
Whole-Genome Investigation of Salmonella Dublin Considering Mountain Pastures as Reservoirs in Southern Bavaria, Germany


Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella Isolates
2.2. Epidemiological Analysis
2.3. Antimicrobial Susceptibility Testing
2.4. Whole-Genome Sequencing
2.5. Genetic Characterization
3. Results
3.1. Identification and Characterization of S. Dublin
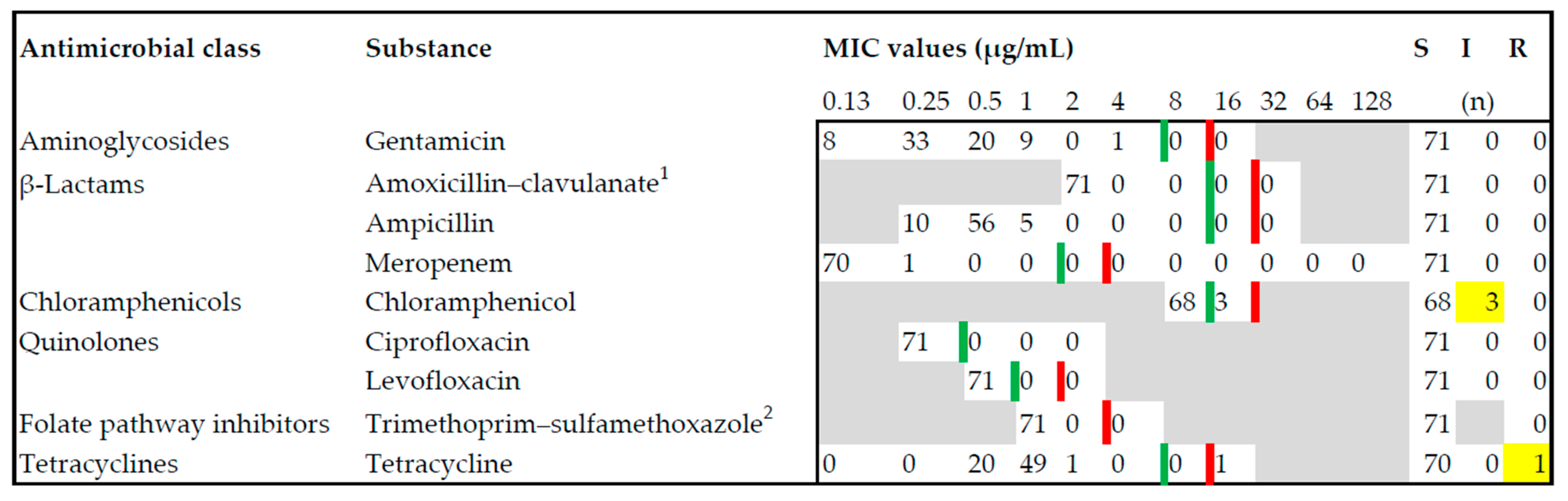
3.2. Antimicrobial Susceptibility
3.3. In Silico Analysis for Virulence and Plasmid Content
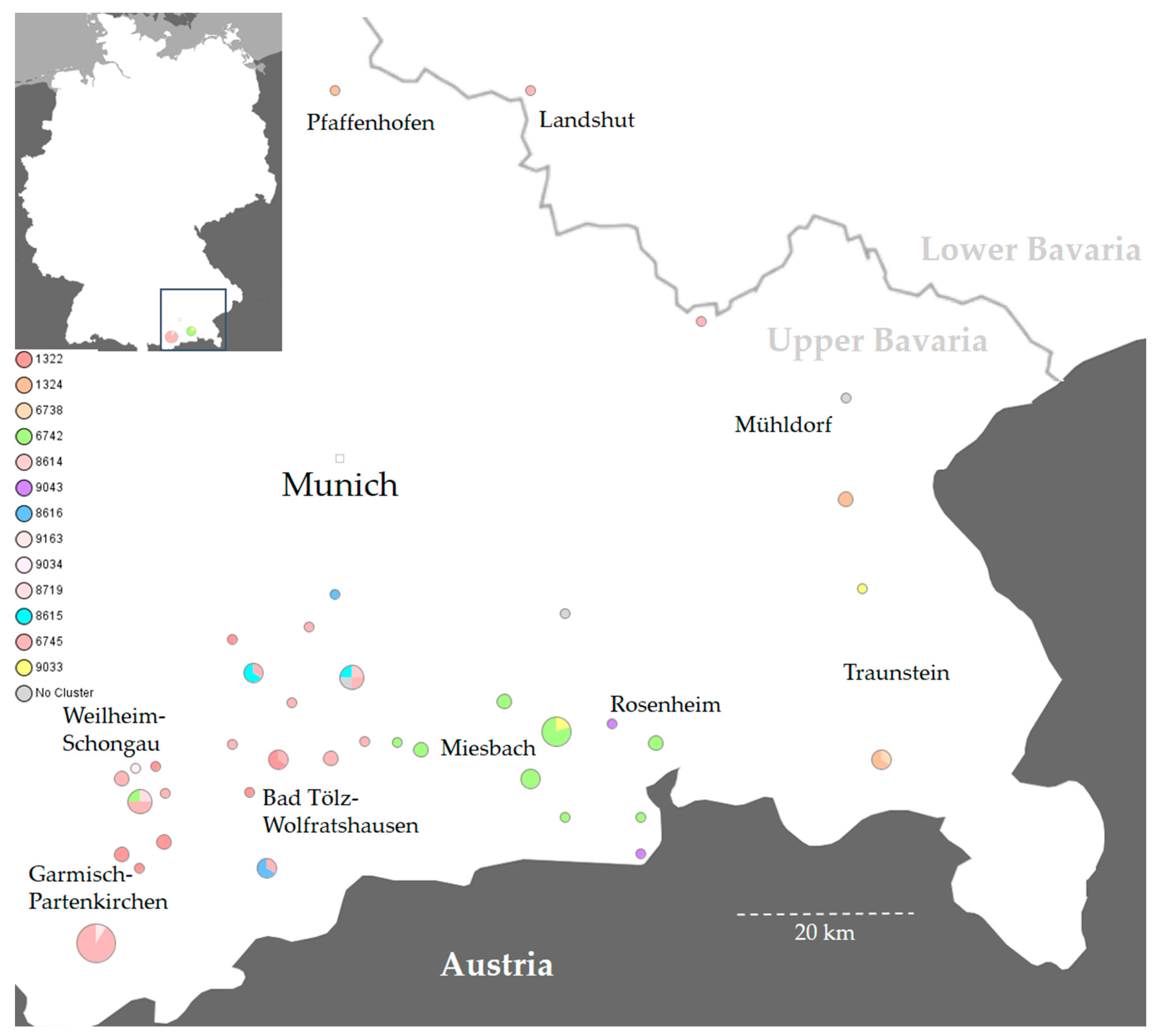
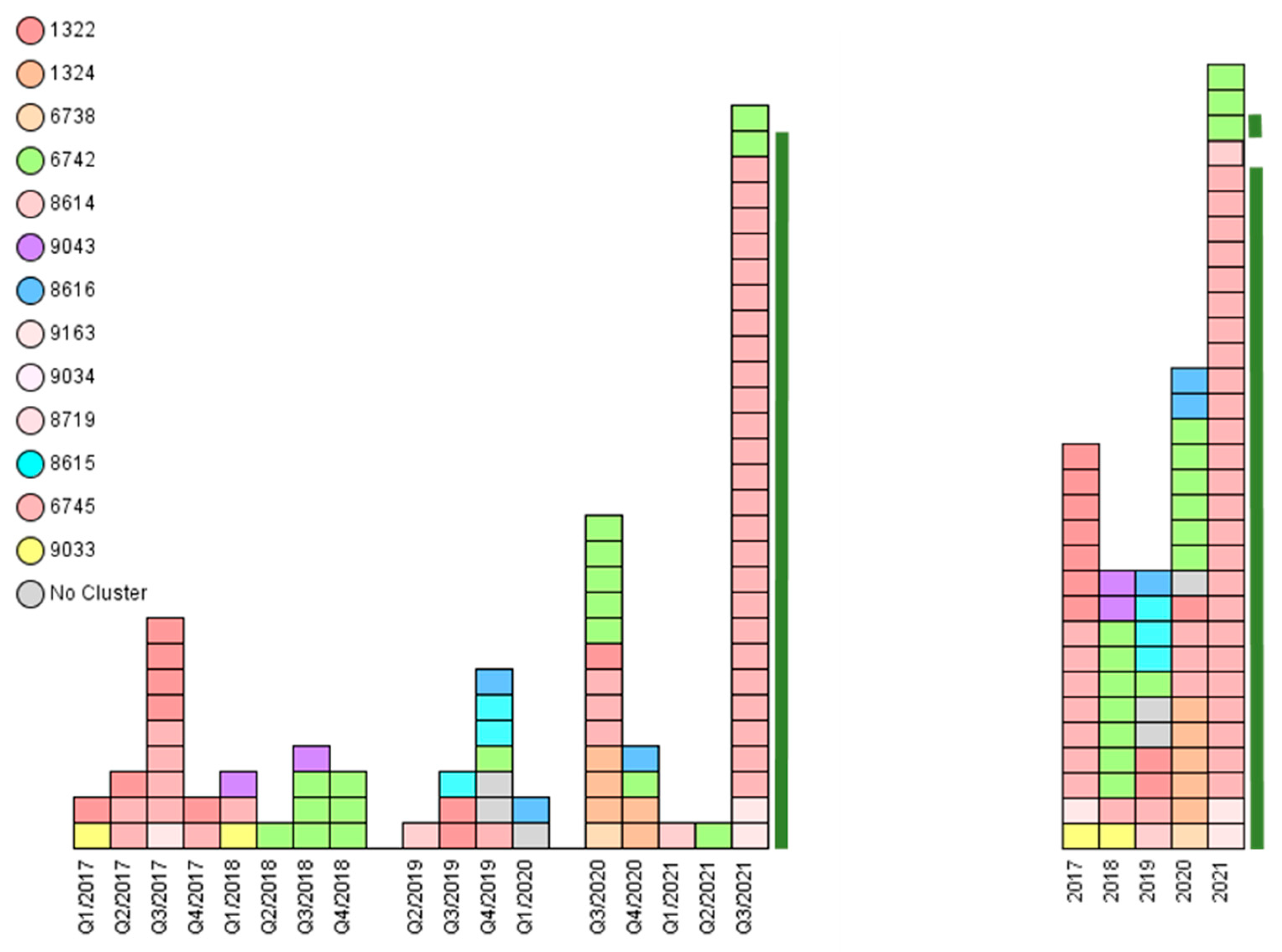
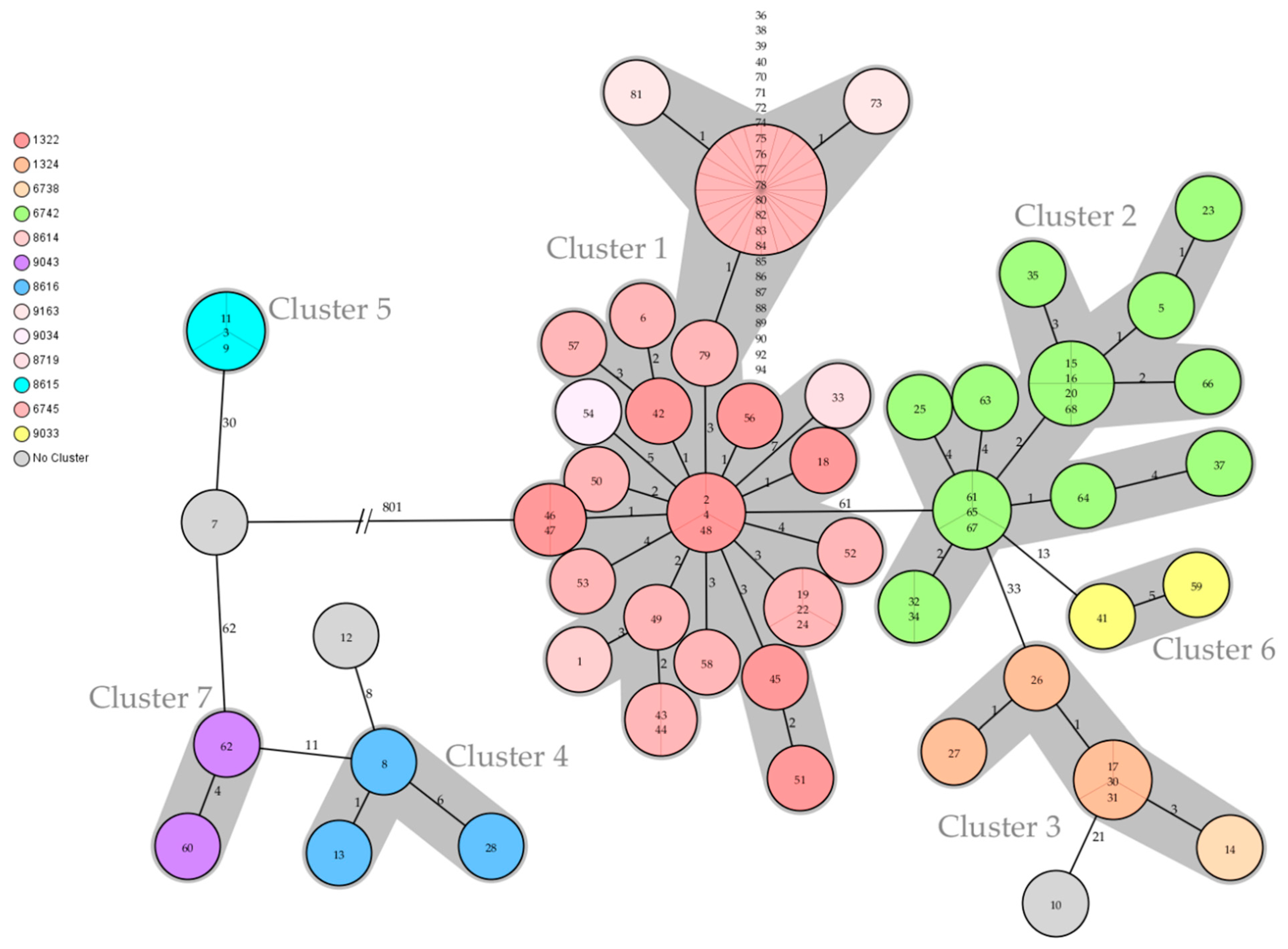
3.4. Genomic and Epidemiological Analysis of Animal Disease Outbreaks
4. Discussion
4.1. Challenges Regarding Routine Laboratory Diagnostics and Animal Disease Outbreak Investigation
4.2. Salmonella Dublin as a Zoonosis
4.3. Epidemiology with a Special Focus on Mountain Pastures
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schielke, A.; Rabsch, W.; Prager, R.; Simon, S.; Fruth, A.; Helling, R.; Schnabel, M.; Siffczyk, C.; Wieczorek, S.; Schroeder, S.; et al. Two consecutive large outbreaks of Salmonella Muenchen linked to pig farming in Germany, 2013 to 2014: Is something missing in our regulatory framework? EuroSurveill 2017, 22, 30528. [Google Scholar] [CrossRef] [PubMed]
