
Simultaneous Detection of Five Foodborne Pathogens Using a Mini Automatic Nucleic Acid Extractor Combined with Recombinase Polymerase Amplification and Lateral Flow Immunoassay


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture Preparation and DNA Extraction
2.2. RPA Primers Design
2.3. Multiple RPA Procedure
2.4. Preparation of AuNPs and Lateral Flow Dipsticks Immunoassay
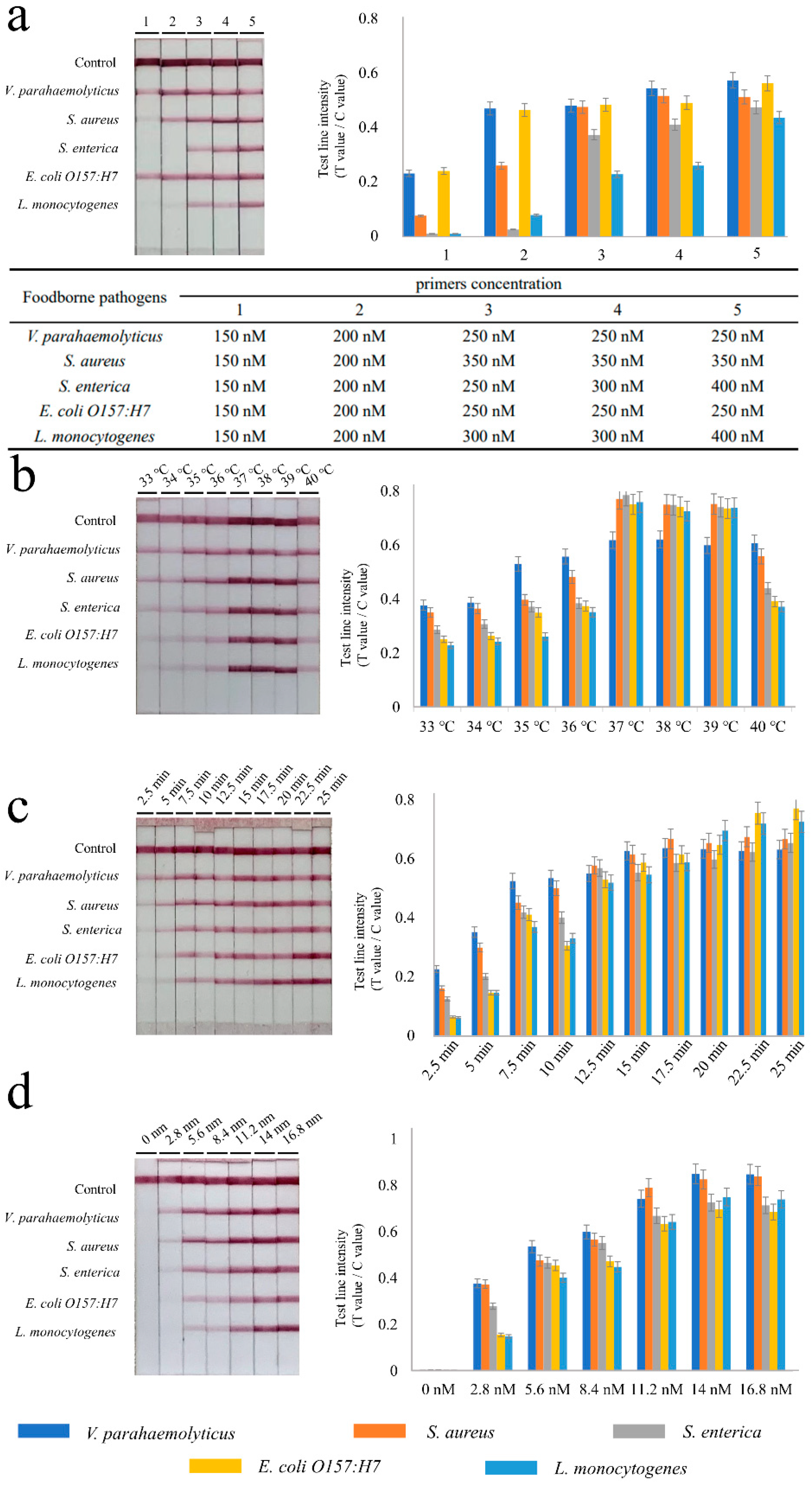
2.5. Optimization of the RPA-LFIA Conditions
2.6. Specificity and Sensitivity of Multiple RPA-LFIA Assay
2.7. Evaluation of Multiple RPA-LFIA in Artificially Contaminated Food Samples
2.8. Field Samples Testing
2.9. Statistical Analysis
3. Results
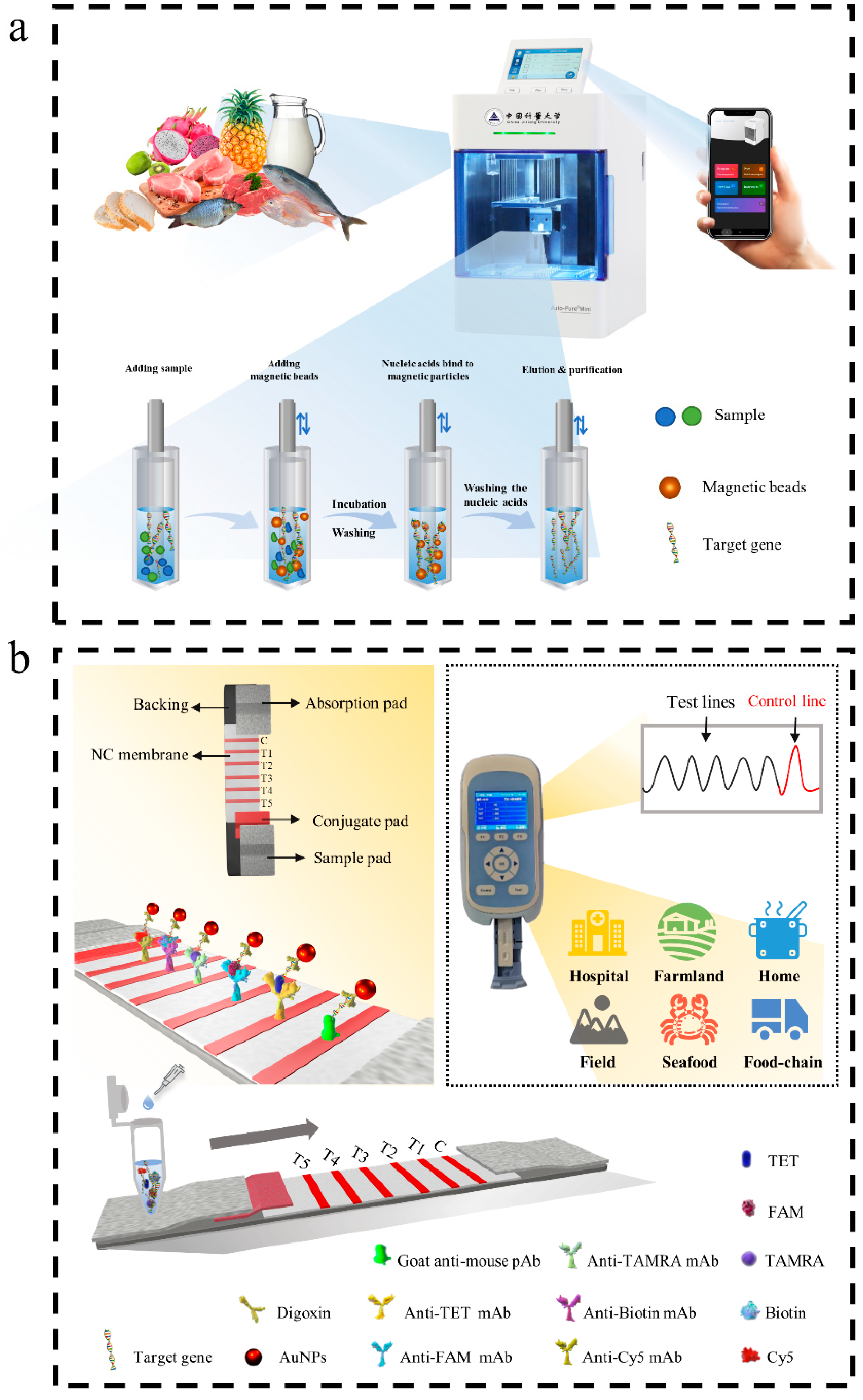
3.1. Assay Principle
3.2. Establishment and Optimization of Multiple RPA-LFIA Assay
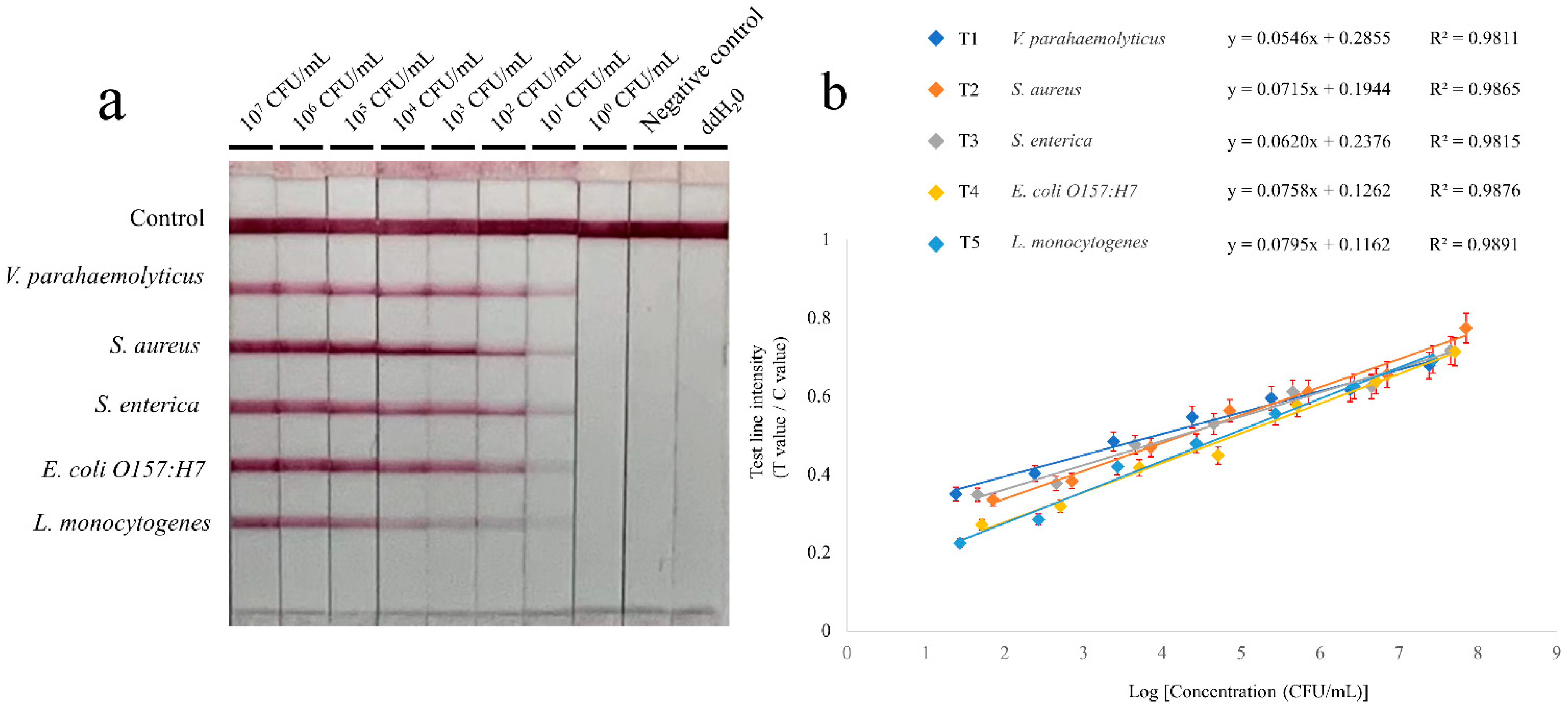
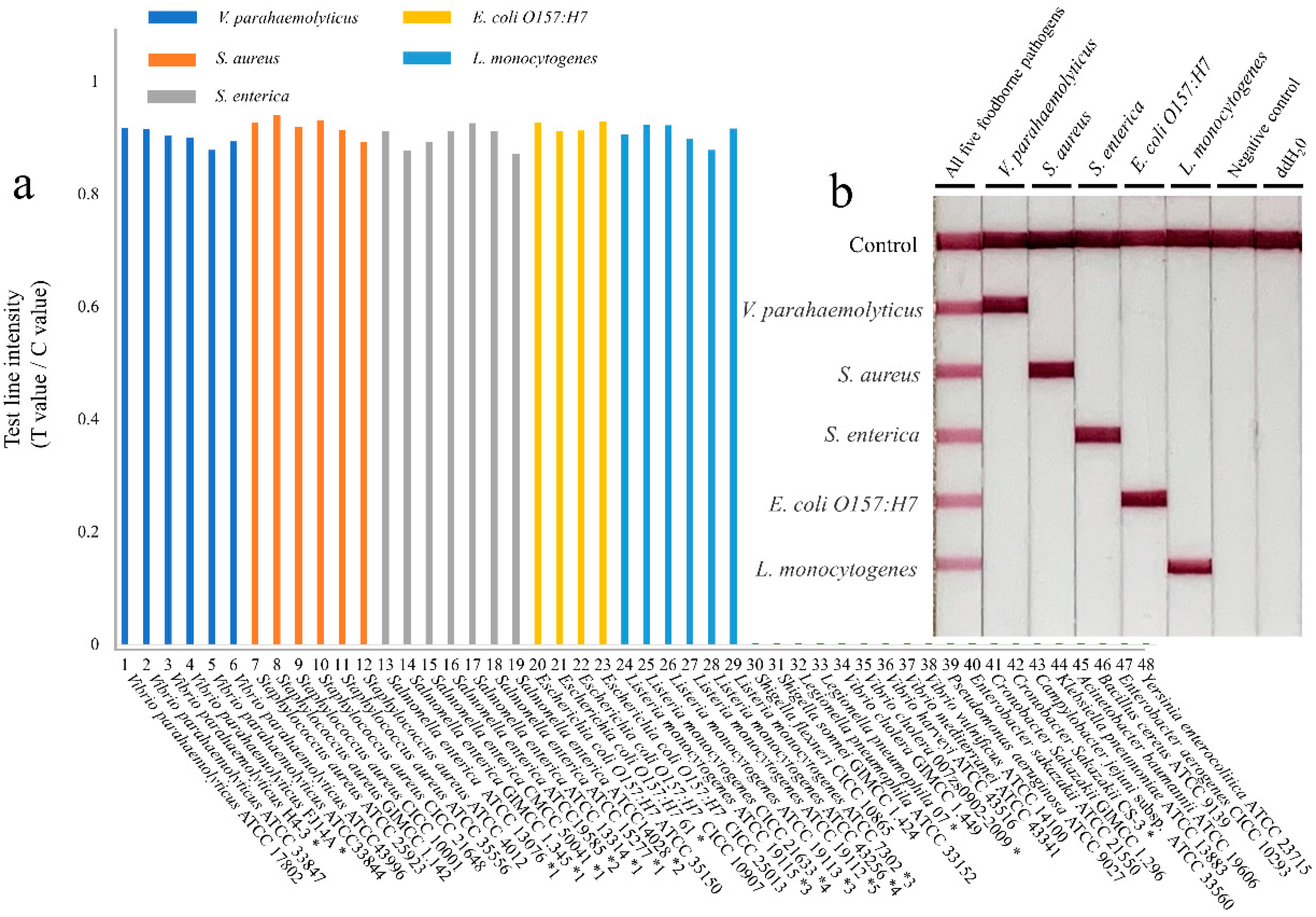
3.3. Sensitivity and Specificity of the Multiple RPA-LFIA Assay
3.4. Application of RPA-LFIA in Spiked Samples
3.5. Detection of Field Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paudyal, N.; Hang, P.; Liao, X.; Xian, Z.; Min, Y. A meta-analysis of major foodborne pathogens in chinese food commodities between 2006 and 2016. Foodborne Pathog. Dis. 2018, 15, 187–197. [Google Scholar] [CrossRef]
