
The Response Regulator RegA Is a Copper Binding Protein That Covalently Dimerizes When Exposed to Oxygen



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overexpression of RegA
2.2. Purification of RegA from E. coli
2.3. Loading RegA with Metal through Buffer Exchange
2.4. ICP-MS
2.5. ESI-MS
2.6. LC-MS/MS
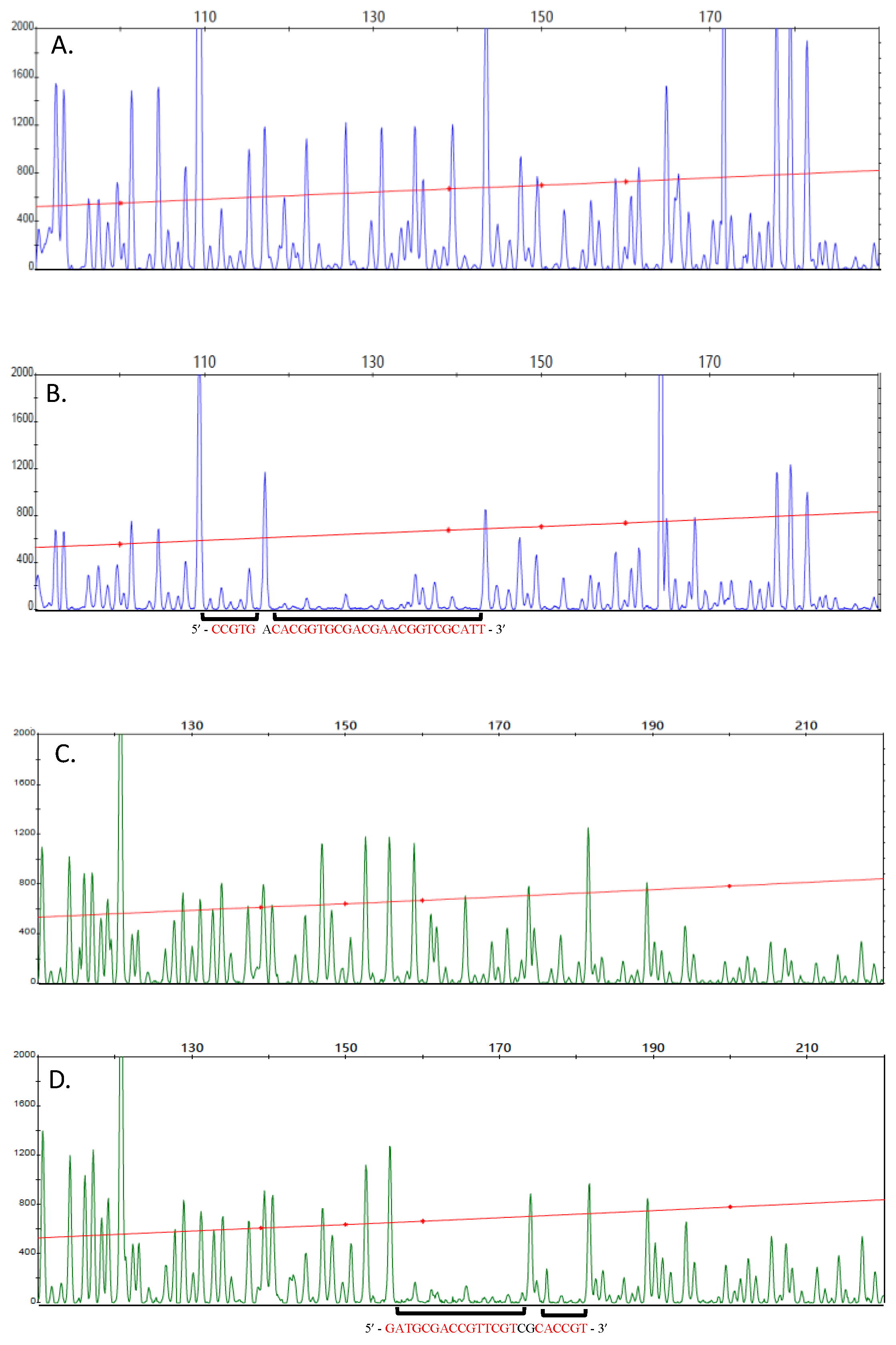
2.7. DNase I FootPrinting Assay
3. Results
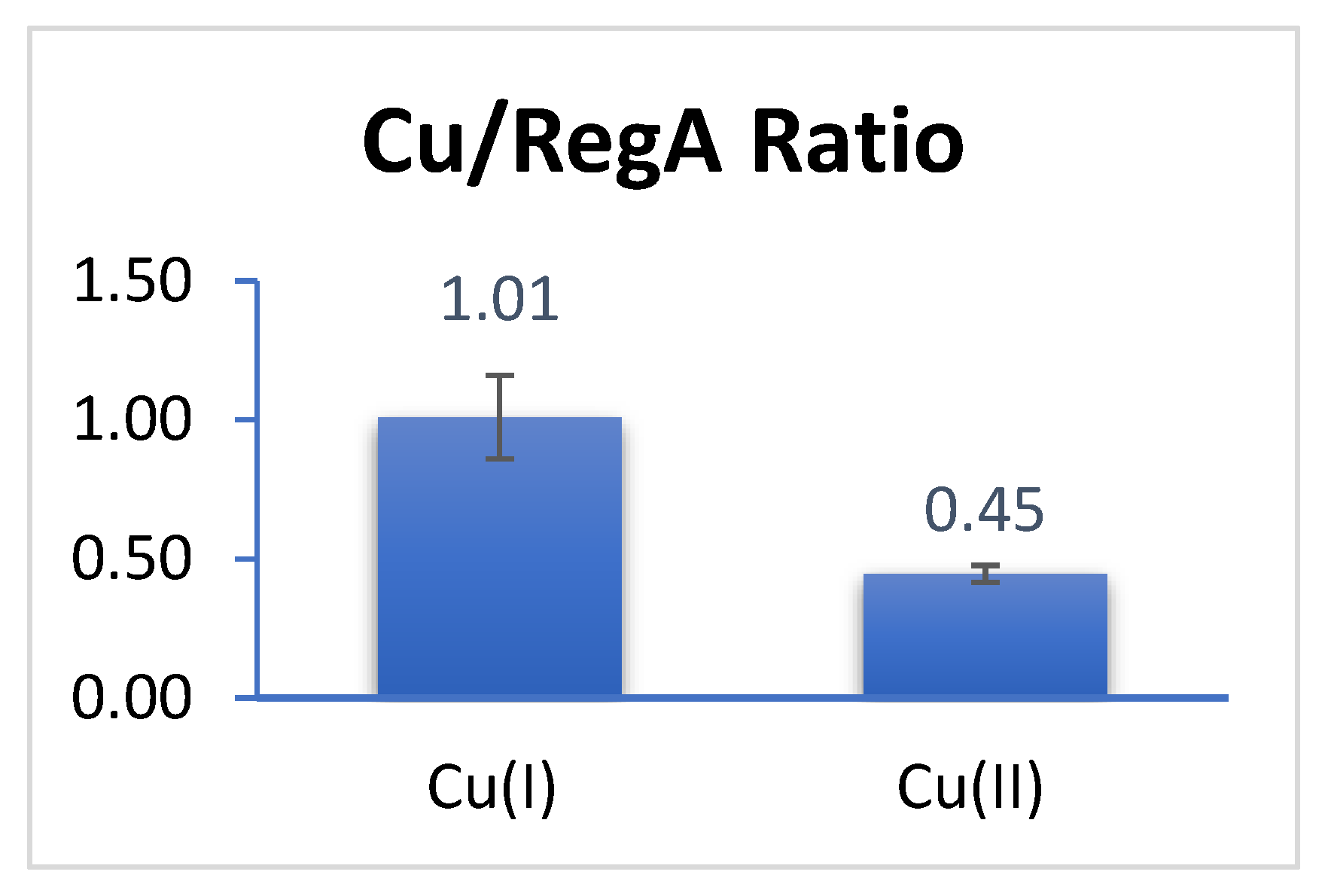
3.1. RegA Binds Copper Anaerobically
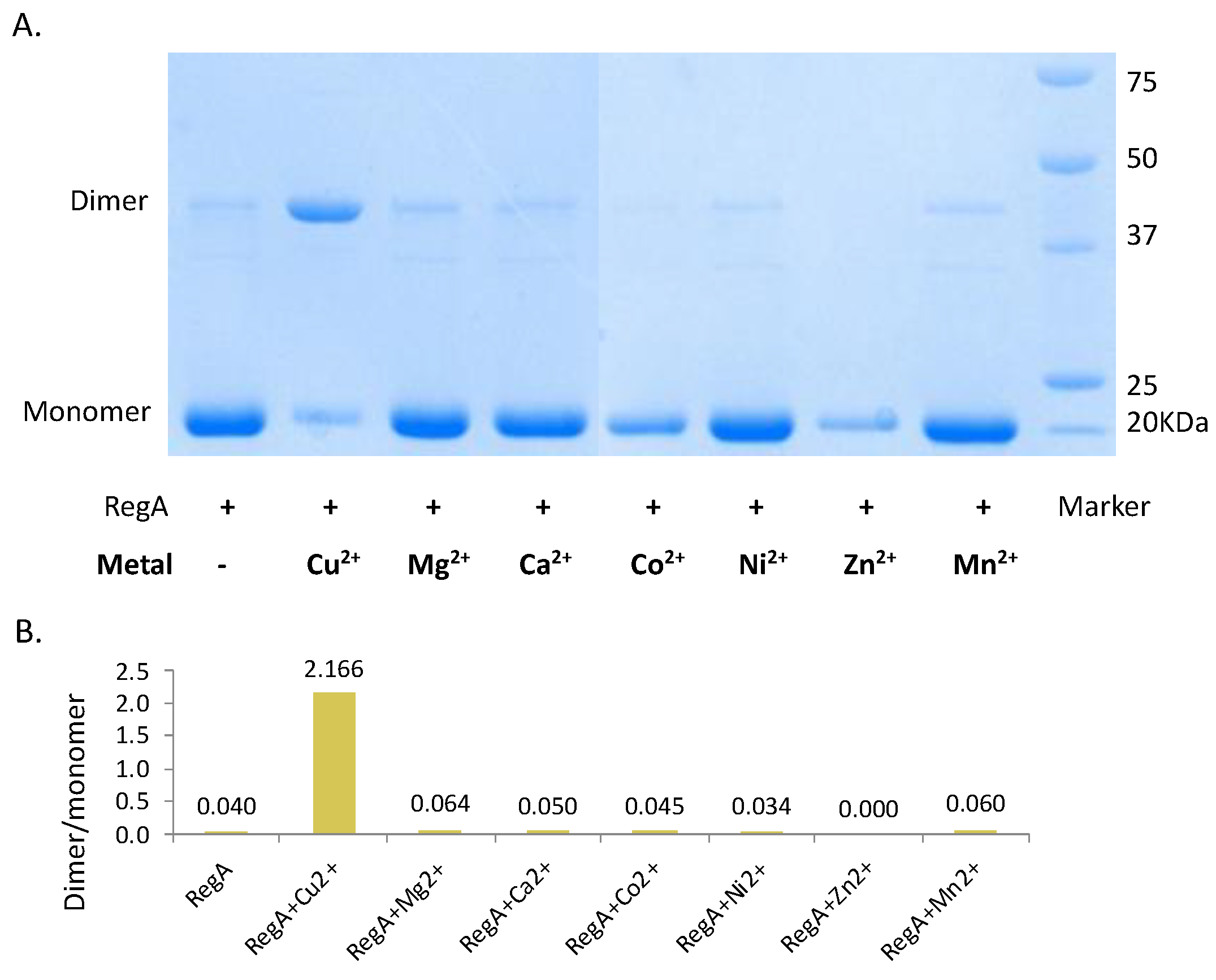
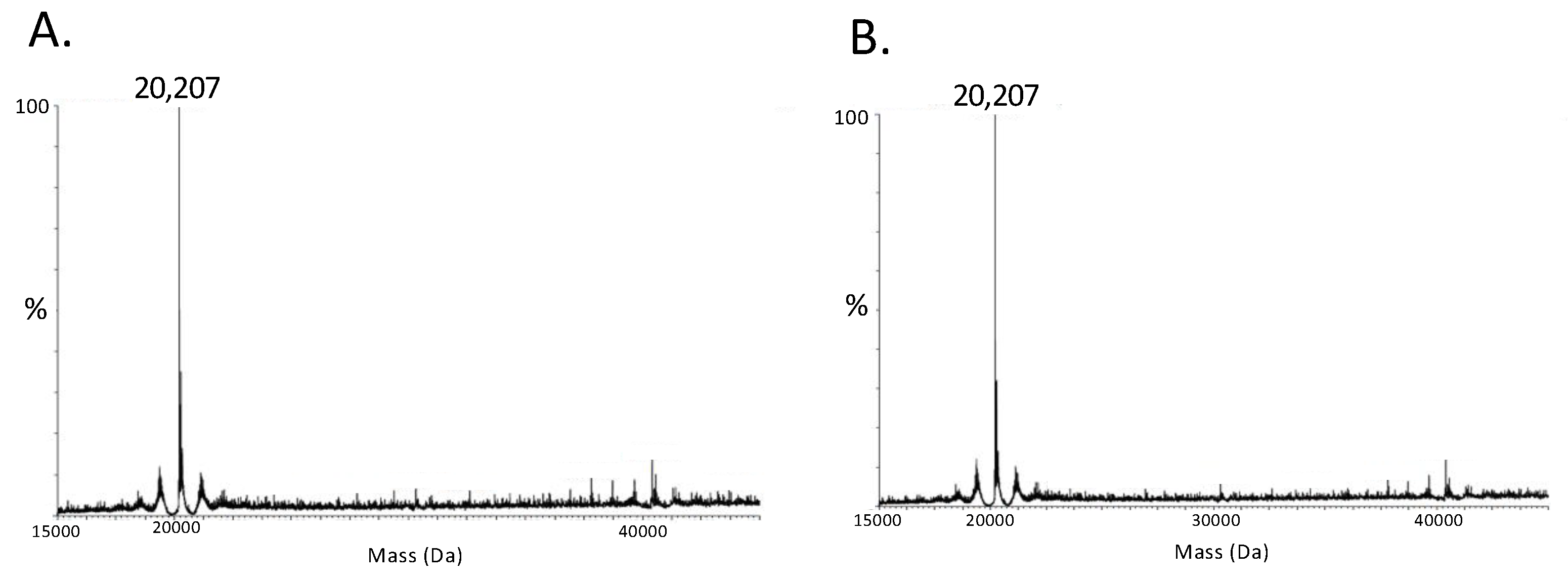
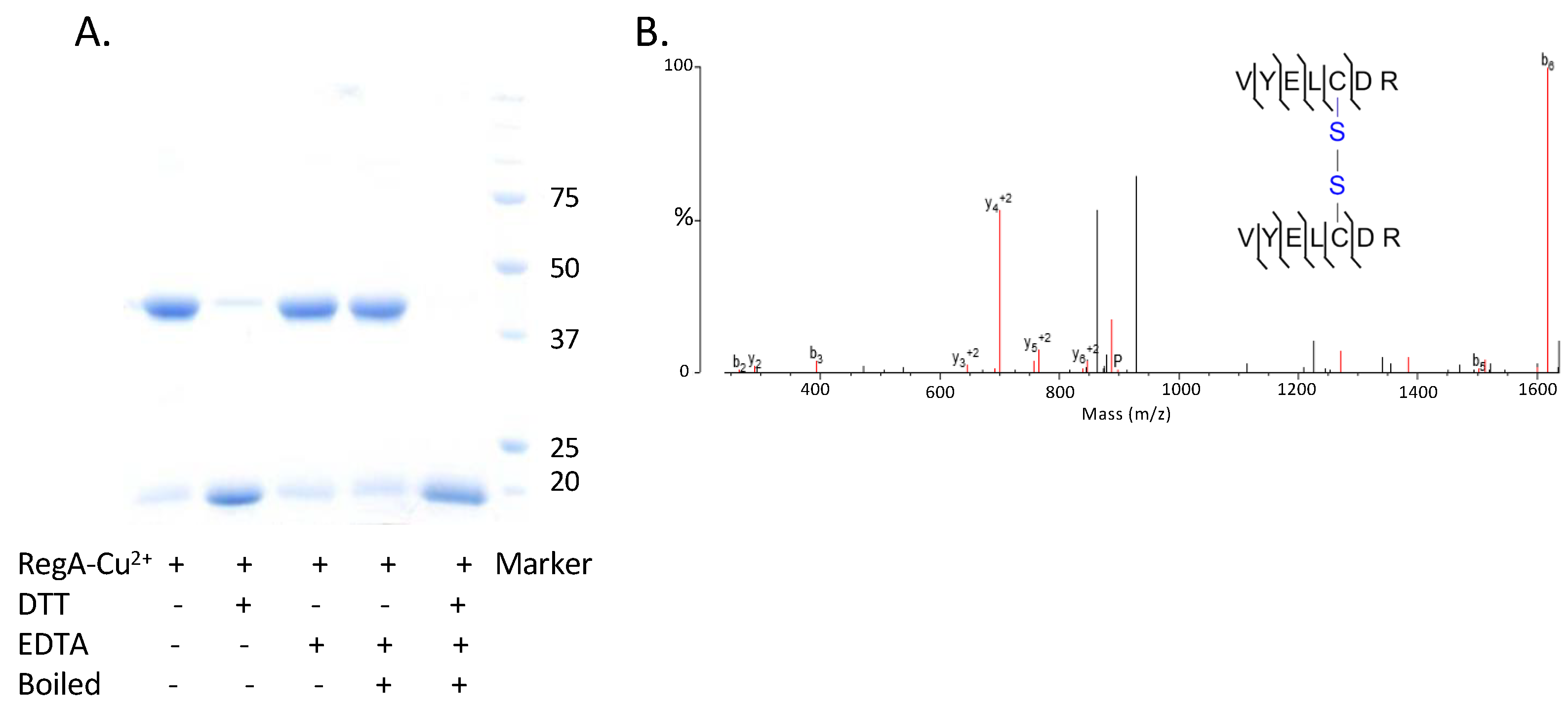
3.2. Cu2+ Promotes the Dimerization of RegA Aerobically
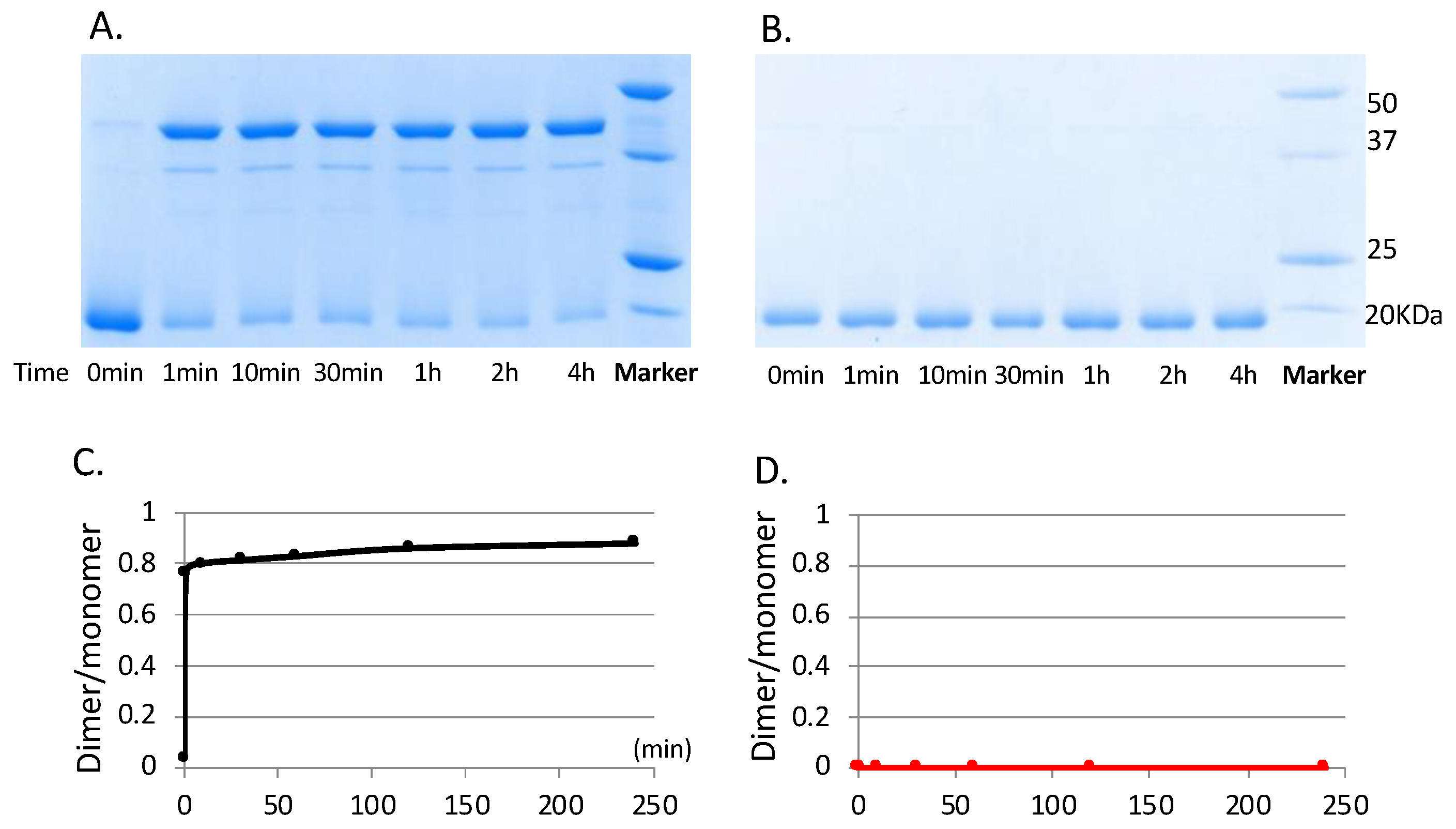
3.3. RegA Dimerization Requires Both Cu2+ and Molecular Oxygen
3.4. Covalently Dimerized RegA Is Capable of Binding DNA
4. Discussion
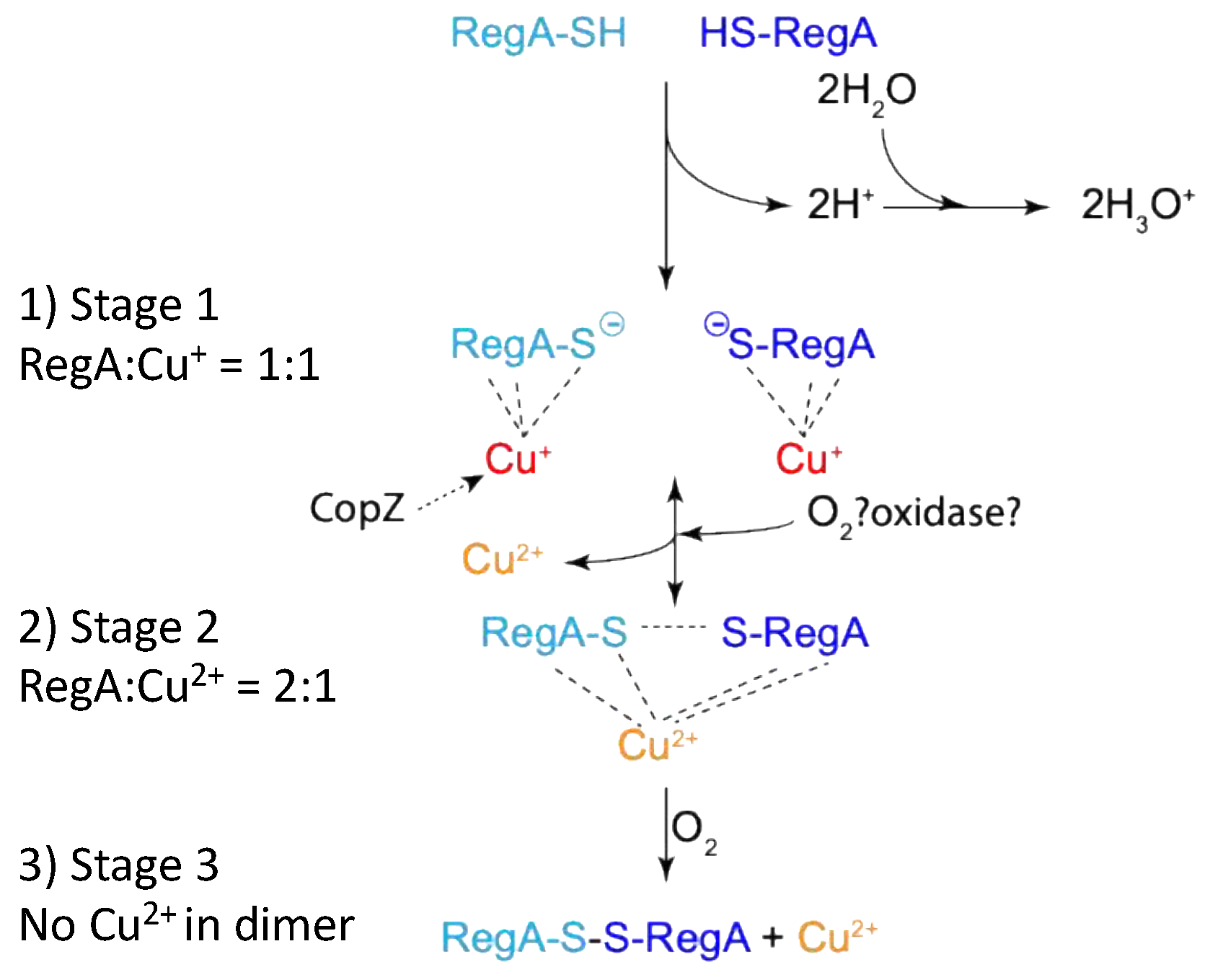
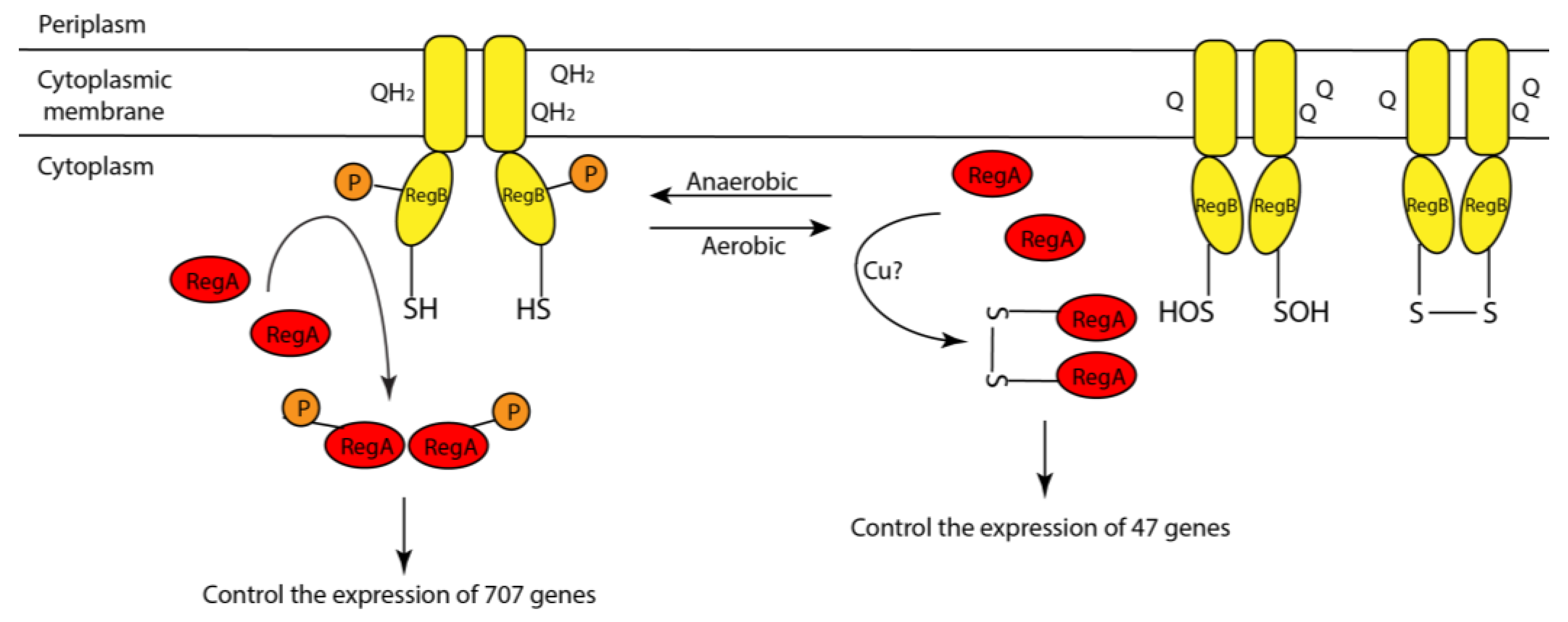
4.1. A Model Describing How Copper and Oxygen Promote RegA Disulfide Bond Formation
4.2. Both Aerobic and Anaerobic Conditions Control RegA Activity
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, R.; Stock, A.M. Biological insights from structures of two-component proteins. Ann. Rev. Microbiol. 2009, 63, 133–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-component signal transduction. Ann. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swem, L.R.; Elsen, S.; Bird, T.H.; Swem, D.L.; Koch, H.G.; Myllykallio, H.; Daldal, F.; Bauer, C.E. The RegB/RegA two-component regulatory system controls synthesis of photosynthesis and respiratory electron transfer components in Rhodobacter capsulatus. J. Mol. Biol. 2001, 309, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Swem, L.R.; Gong, X.; Yu, C.-A.; Bauer, C.E. Identification of a ubiquinone-binding site that affects autophosphorylation of the sensor kinase RegB. J. Biol. Chem. 2006, 281, 6768–6775. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Bauer, C.E. RegB kinase activity is controlled in part by monitoring the ratio of oxidized to reduced ubiquinones in the ubiquinone pool. mBio 2010, 1, e00272-10. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Cheng, Z.; Reddie, K.; Carroll, K.; Hammad, L.A.; Karty, J.A.; Bauer, C.E. RegB kinase activity is repressed by oxidative formation of cysteine sulfenic acid. J. Biol. Chem. 2013, 288, 4755–4762. [Google Scholar] [CrossRef] [Green Version]
- Swem, L.R.; Kraft, B.J.; Swem, D.L.; Setterdahl, A.T.; Masuda, S.; Knaff, D.B.; Zaleski, J.M.; Bauer, C.E. Signal transduction by the global regulator RegB is mediated by a redox-active cysteine. EMBO J. 2003, 22, 4699–4708. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Bauer, C.E. RegB/RegA, a global redox-responding two-component system. Adv. Exp. Med. Biol. 2008, 631, 131–148. [Google Scholar]