- Elmberg, J.; Berg, C.; Lerner, H.; Waldenstrom, J.; Hessel, R. Potential disease transmission from wild geese and swans to livestock, poultry and humans: A review of the scientific literature from a One Health perspective. Infect. Ecol. Epidemiol. 2017, 7, 1300450. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, K.; Nevarez, J.G.; Del Piero, F. Salmonella Enterica Serovar Pomona Infection in Farmed Juvenile American Alligators (Alligator Mississippiensis). Vet. Pathol. 2017, 54, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Zajac, M.; Skarzynska, M.; Lalak, A.; Kwit, R.; Smialowska-Weglinska, A.; Pasim, P.; Szulowski, K.; Wasyl, D. Salmonella in Captive Reptiles and Their Environment-Can We Tame the Dragon? Microorganisms 2021, 9, 1012. [Google Scholar] [CrossRef] [PubMed]
- Bonifait, L.; Thepault, A.; Bauge, L.; Rouxel, S.; Le Gall, F.; Chemaly, M. Occurrence of Salmonella in the Cattle Production in France. Microorganisms 2021, 9, 872. [Google Scholar] [CrossRef] [PubMed]
- Wissmann, J.E.; Kirchhoff, L.; Bruggemann, Y.; Todt, D.; Steinmann, J.; Steinmann, E. Persistence of Pathogens on Inanimate Surfaces: A Narrative Review. Microorganisms 2021, 9, 343. [Google Scholar] [CrossRef]
- Nielsen, L.R. Review of pathogenesis and diagnostic methods of immediate relevance for epidemiology and control of Salmonella Dublin in cattle. Vet. Microbiol. 2013, 162, 1–9. [Google Scholar] [CrossRef]
- Field, H.I. A survey of bovine salmonellosis in mid and west Wales. Br. Vet. J. 1948, 104, 323–339. [Google Scholar] [CrossRef]
- McCaughey, W.; McClelland, T.; Hanna, J. Some observations on Salmonella dublin infection in clinically healthy beef cattle. Br. Vet. J. 1971, 127, 549–556. [Google Scholar] [CrossRef]
