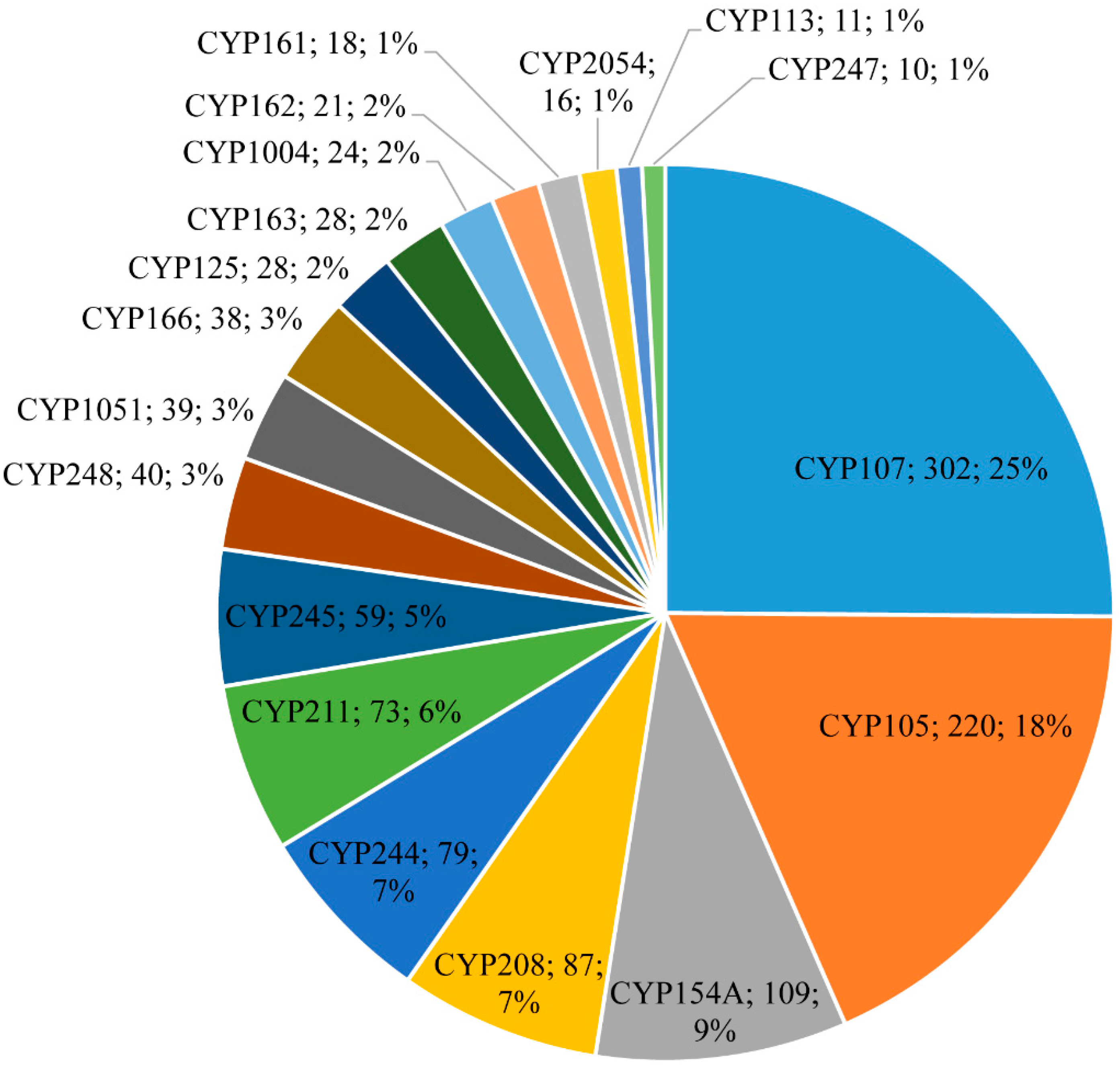
3.2. CYP105 and CYP107 Families Are Bloomed in Salinispora Species
3.3. CYP107 and CYP125 Are Conserved in Salinispora Species
3.4. Unprecedented Number of P450s Involved in smBGCs
3.5. Functional Prediction of Salinispora Species P450s
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Follmer, A.H.; Goldstone, J.V.; Nelson, D.R.; Warrilow, A.G.; Price, C.L.; True, M.Y.; Kelly, S.L.; Poulos, T.L.; Stegeman, J.J. On the occurrence of cytochrome P450 in viruses. Proc. Natl. Acad. Sci. USA 2019, 116, 12343–12352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.E.; Coon, M.J. Oxygen activation by cytochrome P-450. Annu. Rev. Biochem. 1980, 49, 315–356. [Google Scholar] [CrossRef] [PubMed]
- Sono, M.; Roach, M.P.; Coulter, E.D.; Dawson, J.H. Heme-containing oxygenases. Chem. Rev. 1996, 96, 2841–2888. [Google Scholar] [CrossRef]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef]
- Kelly, S.L.; Kelly, D.E. Microbial cytochromes P450: Biodiversity and biotechnology. Where do cytochromes P450 come from, what do they do and what can they do for us? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2013, 368, 20120476. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P.; Munro, A.W. Unusual cytochrome p450 enzymes and reactions. J. Biol. Chem. 2013, 288, 17065–17073. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.C.; Waterman, M.R. Unusual properties of the cytochrome P450 superfamily. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2013, 368, 20120434. [Google Scholar] [CrossRef] [Green Version]
- Girvan, H.M.; Munro, A.W. Applications of microbial cytochrome P450 enzymes in biotechnology and synthetic biology. Curr. Opin. Chem. Biol. 2016, 31, 136–145. [Google Scholar] [CrossRef]
- Urlacher, V.B.; Eiben, S. Cytochrome P450 monooxygenases: Perspectives for synthetic application. Trends Biotechnol. 2006, 24, 324–330. [Google Scholar] [CrossRef]
- Bernhardt, R.; Urlacher, V.B. Cytochromes P450 as promising catalysts for biotechnological application: Chances and limitations. Appl. Microbiol. Biotechnol. 2014, 98, 6185–6203. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, Y.; Guengerich, F.P.; Ma, L.; Li, S.; Zhang, W. Engineering cytochrome P450 enzyme systems for biomedical and biotechnological applications. J. Biol. Chem. 2020, 295, 833–849. [Google Scholar] [CrossRef]
- Guengerich, F.P. A history of the roles of cytochrome P450 enzymes in the toxicity of drugs. Toxicol. Res. 2020, 37, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Esteves, F.; Rueff, J.; Kranendonk, M. The central role of cytochrome P450 in xenobiotic metabolism—A brief review on a fascinating enzyme family. J. Xenobiotics 2021, 11, 94–114. [Google Scholar] [CrossRef] [PubMed]
- Debnath, A.; Calvet, C.M.; Jennings, G.; Zhou, W.; Aksenov, A.; Luth, M.R.; Abagyan, R.; Nes, W.D.; McKerrow, J.H.; Podust, L.M. CYP51 is an essential drug target for the treatment of primary amoebic meningoencephalitis (PAM). PLoS Negl. Trop. Dis. 2017, 11, e0006104. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Friggeri, L.; Waterman, M.R. CYP51 as drug targets for fungi and protozoan parasites: Past, present and future. Parasitology 2018, 145, 1820–1836. [Google Scholar] [CrossRef]
- Jawallapersand, P.; Mashele, S.S.; Kovacic, L.; Stojan, J.; Komel, R.; Pakala, S.B.; Krasevec, N.; Syed, K. Cytochrome P450 monooxygenase CYP53 family in fungi: Comparative structural and evolutionary analysis and its role as a common alternative anti-fungal drug target. PLoS ONE 2014, 9, e107209. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.F.; Tatsuta, K.; Gunji, H.; Ishiyama, T.; Hutchinson, C.R. Substrate specificity of 6-deoxyerythronolide B hydroxylase, a bacterial cytochrome P450 of erythromycin A biosynthesis. Biochemistry 1993, 32, 1905–1913. [Google Scholar] [CrossRef]
- Bischoff, D.; Bister, B.; Bertazzo, M.; Pfeifer, V.; Stegmann, E.; Nicholson, G.J.; Keller, S.; Pelzer, S.; Wohlleben, W.; Süssmuth, R.D. The biosynthesis of vancomycin-type glycopeptide antibiotics—A model for oxidative side-chain cross-linking by oxygenases coupled to the action of peptide synthetases. Chembiochem 2005, 6, 267–272. [Google Scholar] [CrossRef]
- Jennewein, S.; Park, H.; DeJong, J.M.; Long, R.M.; Bollon, A.P.; Croteau, R.B. Coexpression in yeast of Taxus cytochrome P450 reductase with cytochrome P450 oxygenases involved in Taxol biosynthesis. Biotechnol. Bioeng. 2005, 89, 588–598. [Google Scholar] [CrossRef]
- van Beilen, J.B.; Holtackers, R.; Lüscher, D.; Bauer, U.; Witholt, B.; Duetz, W.A. Biocatalytic production of perillyl alcohol from limonene by using a novel Mycobacterium sp. cytochrome P450 alkane hydroxylase expressed in Pseudomonas putida. Appl. Environ. Microbiol. 2005, 71, 1737–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podust, L.M.; Sherman, D.H. Diversity of P450 enzymes in the biosynthesis of natural products. Nat. Prod. Rep. 2012, 29, 1251–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greule, A.; Stok, J.E.; De Voss, J.J.; Cryle, M.J. Unrivalled diversity: The many roles and reactions of bacterial cytochromes P450 in secondary metabolism. Nat. Prod. Rep. 2018, 35, 757–791. [Google Scholar] [CrossRef] [Green Version]
