
The Frequency of Occurrence of Resistance and Genes Involved in the Process of Adhesion and Accumulation of Biofilm in Staphylococcus aureus Strains Isolated from Tracheostomy Tubes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Phenotypic Identification of Staphylococcus aureus
2.3. Antimicrobial Susceptibility Testing
2.4. Genomic DNA Extraction
2.5. Pulsed Field Gel Electrophoresis (PFGE)
2.6. Molecular Confirmation of the Species Staphylococcus aureus
2.7. Methicillin-Resistant Staphylococcus aureus Screening
2.8. Quantification of Biofilm in Microtiter Plates
2.9. Phenotypic Characterization of Slime Producing Ability on Congo Red Agar (CRA)
2.10. MSCRAMM Gene Detection
2.11. icaAD Gene Detection
2.12. Scanning Electron Microscopy (SEM)
3. Results
3.1. Characteristics of Patients
3.2. Bacterial Strains and MRSA Screening
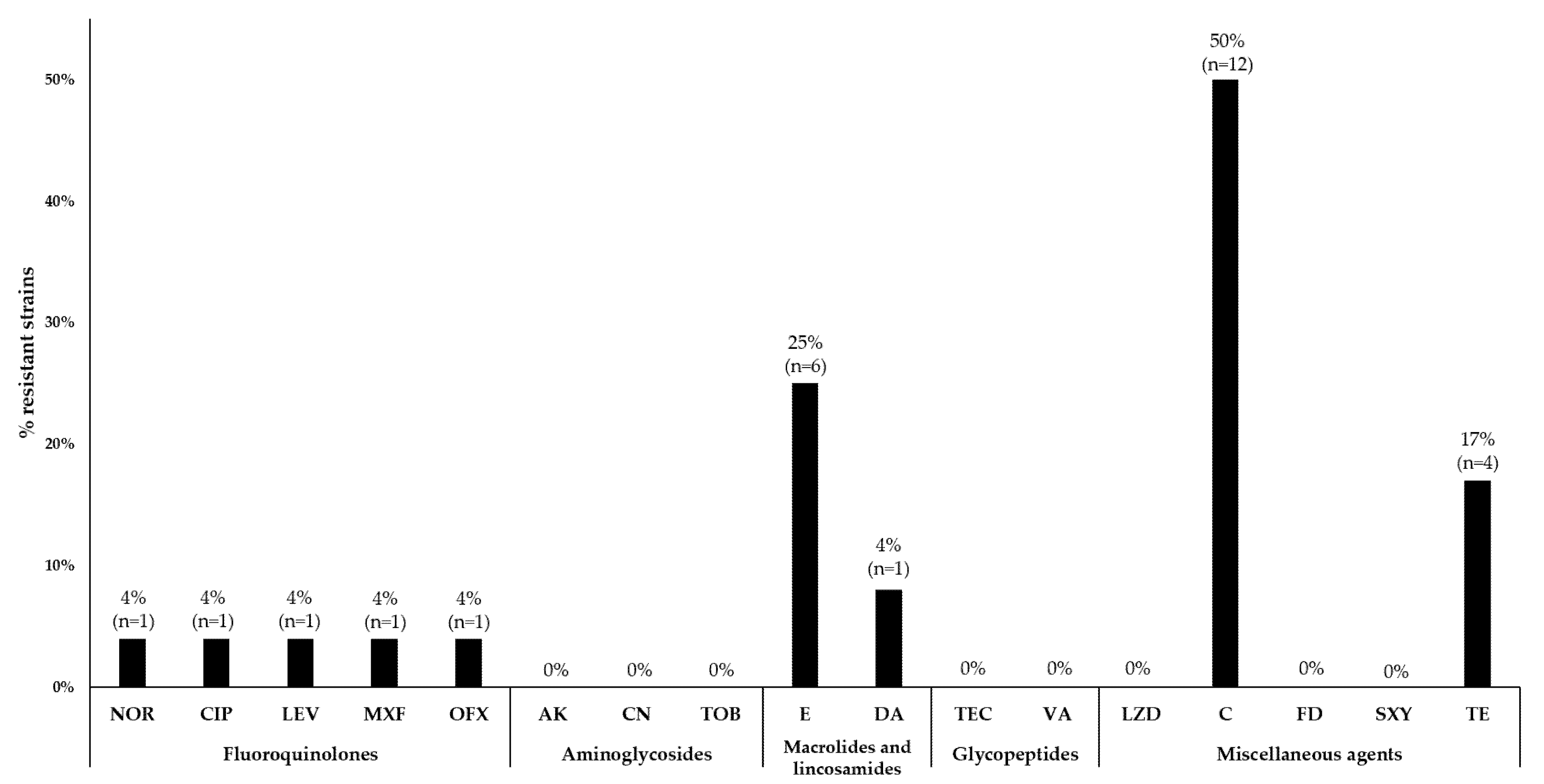
3.3. Antimicrobial Susceptibility Testing
3.4. PFGE
3.5. Quantification of Biofilm in Microtiter Plates
