
Bacteria Halotolerant from Karst Sinkholes as a Source of Biosurfactants and Bioemulsifiers

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Isolation of Bacteria
2.3. Selection of Strains with Biosurfactant (BS) and Bioemulsifying (BE) Activity
2.3.1. Emulsifying Activity (EI24)
2.3.2. Biosurfactant Activity (Drop-Collapse)
2.3.3. Stability of BS and BE Activity under Salinity Conditions
2.3.4. Halotolerance Test
2.3.5. Haemolytic Activity
2.4. Motility Test: Swimming and Swarming
2.5. Molecular Identification of Bacteria with BS and BE Activity Stable in Salt
2.6. BS and BE Production at Different Times (h) in Planktonic Culture
2.7. BS and BE Production at Different Times (h) in Sessile Culture
2.8. Optimization of the Composition of the Culture Medium for BS and BE Activity
2.9. Optimization of Sessile and Planktonic Culture Conditions for BS and BE Activity
2.10. Statistical Analysis
3. Results and Discussion
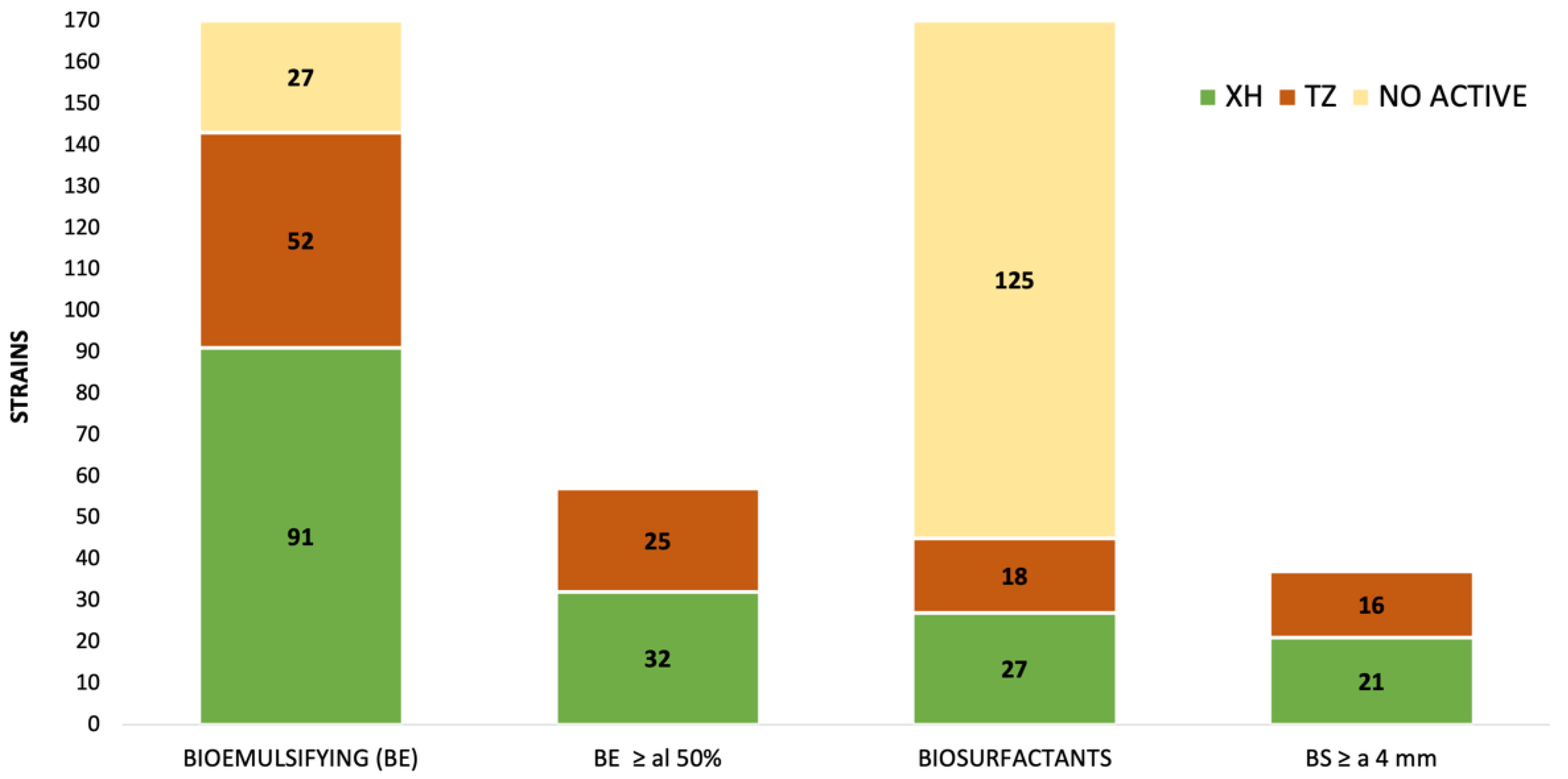
3.1. Isolation of Cultivable Bacteria from Sinkhole
3.2. Biosurfactant and Bioemulsifying Activity of Strains Isolated from Sinkholes
3.3. Stability of the BS and BE Activity under Salinity Conditions
3.4. Haemolytic Activity of Strains with Stable BS and BE Compound under Salinity Conditions
3.5. Motility Test: Swimming and Swarming
3.6. Molecular Identification of Strains with Stable BS and BE Activity in Salinity Conditions
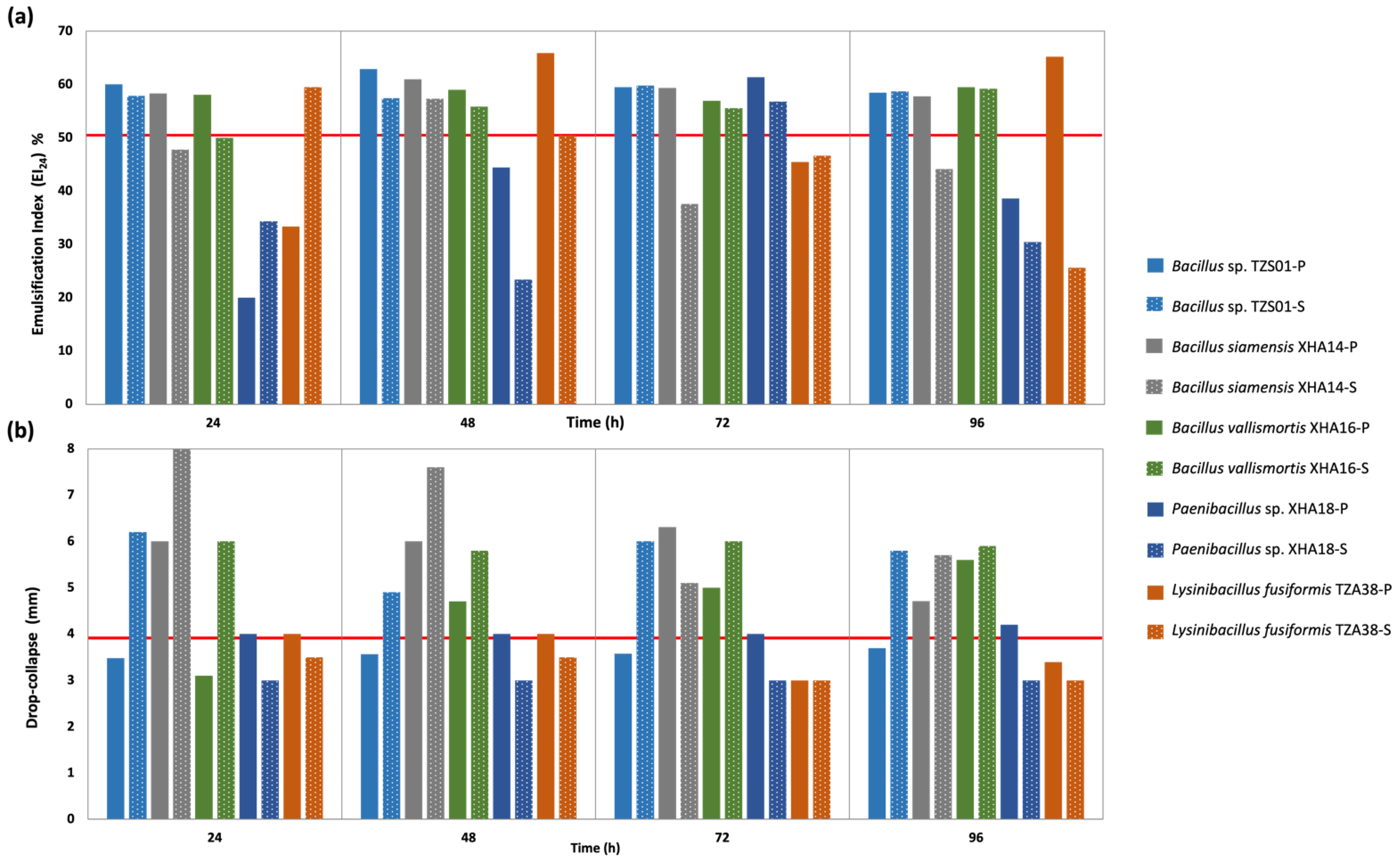
3.7. Expression at Different Times of Metabolites with BS and BE Activity in Planktonic and Sessile Cultures
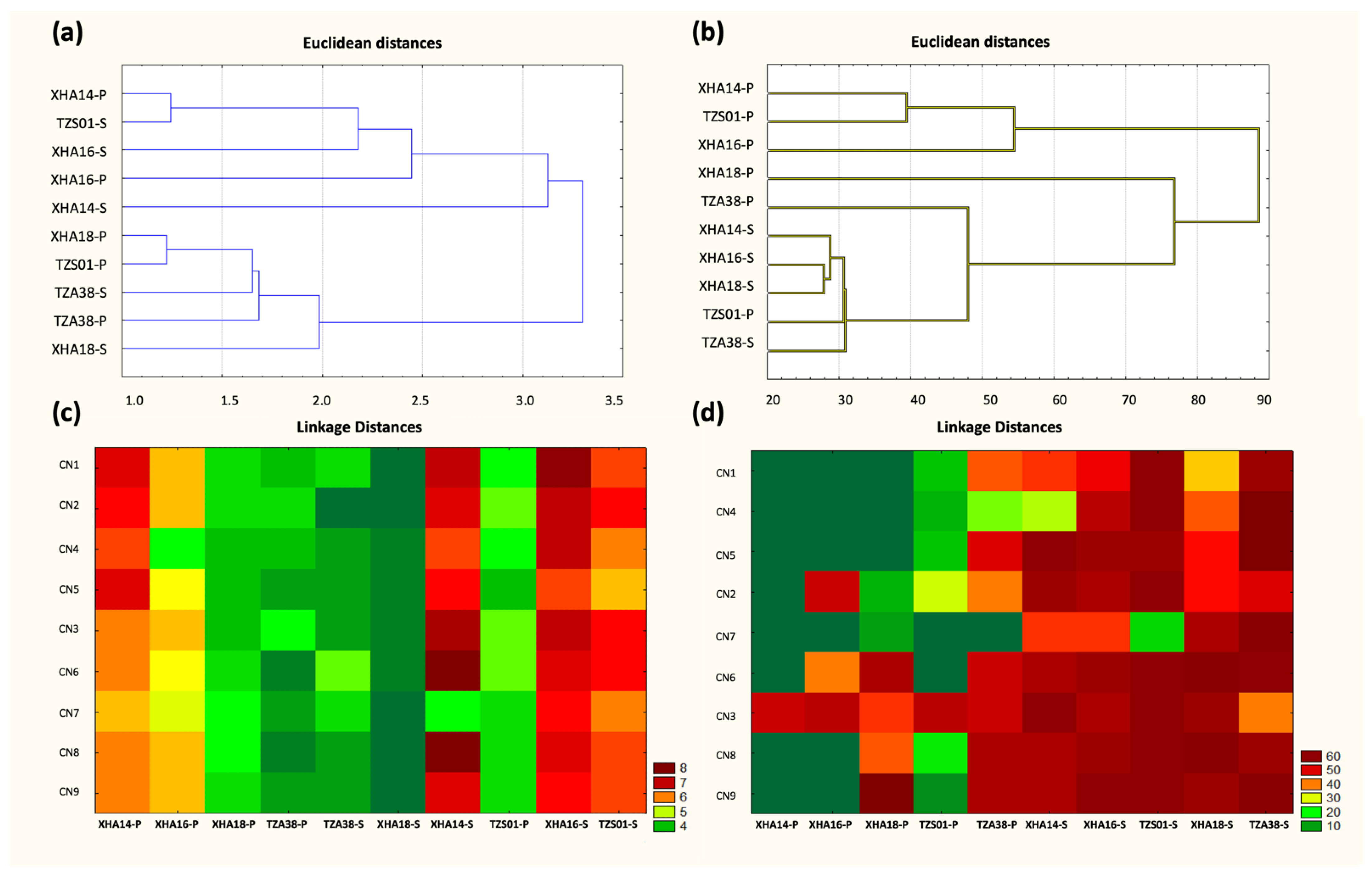
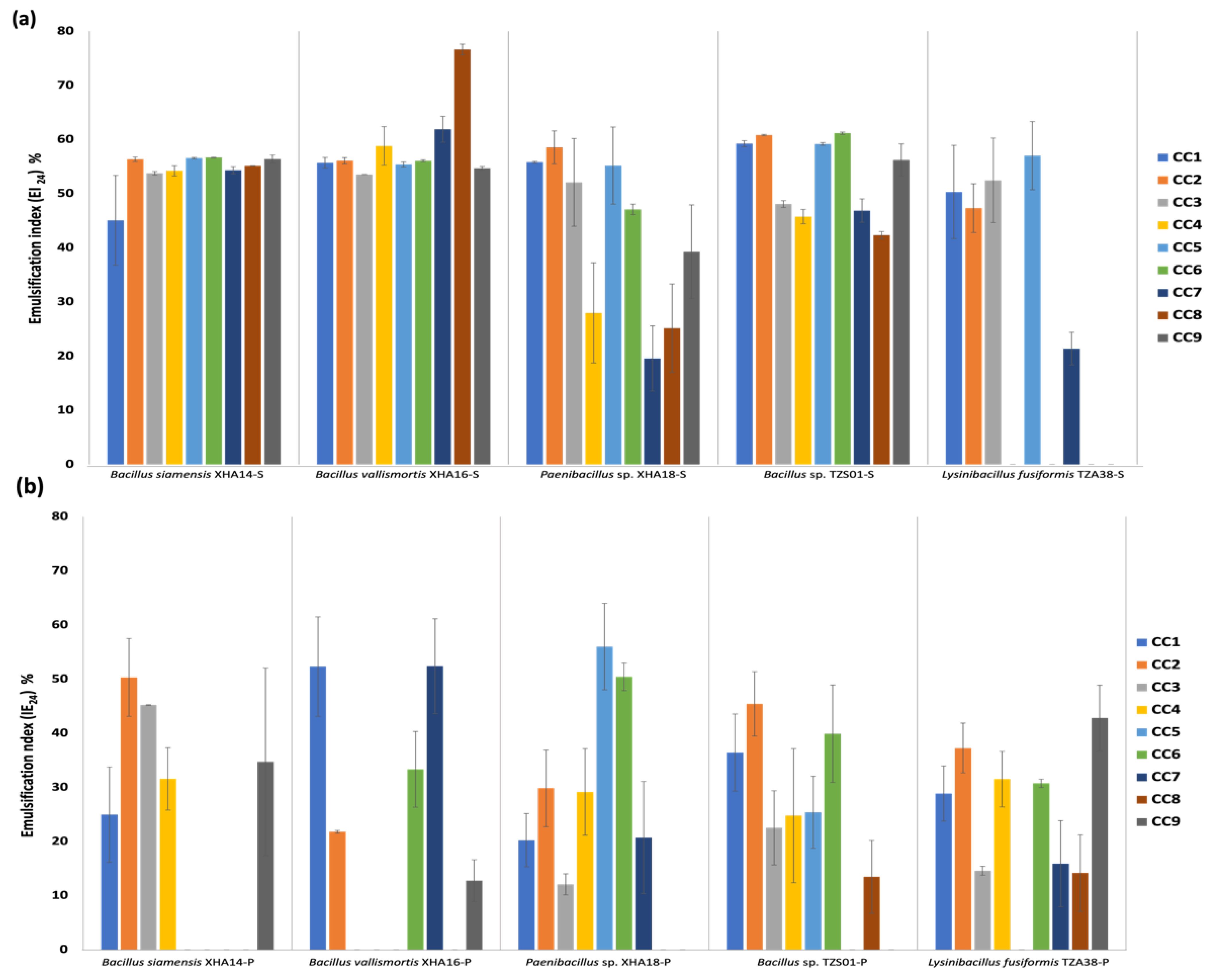
3.8. Optimization of the C/N Source in the Medium to Produce BS and BE Compounds in Sessile and Planktonic Cultures
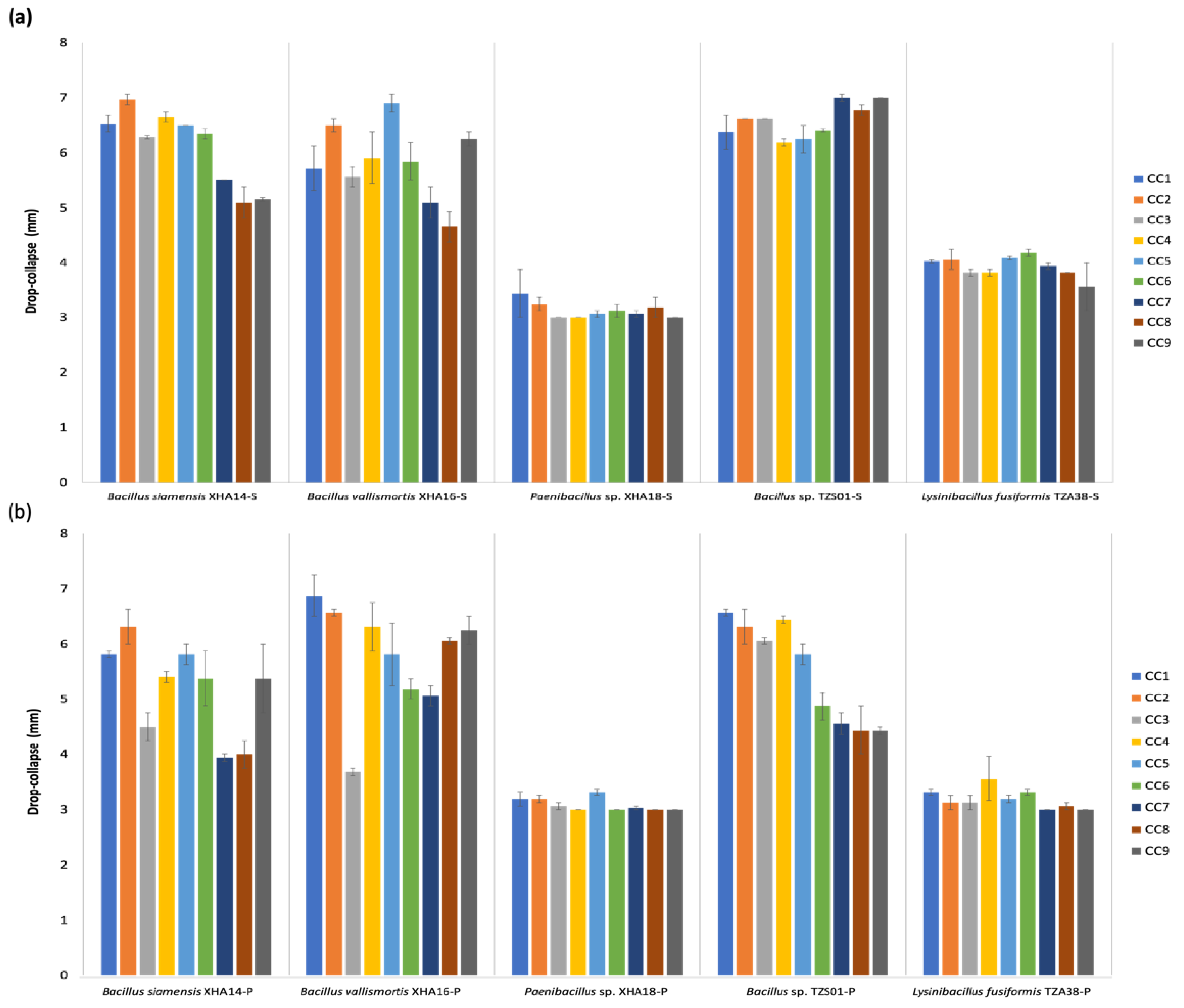
3.9. Optimization of Culture Conditions for the Production of BS and BE Compounds in Sessile and Planktonic Cultures
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perry, E.C.; Velazquez-Oliman, G.; Leal-Bautista, R.M.; Dunning, N. The Icaiche Formation: Major contributor to the stratigraphy, hydrogeochemistry and geomorphology of the northern Yucatán peninsula, Mexico. Bol. Soc. Geol. Mex. 2019, 71, 741–760. [Google Scholar] [CrossRef]
- Bauer-Gottwein, P.; Gondwe, B.R.; Charvet, G.; Marín, L.E.; Rebolledo-Vieyra, M.; Merediz-Alonso, G. The Yucatán Peninsula karst aquifer, Mexico. Hydrogeol. J. 2011, 19, 507–524. [Google Scholar] [CrossRef]
- Schmitter-Soto, J.J.; Comín, F.A.; Escobar-Briones, E.; Herrera-Silveira, J.; Alcocer, J.; Suárez-Morales, E.; Elías-Gutiérrez, M.; Arce-Díaz, V.; Marín, L.E.; Steinich, B. Hydrogeochemical and bilogical characteristics of sinkholes in the Yucatán Peninsula (SE México). Hydrobiologia 2002, 467, 215–228. [Google Scholar] [CrossRef]
- Moreno-Pérez, P.A.; Hernández-Téllez, M.; Ramirez-Durán, N.; Gamboa-Angulo, M.; Isaac-Olivé, K. Microorganisms and spatial distribution of the sinkholes of the Yucatan Peninsula, underestimated biotechnological potential? Water Environ. J. 2019, 34, 41–49. [Google Scholar] [CrossRef]