- Vaishnav, P.; Demain, A.L. Unexpected applications of secondary metabolites. Biotechnol. Adv. 2011, 29, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. In History of Modern Biotechnology I; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–39. [Google Scholar]
- Thirumurugan, D.; Cholarajan, A.; Raja, S.S.; Vijayakumar, R. An Introductory Chapter: Secondary Metabolites. In Secondary Metabolites-Sources and Applications; IntechOpen: London, UK, 2018. [Google Scholar]
- Sharma, A.; Kumari, N.; Menghani, E. Bioactive secondary metabolites: An overview. Int. J. Sci. Eng. Res. 2014, 5, 1395. [Google Scholar]
- Abegaz, B.M.; Kinfe, H.H. Secondary metabolites, their structural diversity, bioactivity, and ecological functions: An overview. Phys. Sci. Rev. 2019, 4. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Medema, M.H.; Kottmann, R.; Yilmaz, P.; Cummings, M.; Biggins, J.B.; Blin, K.; de Bruijn, I.; Chooi, Y.H.; Claesen, J.; Coates, R.C.; et al. Minimum Information about a Biosynthetic Gene cluster. Nat. Chem. Biol. 2015, 11, 625–631. [Google Scholar] [CrossRef]
- Weber, T.; Kim, H.U. The secondary metabolite bioinformatics portal: Computational tools to facilitate synthetic biology of secondary metabolite production. Synth. Syst. Biotechnol. 2016, 1, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.; Abraham, J. Natural products from actinobacteria for drug discovery. In Advances in Pharmaceutical Biotechnology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 333–363. [Google Scholar]
- Jose, P.A.; Maharshi, A.; Jha, B. Actinobacteria in natural products research: Progress and prospects. Microbiol. Res. 2021, 246, 126708. [Google Scholar] [CrossRef] [PubMed]
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lima Procópio, R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Bonet, B.; Teufel, R.; Crusemann, M.; Ziemert, N.; Moore, B.S. Direct capture and heterologous expression of Salinispora natural product genes for the biosynthesis of enterocin. J. Nat. Prod. 2015, 78, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef]
- Penn, K.; Jenkins, C.; Nett, M.; Udwary, D.W.; Gontang, E.A.; McGlinchey, R.P.; Foster, B.; Lapidus, A.; Podell, S.; Allen, E.E.; et al. Genomic islands link secondary metabolism to functional adaptation in marine Actinobacteria. ISME J. 2009, 3, 1193–1203. [Google Scholar] [CrossRef]
- Asolkar, R.N.; Kirkland, T.N.; Jensen, P.R.; Fenical, W. Arenimycin, an antibiotic effective against rifampin- and methicillin-resistant Staphylococcus aureus from the marine actinomycete Salinispora arenicola. J. Antibiot. 2010, 63, 37–39. [Google Scholar] [CrossRef] [Green Version]
- Eustaquio, A.S.; Nam, S.J.; Penn, K.; Lechner, A.; Wilson, M.C.; Fenical, W.; Jensen, P.R.; Moore, B.S. The discovery of salinosporamide K from the marine bacterium “Salinispora” pacifica by genome mining gives insight into pathway evolution. Chembiochem 2011, 12, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.R.; Moore, B.S.; Fenical, W. The marine actinomycete genus Salinispora: A model organism for secondary metabolite discovery. Nat. Prod. Rep. 2015, 32, 738–751. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.R.; Williams, P.G.; Oh, D.C.; Zeigler, L.; Fenical, W. Species-specific secondary metabolite production in marine actinomycetes of the genus Salinispora. Appl. Environ. Microbiol. 2007, 73, 1146–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, S.; Adachi, K.; Matsuo, Y.; Nukina, M.; Shizuri, Y. Salinisporamycin, a novel metabolite from Salinispora arenicola. [corrected]. J. Antibiot. 2009, 62, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Ziemert, N.; Lechner, A.; Wietz, M.; Millán-Aguiñaga, N.; Chavarria, K.L.; Jensen, P.R. Diversity and evolution of secondary metabolism in the marine actinomycete genus Salinispora. Proc. Natl. Acad. Sci. USA 2014, 111, E1130–E1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udwary, D.W.; Zeigler, L.; Asolkar, R.N.; Singan, V.; Lapidus, A.; Fenical, W.; Jensen, P.R.; Moore, B.S. Genome sequencing reveals complex secondary metabolome in the marine actinomycete Salinispora tropica. Proc. Natl. Acad. Sci. USA 2007, 104, 10376–10381. [Google Scholar] [CrossRef] [Green Version]
- Fenical, W.; Jensen, P.R.; Palladino, M.A.; Lam, K.S.; Lloyd, G.K.; Potts, B.C. Discovery and development of the anticancer agent salinosporamide A (NPI-0052). Bioorganic Med. Chem. 2009, 17, 2175–2180. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.R.; Mafnas, C. Biogeography of the marine actinomycete Salinispora. Environ. Microbiol. 2006, 8, 1881–1888. [Google Scholar] [CrossRef]