- Juru, T.; Lema, N.; Chirundu, D.; Garba, R.M.; Rimpong, J.A. Investigating a foodborne illness outbreak at a private girls’ school in mashonaland east province, Zimbabwe, 2015. Pan Afr. Med. J. 2018, 30, 15271. [Google Scholar] [CrossRef]
- Mei, Y.; He, C.; Zeng, W.; Luo, Y.; Liu, C.; Yang, M.; Kuang, Y.; Lin, X.; Huang, Q. Electrochemical biosensors for foodborne pathogens detection based on carbon nanomaterials: Recent advances and challenges. Food Bioproc. Tech. 2022, 15, 498–513. [Google Scholar] [CrossRef]
- Russini, V.; Corradini, C.; De Marchis, M.L.; Bogdanova, T.; Lovari, S.; De Santis, P.; Migliore, G.; Bilei, S.; Bossù, T. Foodborne toxigenic agents investigated in central Italy: An overview of a three-year experience (2018–2020). Toxins 2022, 14, 40. [Google Scholar] [CrossRef]
- Li, H.; Li, W.; Dai, Y.; Jiang, Y.; Liang, J.; Wang, S.; Zhuang, M.; Huang, Z.; Xu, L.; Xue, B.; et al. Characteristics of settings and etiologic agents of foodborne disease outbreaks- China, 2020. China CDC Wkly. 2021, 3, 889–893. [Google Scholar] [CrossRef]
- Gao, W.; Huang, H.; Zhu, P.; Yan, X.; Fan, J.; Jiang, J.; Xu, J. Recombinase polymerase amplification combined with lateral flow dipstick for equipment-free detection of Salmonella in shellfish. Bioprocess Biosyst. Eng. 2018, 41, 603–611. [Google Scholar] [CrossRef]