- Nielsen, L.R.; Schukken, Y.H.; Grohn, Y.T.; Ersboll, A.K. Salmonella Dublin infection in dairy cattle: Risk factors for becoming a carrier. Prev. Vet. Med. 2004, 65, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Allerberger, F.; Liesegang, A.; Grif, K.; Khaschabi, D.; Prager, R.; Danzl, J.; Höck, F.; Öttl, J.; Dierich, M.P.; Berghold, C. Occurrence of Salmonella enterica serovar Dublin in Austria. Wien. Med. Wochenschr. 2003, 153, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Wray, C.; Sojka, W.J. Reviews of the progress of dairy science: Bovine salmonellosis. J. Dairy Res. 1977, 44, 383–425. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.K.; Brennand, J. Bacteraemia in salmonellosis: A 15 year retrospective study from a regional infectious diseases unit. BMJ 1988, 297, 1242–1243. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.F.; Ingram, L.A.; Cieslak, P.R.; Vugia, D.J.; Tobin-D’Angelo, M.; Hurd, S.; Medus, C.; Cronquist, A.; Angulo, F.J. Salmonellosis outcomes differ substantially by serotype. J. Infect. Dis. 2008, 198, 109–114. [Google Scholar] [CrossRef]
- Fang, F.C.; Fierer, J. Human infection with Salmonella dublin. Medicine 1991, 70, 198–207. [Google Scholar] [CrossRef]