- Maldonado, L.A.; Fenical, W.; Jensen, P.R.; Kauffman, C.A.; Mincer, T.J.; Ward, A.C.; Bull, A.T.; Goodfellow, M. Salinispora arenicola gen. nov., sp. nov. and Salinispora tropica sp. nov., obligate marine actinomycetes belonging to the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 2005, 55, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, L.; Jensen, P.R.; Freel, K.C.; Brown, R.; Jones, A.L.; Kim, B.-Y.; Goodfellow, M. Salinispora pacifica sp. nov., an actinomycete from marine sediments. Antonie Van Leeuwenhoek 2013, 103, 1069–1078. [Google Scholar] [CrossRef]
- Contador, C.A.; Rodríguez, V.; Andrews, B.A.; Asenjo, J.A. Use of genome-scale models to get new insights into the marine actinomycete genus Salinispora. BMC Syst. Biol. 2019, 13, 11. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.C.; Gulder, T.A.; Mahmud, T.; Moore, B.S. Shared biosynthesis of the saliniketals and rifamycins in Salinispora arenicola is controlled by the sare1259-encoded cytochrome P450. J. Am. Chem. Soc. 2010, 132, 12757–12765. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-M.A.; Chu, K.; Palaniappan, K.; Ratner, A.; Huang, J.; Huntemann, M.; Hajek, P.; Ritter, S.; Varghese, N.; Seshadri, R. The IMG/M data management and analysis system v. 6.0: New tools and advanced capabilities. Nucleic Acids Res. 2021, 49, D751–D763. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Stamatis, D.; Bertsch, J.; Ovchinnikova, G.; Sundaramurthi, J.C.; Lee, J.; Kandimalla, M.; Chen, I.-M.A.; Kyrpides, N.C.; Reddy, T. Genomes OnLine Database (GOLD) v. 8: Overview and updates. Nucleic Acids Res. 2021, 49, D723–D733. [Google Scholar] [CrossRef] [PubMed]
- Mnguni, F.C.; Padayachee, T.; Chen, W.; Gront, D.; Yu, J.-H.; Nelson, D.R.; Syed, K. More P450s are involved in secondary metabolite biosynthesis in Streptomyces compared to Bacillus, Cyanobacteria and Mycobacterium. Int. J. Mol. Sci. 2020, 21, 4814. [Google Scholar] [CrossRef] [PubMed]
- Syed, P.R.; Chen, W.; Nelson, D.R.; Kappo, A.P.; Yu, J.H.; Karpoormath, R.; Syed, K. Cytochrome P450 Monooxygenase CYP139 Family Involved in the Synthesis of Secondary Metabolites in 824 Mycobacterial Species. Int. J. Mol. Sci. 2019, 20, 2690. [Google Scholar] [CrossRef] [Green Version]
- Syed, K.; Mashele, S.S. Comparative analysis of P450 signature motifs EXXR and CXG in the large and diverse kingdom of fungi: Identification of evolutionarily conserved amino acid patterns characteristic of P450 family. PLoS ONE 2014, 9, e95616. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, O. Substrate recognition sites in cytochrome P450 family 2 (CYP2) proteins inferred from comparative analyses of amino acid and coding nucleotide sequences. J. Biol. Chem. 1992, 267, 83–90. [Google Scholar] [CrossRef]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 nomenclature, 2004. Methods Mol. Biol. 2006, 320, 1–10. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. 1998, 107, 15–24. [Google Scholar] [CrossRef]
- Nzuza, N.; Padayachee, T.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. Diversification of Ferredoxins across Living Organisms. Curr. Issues Mol. Biol. 2021, 43, 1374–1390. [Google Scholar] [CrossRef]
- Nzuza, N.; Padayachee, T.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. Ancient Bacterial Class Alphaproteobacteria Cytochrome P450 Monooxygenases Can Be Found in Other Bacterial Species. Int. J. Mol. Sci. 2021, 22, 5542. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryś, J.D.; Gront, D. VisuaLife: Library for interactive visualization in rich web applications. Bioinformatics 2021, 37, 3662–3663. [Google Scholar] [CrossRef] [PubMed]
- Msomi, N.N.; Padayachee, T.; Nzuza, N.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. In silico analysis of P450s and their role in secondary metabolism in the bacterial class Gammaproteobacteria. Molecules 2021, 26, 1538. [Google Scholar] [CrossRef]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef] [Green Version]
- Padayachee, T.; Nzuza, N.; Chen, W.; Nelson, D.R.; Syed, K. Impact of lifestyle on cytochrome P450 monooxygenase repertoire is clearly evident in the bacterial phylum Firmicutes. Sci. Rep. 2020, 10, 13982. [Google Scholar] [CrossRef]
- Khumalo, M.J.; Nzuza, N.; Padayachee, T.; Chen, W.; Yu, J.-H.; Nelson, D.; Syed, K. Comprehensive analyses of cytochrome P450 monoxygenases and secondary metabolite biosynthetic gene clusters in Cyanobacteria. Int. J. Mol. Sci. 2020, 21, 656. [Google Scholar] [CrossRef] [Green Version]