- Adley, C.C. Past, present and future of sensors in food production. Foods 2014, 3, 491–510. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Liu, S.; Wang, J.; Nan, H.; Liu, L.; Sun, X.; Li, D.; Liu, M.; Wang, J.; Tan, K. Rapid detection of Staphylococcus aureus in food using a recombinase polymerase amplification-based assay. Food Anal. Methods 2018, 11, 2847–2856. [Google Scholar] [CrossRef]
- Castro, A.; Palhau, C.; Cunha, S.; Camarinha, S.; Teixeira, P. Virulence and resistance profile of Staphylococcus aureus isolated from food. Acta Aliment. Hung 2017, 46, 231–237. [Google Scholar] [CrossRef]
- Castro, V.S.; Figueiredo, E.E.d.S.; Stanford, K.; McAllister, T.; Conte-Junior, C.A. Shiga-toxin producing Escherichia coli in Brazil: A systematic review. Microorganisms 2019, 7, 137. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Wang, Y.; Su, H.; Ding, H.; Sun, X.; Gao, H.; Geng, Y.; Wang, Z. Rapid analysis of Escherichia coli O157:H7 using isothermal recombinase polymerase amplification combined with triple-labeled nucleotide probes. Mol. Cell. Probes 2020, 50, 101501. [Google Scholar] [CrossRef]
- Horlbog, J.A.; Stevens, M.J.A.; Stephan, R.; Guldimann, C. Global transcriptional response of three highly acid-tolerant field strains of Listeria monocytogenes to HCl stress. Microorganisms 2019, 7, 455. [Google Scholar] [CrossRef] [Green Version]
- Nfdsa, C.; Mmpsn, A.; Jmcsm, B.; Cf, C.; Dm, A. Emerging electrochemical biosensing approaches for detection of Listeria monocytogenes in food samples: An overview- ScienceDirect. Trends Food Sci. Technol. 2020, 99, 621–633. [Google Scholar]