- Suárez-Moo, P.; Remes-Rodríguez, C.A.; Márquez-Velázquez, N.A.; Falcón, L.I.; García-Maldonado, J.Q.; Prieto-Davó, A. Changes in the sediment microbial community structure of coastal and inland sinkholes of a karst ecosystem from the Yucatan peninsula. Sci. Rep. 2022, 12, 1110. [Google Scholar] [CrossRef]
- De la Rosa-García, S.C.; Muñoz-García, A.A.; Barahona-Pérez, L.F.; Gamboa-Angulo, M.M. Antimicrobial properties of moderately halotolerant bacteria from sinkholes of the Yucatan peninsula. Lett. Appl. Microbiol. 2007, 45, 289–294. [Google Scholar] [CrossRef]
- Paria, S. Surfactant-enhanced remediation of organic contaminated soil and water. Adv. Colloid Interface Sci. 2008, 138, 24–58. [Google Scholar] [CrossRef]
- Mnif, I.; Sahnoun, R.; Ellouz-Chaabouni, S.; Ghribi, D. Application of bacterial biosurfactants for enhanced removal and biodegradation of diesel oil in soil using a newly isolated consortium. Process Saf. Environ. Prot. 2017, 109, 72–81. [Google Scholar] [CrossRef]
- Smyth, T.J.P.; Perfumo, A.; McClean, S.; Marchant, R.; Banat, I.M. Isolation an analysis of lipopeptides and high molecular weight biosurfactants. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3688–3704. [Google Scholar]
- Bach, H.; Gutnick, D.L. Potential applications of bioemulsifiers in the oil industry. In Studies in Surface Science and Catalysis; Elsevier: Amsterdam, The Netherlands, 2004; Volume 151, pp. 233–281. [Google Scholar]
- Banat, I.M.; Franzetti, A.; Gandolfi, I.; Bestetti, G.; Martinotti, M.G.; Fracchia, L.; Smyth, T.J.; Marchant, R. Microbial biosurfactants production, applications and future potential. Appl. Microbiol. Biotechnol. 2010, 87, 427–444. [Google Scholar] [CrossRef]
- Suryawanshi, Y.; Kaur, G.; Mandavkar, A.; Jena, B. Biosurfactants from soil microorganisms as a possible detergent replacement. Int. J. Res. Appl. Sci. Biotechnol. 2021, 8, 136–143. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Singh, A.; Ward, O.P. Physiological aspects. Part 1 in a series of papers devoted to surfactants in microbiology and biotechnology. Biotechnol. Adv. 2006, 24, 604–620. [Google Scholar] [CrossRef] [PubMed]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Piotrowska-Seget, Z.; Cameotra, S.S. Environmental applications of biosurfactants: Recent advances. Int. J. Mol. Sci. 2011, 12, 633–654. [Google Scholar] [CrossRef] [PubMed]
- Banat, I.M.; Satpute, S.K.; Cameotra, S.S.; Patil, R.; Nyayanit, N.V. Cost effective technologies and renewable substrates for biosurfactants’ production. Front. Microbiol. 2014, 5, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panjiar, N.; Sachan, S.G.; Sachan, A. Screening of bioemulsifier-producing micro-organisms isolated from oil-contaminated sites. Ann. Microbiol. 2015, 65, 753–764. [Google Scholar] [CrossRef]