- Senate, L.M.; Tjatji, M.P.; Pillay, K.; Chen, W.; Zondo, N.M.; Syed, P.R.; Mnguni, F.C.; Chiliza, Z.E.; Bamal, H.D.; Karpoormath, R.; et al. Similarities, variations, and evolution of cytochrome P450s in Streptomyces versus Mycobacterium. Sci. Rep. 2019, 9, 3962. [Google Scholar] [CrossRef] [Green Version]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Ngcobo, N.S.; Chiliza, Z.E.; Chen, W.; Yu, J.-H.; Nelson, D.R.; Tuszynski, J.A.; Preto, J.; Syed, K. Comparative Analysis, Structural Insights, and Substrate/Drug Interaction of CYP128A1 in Mycobacterium Tuberculosis. Int. J. Mol. Sci. 2020, 21, 4816. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.; Leung, J.; Swanson, S.; Idler, K.; McAlpine, J. An erythromycin derivative produced by targeted gene disruption in Saccharopolyspora erythraea. Science 1991, 252, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Trefzer, A.; Jungmann, V.; Molnár, I.; Botejue, A.; Buckel, D.; Frey, G.; Hill, D.S.; Jörg, M.; Ligon, J.M.; Mason, D. Biocatalytic conversion of avermectin to 4″-oxo-avermectin: Improvement of cytochrome p450 monooxygenase specificity by directed evolution. Appl. Environ. Microbiol. 2007, 73, 4317–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, Y.; Kabumoto, H.; Nishimura, K.; Fujii, T.; Yanai, S.; Takeda, K.; Tamura, N.; Arisawa, A.; Tamura, T. Purification, characterization, and directed evolution study of a vitamin D3 hydroxylase from Pseudonocardia autotrophica. Biochem. Biophys. Res. Commun. 2009, 385, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Prior, J.E.; Shokati, T.; Christians, U.; Gill, R.T. Identification and characterization of a bacterial cytochrome P450 for the metabolism of diclofenac. Appl. Microbiol. Biotechnol. 2010, 85, 625–633. [Google Scholar] [CrossRef]
- Moody, S.C.; Loveridge, E.J. CYP105-diverse structures, functions and roles in an intriguing family of enzymes in Streptomyces. J. Appl. Microbiol. 2014, 117, 1549–1563. [Google Scholar] [CrossRef] [Green Version]
- McLean, K.J.; Lafite, P.; Levy, C.; Cheesman, M.R.; Mast, N.; Pikuleva, I.A.; Leys, D.; Munro, A.W. The Structure of Mycobacterium tuberculosis CYP125: Molecular basis for cholesterol binding in a P450 needed for host infection. J. Biol. Chem. 2009, 284, 35524–35533. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, H.; Guan, S.; Johnston, J.B.; Chow, E.D.; Kells, P.M.; Burlingame, A.L.; Cox, J.S.; Podust, L.M.; de Montellano, P.R.O. Mycobacterium tuberculosis CYP125A1, a steroid C27 monooxygenase that detoxifies intracellularly generated cholest-4-en-3-one. Mol. Microbiol. 2010, 77, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Bergstrand, L.H.; Cardenas, E.; Holert, J.; Van Hamme, J.D.; Mohn, W.W. Delineation of steroid-degrading microorganisms through comparative genomic analysis. mBio 2016, 7, e00166-16. [Google Scholar] [CrossRef] [Green Version]
- Cryle, M.J.; Schlichting, I. Structural insights from a P450 Carrier Protein complex reveal how specificity is achieved in the P450BioI ACP complex. Proc. Natl. Acad. Sci. USA 2008, 105, 15696–15701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracco, P.; Janssen, D.B.; Schallmey, A. Selective steroid oxyfunctionalisation by CYP154C5, a bacterial cytochrome P450. Microb. Cell Factories 2013, 12, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subedi, P.; Kim, K.-H.; Hong, Y.-S.; Lee, J.-H.; Oh, T.-J. Enzymatic characterization and comparison of two steroid hydroxylases CYP154C3-1 and CYP154C3-2 from Streptomyces species. J. Microbiol. Biotechnol. 2021, 31, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-Y.; Park, H.-Y.; Kim, B.-G. Characterization of bi-functional CYP154 from Nocardia farcinica IFM10152 in the O-dealkylation and ortho-hydroxylation of formononetin. Enzym. Microb. Technol. 2010, 47, 327–334. [Google Scholar] [CrossRef]
- Chen, H.; Walsh, C.T. Coumarin formation in novobiocin biosynthesis: Beta-hydroxylation of the aminoacyl enzyme tyrosyl-S-NovH by a cytochrome P450 NovI. Chem. Biol. 2001, 8, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Uhlmann, S.; Sussmuth, R.D.; Cryle, M.J. Cytochrome p450sky interacts directly with the nonribosomal peptide synthetase to generate three amino acid precursors in skyllamycin biosynthesis. ACS Chem. Biol. 2013, 8, 2586–2596. [Google Scholar] [CrossRef]
- Lauer, B.; Russwurm, R.; Bormann, C. Molecular characterization of two genes from Streptomyces tendae Tu901 required for the formation of the 4-formyl-4-imidazolin-2-one-containing nucleoside moiety of the peptidyl nucleoside antibiotic nikkomycin. Eur. J. Biochem. 2000, 267, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Niu, G.; Li, R.; Liu, G.; Tan, H. Identification and characterization of sanH and sanI involved in the hydroxylation of pyridyl residue during nikkomycin biosynthesis in Streptomyces ansochromogenes. Curr. Microbiol. 2007, 55, 537–542. [Google Scholar] [CrossRef]
- Mendes, M.V.; Anton, N.; Martin, J.F.; Aparicio, J.F. Characterization of the polyene macrolide P450 epoxidase from Streptomyces natalensis that converts de-epoxypimaricin into pimaricin. Biochem. J. 2005, 386, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, P.; Lynch, S.; Flood, E.; Finnan, S.; Oliynyk, M. Amphotericin biosynthesis in Streptomyces nodosus: Deductions from analysis of polyketide synthase and late genes. Chem. Biol. 2001, 8, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Shafiee, A.; Hutchinson, C.R. Macrolide antibiotic biosynthesis: Isolation and properties of two forms of 6-deoxyerythronolide B hydroxylase from Saccharopolyspora erythraea (Streptomyces erythreus). Biochemistry 1987, 26, 6204–6210. [Google Scholar] [CrossRef] [PubMed]