- Shin, H.H.; Hwang, B.H.; Cha, H.J. Multiplex 16S rRNA-derived geno-biochip for detection of 16 bacterial pathogens from contaminated foods. Biotechnol. J. 2016, 11, 1405–1414. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ma, L.; Ma, L.; Hua, M.Z.; Wang, S.; Lu, X. Rapid detection of methicillin-resistant Staphylococcus aureus in pork using a nucleic acid-based lateral flow immunoassay. Int. J. Food Microbiol. 2017, 243, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Crannell, Z.; Castellanos-Gonzalez, A.; Nair, G.; Mejia, R.; White, A.C.; Richards-Kortum, R. Multiplexed recombinase polymerase amplification assay to detect intestinal protozoa. Anal Chem. 2016, 88, 1610–1616. [Google Scholar] [CrossRef]
- Kérouanton, A.; Marault, M.; Petit, L.; Grout, J.; Dao, T.T.; Brisabois, A. Evaluation of a multiplex PCR assay as an alternative method for Listeria monocytogenes serotyping. J. Microbiol. Methods 2010, 80, 134–137. [Google Scholar] [CrossRef]
- Espineira, M.; Vieites, J.M. Rapid method for controlling the correct labeling of products containing common octopus (Octopus vulgaris) and main substitute species (Eledone cirrhosa and Dosidicus gigas) by fast real-time PCR. Food Chem. 2012, 135, 2439–2444. [Google Scholar] [CrossRef]