- Shavandi, M.; Mohebalia, G.; Haddadib, A.; Shakaramia, H.; Nuhic, A. Emulsification potential of a newly isolated biosurfactant-producing bacterium, Rhodococcus sp. strain TA6. Colloids Surf. B 2011, 82, 477–482. [Google Scholar] [CrossRef]
- Bodour, A.; Drees, K.; Maier, R. Distribution of biosurfactant-producing bacteria in undisturbed and contaminated arid southwestern soils. Appl. Environ. Microbiol. 2003, 69, 3280–3287. [Google Scholar]
- Oren, A. Microbial life at high salt concentrations: Phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Kirsop, B.E.; Doyle, A. Maintenance of micro-organisms and cultured cells. In A Manual of Laboratory Methods, 2nd ed.; Academic: London, UK, 1991. [Google Scholar]
- Cooper, D.G.; Goldenberg, B.G. Surface-active agents from two Bacillus species. Appl. Environ. Microbiol. 1987, 53, 224–229. [Google Scholar]
- Maneerat, S.; Phetrong, K. Isolation of biosurfactant-producing marine bacteria and characteristics of selected biosurfactant. Songklanakarin J. Sci. Technol. 2007, 29, 781–791. [Google Scholar]
- Cao, Y.; Zhang, B.; Zhu, Z.; Song, X.; Cai, Q.; Chen, B.; Ye, X. Microbial eco-physiological strategies for salinity-mediated crude oil biodegradation. Sci. Total Environ. 2020, 727, 138723. [Google Scholar] [CrossRef]
- Youssef, N.H.; Duncan, K.E.; Nagle, D.P.; Savage, K.N.; Knapp, R.M.; McInerney, M.J. Comparison of methods to detect biosurfactant production by diverse microorganisms. J. Microbiol. Methods. 2004, 56, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Bindel Connelly, M.; Young, G.M.; Sloma, A. Extracellular proteolytic activity plays a central role in swarming motility in Bacillus subtilis. J. Bacteriol. 2004, 186, 4159–4167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, A.; Brader, G.; Corretto, E.; Aleti, G.; Abaidullah, M.; Sessitsch, A.; Hafeez, F.Y. Qualitative analysis of biosurfactants from Bacillus species exhibiting antifungal activity. PLoS ONE 2018, 13, e0198107. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Boyd, K.G.; Adams, D.R.; Burgess, J.G. Biofilm-specific cross-species induction of antimicrobial compounds in bacilli. Appl. Environ. Microbiol. 2003, 69, 3719–3727. [Google Scholar] [CrossRef] [Green Version]
- Jumpathong, W.; Intra, B.; Euanorasetr, J.; Wanapaisan, P. Biosurfactant-Producing Bacillus velezensis PW192 as an Anti-Fungal Biocontrol Agent against Colletotrichum gloeosporioides and Colletotrichum musae. Microorganisms 2022, 10, 1017. [Google Scholar] [CrossRef]
- Saimmai, A.; Sobhon, V.; Maneerat, S. Molasses as a whole medium for biosurfactants production by Bacillus strains and their application. Biotechnol. Appl. Biochem 2011, 165, 315–335. [Google Scholar] [CrossRef]
- Hippolyte, M.T.; Augustin, M.; Hervé, T.M.; Robert, N.; Devappa, S. Application of response surface methodology to improve the production of antimicrobial biosurfactants by Lactobacillus paracasei subsp. tolerans N2 using sugar cane molasses as substrate. Bioresour. Bioprocess. 2018, 5, 48. [Google Scholar]