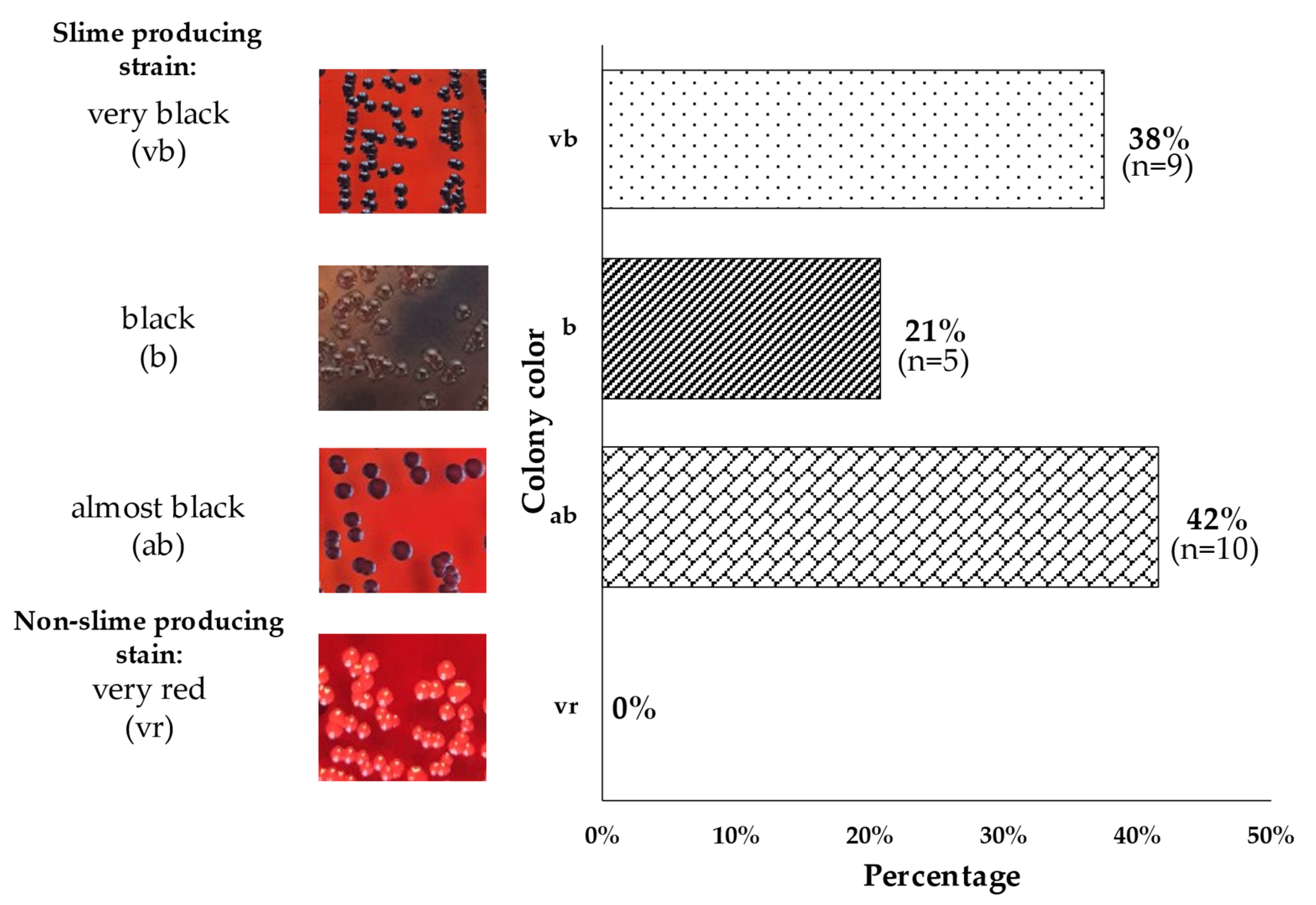
3.6. Phenotypic Characterization of Slime Producing Ability on CRA
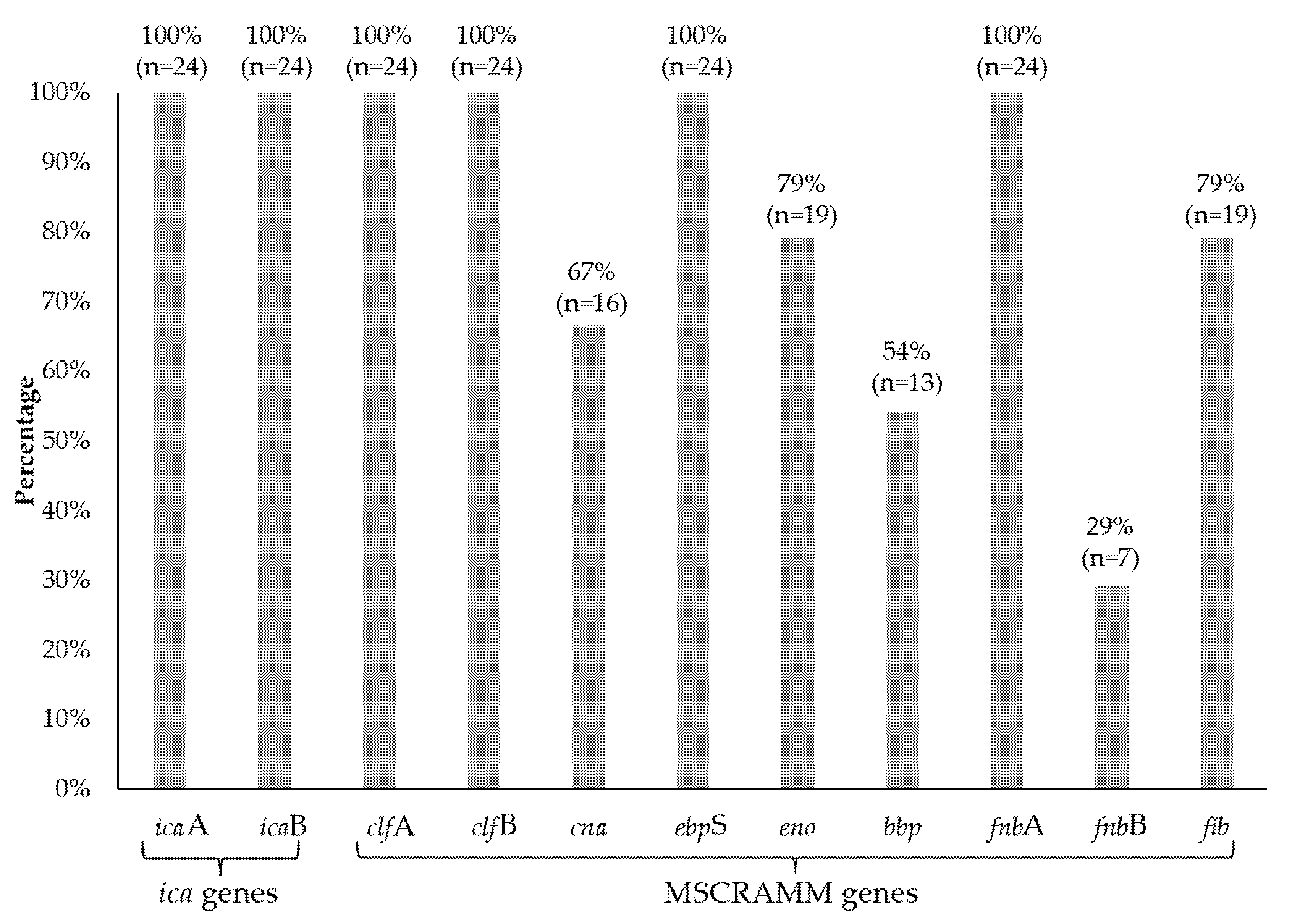
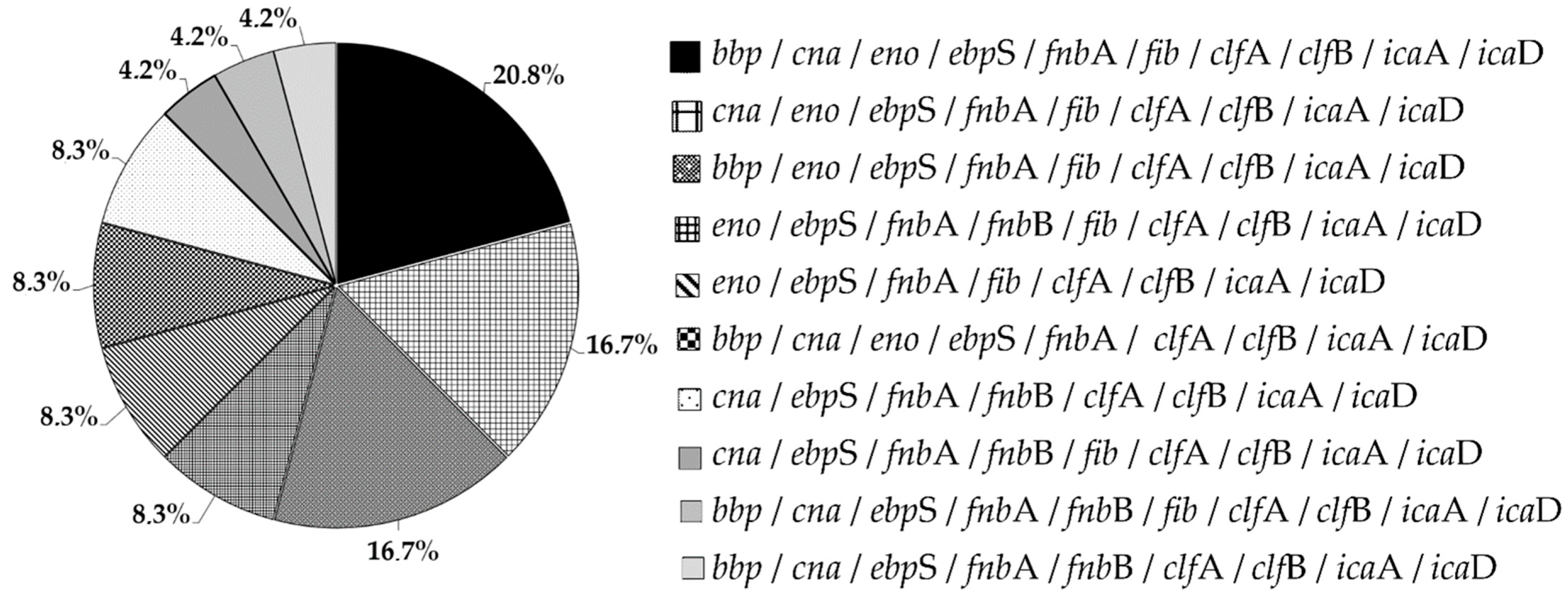
3.7. MSCRAMMs
3.8. icaAD Genes
3.9. Scanning Electron Microscopy (SEM)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laux, C.; Peschel, A.; Krismer, B. Staphylococcus aureus Colonization of the Human Nose and Interaction with Other Microbiome Members. Microbiol. Spectr. 2019, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezende-Pereira, G.; Albuquerque, J.P.; Souza, M.C.; Nogueira, B.A.; Silva, M.G.; Hirata, R.; Mattos-Guaraldi, A.L.; Duarte, R.S.; Neves, F.P.G. Biofilm Formation on Breast Implant Surfaces by Major Gram-Positive Bacterial Pathogens. Aesthetic Surg. J. 2021, 41, 1144–1151. [Google Scholar] [CrossRef]