- Stassi, D.; Donadio, S.; Staver, M.J.; Katz, L. Identification of a Saccharopolyspora erythraea gene required for the final hydroxylation step in erythromycin biosynthesis. J. Bacteriol. 1993, 175, 182–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merson-Davies, L.A.; Cundliffe, E. Analysis of five tylosin biosynthetic genes from the tyllBA region of the Streptomyces fradiae genome. Mol. Microbiol. 1994, 13, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Fouces, R.; Mellado, E.; Diez, B.; Barredo, J.L. The tylosin biosynthetic cluster from Streptomyces fradiae: Genetic organization of the left region. Microbiology 1999, 145 Pt 4, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, J.; Wang, H.; Xie, Y.; Ju, J.; Yan, Y.; Zhang, H. Structural analysis of HmtT and HmtN involved in the tailoring steps of himastatin biosynthesis. FEBS Lett. 2013, 587, 1675–1680. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Z.; Huang, H.; Luo, M.; Zuo, D.; Wang, B.; Sun, A.; Cheng, Y.Q.; Zhang, C.; Ju, J. Biosynthesis of himastatin: Assembly line and characterization of three cytochrome P450 enzymes involved in the post-tailoring oxidative steps. Angew. Chem. 2011, 50, 7797–7802. [Google Scholar] [CrossRef]
- Aparicio, J.F.; Molnar, I.; Schwecke, T.; Konig, A.; Haydock, S.F.; Khaw, L.E.; Staunton, J.; Leadlay, P.F. Organization of the biosynthetic gene cluster for rapamycin in Streptomyces hygroscopicus: Analysis of the enzymatic domains in the modular polyketide synthase. Gene 1996, 169, 9–16. [Google Scholar] [CrossRef]
- Molnar, I.; Aparicio, J.F.; Haydock, S.F.; Khaw, L.E.; Schwecke, T.; Konig, A.; Staunton, J.; Leadlay, P.F. Organisation of the biosynthetic gene cluster for rapamycin in Streptomyces hygroscopicus: Analysis of genes flanking the polyketide synthase. Gene 1996, 169, 1–7. [Google Scholar] [CrossRef]
- Zocher, G.; Richter, M.E.; Mueller, U.; Hertweck, C. Structural fine-tuning of a multifunctional cytochrome P450 monooxygenase. J. Am. Chem. Soc. 2011, 133, 2292–2302. [Google Scholar] [CrossRef]
- Johnston, J.B.; Kells, P.M.; Podust, L.M.; Ortiz de Montellano, P.R. Biochemical and structural characterization of CYP124: A methyl-branched lipid omega-hydroxylase from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2009, 106, 20687–20692. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Category | Salinispora Species | Streptomyces Species | Mycobacterial Species | Cyanobacterial Species | Firmicutes Species | Alphaproteobacterial Species | Gammaproteobacterial Species |
|---|---|---|---|---|---|---|---|
| Species analysed | 126 | 203 | 60 | 114 | 972 | 599 | 1261 |
| Species without P450s | 0 | 0 | 0 | 0 | 743 | 370 | 1091 |
| Species with P450s | 126 | 203 | 60 | 114 | 229 | 229 | 169 |
| Percentage of species with P450s | 100 | 100 | 100 | 100 | 24 | 38 | 13 |
| No. of P450s | 2643 | 5460 | 1784 | 341 | 712 | 873 | 277 |
| No. of families | 45 | 253 | 77 | 36 | 14 | 143 | 81 |
| No. of subfamilies | 103 | 698 | 132 | 79 | 53 | 214 | 102 |
| Dominant P450 family | CYP105 | CYP107 | CYP125 | CYP110 | CYP107 | CYP202 | CYP133 & CYP107 |
| Average No. of P450s | 21 | 27 | 30 | 3 | 3 | 4 | 2 |
| P450 diversity percentage | 0.01 | 0.02 | 0.07 | 0.09 | 0.01 | 0.07 | 0.17 |
| No. of P450s part of BGCs | 1236 | 1231 | 204 | 27 | 126 | 21 | 49 |
| No. of P450 families part of BGCs | 35 | 135 | 31 | 6 | 10 | 16 | 22 |
| Percentage of P450s part of BGCs | 47 | 23 | 11 | 8 | 18 | 2 | 18 |
| Reference(s) | This study | [56,72] | [72,73] | [71] | [70] | [64] | [68] |
| Species Name | No. of P450s | No. of P450 Fragments | No. of P450 Families | No. of Subfamilies |
|---|---|---|---|---|
| Salinispora arenicola CNH996 | 26 | 6 | 14 | 25 |
| Salinispora arenicola CNH996B | 27 | 14 | 25 | |
| Salinispora arenicola CNY280 | 35 | 18 | 32 | |
| Salinispora arenicola CNH877 | 34 | 15 | 30 | |
| Salinispora arenicola CNS848 | 32 | 16 | 29 | |
| Salinispora arenicola CNT798 | 31 | 14 | 27 | |
| Salinispora arenicola CNH643 | 31 | 1 | 14 | 28 |
| Salinispora arenicola CNS-991 | 31 | 15 | 28 | |
| Salinispora arenicola CNT799 | 31 | 14 | 28 | |
| Salinispora arenicola CNY679 | 31 | 1 | 14 | 27 |
| Salinispora arenicola CNT850 | 31 | 13 | 27 | |
| Salinispora arenicola CNT800 | 31 | 14 | 28 | |
| Salinispora arenicola CNY011 | 31 | 14 | 26 | |
| Salinispora arenicola CNY230 | 30 | 17 | 30 | |
| Salinispora arenicola CNH713 | 30 | 14 | 27 | |
| Salinispora arenicola CNH905 | 31 | 1 | 14 | 28 |
| Salinispora arenicola CNT857 | 30 | 14 | 28 | |
| Salinispora arenicola CNY281 | 29 | 1 | 17 | 29 |
| Salinispora arenicola CNH941 | 29 | 14 | 26 | |
| Salinispora arenicola CNB527 | 29 | 4 | 15 | 27 |
| Salinispora arenicola CNT859 | 29 | 13 | 26 | |
| Salinispora arenicola CNT005 | 28 | 16 | 28 | |
| Salinispora arenicola CNH964 | 28 | 1 | 14 | 24 |
| Salinispora arenicola CNP193 | 28 | 14 | 26 | |
| Salinispora arenicola CNP105 | 28 | 2 | 14 | 25 |
| Salinispora arenicola CNH646 | 28 | 14 | 26 | |