- Torsvik, V.L.; Øvreås, L.; Thingstad, T.F. Prokaryotic diversity—Magnitude, dynamics, and controlling factors. Science 2002, 296, 1064–1066. [Google Scholar] [CrossRef] [Green Version]
- De la Rosa-García, S. Evaluación del Potencial Antimicrobiano de Microorganismos Aislados de Cenotes de la Península de Yucatán. Ph.D. Thesis, Tesis de Doctorado en Ciencias y Biotecnología en Plantas. CICY, Mérida, Mexico, 2007; p. 184. (In Spanish). [Google Scholar]
- Joy, S.; Rahman, P.; Sharma, S. Biosurfactant production and concomitant hydrocarbon degradation potentials of bacteria isolated from extreme and hydrocarbon contaminated environments. Chem. Eng. J. 2017, 317, 232–241. [Google Scholar] [CrossRef]
- Varjani, S.; Upasani, V. Critical review on biosurfactant analysis, purification and characterization using rhamnolipid as a model biosurfactant. Bioresour. Technol. 2017, 232, 389–397. [Google Scholar] [CrossRef]
- Nanjani, S.G.; Soni, H. Diversity and EPS Production Potential of Halotolerant Bacteria from Veraval and Dwarka. IOSR J. Pharm. Biol. Sci. 2012, 2, 20–25. [Google Scholar] [CrossRef]
- Agwu, O.; Oluwagunke, T. Halotolerance of Heterotrophic Bacteria Isolated from Tropical Coastal Waters. Int. J. Sci. Basic Appl. 2014, 16, 224–231. [Google Scholar]
- Lopez-Adrian, S.; Herrera-Silviera, J.A. Plankton composition in a sinkholes, Yucatan, Mexico. Verh. Internat. Verein. Limnol. 1994, 25, 1402–1405. [Google Scholar]
- Fenical, W. Chemical Studies of Marine Bacteria: Developing a New Resource. Chem. Rev. 1993, 93, 1673–1683. [Google Scholar] [CrossRef]
- Shivanand, P.; Mugeraya, G. Halophilic bacteria and their compatible solutes—Osmoregulation and potential applications. Curr. Sci. 2011, 100, 1516–1521. [Google Scholar]
- Vaidya, S.; Dev, K.; Sourirajan, A. Distinct Osmoadaptation Strategies in the Strict Halophilic and Halotolerant Bacteria Isolated from Lunsu Salt Water Body of North West Himalayas. Curr. Microbiol. 2018, 75, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Hrenovic, J.; Ivankovic, T. Survival of Escherichia coli and Acinetobacter junii at various concentrations of sodium chloride. EurAsian J. BioSci. 2009, 3, 144–151. [Google Scholar] [CrossRef]
- Kargi, F.; Dinçer, A.R. Use of halophilic bacteria in biological treatment of saline wastewater by fed-batch operation. Water Environ. Res. 2000, 72, 170–174. [Google Scholar] [CrossRef]
- Mainka, T.; Weirathmüller, D.; Herwig, C.; Pflügl, S. Potential applications of halophilic microorganisms for biological treatment of industrial process brines contaminated with aromatics. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab015. [Google Scholar] [CrossRef]
- Vinardell, M.P.; Infante, M.R. The relationship between the chain length of non-ionic surfactants and their hemolytic action on human erythrocytes. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1999, 124, 117–120. [Google Scholar] [CrossRef]