- Wu, J.; Dragnea, V.; Bauer, C.E. Redox Responding Sensor Kinases. In Two Component Systems in Bacteria; Gross, R., Beier, D., Eds.; Caister Academic Press: Würzburg, Germany, 2012; pp. 41–56. [Google Scholar]
- Schindel, H.; Bauer, C.E. The RegA regulon exhibits variability in response to altered growth conditions and differs markedly between Rhodobacter species. Microb. Genom. 2016, 2, e000081. [Google Scholar] [CrossRef]
- Masuda, S.; Matsumoto, Y.; Nagashima, K.V.P.; Shimada, K.; Inoue, K.; Bauer, C.E.; Matsuura, K. Structural and functional analyses of photosynthetic regulatory genes regA and regB from Rhodovulum sulfidophilum, Roseobacter denitrificans, and Rhodobacter capsulatus. J. Bacteriol. 1999, 181, 4205–4215. [Google Scholar] [CrossRef] [Green Version]
- Elsen, S.; Swem, L.R.; Swem, D.L.; Bauer, C.E. RegB/RegA, a highly conserved redox-responding global two-component regulatory system. Microbiol. Mol. Biol. Rev. 2004, 68, 263–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swem, D.L.; Swem, L.R.; Setterdahl, A.; Bauer, C.E. Involvement of SenC in assembly of cytochrome c oxidase in Rhodobacter capsulatus. J. Bacteriol. 