- Helms, M.; Vastrup, P.; Gerner-Smidt, P.; Molbak, K. Short and long term mortality associated with foodborne bacterial gastrointestinal infections: Registry based study. BMJ 2003, 326, 357. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.R.; Friedman, C.R.; Crim, S.M.; Judd, M.; Barrett, K.A.; Tolar, B.; Folster, J.P.; Griffin, P.M.; Brown, A.C. Epidemiology of Salmonella enterica Serotype Dublin Infections among Humans, United States, 1968–2013. Emerg. Infect. Dis. 2017, 23, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- McDonough, P.L.; Fogelman, D.; Shin, S.J.; Brunner, M.A.; Lein, D.H. Salmonella enterica serotype Dublin infection: An emerging infectious disease for the northeastern United States. J. Clin. Microbiol. 1999, 37, 2418–2427. [Google Scholar] [CrossRef] [Green Version]
- van Asten, A.J.; van Dijk, J.E. Distribution of “classic” virulence factors among Salmonella spp. FEMS Immunol. Med. Microbiol. 2005, 44, 251–259. [Google Scholar] [CrossRef]
- Van Schaik, G.; Klinkenberg, D.; Veling, J.; Stegeman, A. Transmission of Salmonella in dairy herds quantified in the endemic situation. Vet. Res. 2007, 38, 861–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methner, U. Salmonellose der Rinder. In Tiergesundheitsjahresbericht 2017; Korr, F., Ed.; Tiergesundheitsjahresbericht; Friedrich-Loeffler-Inst.: Greifswald, Germany, 2018; Volume 18, p. 105111. [Google Scholar]
- Methner, U. Salmonellose der Rinder. In Tiergesundheitsjahresbericht 2019; Tiergesundheitsjahresbericht; Friedrich-Loeffler-Inst.: Greifswald, Germany, 2020; Volume 20, pp. 92–98. [Google Scholar]
- Robert-Koch-Institut. SurvStat@RKI 2.0. Available online: https://survstat.rki.de (accessed on 24 October 2021).