| Salinispora arenicola CNR425 | 28 | 15 | 28 | |
| Salinispora arenicola CNS-205 | 28 | 1 | 15 | 28 |
| Salinispora arenicola ATCC BAA-917 | 27 | 13 | 11 | 21 |
| Salinispora arenicola CNY685 | 26 | 6 | 14 | 26 |
| Salinispora arenicola CNS325 | 26 | 13 | 26 | |
| Salinispora arenicola CNS744 | 26 | 13 | 26 | |
| Salinispora arenicola CNY694 | 26 | 6 | 13 | 26 |
| Salinispora arenicola CNY260 | 26 | 14 | 26 | |
| Salinispora arenicola CNT-088 | 26 | 1 | 13 | 24 |
| Salinispora arenicola CNB458 | 26 | 4 | 13 | 26 |
| Salinispora arenicola CNS296 | 25 | 1 | 14 | 25 |
| Salinispora arenicola CNY231 | 25 | 14 | 26 | |
| Salinispora arenicola CNY282 | 25 | 13 | 25 | |
| Salinispora arenicola CNS299 | 25 | 1 | 14 | 25 |
| Salinispora arenicola CNQ748 | 25 | 13 | 25 | |
| Salinispora arenicola CNY244 | 25 | 13 | 25 | |
| Salinispora arenicola CNS820 | 25 | 13 | 25 | |
| Salinispora arenicola CNS673 | 25 | 13 | 25 | |
| Salinispora arenicola CNY237 | 24 | 12 | 24 | |
| Salinispora arenicola CNS342 | 24 | 1 | 13 | 24 |
| Salinispora arenicola CNH718 | 24 | 1 | 13 | 24 |
| Salinispora arenicola CNX891 | 24 | 3 | 15 | 24 |
| Salinispora arenicola CNY256 | 24 | 13 | 25 | |
| Salinispora arenicola CNS243 | 24 | 1 | 13 | 24 |
| Salinispora arenicola CNY234 | 24 | 1 | 13 | 24 |
| Salinispora arenicola CNY690 | 24 | 4 | 13 | 24 |
| Salinispora arenicola CNQ884 | 23 | 1 | 13 | 25 |
| Salinispora arenicola CNR107 | 22 | 12 | 22 | |
| Salinispora arenicola CNR921 | 22 | 12 | 22 | |
| Salinispora arenicola CNH962 | 22 | 1 | 12 | 22 |
| Salinispora arenicola CNX481 | 22 | 2 | 12 | 22 |
| Salinispora arenicola CNH963 | 22 | 1 | 12 | 22 |
| Salinispora arenicola CNX814 | 22 | 1 | 12 | 21 |
| Salinispora arenicola CNY486 | 22 | 1 | 13 | 24 |
| Salinispora arenicola CNX508 | 21 | 1 | 12 | 21 |
| Salinispora arenicola CNX482 | 21 | 1 | 12 | 21 |
| Salinispora pacifica CNS996 | 21 | 1 | 15 | 21 |
| Salinispora pacifica CNS237 | 20 | 1 | 12 | 19 |
| Salinispora pacifica CNY646 | 20 | 1 | 13 | 19 |
| Salinispora tropica CNT261 | 20 | 2 | 10 | 18 |
| Salinispora pacifica DSM 45548 | 19 | 7 | 10 | |
| Salinispora pacifica CNT045 | 19 | 1 | 13 | 19 |
| Salinispora pacifica CNT124 | 19 | 13 | 19 | |
| Salinispora pacifica DSM 45543 | 19 | 12 | 18 | |
| Salinispora tropica CNB536 | 19 | 11 | 19 | |
| Salinispora tropica CNH898 | 18 | 1 | 11 | 20 |
| Salinispora pacifica CNT403 | 18 | 1 | 12 | 17 |
| Salinispora pacifica CNS860 | 18 | 2 | 11 | 16 |
| Salinispora pacifica CNS863 | 18 | 2 | 12 | 17 |
| Salinispora tropica CNY012 | 18 | 2 | 10 | 18 |
| Salinispora pacifica CNT584 | 17 | 11 | 17 | |
| Salinispora pacifica DSM 45549 | 17 | 1 | 11 | 16 |
| Salinispora pacifica CNR114 | 17 | 1 | 13 | 17 |
| Salinispora tropica CNR699 | 17 | 2 | 10 | 16 |
| Salinispora pacifica CNT854 | 18 | 13 | 18 | |
| Salinispora pacifica CNT150 | 17 | 1 | 11 | 16 |
| Salinispora pacifica CNT131 | 17 | 1 | 11 | 15 |
| Salinispora pacifica DSM 45544 | 16 | 11 | 16 | |
| Salinispora pacifica CNT003 | 16 | 1 | 10 | 15 |
| Salinispora tropica CNY681 | 16 | 1 | 10 | 16 |
| Salinispora tropica CNS197 | 16 | 1 | 10 | 16 |
| Salinispora tropica CNY678 | 16 | 1 | 10 | 16 |
| Salinispora tropica CNT250 | 16 | 1 | 10 | 16 |
| Salinispora tropica CNB-440 | 16 | 1 | 10 | 16 |
| Salinispora tropica CNS416 | 15 | 9 | 15 | |
| Salinispora pacifica CNT001 | 15 | 1 | 11 | 15 |
| Salinispora pacifica CNY498 | 15 | 1 | 11 | 15 |
| Salinispora pacifica CNR909 | 15 | 1 | 10 | 15 |
| Salinispora tropica CNB476 | 15 | 1 | 9 | 15 |
| Salinispora pacifica CNR894 | 15 | 11 | 15 | |
| Salinispora pacifica CNY363 | 15 | 11 | 15 | |
| Salinispora pacifica CNS055 | 15 | 9 | 15 | |
| Salinispora pacifica CNT603 | 15 | 1 | 11 | 15 |
| Salinispora pacifica CNT138 | 14 | 10 | 14 | |
| Salinispora pacifica DSM 45547 | 14 | 1 | 10 | 14 |
| Salinispora pacifica CNH732 | 14 | 1 | 10 | 14 |
| Salinispora pacifica CNY703 | 14 | 9 | 13 | |
| Salinispora pacifica CNQ768 | 14 | 1 | 10 | 14 |
| Salinispora pacifica CNY673 | 14 | 10 | 14 | |
| Salinispora pacifica CNT855 | 14 | 9 | 14 | |
| Salinispora pacifica CNY239 | 14 | 1 | 10 | 14 |
| Salinispora pacifica CNR942 | 14 | 1 | 10 | 14 |
| Salinispora pacifica DSM 45546 | 14 | 1 | 10 | 14 |
| Salinispora pacifica CNT609 | 14 | 1 | 10 | 14 |
| Salinispora pacifica CNY331 | 14 | 10 | 14 | |
| Salinispora tropica CNR416 | 14 | 9 | 14 | |
| Salinispora pacifica CNY330 | 13 | 9 | 13 | |
| Salinispora pacifica CNT851 | 13 | 1 | 9 | 13 |
| Salinispora pacifica CNT796 | 13 | 1 | 9 | 13 |
| Salinispora pacifica CNS103 | 13 | 9 | 13 | |
| Salinispora pacifica CNY202 | 13 | 1 | 9 | 13 |
| Salinispora pacifica CNT133A | 13 | 9 | 13 | |
| Salinispora arenicola CNY666 | 13 | 5 | 8 | 13 |
| Salinispora pacifica CNT029 | 13 | 1 | 9 | 13 |
| Salinispora pacifica CNT084 | 13 | 9 | 13 | |
| Salinispora pacifica CNR510 | 13 | 9 | 13 | |
| Salinispora pacifica CNT569 | 12 | 1 | 9 | 13 |
| Salinispora pacifica CNT-133 | 11 | 11 | 7 | 9 |
| Salinispora pacifica CNS801 | 10 | 7 | 10 | |