2005, 187, 8081–8087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohmeyer, E.; Schröder, S.; Pawlik, G.; Trasnea, P.-I.; Peters, A.; Daldal, F.; Koch, H.-G. The ScoI homologue SenC is a copper binding protein that interacts directly with the cbb3-type cytochrome oxidase in Rhodobacter capsulatus. Biochim. Biophys. Acta 2012, 1817, 2005–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trasnea, P.I.; Utz, M.; Khalfaoui-Hassani, B.; Lagies, S.; Daldal, F.; Koch, H.G. Cooperation between two periplasmic copper chaperones is required for full activity of the cbb3 -type cytochrome c oxidase and copper homeostasis in Rhodobacter capsulatus. Mol. Microbiol. 2016, 100, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwan, A.G.; Lewin, A.; Davy, S.L.; Boetzel, R.; Leech, A.; Walker, D.; Wood, T.; Moore, G.R. PrrC from Rhodobacter sphaeroides, a homologue of eukaryotic Sco proteins, is a copper-binding protein and may have a thiol-disulfide oxidoreductase activity. FEBS Lett. 2002, 518, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Laguri, C.; Phillips-Jones, M.K.; Williamson, M.P. Solution structure and DNA binding of the effector domain from the global regulator PrrA (RegA) from Rhodobacter sphaeroides: Insights into DNA binding specificity. Nucleic Acids Res. 