- Ogunshe, A.A.O.; Falode, O.A. The Intriguing Extrapolations of Haemolysis Assay as Screening Criterion for Selecting Biosurfactant-Producing Microorganisms in Petroleum Industries Process-Conditions. J. Pet. Environ. Biotechnol. 2021, 8, 431. [Google Scholar]
- Luo, C.; Liu, X.; Zhou, X.; Guo, J.; Truong, J.; Wang, X.; Chen, Z. Unusual biosynthesis and structure of locillomycins from Bacillus subtilis 916. Appl. Environ. Microbiol. 2015, 81, 6601–6609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, M.S.; Naresh, B.; Leela, T.; Prashanthi, M.; Madhusudhan, N.C.; Dhanasri, G.; Devi, P. Biodegradation of phenanthrene with biosurfactant production by a new strain of Brevibacillus sp. Bioresour. Technol. 2010, 101, 7980–7983. [Google Scholar] [CrossRef]
- Vigneshwaran, C.; Sivasubramanian, V.; Vasantharaj, K.; Krishnanand, N.; Jerold, M. Potential of Brevibacillus sp. AVN 13 isolated from crude oil contaminated soil for biosurfactant production and its optimization studies. J. Environ. Chem. Eng. 2018, 6, 4347–4356. [Google Scholar] [CrossRef]
- Hussain, T.; Khan, A.A. Determining the antifungal activity and characterization of Bacillus siamensis AMU03 against Macrophomina phaseolina (Tassi) Goid. Indian Phytopathol. 2020, 73, 507–516. [Google Scholar] [CrossRef]
- Zouari, I.; Masmoudi, F.; Medhioub, K.; Tounsi, S.; Trigui, M. Biocontrol and plant growth-promoting potentiality of bacteria isolated from compost extract. Antonie Van Leeuwenhoek 2020, 113, 2107–2122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, W.; Xue, Q. Biosurfactant production and oil degradation by Bacillus siamensis and its potential applications in enhanced heavy oil recovery. Int. Biodeterior. Biodegrad. 2022, 169, 105388. [Google Scholar] [CrossRef]
- Ahsan, N.; Shimizu, M. Lysinibacillus species: Their potential as Eeffective Bbioremediation, biostimulant, and biocontrol agents. Rev. Agric. Sci. 2021, 9, 103–116. [Google Scholar]
- John, W.C.; Ogbonna, I.O.; Gberikon, G.M.; Iheukwumere, C.C. Evaluation of biosurfactant production potential of Lysinibacillus fusiformis MK559526 isolated from automobile-mechanic-workshop soil. Braz. J. Microbiol. 2021, 52, 663–674. [Google Scholar]
- Chakraborty, J.; Chakrabarti, S.; Das, S. Characterization and antimicrobial properties of lipopeptide biosurfactants produced by Bacillus subtilis SJ301 and Bacillus vallismortis JB201. Appl. Biochem. Microbiol. 2014, 50, 609–618. [Google Scholar] [CrossRef]
- Das, M.; Borah, D.; Patowary, K.; Borah, M.; Khataniar, A.; Kakoti, B.B. Antimicrobial activity of silver nanoparticles synthesised by using microbial biosurfactant produced by a newly isolated Bacillus vallismortis MDU6 strain. IET Nanobiotechnol. 2019, 13, 967973. [Google Scholar] [CrossRef] [PubMed]