- Hasan Abdul Cader, S.; Fahim Ahmed Shah, F.A. Tracheostomy colonisation and microbiological isolates of patients in intensive care units-a retrospective study. World J. Otorhinolaryngol. Head Neck Surg. 2020, 6, 49–52. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- El Cheik, M.R.; Barbosa, J.M.; Caixêta, J.A.S.; Avelino, M.A.G. Microbiology of Tracheal Secretions: What to Expect with Children and Adolescents With Tracheostomies. Int. Arch. Otorhinolaryngol. 2018, 22, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Abdollahi, A.; Shoar, S.; Shoar, N. Microorganisms’ colonization and their antibiotic resistance pattern in oro—tracheal tube. Iran. J. Microbiol. 2013, 5, 102–107. [Google Scholar]
- Lepainteur, M.; Ogna, A.; Clair, B.; Dinh, A.; Tarragon, C.; Prigent, H.; Davido, B.; Barbot, F.; Vaugier, I.; Afif, M.; et al. Risk Factors for Respiratory Tract Bacterial Colonization in Adults with Neuromuscular or Neurological Disorders and Chronic Tracheostomy. Respir. Med. 2019, 152, 32–36. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Jensen, P.Ø.; Fiandaca, M.J.; Pedersen, J.; Hansen, C.R.; Andersen, C.B.; Pressler, T.; Givskov, M.; Høiby, N. Pseudomonas aeruginosa biofilms in the respiratory tract of cystic fibrosis patients. Pediatric Pulmonol. 2009, 44, 547–548. [Google Scholar] [CrossRef]
- Piechota, M.; Kot, B.; Frankowska-Maciejewska, A.; Gruzewska, A.; Woźniak-Kosek, A. Biofilm Formation by Methicillin-Resistant and Methicillin-Sensitive Staphylococcus aureus Strains from Hospitalized Patients in Poland. BioMed Res. Int. 2018, 2018, 4657396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4, 529–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folliero, V.; Franci, G.; Dell’Annunziata, F.; Giugliano, R.; Foglia, F.; Sperlongano, R.; De Filippis, A.; Finamore, E. Evaluation of Antibiotic Resistance and Biofilm Production among Clinical Strain Isolated from Medical Devices. Int. J. Microbiol. 2021, 2021, 9033278. [Google Scholar] [CrossRef] [PubMed]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.; Penadés, J.R. Bap, a Staphylococcus aureus Surface Protein Involved in Biofilm Formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. Presence of icaA and icaDGenes and Slime Production in a Collection of Staphylococcal Strains from Catheter-Associated Infections. J. Clin. Microbiol. 2001, 39, 2151–2156. [Google Scholar] [CrossRef] [Green Version]
- Pereiraab, E.M.; Schuencka, E.P.; Malvara, K.L.; Iorioa, N.L.P.; Matosa, P.D.M.; Olendzkic, A.N.; Oelemanna, W.M.R.; dos Santosa, K.R.N. Staphylococcus aureus, Staphylococcus epidermidis and Staphylococcus haemolyticus: Methicillin-resistant isolates are detected directly in blood cultures by multiplex PCR. Microbiol. Res. 2010, 165, 243–249. [Google Scholar] [CrossRef]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing Breakpoint tables for Interpretation of MICs and Zone Diameters Version 11.0, Valid from 2021-01-01. Available online: http://www.eucast.org (accessed on 1 January 2021).