- Law, J.W.F.; Ab Mutalib, N.S.; Chan, K.G.; Lee, L.H. Rapid methods for the detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2015, 5, 770. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Y.; Lan, R.; Xu, H.; Ma, A.; Li, D.; Dai, H.; Yuan, X.; Xu, J.; Ye, C. Multiple endonuclease restriction real-time loop-mediated isothermal amplification: A novel analytically rapid, sensitive, multiplex loop-mediated isothermal amplification detection technique. J. Mol. Diagn. 2015, 17, 392–401. [Google Scholar] [CrossRef]
- James, A.; Macdonald, J. Recombinase polymerase amplification: Emergence as a critical molecular technology for rapid, low-resource diagnostics. Expert Rev. Mol. Diagn 2015, 15, 1475–1489. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ye, Z.; Ying, Y. New trends in impedimetric biosensors for the detection of foodborne pathogenic bacteria. Sensors 2012, 12, 3449–3471. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, Y.; Yang, Q.; Fan, H.; Liang, W. A Rapid and sensitive detection method for Pseudomonas aeruginosa using visualized recombinase polymerase amplification and lateral flow strip technology. Front. Cell. Infect. Microbiol. 2021, 11, 698929. [Google Scholar] [CrossRef]
- Wang, P.; Liao, L.; Ma, C.; Zhang, X.; Yu, J.; Yi, L.; Liu, X.; Shen, H.; Gao, S.; Lu, Q. Duplex on-site detection of Vibrio cholerae and Vibrio vulnificus by recombinase polymerase amplification and three-segment lateral flow strips. Biosensors 2021, 11, 151. [Google Scholar] [CrossRef]
- Bao, H.; Yuan, M.; Xiao, C.; Liu, D.; Lai, W. Development of a signal-enhanced LFIA based on tyramine-induced AuNPs aggregation for sensitive detection of danofloxacin. Food Chem. 2022, 375, 131875. [Google Scholar] [CrossRef]
- Ma, B.; Li, J.; Chen, K.; Yu, X.; Sun, C.; Zhang, M. Multiplex recombinase polymerase amplification assay for the simultaneous detection of three foodborne pathogens in seafood. Foods 2020, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Ma, B.; Li, J.; Chen, E.; Xu, Y.; Yu, X.; Sun, C.; Zhang, M. A rapid and sensitive europium nanoparticle-based lateral flow immunoassay combined with recombinase polymerase amplification for simultaneous detection of three food-borne pathogens. Int. J. Environ. Res. Public Health 2021, 18, 4574. [Google Scholar] [CrossRef]
- Borse, V.B.; Konwar, A.N.; Jayant, R.D.; Patil, P.O. Perspectives of characterization and bioconjugation of gold nanoparticles and their application in lateral flow immunosensing. Drug Deliv. Transl. Res. 2020, 10, 878–902. [Google Scholar] [CrossRef]
- Bacteriological Analytical Manual. Available online: https://www.fda.gov/food/laboratory-methods-food/bacteriological-analytical-manual-bam (accessed on 28 September 2021).