- Brint, J.M.; Ohman, D.E. Synthesis of multiple exoproducts in Pseudomonas aeruginosa is under the control of RhlR-RhlI, another set of regulators in strain PAO1 with homology to the autoinducer-responsive LuxR-LuxI family. J. Bacteriol. 1995, 177, 7155–7163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Jeon, J.W.; Lee, H.W.; Park, Y.I.; Seo, W.T.; Oh, H.M.; Yoon, B.D. Extracellular production of a glycolipid biosurfactant, mannosylerythritol lipid, from Candida antarctica. Biotechnol. Lett. 2002, 24, 225–229. [Google Scholar] [CrossRef]
- Cha, M.; Lee, N.; Kim, M.; Kim, M.; Lee, S. Heterologous production of Pseudomonas aeruginosa EMS1 biosurfactant in Pseudomonas putida. Bioresour. Technol. 2008, 99, 2192–2199. [Google Scholar] [CrossRef]
- Franzetti, A.; Caredda, P.; La Colla, P.; Pintus, M.; Tamburini, E.; Papacchini, M.; Bestetti, G. Cultural factors affecting biosurfactant production by Gordonia sp. BS29. Int. Biodeterior. Biodegrad. 2009, 63, 943–947. [Google Scholar] [CrossRef]
- Franzetti, A.; Gandolfi, I.; Bestetti, G.; Smyth, T.J.; Banat, I.M. Production and applications of trehalose lipid biosurfactants. Eur. J. Lipid Sci. Technol. 2010, 112, 617–627. [Google Scholar] [CrossRef]
- Perrier-Cornet, J. Comparison of emulsification efficiency of protein-stabilized oil-in-water emulsions using jet, high pressure and colloid mill homogenization. J. Food Eng. 2005, 66, 211–217. [Google Scholar] [CrossRef]
- Perfumo, A.; Smyth, T.; Marchant, R.; Banat, I. Production and roles of biosurfactants and bioemulsifiers in accessing hydrophobic substrates. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1501–1512. [Google Scholar]
- Teixeira Souza, K.S.; Gudiña, E.J.; Schwan, R.F.; Rodrigues, L.R.; Dias, D.R.; Teixeira, J.A. Improvement of biosurfactant production by Wickerhamomyces anomalus CCMA 0358 and its potential application in bioremediation. J. Hazard. Mater. 2018, 346, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Nurfarahin, A.H.; Mohamed, M.S.; Phang, L.Y. Culture medium development for microbial-derived surfactants production—an overview. Molecules 2018, 23, 1049. [Google Scholar] [CrossRef] [Green Version]
- Qazi, M.A.; Malik, Z.A.; Qureshi, G.D.; Hameed, A.; Ahmed, S. Yeast extractas the most preferable substrate for optimized biosurfactant production by rhlB Ggene Positive Pseudomonas putida SOL-10 isolate. Bioremediat. Biodegrad. 2013, 4, 2. [Google Scholar]
- Al-Ajlani, M.M.; Sheikh, M.A.; Ahmad, Z.; Hasnain, S. Production of surfactin from Bacillus subtilis MZ-7 grown on pharmamedia commercial medium. Microbial. Cell. Factories 2007, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Mawgoud, A.M.; Aboulwafa, M.M.; Hassouna, N.A.H. Microbial production of surfactants: Screening and identification of two promising isolates and their biosurfactants. Egypt. J. Biotechnol. 2007, 27, 166–185. [Google Scholar]
- Lee, S.; Lee, J.; Yu, H.; Lim, J. Synthesis of environment friendly biosurfactants and characterization of interfacial properties for cosmetic and household products formulations. Colloids Sur. A Physicochem. Eng. 2018, 536, 224–233. [Google Scholar] [CrossRef]