- The Federal Ministry of Justice. Regulation on the Protection against Salmonellosis in Cattle (Cattle Salmonellosis Regulation) of 14. November 1991 (BGBl. I p. 2118), Amended by Article 2 of 17. April 2014 (BGBl. I p. 388); Bundesministerium der Justiz: Berlin, Germany, 1991.
- Agren, E.C.; Wahlstrom, H.; Vesterlund-Carlson, C.; Lahti, E.; Melin, L.; Soderlund, R. Comparison of whole genome sequencing typing results and epidemiological contact information from outbreaks of Salmonella Dublin in Swedish cattle herds. Infect. Ecol. Epidemiol. 2016, 6, 31782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Committee for Standardization. Microbiology of the Food Chain-Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella-Part 1: Detection of Salmonella Spp., German version EN ISO 6579-1:2017 + A1:2020; Beuth Verlag GmbH: Berlin, Germany, 2020; p. 66. [Google Scholar] [CrossRef]
- Grimont, P.A.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars; WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007; Volume 9, pp. 1–166. [Google Scholar]
- Garcia-Soto, S.; Tomaso, H.; Linde, J.; Methner, U. Epidemiological Analysis of Salmonella Enterica Subsp. Enterica Serovar Dublin in German Cattle Herds Using Whole-Genome Sequencing. Microbiol. Spectr. 2021, 9, e0033221. [Google Scholar] [CrossRef] [PubMed]
- The Bavarian State Ministry of Food, Agriculture and Forestry. Herkunftssicherungs- und Informationssystem für Tiere (HIT). Available online: https://www3.hi-tier.de/HitCom/login.asp (accessed on 28 February 2022).
- Friedrich-Loeffler-Institut. TierSeuchenNachrichten-System (TSN). Available online: https://tsn.fli.de/Auth/Forms/Login.Aspx?ReturnUrl=%2f (accessed on 28 February 2022).
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; CLSI Supplement VET01S; Clinical and Laboratory Standards Institute: New York, NY, USA, 2020. [Google Scholar]
- Technical University of Denmark. SeqSero 1.2. Available online: https://cge.cbs.dtu.dk/services/SeqSero/ (accessed on 20 October 2021).
- Zhang, S.; Yin, Y.; Jones, M.B.; Zhang, Z.; Deatherage Kaiser, B.L.; Dinsmore, B.A.; Fitzgerald, C.; Fields, P.I.; Deng, X. Salmonella serotype determination utilizing high-throughput genome sequencing data. J. Clin. Microbiol. 2015, 53, 1685–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Technical University of Denmark. ResFinder 4.1. Available online: https://cge.cbs.dtu.dk/services/ResFinder/ (accessed on 7 February 2022).
- Institute of Pathogen Biology; CAMS & PUMC. Virulence Factor Database (VFDB). Available online: http://www.mgc.ac.cn/VFs/main.htm (accessed on 3 February 2022).
- Technical University of Denmark. PlasmidFinder 2.1. Available online: https://cge.cbs.dtu.dk/services/PlasmidFinder/ (accessed on 4 February 2022).
- Uelze, L.; Becker, N.; Borowiak, M.; Busch, U.; Dangel, A.; Deneke, C.; Fischer, J.; Flieger, A.; Hepner, S.; Huber, I.; et al. Toward an Integrated Genome-Based Surveillance of Salmonella enterica in Germany. Front. Microbiol. 2021, 12, 626941. [Google Scholar] [CrossRef]
- Richardson, A.; Fawcett, A.R. Salmonella dublin infection in calves: The value of rectal swabs in diagnosis and epidemiological studies. Br. Vet. J. 1973, 129, 151–156. [Google Scholar] [CrossRef]
- Uzzau, S.; Brown, D.J.; Wallis, T.; Rubino, S.; Leori, G.; Bernard, S.; Casadesus, J.; Platt, D.J.; Olsen, J.E. Host adapted serotypes of Salmonella enterica. Epidemiol. Infect. 2000, 125, 229–255. [Google Scholar] [CrossRef]