| Salinispora pacifica CNT148 | 10 | 7 | 10 |
| P450 Family | P450 Count | Percentage Count | Subfamily | Count | Percentage Count |
|---|---|---|---|---|---|
| CYP1004 | 34 | 1.29% | A | 17 | 0.64 |
| B | 17 | 0.64 | |||
| CYP1005 | 127 | 4.81% | A | 127 | 4.79 |
| CYP1037 | 2 | 0.08% | B | 2 | 0.08 |
| CYP1051 | 60 | 2.27% | A | 60 | 2.26 |
| CYP1056 | 2 | 0.08% | B | 2 | 0.08 |
| CYP105 | 600 | 22.70% | AB | 124 | 4.67 |
| AH | 4 | 0.15 | |||
| B | 1 | 0.04 | |||
| BL | 78 | 2.94 | |||
| BN | 1 | 0.04 | |||
| CH | 44 | 1.66 | |||
| CN | 62 | 2.34 | |||
| CP | 62 | 2.34 | |||
| CT | 41 | 1.55 | |||
| EJ | 3 | 0.11 | |||
| G | 62 | 2.34 | |||
| H | 3 | 0.11 | |||
| J | 52 | 1.96 | |||
| W | 63 | 2.37 | |||
| CYP107 | 551 | 20.85% | AW | 65 | 2.45 |
| AX | 75 | 2.83 | |||
| AY | 116 | 4.37 | |||
| CL | 3 | 0.11 | |||
| CT | 6 | 0.23 | |||
| E | 38 | 1.43 | |||
| EP | 2 | 0.08 | |||
| EU | 44 | 1.66 | |||
| FH | 25 | 0.94 | |||
| FJ | 20 | 0.75 | |||
| FS | 61 | 2.30 | |||
| GU | 1 | 0.04 | |||
| HF | 2 | 0.08 | |||
| LA | 6 | 0.23 | |||
| N | 2 | 0.08 | |||
| NE | 2 | 0.08 | |||
| NF | 2 | 0.08 | |||
| NG | 4 | 0.15 | |||
| NH | 8 | 0.30 | |||
| Q | 63 | 2.37 | |||
| Z | 6 | 0.23 | |||
| CYP1114 | 1 | 0.04% | C | 1 | 0.04 |
| CYP113 | 24 | 0.91% | B | 6 | 0.23 |
| D | 1 | 0.04 | |||
| E | 10 | 0.38 | |||
| R | 2 | 0.08 | |||
| S | 2 | 0.08 | |||
| T | 1 | 0.04 | |||
| X | 2 | 0.08 | |||
| CYP1197 | 1 | 0.04% | A | 1 | 0.04 |
| CYP1198 | 43 | 1.63% | B | 43 | 1.62 |
| CYP1207 | 4 | 0.15% | A | 4 | 0.15 |
| CYP1223 | 6 | 0.23% | D | 2 | 0.08 |
| A | 4 | 0.15 | |||
| CYP1226 | 2 | 0.08% | A | 2 | 0.08 |
| CYP124 | 15 | 0.57% | M | 15 | 0.57 |
| CYP125 | 164 | 6.21% | A | 128 | 4.82 |
| G | 36 | 1.36 | |||
| CYP1269 | 2 | 0.08% | A | 2 | 0.08 |
| CYP1278 | 11 | 0.42% | A | 5 | 0.19 |
| B | 6 | 0.23 | |||
| CYP1437 | 1 | 0.04% | C | 1 | 0.04 |
| CYP146 | 1 | 0.04% | A | 1 | 0.04 |
| CYP1522 | 1 | 0.04% | A | 1 | 0.04 |
| CYP154 | 155 | 5.86% | AJ | 4 | 0.15 |
| J | 1 | 0.04 | |||
| M | 150 | 5.65 | |||
| CYP1611 | 1 | 0.04% | B | 1 | 0.04 |
| CYP161 | 28 | 1.06% | N | 23 | 0.87 |
| 0.00% | T | 5 | 0.19 | ||
| CYP162 | 39 | 1.48% | A | 11 | 0.41 |
| B | 2 | 0.08 | |||
| G | 2 | 0.08 | |||
| H | 1 | 0.04 | |||
| J | 1 | 0.04 | |||
| K | 1 | 0.04 | |||
| L | 1 | 0.04 | |||
| M | 1 | 0.04 | |||
| N | 1 | 0.04 | |||
| P | 18 | 0.68 | |||
| CYP163 | 39 | 1.48% | A | 2 | 0.08 |
| B | 37 | 1.39 | |||
| CYP164 | 4 | 0.15% | C | 4 | 0.15 |
| CYP166 | 62 | 2.35% | A | 62 | 2.34 |
| CYP173 | 1 | 0.04% | K | 1 | 0.04 |
| CYP1902 | 2 | 0.08% | A | 2 | 0.08 |
| CYP2054 | 22 | 0.83% | A | 22 | 0.83 |
| CYP205 | 1 | 0.04% | A | 1 | 0.04 |
| CYP208 | 126 | 4.77% | A | 126 | 4.75 |
| CYP2091 | 1 | 0.04% | A | 1 | 0.04 |
| CYP2098 | 2 | 0.08% | A | 2 | 0.08 |
| CYP211 | 225 | 8.51% | B | 124 | 4.67 |
| C | 101 | 3.81 | |||
| CYP2296 | 1 | 0.04% | A | 1 | 0.04 |
| CYP244 | 107 | 4.05% | A | 107 | 4.03 |
| CYP245 | 83 | 3.14% | A | 83 | 3.13 |
| CYP247 | 21 | 0.79% | A | 21 | 0.79 |
| CYP248 | 63 | 2.38% | A | 63 | 2.37 |
| CYP2611 | 1 | 0.04% | B | 1 | 0.04 |
| CYP283 | 1 | 0.04% | A | 1 | 0.04 |
| CYP285 | 4 | 0.15% | A | 2 | 0.08 |
| D | 2 | 0.08 | |||
| CYP294A4 | 2 | 0.08% | A | 2 | 0.08 |
| smBGC Type | smBGC Type Count | smBGC Type Variety | P450s | P450 Count |
|---|---|---|---|---|
| Bacteriocin | 47 | 46 | CYP107AW | 46 |
| 1 | CYP283A | 1 | ||
| betalactone | 2 | 1 | CYP162A6,CYP107HF1 | 2 |
| 1 | CYP113S1 | 1 | ||
| butyrolactone | 1 | 1 | CYP105CT1,CYP154M5 | 2 |
| Indole | 54 | 51 | CYP244A,CYP245A | 102 |
| 3 | CYP244A | 3 | ||
| ladderane | 18 | 4 | CYP154M15,CYP125G6,CYP107FS2,CYP105CN1,CYP105CP2 | 20 |
| 8 | CYP107AX-fragment | 8 | ||
| 6 | CYP107AX | 6 | ||
| lanthipeptide | 2 | 1 | CYP1223A5 | 1 |
| 1 | CYP105CP2,CYP105CN1,CYP107FS2,CYP248A2,CYP105W2 | 5 | ||
| LAP | 1 | 1 | CYP154AJ2 | 1 |
| lipolanthine | 2 | 2 | CYP1223A5 | 2 |
| NRPS | 205 | 1 | CYP1004B1,CYP1004A1 | 2 |
| 8 | CYP1004B,CYP1004A,CYP125G | 24 | ||
| 1 | CYP105CH2-fragment,CYP105CH1-fragment | 2 | ||
| 1 | CYP105CN1 | 1 | ||
| 1 | CYP105CN1,CYP105CP2 | 2 | ||
| 1 | CYP105CN1,CYP107FS2,CYP125G6,CYP154M15 | 4 | ||
| 1 | CYP105CN1,CYP107FS2,CYP247A7 | 3 | ||
| 1 | CYP105CP2 | 1 | ||
| 7 | CYP105CP2,CYP105CN1,CYP107FS2 | 21 | ||
| 10 | CYP105CP2,CYP105CN1,CYP107FS2,CYP125G6,CYP154M15 | 50 | ||
| 2 | CYP105CP2,CYP105CN1,CYP107FS2,CYP248A2 | 8 | ||
| 1 | CYP105CP2,CYP105CN1,CYP107FS2,CYP248A2,CYP105W2 | 5 | ||
| 3 | CYP105W | 3 | ||
| 38 | CYP107AY | 38 | ||
| 1 | CYP107AY14,CYP244A-fragment2 | 2 | ||
| 6 | CYP107AY2,CYP105CT1,CYP154M5 | 18 | ||
| 1 | CYP107AY2,CYP163B16 | 2 | ||
| 1 | CYP107AY7,CYP244A5,CYP245A11 | 3 | ||
| 1 | CYP107AY9,CYP162B3 | 2 | ||
| 1 | CYP107AY9,CYP244A10 | 2 | ||
| 2 | CYP107CL2,CYP1056B2 | 4 | ||
| 5 | CYP107CT3 | 5 | ||
| 1 | CYP107CT3,CYP107AY7 | 2 | ||
| 1 | CYP107FS2 | 1 | ||
| 6 | CYP107FS2,CYP105CN1,CYP105CP2 | 18 | ||
| 2 | CYP107NH1,CYP247A8,CYP107Z27 | 6 | ||
| 1 | CYP107Z27,CYP247A8,CYP107NH1 | 3 | ||