2003, 31, 6778–6787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, S.; Bird, T.H.; Bauer, C.E. DNA binding characteristics of RegA. A constitutively active anaerobic activator of photosynthesis gene expression in Rhodobacter capsulatus. J. Biol. Chem. 1998, 273, 18509–18513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willett, J.; Smart, J.L.; Bauer, C.E. RegA Control of bacteriochlorophyll and carotenoid synthesis in Rhodobacter capsulatus. J. Bacteriol. 2007, 189, 7765–7773. [Google Scholar] [CrossRef] [Green Version]
- Bird, T.H.; Du, S.; Bauer, C.E. Autophosphorylation, phosphotransfer, and DNA-binding properties of the RegB/RegA two-component regulatory system in Rhodobacter capsulatus. J. Biol. Chem. 1999, 274, 16343–16348. [Google Scholar] [CrossRef] [Green Version]
- Bauer, C.E.; Elsen, S.; Bird, T.H. Mechanisms for redox control of gene expression. Annu. Rev. Microbiol. 1999, 53, 495–523. [Google Scholar] [CrossRef]
- Kim, S.O.; Merchant, K.; Nudelman, R.; Beyer, W.F., Jr.; Keng, T.; DeAngelo, J.; Hausladen, A.; Stamler, J.S. OxyR: A molecular code for redox-related signaling. Cell 2002, 109, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Masuda, S.; Dong, C.; Swem, D.; Setterdahl, A.T.; Knaff, D.B.; Bauer, C.E. Repression of photosynthesis gene expression by formation