- Shan, S.; Lai, W.; Xiong, Y.; Wei, H.; Xu, H. Novel strategies to enhance lateral flow immunoassay sensitivity for detecting foodborne pathogens. J. Agric. Food Chem. 2015, 63, 745–753. [Google Scholar] [CrossRef]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trends Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef]
- Mayboroda, O.; Katakis, I.; O’Sullivan, C.K. Multiplexed isothermal nucleic acid amplification. Anal. Biochem. 2018, 545, 20–30. [Google Scholar] [CrossRef]
- Ahmed, A.; Van der Linden, H.; Hartskeerl, R.A. Development of a recombinase polymerase amplification assay for the detection of pathogenic Leptospira. Int. J. Environ. Res. 2014, 11, 4953–4964. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Q.Y.; Tan, B.; Shi, P.F.; Qiao, L.J.; Li, Z.J.; Liu, K.X.; Cao, Z.G.; Zhang, S.Q.; Sun, F.Y. A lateral flow dipstick combined with reverse transcription recombinase polymerase amplification for rapid and visual detection of the BVDV and BPIV3. J. Virol. Methods 2022, 299, 114343. [Google Scholar] [CrossRef]
- Lau, H.Y.; Wang, Y.; Wee, E.J.H.; Botella, J.R.; Trau, M. Field demonstration of a multiplexed point-of-care diagnostic platform for plant pathogens. Anal. Chem. 2016, 88, 8074–8081. [Google Scholar] [CrossRef]
- Kersting, S.; Rausch, V.; Bier, F.F.; von Nickisch-Rosenegk, M. Multiplex isothermal solid-phase recombinase polymerase amplification for the specific and fast DNA-based detection of three bacterial pathogens. Mikrochim. Acta 2014, 181, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-H.; Hwang, H.J.; Kim, J.H. Ultra-fast on-site molecular detection of foodborne pathogens using a combination of convection polymerase chain reaction and nucleic acid lateral flow immunoassay. Foodborne Pathog. Dis. 2019, 16, 144–151. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, S.; Zhao, Y.; Yang, X.; Fu, S.; McKillip, J.L.; Fox, E.M.; Man, C. Multiplex loop-mediated isothermal amplification-based lateral flow dipstick for simultaneous detection of 3 food-borne pathogens in powdered infant formula. Int. J. Dairy Sci. 2020, 103, 4002–4012. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, J.-L. Development of a multiplex real-time recombinase polymerase amplification (RPA) assay for rapid quantitative detection of Campylobacter coli and jejuni from eggs and chicken products. Food Control 2017, 73, 1247–1255. [Google Scholar] [CrossRef] [Green Version]
- Choi, G.; Jung, J.H.; Park, B.H.; Oh, S.J.; Seo, J.H.; Choi, J.S.; Kim, D.H.; Seo, T.S. A centrifugal direct recombinase polymerase amplification (direct-RPA) microdevice for multiplex and real-time identification of food poisoning bacteria. Lab Chip 2016, 16, 2309–2316. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ID of Strains | Multiple RPA-LFIA Test Results | ||||
|---|---|---|---|---|---|---|
| toxR | nuc | fimY | rfbE | hlyA | ||
| Vibrio parahaemolyticus | ATCC 17802 | + | - | - | - | - |
| Vibrio parahaemolyticus | ATCC 33847 | + | - | - | - | - |
| Vibrio parahaemolyticus | H4-3 * | + | - | - | - | - |
| Vibrio parahaemolyticus | FJ14A * | + | - | - | - | - |
| Vibrio parahaemolyticus | ATCC33844 | + | - | - | - | - |
| Vibrio parahaemolyticus | ATCC43996 | + | - | - | - | - |
| Staphylococcus aureus | ATCC 25923 | - | + | - | - | - |
| Staphylococcus aureus | GIMCC 1.142 | - | + | - | - | - |
| Staphylococcus aureus | CICC 10001 | - | + | - | - | - |
| Staphylococcus aureus | CICC 21648 | - | + | - | - | - |
| Staphylococcus aureus | ATCC 35556 | - | + | - | - | - |
| Staphylococcus aureus | ATCC 4012 | - | + | - | - | - |