| 1 | CYP113D13,CYP163B22 | 2 | ||
| 1 | CYP1196A2 | 1 | ||
| 1 | CYP1198B1 | 1 | ||
| 1 | CYP1198B1,CYP107AY2 | 2 | ||
| 2 | CYP1207A12 | 2 | ||
| 1 | CYP125G1,CYP1004A1,CYP1004B1 | 3 | ||
| 1 | CYP125G6,CYP154M15 | 2 | ||
| 4 | CYP1278A4 | 4 | ||
| 1 | CYP1437C1 | 1 | ||
| 1 | CYP154AJ3 | 1 | ||
| 1 | CYP154J2,CYP244A5,CYP245A11 | 3 | ||
| 5 | CYP154M1,CYP208A4 | 10 | ||
| 6 | CYP154M | 6 | ||
| 3 | CYP154M,CYP208A | 6 | ||
| 1 | CYP154M16,CYP211C6 | 2 | ||
| 1 | CYP154M21,CYP154M13 | 2 | ||
| 1 | CYP154M21,CYP154M13,CYP105W2,CYP248A2 | 4 | ||
| 3 | CYP154M21,CYP154M13,CYP105W2,CYP248A2,CYP154M20 | 15 | ||
| 1 | CYP154M21,CYP154M13,CYP105W2,CYP248A2,CYP154M20,CYP162P1 | 6 | ||
| 12 | CYP162 | 12 | ||
| 1 | CYP163A10,CYP162K1 | 2 | ||
| 15 | CYP163B | 15 | ||
| 3 | CYP164C2 | 3 | ||
| 5 | CYP208A21,CYP154M16 | 10 | ||
| 2 | CYP208A4,CYP154M1 | 4 | ||
| 8 | CYP244A,CYP107AY | 16 | ||
| 5 | CYP244A5,CYP245A11 | 10 | ||
| 2 | CYP244A,CYP107AY | 4 | ||
| 2 | CYP244A | 2 | ||
| 1 | CYP245A11 | 1 | ||
| 3 | CYP247A | 3 | ||
| 1 | CYP247A8,CYP107NH1 | 2 | ||
| 1 | CYP247A8,CYP107Z27 | 2 | ||
| 1 | CYP248A2 | 1 | ||
| 1 | CYP248A2,CYP105W2 | 2 | ||
| 1 | CYP285D2 | 1 | ||
| NRPS-like | 30 | 5 | CYP107EU | 5 |
| 1 | CYP107EU1,CYP1198B1,CYP105CH1 | 3 | ||
| 1 | CYP107FH3,CYP161N4,CYP107AY9 | 3 | ||
| 7 | CYP107FH3,CYP2054A3,CYP161N4 | 21 | ||
| 2 | CYP161N4,CYP2054A3,CYP107FH3 | 6 | ||
| 6 | CYP162A8 | 6 | ||
| 6 | CYP166A4 | 6 | ||
| 1 | CYP166A4,CYP107Q4,CYP105G5 | 3 | ||
| 1 | CYP285A9-fragment,CYP285A9-fragment | 2 | ||
| oligosaccharide | 35 | 1 | CYP105CP2 | 1 |
| 1 | CYP105W2,CYP107FS2,CYP105CN1,CYP105CP2 | 4 | ||
| 1 | CYP105W2,CYP107NH1 | 2 | ||
| 1 | CYP105W2,CYP154M20,CYP154M13,CYP154M21,CYP248A2 | 5 | ||
| 5 | CYP105W2,CYP248A2 | 10 | ||
| 1 | CYP105W2,CYP248A2,CYP107FS2 | 3 | ||
| 9 | CYP105W2/3,CYP248A2,CYP107FS2,CYP105CN1,CYP105CP2 | 45 | ||
| 1 | CYP1269A2 | 1 | ||
| 8 | CYP154M20,CYP248A2,CYP105W2,CYP154M13,CYP154M21 | 40 | ||
| 1 | CYP2091A1 | 1 | ||
| 3 | CYP248A2 | 3 | ||
| 3 | CYP248A2,CYP105W2/3 | 6 | ||
| other | 4 | 2 | CYP247A7 | 2 |
| 2 | CYP105AH4 | 2 | ||
| 1 | CYP1004A3,CYP1004B4,CYP113E2,CYP163B18 | 4 | ||
| T1PKS | 223 | 1 | CYP105AH4 | 1 |
| 1 | CYP105BN4 | 1 | ||
| 17 | CYP105CH1/2 | 17 | ||
| 1 | CYP105CN1 | 1 | ||
| 4 | CYP105G5 | 4 | ||
| 16 | CYP105G5,CYP107Q4 | 32 | ||
| 2 | CYP105H11 | 2 | ||
| 1 | CYP107AY13 | 1 | ||
| 24 | CYP107E | 24 | ||
| 1 | CYP107E3,CYP125G1,CYP1004A1,CYP1004B1 | 4 | ||
| 8 | CYP107EU1 | 8 | ||
| 2 | CYP107FH4 | 2 | ||
| 1 | CYP107NE1 | 1 | ||
| 3 | CYP107Q4 | 3 | ||
| 17 | CYP107Q4,CYP105G5 | 34 | ||
| 6 | CYP113E1/2 | 6 | ||
| 1 | CYP113E2,CYP107EP2 | 2 | ||
| 2 | CYP1198B2 | 2 | ||
| 1 | CYP125G1 | 1 | ||
| 1 | CYP1278B-fragment2 | 1 | ||
| 5 | CYP154M5,CYP105CT1 | 10 | ||
| 1 | CYP154M5,CYP105CT1,CYP105G5,CYP105CP2 | 4 | ||
| 1 | CYP154M5,CYP105CT1,CYP107AY2-fragment | 3 | ||
| 1 | CYP154M5,CYP105CT2 | 2 | ||
| 1 | CYP1611B1,CYP2098A1 | 2 | ||
| 29 | CYP166A4 | 29 | ||
| 1 | CYP166A4,CYP107Q4,CYP105G5 | 3 | ||
| 70 | CYP208A | 70 | ||
| 1 | CYP208A28,CYP154M18 | 2 | ||
| 1 | CYP211C5 | 1 | ||
| 2 | CYP294A4 | 2 | ||
| T2PKS | 76 | 2 | CYP107NG1 | 2 |
| 1 | CYP107NH1 | 1 | ||
| 1 | CYP125G4 | 1 | ||
| 1 | CYP161T1 | 1 | ||
| 69 | CYP211C | 69 | ||
| 1 | CYP2296A2,CYP166A4,CYP173K1 | 3 | ||
| 1 | CYP244A5,CYP211C6 | 2 | ||
| T3PKS | 8 | 7 | CYP161N4,CYP2054A3,CYP107FH3 | 21 |
| 1 | CYP107FH3 | 1 | ||
| Terpene | 61 | 39 | CYP1051A | 39 |
| 2 | CYP105CT1 | 2 | ||
| 7 | CYP105CT1,CYP154M5 | 14 | ||
| 6 | CYP107AY | 6 | ||
| 4 | CYP107AY9,CYP244A10 | 8 | ||
| 1 | CYP107E37 | 1 | ||
| 1 | CYP154AJ2 | 1 | ||
| 1 | CYP154M5 | 1 | ||
| transAT-PKS | 1 | 1 | CYP113 × 1 | 1 |
| transAT-PKS-like | 8 | 8 | CYP163B | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malinga, N.A.; Nzuza, N.; Padayachee, T.; Syed, P.R.; Karpoormath, R.; Gront, D.; Nelson, D.R.; Syed, K. An Unprecedented Number of Cytochrome P450s Are Involved in Secondary Metabolism in Salinispora Species. Microorganisms 2022, 10, 871. https://doi.org/10.3390/microorganisms10050871
Malinga NA, Nzuza N, Padayachee T, Syed PR, Karpoormath R, Gront D, Nelson DR, Syed K. An Unprecedented Number of Cytochrome P450s Are Involved in Secondary Metabolism in Salinispora Species. Microorganisms. 2022; 10(5):871. https://doi.org/10.3390/microorganisms10050871
Chicago/Turabian StyleMalinga, Nsikelelo Allison, Nomfundo Nzuza, Tiara Padayachee, Puleng Rosinah Syed, Rajshekhar Karpoormath, Dominik Gront, David R. Nelson, and Khajamohiddin Syed. 2022. "An Unprecedented Number of Cytochrome P450s Are Involved in Secondary Metabolism in Salinispora Species" Microorganisms 10, no. 5: 871. https://doi.org/10.3390/microorganisms10050871
APA StyleMalinga, N. A., Nzuza, N., Padayachee, T., Syed, P. R., Karpoormath, R., Gront, D., Nelson, D. R., & Syed, K. (2022). An Unprecedented Number of Cytochrome P450s Are Involved in Secondary Metabolism in Salinispora Species. Microorganisms, 10(5), 871. https://doi.org/10.3390/microorganisms10050871