- Herikstad, H.; Motarjemi, Y.; Tauxe, R.V. Salmonella surveillance: A global survey of public health serotyping. Epidemiol. Infect. 2002, 129, 1–8. [Google Scholar] [CrossRef]
- Wattiau, P.; Boland, C.; Bertrand, S. Methodologies for Salmonella enterica subsp. enterica subtyping: Gold standards and alternatives. Appl. Environ. Microbiol. 2011, 77, 7877–7885. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; den Bakker, H.C.; Li, S.; Chen, J.; Dinsmore, B.A.; Lane, C.; Lauer, A.C.; Fields, P.I.; Deng, X. SeqSero2: Rapid and Improved Salmonella Serotype Determination Using Whole-Genome Sequencing Data. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Ibrahim, G.M.; Morin, P.M. Salmonella serotyping using whole genome sequencing. Front. Microbiol. 2018, 9, 2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, M.; Le Hello, S.; Leekitcharoenphon, P.; Hendriksen, R. The invasome of Salmonella Dublin as revealed by whole genome sequencing. BMC Infect. Dis. 2017, 17, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srednik, M.E.; Lantz, K.; Hicks, J.A.; Morningstar-Shaw, B.R.; Mackie, T.A.; Schlater, L.K. Antimicrobial resistance and genomic characterization of Salmonella Dublin isolates in cattle from the United States. PLoS ONE 2021, 16, e0249617. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, e07209. [Google Scholar] [CrossRef]
- Paudyal, N.; Pan, H.; Elbediwi, M.; Zhou, X.; Peng, X.; Li, X.; Fang, W.; Yue, M. Characterization of Salmonella Dublin isolated from bovine and human hosts. BMC Microbiol. 2019, 19, 226. [Google Scholar] [CrossRef]
- Carroll, L.M.; Wiedmann, M.; den Bakker, H.; Siler, J.; Warchocki, S.; Kent, D.; Lyalina, S.; Davis, M.; Sischo, W.; Besser, T.; et al. Whole-Genome Sequencing of Drug-Resistant Salmonella enterica Isolates from Dairy Cattle and Humans in New York and Washington States Reveals Source and Geographic Associations. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Fenske, G.J.; Thachil, A.; McDonough, P.L.; Glaser, A.; Scaria, J. Geography Shapes the Population Genomics of Salmonella enterica Dublin. Genome Biol. Evol. 2019, 11, 2220–2231. [Google Scholar] [CrossRef] [Green Version]
- Kudirkiene, E.; Sorensen, G.; Torpdahl, M.; de Knegt, L.V.; Nielsen, L.R.; Rattenborg, E.; Ahmed, S.; Olsen, J.E. Epidemiology of Salmonella enterica Serovar Dublin in Cattle and Humans in Denmark, 1996 to 2016: A Retrospective Whole-Genome-Based Study. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- The Federal Ministry of Food and Agriculture (BMEL). Report of the Federal Ministry of Food and Agriculture on the Evaluation of the Antibiotics Minimisation Concept introduced with the 16th Act to Amend the Medicinal Products Act. (16th AMG Amendment); BMEL: Bonn, Germany, 2019.
- The European Parliament and the Council of the European Union. Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on veterinary medicinal products and repealing Directive 2001/82/EC. Article 107. 2018. [Google Scholar]
- Foster, N.; Tang, Y.; Berchieri, A.; Geng, S.; Jiao, X.; Barrow, P. Revisiting Persistent Salmonella Infection and the Carrier State: What Do We Know? Pathogens 2021, 10, 1299. [Google Scholar] [CrossRef]
- Methner, U. Salmonellose der Rinder. Amtstierärztlicher Dienst Und Lebensm. 2012, 4, 1–4. [Google Scholar]