- Abouseoud, M.; Maachi, R.; Amrane, A.; Boudergua, S.; Nabi, A. Evaluation of different carbon and nitrogen sources in production of biosurfactant by Pseudomonas fluorescens. Desalination 2008, 223, 143–151. [Google Scholar] [CrossRef]
- Fonseca, R.R.; Silva, A.J.R.; De França, F.P.; Cardoso, V.L.; Sérvulo, E.F.C. Optimizing carbon/nitrogen ratio for biosurfactant production by a Bacillus subtilis strain. Appl. Biochem. Biotecnol. 2007, 7, 471–486. [Google Scholar]
- Al-Bahry, S.N.; Al-Wahaibi, Y.M.; Elshafie, A.E.; Al-Bemani, A.S.; Joshi, S.J. Isolation of biosurfactant producing spore-forming bacteria from Oman: Potential applications in bioremediation. Int. J. Earth Sci. 2014, 8, 77–79. [Google Scholar]
- Shaligram, S.; Kumbhare, S.V.; Dhotre, D.P.; Muddeshwar, M.G.; Kapley, A.; Joseph, N.; Pawar, S.P. Genomic and functional features of the biosurfactant producing Bacillus sp. AM13. Funct. Integr. Genom. 2016, 16, 557–566. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C/N Source Concentration | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Composition (%) | CN1 | CN2 | CN3 | CN4 | CN5 | CN6 | CN7 | CN8 | CN9 |
| Molasses | 1 | 1 | 1 | 2 | 2 | 2 | 3 | 3 | 3 |
| Yeast extract | 0.55 | 1.25 | 2 | 0.55 | 1.25 | 2 | 0.55 | 1.25 | 2 |
| Culture Conditions | |||||||||
| Culture condition | CC1 | CC2 | CC3 | CC4 | CC5 | CC6 | CC7 | CC8 | CC9 |
| NaCl (%) | 0.5 | 0.5 | 4.5 | 4.5 | 0.5 | 0.5 | 4.5 | 4.5 | 2.5 |
| pH | 6 | 8 | 6 | 8 | 6 | 8 | 6 | 8 | 7 |
| Planktonic agitation (rpm) | 180 | 180 | 180 | 180 | 120 | 120 | 120 | 120 | 150 |
| Sessile agitation (rpm) | 4 | 4 | 4 | 4 | 10 | 10 | 10 | 10 | 7 |
| Identification | Emulsification Index (%) | Drop-Collapse (mm) | Haemolytic Activity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Similarity (%) | GenBank Accessions | Nearest Phylogenetic Neighbour | Tolerance NaCl (%) | TSB | NaCl 5% | NaCl 10% | TSB | NaCl 5% | NaCl 10% | CFS (mm) |
| TZA46 | 97.73 | ON600577 | Brevibacillus sp. | 2.5 | 65.51 | 63.62 | 57.40 | 3.0 | 3.0 | 3.0 | 1.8 |
| XHA18 | 98.37 | ON600579 | Paenibacillus sp. * | 7.5 | 65.15 | 61.59 | 61.24 | 3.5 | 4.9 | 4.2 | (−) |
| TZA38 | 99.18 | ON600589 | Lysinibacillus fusiformis * | 5.0 | 65.10 | 63.20 | 60.33 | 4.1 | 3.1 | 3.4 | (−) |
| TZA34 | 94.00 | ON586647 | Pseudomonas aeruginosa | 7.5 | 64.38 | 64.43 | 55.79 | 3.0 | 4.0 | 4.0 | 2.1 |
| TZA10 | 88.00 | ON600528 | Pseudomonas sp. | 10.0 | 64.38 | 53.79 | 50.95 | 3.0 | 3.0 | 3.0 | 1.7 |
| XHA90 | 97.46 | ON600581 | Bacillus sp. | 20.0 | 63.94 | 62.22 | 62.04 | 3.0 | 4.0 | 3.0 | (−) |
| XHA66 | 96.84 | ON606036 | Cytobacillus sp. | 10.0 | 63.57 | 62.89 | 61.16 | 3.0 | 3.0 | 3.0 | (−) |
| XHA28 | 88.60 | ON600618 | Family Bacillaceae | 17.5 | 63.11 | 62.07 | 38.16 | 3.0 | 3.0 | 3.0 | (−) |
| TZA26 | 98.63 | ON600536 | Staphylococcus epidermidis | 15.0 | 61.69 | 55.82 | 6.32 | 4.1 | 3.0 | 3.0 | (−) |