- Murchan, S.; Kaufmann, M.E.; Deplano, A.; de Ryck, R.; Struelens, M.; Zinn, C.E.; Fussing, V.; Salmenlinna, S.; Vuopio-Varkila, J.; El Solh, N.; et al. Harmonization of Pulsed-Field Gel Electrophoresis Protocols for Epidemiological Typing of Strains of Methicillin-Resistant Staphylococcus aureus: A single approach developed by consensus in 10 European laboratories and its application for tracing the spread of related strains. J. Clin. Microbiol. 2003, 41, 1574–1585. [Google Scholar]
- Ochońska, D.; Ścibik, Ł.; Brzychczy-Włoch, M. Biofilm Formation of Clinical Klebsiella pneumoniae Strains Isolated from Tracheostomy Tubes and Their Association with Antimicrobial Resistance, Virulence and Genetic Diversity. Pathogens 2021, 10, 1345. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Acta Pathol. Microbiol. Scand. 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arciola, C.R.; Campoccia, D.; Gamberini, S.; Cervellati, M.; Donati, E.; Montanaro, L. Detection of slime production by means of an optimised Congo Red agar plate test based on a colourimetric scale in Staphylococcus epidermidis clinical isolates genotyped for ica locus. Biomaterials 2002, 23, 4233–4239. [Google Scholar] [CrossRef]
- Tristan, A.; Ying, L.; Bes, M.; Etienne, J.; Vandenesch, F.; Lina, G. Use of multiplex PCR to identify Staphylococcus aureus adhesins involved in human hematogenous infections. J. Clin. Microbiol. 2003, 41, 4465–4467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajerski, W.; Duch, J.; Ochonska, D.; Gołda-Cępa, M.; Brzychczy-Włoch, M.; Kotarba, A. Bacterial attachment to oxygen-functionalized graphenic surfaces. Mater. Sci. Eng. C 2020, 113, 110972. [Google Scholar] [CrossRef] [PubMed]
- Ghasemian, A.; Peerayeh, S.N.; Bakhshi, B.; Mirzaee, M. The Microbial Surface Components Recognizing Adhesive Matrix Molecules (MSCRAMMs) Genes among Clinical Isolates of Staphylococcus aureus from Hospitalized Children. Iran. J. Pathol. 2015, 10, 258–264. [Google Scholar]
- Foster, T.J. The MSCRAMM Family of Cell-Wall-Anchored Surface Proteins of Gram-Positive Cocci. Trends Microbiol. 2019, 27, 927–941. [Google Scholar] [CrossRef]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Golding, G.R.; Campbell, J.; Spreitzer, D.; Chui, L. Pulsed-field gel electrophoresis of Staphylococcus aureus. Methods Mol. Biol. 2015, 1301, 85–93. [Google Scholar]
- Jain, S.; Chowdhury, R.; Datta, M.; Chowdhury, G.; Mukhopadhyay, A.K. Characterization of the clonal profile of methicillin resistant Staphylococcus aureus isolated from patients with early post-operative orthopedic implant based infections. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 8. [Google Scholar] [CrossRef]
- Zariza, S.; Yeo, C.C.; Faizal, G.M.; Chew, C.H.; Zakaria, Z.A.; Al-Obaidi, M.M.J.; Amin, N.S.; Nasir, M.D.M. Nasal colonisation, antimicrobial susceptibility and genotypic pattern of Staphylococcus aureus among agricultural biotechnology students in Besut, Terengganu, east coast of Malaysia. Trop. Med. Int. Health 2018, 23, 905–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, B.R.; Skurray, R.A. Antimicrobial resistance of Staphylococcus aureus: Genetic basis. Microbiol. Rev. 1987, 51, 88–134. [Google Scholar] [CrossRef] [PubMed]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.J.; Fatma, T.; Rattan, A. Detection of biofilm formation among the clinical isolates of Staphylococi: An evaluation of three different screening methods. Indian J. Med. Microbiol. 2006, 24, 25–29. [Google Scholar] [CrossRef]