- Vaillant, V.; Haeghebaert, S.; Desenclos, J.C.; Bouvet, P.; Grimont, F.; Grimont, P.A.; Burnens, A.P. Outbreak of Salmonella dublin infection in France, November–December 1995. EuroSurveill 1996, 1, 9–10. [Google Scholar] [CrossRef] [PubMed]
- The Bavarian State Ministry of Food, Agriculture and Forestry. Alm-und Alpwirtschaft in Bayern; Bayerisches Staatsministerium für Ernährung: Landwirtschaft und Forsten, Bayern, 2010. [Google Scholar]
- Krogmeier, D.; Kimmerle, A.; Schmidt, E.; Götz, K. The effect of Alpine pasturing of heifers on the longevity and on different yield traits of cows in Simmental and Brown Swiss cattle. Züchtungskunde 2015, 87, 107–119. [Google Scholar]
- Wray, C.; Todd, N.; McLaren, I.M.; Beedell, Y.E. The epidemiology of Salmonella in calves: The role of markets and vehicles. Epidemiol. Infect. 1991, 107, 521–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronstol, H.; Osborne, A.D.; Pethiyagoda, S. Experimental Salmonella infection in calves. 2. Virulence and the spread of infection. J. Hyg. 1974, 72, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, C.; Davies, R.H. Salmonella Infections in Cattle. In Salmonella in Domestic Animals; Wray, C., Wray, A., Eds.; CABI Publishing: Oxon, UK, 2000; pp. 169–190. [Google Scholar]
- Maynaud, G.; Pourcher, A.M.; Ziebal, C.; Cuny, A.; Druilhe, C.; Steyer, J.P.; Wery, N. Persistence and Potential Viable but Non-culturable State of Pathogenic Bacteria during Storage of Digestates from Agricultural Biogas Plants. Front. Microbiol. 2016, 7, 1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boqvist, S.; Vagsholm, I. Risk factors for hazard of release from Salmonella-control restriction on Swedish cattle farms from 1993 to 2002. Prev. Vet. Med. 2005, 71, 35–44. [Google Scholar] [CrossRef]
- Nielsen, L.R.; Kudahl, A.B.; Ostergaard, S. Age-structured dynamic, stochastic and mechanistic simulation model of Salmonella Dublin infection within dairy herds. Prev. Vet. Med. 2012, 105, 59–74. [Google Scholar] [CrossRef]
- de Knegt, L.V.; Kudirkiene, E.; Rattenborg, E.; Sorensen, G.; Denwood, M.J.; Olsen, J.E.; Nielsen, L.R. Combining Salmonella Dublin genome information and contact-tracing to substantiate a new approach for improved detection of infectious transmission routes in cattle populations. Prev. Vet. Med. 2020, 181, 104531. [Google Scholar] [CrossRef]
- Mohammed, M.; Cormican, M. Whole genome sequencing provides insights into the genetic determinants of invasiveness in Salmonella Dublin. Epidemiol. Infect. 2016, 144, 2430–2439. [Google Scholar] [CrossRef] [Green Version]
- Watson, W.A.; Wood, B.; Richardson, A. Salmonella dublin infection in a beef herd. Br. Vet. J. 1971, 127, 294–298. [Google Scholar] [CrossRef]
- Liebana, E.; Garcia-Migura, L.; Clouting, C.; Cassar, C.A.; Clifton-Hadley, F.A.; Lindsay, E.A.; Threlfall, E.J.; Chappell, S.A.; Davies, R.H. Investigation of the genetic diversity among isolates of Salmonella enterica serovar Dublin from animals and humans from England, Wales and Ireland. J. Appl. Microbiol. 2002, 93, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Dietz, H.H.; Chriel, M.; Andersen, T.H.; Jorgensen, J.C.; Torpdahl, M.; Pedersen, H.; Pedersen, K. Outbreak of Salmonella Dublin-associated abortion in Danish fur farms. Can. Vet. J. 2006, 47, 1201–1205. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