| TZA51 | 99.32 | ON595357 | Lysinibacillus fusiformis | 5.0 | 61.61 | 61.91 | 58.63 | 3.0 | 3.0 | 3.0 | 0.2 |
| TZRP2 | 98.83 | ON600664 | Serratia rubidaea | 2.5 | 61.58 | 54.95 | 51.80 | 3.0 | 4.0 | 4.0 | (−) |
| TZA11 | 99.59 | ON600582 | Lysinibacillus sphaericus | 12.5 | 60.29 | 54.53 | 53.79 | 3.0 | 3.0 | 3.0 | (−) |
| XHA53 | 98.77 | ON599339 | Staphylococcus epidermidis | 10.0 | 59.66 | 53.68 | 53.32 | 4.5 | 4.0 | 3.0 | 1.8 |
| XHA78 | 98.83 | ON600578 | Pseudomonas parafulva | 5.0 | 59.60 | 41.27 | 36.89 | 3.0 | 3.0 | 3.0 | (−) |
| XHA33 | 98.60 | ON600531 | Staphylococcus sp. | 15.0 | 59.50 | 58.10 | 41.67 | 3.5 | 4.0 | 4.0 | 1.5 |
| TZA50 | 90.70 | ON606021 | Family Rhizobiaceae | 2.5 | 58.27 | 58.24 | 54.96 | 4.9 | 4.0 | 4.0 | (−) |
| TZS01 | 98.17 | ON599903 | Bacillus sp. * | 12.5 | 57.80 | 53.36 | 55.58 | 5.9 | 6.7 | 6.7 | 1.2 |
| XHA06 | 98.27 | ON600721 | Pseudomonas sp. | 12.5 | 57.73 | 60.94 | 54.95 | 6.2 | 5.3 | 4.7 | 1.7 |
| XHA16 | 98.77 | ON600576 | Bacillus vallismortis * | 12.5 | 57.73 | 60.94 | 54.95 | 7 | 7.8 | 7.2 | 1.7 |
| XHA14 | 99.11 | ON600533 | Bacillus siamensis * | 12.5 | 57.63 | 57.03 | 53.71 | 6.7 | 7.3 | 7.2 | 2 |
| TZA04 | 98.84 | ON600529 | Bacillus amyloliquefaciens | 12.5 | 56.11 | 54.95 | 52.83 | 5.6 | 4.0 | 4.0 | 1.6 |
| TZA15 | 82.30 | ON606020 | Family Sphingobacteriaceae | 7.5 | 55.95 | 64.37 | 44.94 | 3.0 | 4.0 | 4.5 | (−) |
| TZA47 | 95.69 | ON606022 | Brevibacillus sp. | 2.5 | 53.02 | 49.87 | 41.35 | 3.8 | 5.0 | 5.3 | (−) |
| XHA01 | 98.90 | ON600876 | Pseudomonas aeruginosa | 7.5 | 3.29 | 59.91 | 60.07 | 4.4 | 6.0 | 6.0 | 2.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maldonado Desena, F.; De la Cruz Ceferino, N.; Gómez Cornelio, S.; Alvarez Villagomez, C.; Herrera Candelario, J.L.; De la Rosa García, S. Bacteria Halotolerant from Karst Sinkholes as a Source of Biosurfactants and Bioemulsifiers. Microorganisms 2022, 10, 1264. https://doi.org/10.3390/microorganisms10071264
Maldonado Desena F, De la Cruz Ceferino N, Gómez Cornelio S, Alvarez Villagomez C, Herrera Candelario JL, De la Rosa García S. Bacteria Halotolerant from Karst Sinkholes as a Source of Biosurfactants and Bioemulsifiers. Microorganisms. 2022; 10(7):1264. https://doi.org/10.3390/microorganisms10071264
Chicago/Turabian StyleMaldonado Desena, Félix, Navila De la Cruz Ceferino, Sergio Gómez Cornelio, Carina Alvarez Villagomez, José Luis Herrera Candelario, and Susana De la Rosa García. 2022. "Bacteria Halotolerant from Karst Sinkholes as a Source of Biosurfactants and Bioemulsifiers" Microorganisms 10, no. 7: 1264. https://doi.org/10.3390/microorganisms10071264
APA StyleMaldonado Desena, F., De la Cruz Ceferino, N., Gómez Cornelio, S., Alvarez Villagomez, C., Herrera Candelario, J. L., & De la Rosa García, S. (2022). Bacteria Halotolerant from Karst Sinkholes as a Source of Biosurfactants and Bioemulsifiers. Microorganisms, 10(7), 1264. https://doi.org/10.3390/microorganisms10071264