- Nasr, R.A.; AbuShady, H.M.; Hussein, H.S. Biofilm formation and presence of icaAD gene in clinical isolates of staphylococci. Egypt. J. Med. Hum. Genet. 2012, 13, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Wertheim, H.F.L.; Walsh, E.; Choudhurry, R.; Melles, D.C.; Boelens, H.A.M.; Miajlovic, H.; Verbrugh, H.A.; Foster, T.; van Belkum, A. Key role for clumping factor B in Staphylococcus aureus nasal colonization of humans. PLoS Med. 2008, 5, e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galant, K.; Giedrys-Kalemba, S.; Johaniuk, A.; Roszkowska, P.; Jursa-Kulesza, J. Occurence of factors associated with ability to biofilm formation among methicillin-sensitive and methicillin-resistant Staphylococcus aureus strains isolated from patients with bone infections. Forum Zakażeń 2016, 7, 385–388. [Google Scholar] [CrossRef]
- Rhem, M.N.; Lech, E.M.; Patti, J.N.; McDevitt, D.; Höök, M.; Jones, D.B.; Wilhelmus, K.R. The collagen-binding adhesin is a virulence factor in Staphylococcus aureus keratitis. Infect. Immun. 2000, 68, 3776–3779. [Google Scholar] [CrossRef] [Green Version]
- Josefsson, E.; Hartford, O.; O’Brien, L.; Patti, J.M.; Foster, T. Protection against experimental Staphylococcus aureus arthritis by vaccination with clumping factor A, a novel virulence determinant. J. Infect. Dis. 2001, 184, 1572–1580. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, C.R.W.; Postol, E.; Nomizo, R.; Reis, L.F.L.; Brentani, R.R. Identification of enolase as a laminin-binding protein on the Surface of Staphylococcus aureus. Microbes Infect. 2004, 6, 604–608. [Google Scholar] [CrossRef]
- Downer, R.; Roche, F.; Park, P.W.; Mecham, R.P.; Foster, T.J. The Elastin-binding Protein of Staphylococcus aureus (EbpS) Is Expressed at the Cell Surface as an Integral Membrane Protein and Not as a Cell Wall-associated Protein. J. Biol. Chem. 2002, 277, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Nourbakhsh, F.; Namvar, A.E. Detection of genes involved in biofilm formation in Staphylococcus aureus isolates. GMS Hyg. Infect. Control 2016, 11, Doc07. [Google Scholar] [PubMed]
- Grinholc, M.; Wegrzyn, G.; Kurlenda, J. Evaluation of biofilm production and prevalence of the icaD gene in methicillin-resistant and methicillin-susceptible Staphylococcus aureus strains isolated from patients with nosocomial infections and carriers. FEMS Immunol. Med. Microbiol. 2007, 50, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Gotz, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, V.G.; Fey, P.D.; Reller, L.B.; Chamis, A.L.; Corey, G.R.; Rupp, M.E. The intercellular adhesin locus ica is present in clinical isolates of Staphylococcus aureus from bacteremic patients with infected and uninfected prosthetic joints. Med. Microbiol. Immunol. 2001, 189, 127–131. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species/Gene | Sequence (5′ → 3′) | Product Size (pz) | Source |