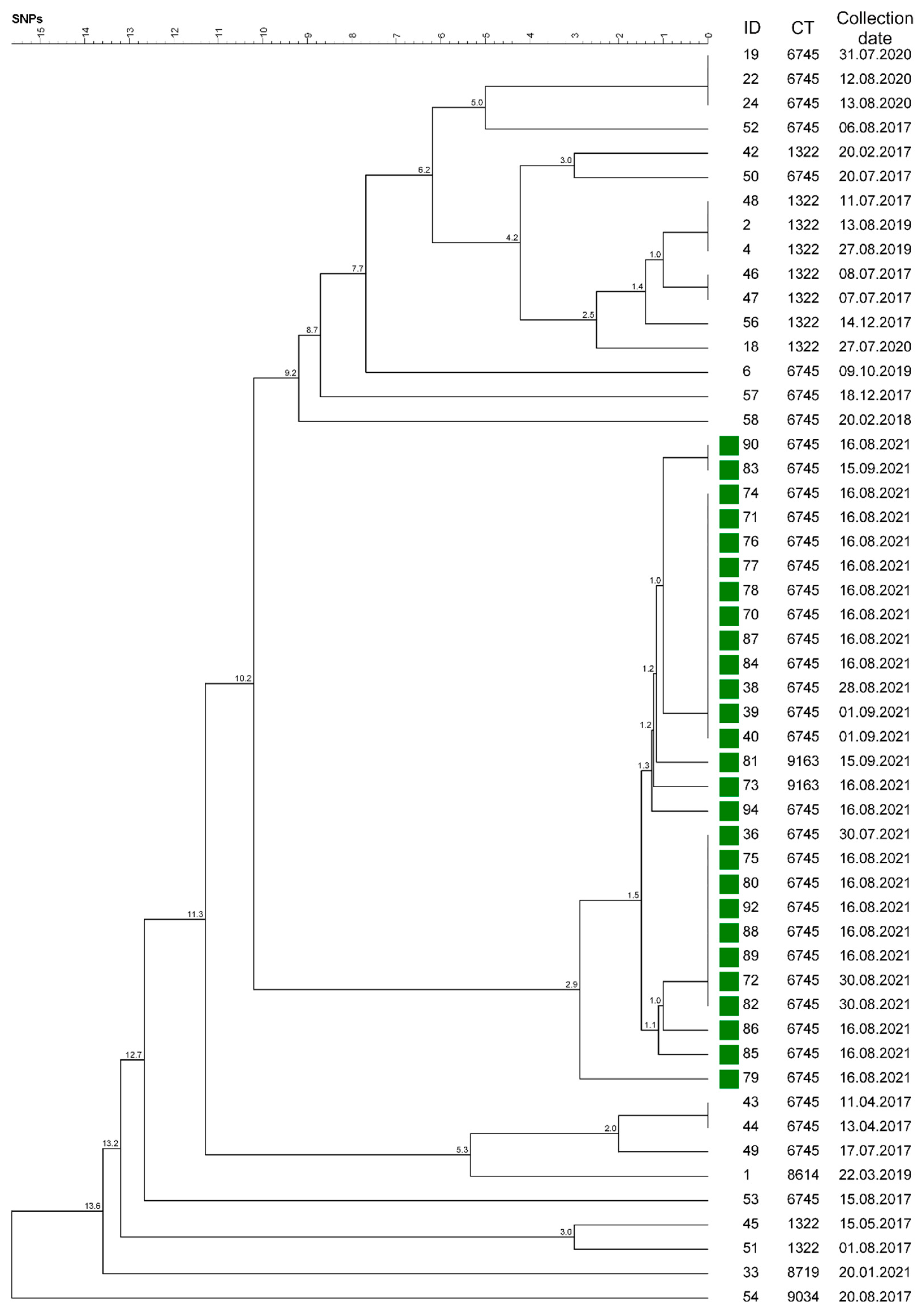
| Cluster | Complex Type | Isolate Number | Year | Host | Number of Farms | District |
|---|---|---|---|---|---|---|
| 1 | 1322 | 42, 51, 56 | 2017 | cattle | 3 | TÖL |
| 1 | 1322 | 45 | 2017 | cattle | 1 | WM |
| 1 | 1322 | 46, 47, 48 | 2017 | cattle | 3 | GAP |
| 1 | 1322 | 2, 4 | 2019 | cattle, dog | 1 | GAP |
| 1 | 1322 | 18 | 2020 | cattle | 1 | TÖL |
| 1 | 6745 | 43, 49, 50, 53 | 2017 | cattle | 4 | TÖL |
| 1 | 6745 | 44 | 2017 | cattle | 1 | MÜ |
| 1 | 6745 | 57 | 2017 | cattle | 1 | WM |
| 1 | 6745 | 52 | 2017 | cattle | 1, mp-GAP | GAP |
| 1 | 6745 | 58 | 2018 | cattle | 1 | TÖL |
| 1 | 6745 | 6 | 2019 | cattle | 1 | TÖL |
| 1 | 6745 | 19, 24 | 2020 | cattle | 2 | TÖL |
| 1 | 6745 | 22 | 2020 | pig | 1 | LA |
| 1 | 6745 | 36 | 2021 | cattle | 1 | TÖL |
| 1 | 6745 | 39, 40, 70–72, 74–78, 80, 82, 84, 85–90, 92 | 2021 | cattle | 6, mp-GAP | GAP |
| 1 | 6745 | 38, 83, 94 | 2021 | dog | mp-GAP | GAP |
| 1 | 6745 | 79 | 2021 | cattle | mp-GAP | GAP |
| 1 | 8614 | 1 | 2019 | cattle | 1 | TÖL |
| 1 | 8719 | 33 | 2021 | cattle | 1 | GAP |
| 1 | 9034 | 54 | 2017 | cattle | 1 | WM |
| 1 | 9163 | 73, 81 | 2021 | cattle | 2, mp-GAP | GAP |
| 2 | 6742 | 61, 66 | 2018 | bovine | 1 | RO |
| 2 | 6742 | 63, 64, 65, 67, 68 | 2018 | cattle | 5 | MB |
| 2 | 6742 | 5 | 2019 | cattle | 1 | RO |
| 2 | 6742 | 15 *, 16 *, 23 | 2020 | cattle | 3, mp-MB * | RO |
| 2 | 6742 | 20 *, 25, 32 | 2020 | cattle | 3, mp-MB * | MB |
| 2 | 6742 | 34, 35 | 2021 | cattle | 2 | MB |
| 2 | 6742 | 37 | 2021 | cattle | 1, mp-GAP | GAP |
| 3 | 6738, 1324 | 14, 17 | 2020 | cattle | 1 | TS |
| 3 | 1324 | 26, 30, 31 | 2020 | cattle | 2 | TS |
| 3 | 1324 | 27 | 2020 | pig | 1 | PAF |
| 4 | 8616 | 8 | 2019 | cattle | 1 | TÖL |
| 4 | 8616 | 13 | 2020 | cattle | 1 | M |
| 4 | 8616 | 28 | 2020 | bovine | 1 | TÖL |
| 5 | 8615 | 3, 9, 11 | 2019 | cattle | 2 | TÖL |
| 6 | 9033 | 41 | 2017 | cattle | 1 | RO |
| 6 | 9033 | 59 | 2018 | cattle | 1 | TS |
| 7 | 9043 | 60, 62 | 2018 | cattle | 2 | RO |
| none | 8620 | 7 | 2019 | cattle | 1 | RO |
| none | 8623 | 12 | 2019 | cattle | 1 | TÖL |
| none | 8626 | 10 | 2019 | cattle | 1 | MÜ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klose, C.; Scuda, N.; Ziegler, T.; Eisenberger, D.; Hanczaruk, M.; Riehm, J.M. Whole-Genome Investigation of Salmonella Dublin Considering Mountain Pastures as Reservoirs in Southern Bavaria, Germany. Microorganisms 2022, 10, 885. https://doi.org/10.3390/microorganisms10050885
Klose C, Scuda N, Ziegler T, Eisenberger D, Hanczaruk M, Riehm JM. Whole-Genome Investigation of Salmonella Dublin Considering Mountain Pastures as Reservoirs in Southern Bavaria, Germany. Microorganisms. 2022; 10(5):885. https://doi.org/10.3390/microorganisms10050885
Chicago/Turabian StyleKlose, Corinna, Nelly Scuda, Tobias Ziegler, David Eisenberger, Matthias Hanczaruk, and Julia M. Riehm. 2022. "Whole-Genome Investigation of Salmonella Dublin Considering Mountain Pastures as Reservoirs in Southern Bavaria, Germany" Microorganisms 10, no. 5: 885. https://doi.org/10.3390/microorganisms10050885
APA StyleKlose, C., Scuda, N., Ziegler, T., Eisenberger, D., Hanczaruk, M., & Riehm, J. M. (2022). Whole-Genome Investigation of Salmonella Dublin Considering Mountain Pastures as Reservoirs in Southern Bavaria, Germany. Microorganisms, 10(5), 885. https://doi.org/10.3390/microorganisms10050885