of a disulfide bond in CrtJ. Proc. Natl. Acad. Sci. USA 2002, 99, 7078–7083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Wu, J.; Setterdahl, A.; Reddie, K.; Carroll, K.; Hammad, L.A.; Karty, J.A.; Bauer, C.E. Activity of the tetrapyrrole regulator CrtJ is controlled by oxidation of a redox active cysteine located in the DNA binding domain. Mol. Microbiol. 2012, 85, 734–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, M.; Bauer, C.E. The vitamin B12 -dependent photoreceptor AerR relieves photosystem gene repression by extending the interaction of CrtJ with photosystem promoters. mBio 2017, 8, e00261-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiley, P.J.; Beinert, H. Oxygen sensing by the global regulator, FNR: The role of the iron-sulfur cluster. FEMS Microbiol. Rev. 1998, 22, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Peuser, V.; Remes, B.; Klug, G. Role of the Irr protein in the regulation of iron metabolism in Rhodobacter sphaeroides. PLoS ONE 2012, 7, e42231. [Google Scholar] [CrossRef] [Green Version]
- Utz, M.; Andrei, A.; Milanov, M.; Trasnea, P.; Marckmann, D.; Daldal, F.; Koch, H. The Cu chaperone CopZ is required for Cu homeostasis in Rhodobacter capsulatus and influences cytochrome cbb3 oxidase assembly. Mol. Microbiol. 2019, 111, 764–783. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Liu, T.; Ramesh, A.; Ma, Z.; Ward, S.K.; Zhang, L.; George, G.; Talaat, A.M.; Sacchettini, J.C.; Giedroc, D. CsoR is a novel Mycobacterium tuberculosis copper-sensing transcriptional regulator. Nat. Chem. Biol. 2007, 3, 60–68. [Google Scholar] [CrossRef]
- Ekici, S.; Yang, H.; Koch, H.G.; Daldal, F. Novel transporter required for biogenesis of cbb3-type cytochrome c oxidase in Rhodobacter capsulatus. mBio 2012, 3, e00293-11. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Cowart, D.M.; Scott, R.A.; Giedroc, D.P. Molecular Insights into the Metal Selectivity of the Copper(I)-Sensing Repressor CsoR from Bacillus subtilis. Biochemistry 2009, 48, 3325–3334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekici, S.; Turkarslan, S.; Pawlik, G.; Dancis, A.; Baliga, N.S.; Koch, H.-G.; Daldal, F. Intracytoplasmic copper homeostasis controls cytochrome c oxidase production. mBio 2014, 5, e01055-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumaa, P. Copper chaperones. The concept of conformational control in the metabolism of copper. FEBS Lett. 2013, 587, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, A.K.; Smith, D.; Gray, J.; Carr, H.S.; Liu, A.; Winge, D.R.; Hosler, J.P. Mutagenic analysis of Cox11 of Rhodobacter sphaeroides: Insights into the assembly of Cu(B) of cytochrome c oxidase. Biochemistry 2010, 49, 5651–5661. [Google Scholar] [CrossRef] [Green Version]