|---|---|---|---|---|
| Molecular confirmation of the species Staphylococcus aureus and detection of mecA gene | ||||
| 1 | S. aureus | AATCTTTGTCGGTACACGATATTCTTCACG | 108 | [18] |
| CGTAATGAGATTTCAGTAGATAATACAACA | ||||
| 2 | S. epidermidis | ATCAAAAAGTTGGCGAACCTTTTCA | 124 | |
| CAAAAGAGCGTGGAGAAAAGTATCA | ||||
| 3 | S. haemolyticus | GGTCGCTTAGTCGGAACAAT | 271 | |
| CACGAGCAATCTCATCACCT | ||||
| 4 | mecA | TAGAAATGACTGAACGTCCG | 154 | |
| TTGCGATCAATGTTACCGTAG | ||||
| Operon ica | ||||
| 5 | icaA | ACACTTGCTGGCGCAGTCAA | 188 | [11] |
| TCTGGAACCAACATCCAACA | ||||
| 6 | icaD | ATGGTCAAGCCCAGACAGAG | 198 | |
| AGTATTTTCAATGTTTAAAGCAA | ||||
| MSCRAMM | ||||
| 7 | bbp | AACTACATCTAGTACTCAACAACAG | 575 | [25] |
| ATGTGCTTGAATAACACCATCATCT | ||||
| 8 | cna | GTCAAGCAGTTATTAACACCAGAC | 423 | |
| AATCAGTAATTGCACTTTGTCCACTG | ||||
| 9 | eno | ACGTGCAGCAGCTGACT | 302 | |
| CAACAGCATYCTTCAGTACCTTC | ||||
| 10 | ebpS | CATCCAGAACCAATCGAAGAC | 186 | |
| CTTAACAGTTACATCATCATGTTTATCTTTG | ||||
| 11 | fnbA | GTGAAGTTTTAGAAGGTGGAAAGATTAG | 643 | |
| GCTCTTGTAAGACCATTTTTCTTCAC | ||||
| 12 | fnbB | GTAACAGCTAATGGTCGAATTGATACT | 524 | |
| CAAGTTCGATAGGAGTACTATGTTC | ||||
| 13 | fib | CTACAACTACAATTGCCGTCAACAG | 404 | |
| GCTCTTGTAAGACCATTTTCTTCAC | ||||
| 14 | clfA | ATTGGCGTGGCTTCAGTGCT | 292 | |
| CGTTTCTTCCGTAGTTGCATTTG | ||||
| 15 | clfB | ACATCAGTAATAGTAGGGGGCAAC | 205 | |
| TTCGCACTGTTTGTGTTTGCAC | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drożdż, K.; Ochońska, D.; Ścibik, Ł.; Gołda-Cępa, M.; Biegun, K.; Brzychczy-Włoch, M. The Frequency of Occurrence of Resistance and Genes Involved in the Process of Adhesion and Accumulation of Biofilm in Staphylococcus aureus Strains Isolated from Tracheostomy Tubes. Microorganisms 2022, 10, 1210. https://doi.org/10.3390/microorganisms10061210
Drożdż K, Ochońska D, Ścibik Ł, Gołda-Cępa M, Biegun K, Brzychczy-Włoch M. The Frequency of Occurrence of Resistance and Genes Involved in the Process of Adhesion and Accumulation of Biofilm in Staphylococcus aureus Strains Isolated from Tracheostomy Tubes. Microorganisms. 2022; 10(6):1210. https://doi.org/10.3390/microorganisms10061210
Chicago/Turabian StyleDrożdż, Kamil, Dorota Ochońska, Łukasz Ścibik, Monika Gołda-Cępa, Katarzyna Biegun, and Monika Brzychczy-Włoch. 2022. "The Frequency of Occurrence of Resistance and Genes Involved in the Process of Adhesion and Accumulation of Biofilm in Staphylococcus aureus Strains Isolated from Tracheostomy Tubes" Microorganisms 10, no. 6: 1210. https://doi.org/10.3390/microorganisms10061210
APA StyleDrożdż, K., Ochońska, D., Ścibik, Ł., Gołda-Cępa, M., Biegun, K., & Brzychczy-Włoch, M. (2022). The Frequency of Occurrence of Resistance and Genes Involved in the Process of Adhesion and Accumulation of Biofilm in Staphylococcus aureus Strains Isolated from Tracheostomy Tubes. Microorganisms, 10(6), 1210. https://doi.org/10.3390/microorganisms10061210