- Mosley, C.S.; Suzuki, J.Y.; Bauer, C.E. Identification and molecular genetic characterization of a sensor kinase responsible for coordinately regulating light harvesting and reaction center gene expression in response to anaerobiosis. J. Bacteriol. 1994, 176, 7566–7573. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, N.; Bauer, C.E. The Response Regulator RegA Is a Copper Binding Protein That Covalently Dimerizes When Exposed to Oxygen. Microorganisms 2022, 10, 934. https://doi.org/10.3390/microorganisms10050934
Ke N, Bauer CE. The Response Regulator RegA Is a Copper Binding Protein That Covalently Dimerizes When Exposed to Oxygen. Microorganisms. 2022; 10(5):934. https://doi.org/10.3390/microorganisms10050934
Chicago/Turabian StyleKe, Nijia, and Carl E. Bauer. 2022. "The Response Regulator RegA Is a Copper Binding Protein That Covalently Dimerizes When Exposed to Oxygen" Microorganisms 10, no. 5: 934. https://doi.org/10.3390/microorganisms10050934
APA StyleKe, N., & Bauer, C. E. (2022). The Response Regulator RegA Is a Copper Binding Protein That Covalently Dimerizes When Exposed to Oxygen. Microorganisms, 10(5), 934. https://doi.org/10.3390/microorganisms10050934