| Salmonella enterica | ATCC 13076 *1 | - | - | + | - | - |
| Salmonella enterica | GIMCC 1.345 *1 | - | - | + | - | - |
| Salmonella enterica | CMCC 50041 *1 | - | - | + | - | - |
| Salmonella enterica | ATCC19585 *2 | - | - | + | - | - |
| Salmonella enterica | ATCC 13314 *1 | - | - | + | - | - |
| Salmonella enterica | ATCC 15277 *1 | - | - | + | - | - |
| Salmonella enterica | ATCC14028 *2 | - | - | + | - | - |
| Escherichia coli O157:H7 | ATCC 35150 | - | - | - | + | - |
| Escherichia coli O157:H7 | 61 * | - | - | - | + | - |
| Escherichia coli O157:H7 | CICC 10907 | - | - | - | + | - |
| Escherichia coli O157:H7 | CICC 25013 | - | - | - | + | - |
| Listeria monocytogenes | ATCC 19115 *3 | - | - | - | - | + |
| Listeria monocytogenes | CICC 21633 *4 | - | - | - | - | + |
| Listeria monocytogenes | ATCC 19113 *3 | - | - | - | - | + |
| Listeria monocytogenes | ATCC 19112 *5 | - | - | - | - | + |
| Listeria monocytogenes | ATCC 43256 *4 | - | - | - | - | + |
| Listeria monocytogenes | ATCC 7302 *3 | - | - | - | - | + |
| Shigella flexneri | CICC 10865 | - | - | - | - | - |
| Shigella sonnei | GIMCC 1.424 | - | - | - | - | - |
| Legionella pneumophila | ATCC 33152 | - | - | - | - | - |
| Legionella pneumophila | 07 * | - | - | - | - | - |
| Vibrio cholera | GIMCC 1.449 | - | - | - | - | - |
| Vibrio cholera | 007zs0902-2009 * | - | - | - | - | - |
| Vibrio harveyi | ATCC 43516 | - | - | - | - | - |
| Vibrio mediterranei | ATCC 43341 | - | - | - | - | - |
| Vibrio vulnificus | ATCC 14100 | - | - | - | - | - |
| Pseudomonas aeruginosa | ATCC 9027 | - | - | - | - | - |
| Enterobacter sakazakii | ATCC 21550 | - | - | - | - | - |
| Cronobacter Sakazakii | GIMCC 1.296 | - | - | - | - | - |
| Cronobacter Sakazakii | CS-3 * | - | - | - | - | - |
| Campylobacter jejuni subsp. | ATCC 33560 | - | - | - | - | - |
| Klebsiella pneumoniae | ATCC 13883 | - | - | - | - | - |
| Acinetobacter baumannii | ATCC 19606 | - | - | - | - | - |
| Bacillus cereus | ATCC 9139 | - | - | - | - | - |
| Enterobacter aerogenes | CICC 10293 | - | - | - | - | - |
| Yersinia enterocolitica | ATCC 23715 | - | - | - | - | - |
| Target Name | Target Name | Sequence (5′-3′) | Modifications | Amplification Size | Reference |
|---|---|---|---|---|---|
| V. parahaemolyticus | toxR-RPA F(forward primer) | TTTGTTTGGCGTGAGCAAGGTTTTGAGGTG | 5′-TET | 230 bp | [26] |
| toxR-RPA R(reverse primer) | GCAGAGGCGTCATTGTTATCAGAAGCAGGT | 5′-Digoxin | |||
| S. aureus | nuc-RPA F(forward primer) | CTTATAGGGATGGCTATCAGTAATGTTTCG | 5′-FAM | 158 bp | |
| nuc-RPA R(reverse primer) | CCACTTCTATTTACGCCGTTATCTGTTTGT | 5′-Digoxin | |||
| S. enterica | fimY-RPA F(forward primer) | TATCAGATAAAACCTCCGCTATAACACAGT | 5′-TAMRA | 133 bp | |
| fimY-RPA R(reverse primer) | CTTTCCGATAAGCGAGGTTTGGAGGCTGAT | 5′-Digoxin | |||
| E. coli O157:H7 | rfbE-RPA F(forward primer) | TATCTGCAAGGTGATTCCTTGATGGTCTCA | 5′-Biotin | 176 bp | [27] |
| rfbE-RPA R(reverse primer) | AGGCCAGTTACCATCCTCAGCTATAGGGTG | 5′-Digoxin | |||
| L. monocytogenes | hlyA-RPA F(forward primer) | CGATCACTCTGGAGGATACGTTGCTCAATT | 5′-Cy5 | 154 bp | |
| hlyA-RPA R(reverse primer) | TTACCAGGCAAATAGATGGACGATGTGAAA | 5′-Digoxin |
| Samples | No. of Samples | V. parahaemolyticus | S. aureus | S. enterica | E. coli O157:H7 | L. monocytogenes | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| RPA-LFIA | BAM | RPA-LFIA | BAM | RPA-LFIA | BAM | RPA-LFIA | BAM | RPA-LFIA | BAM | ||
| Milk | 12 | - | - | - | - | - | - | - | - | - | - |
| Raw pork | 10 | - | - | - | - | - | - | 1 | 1 | - | - |
| Eggs | 9 | - | - | - | - | - | - | - | - | - | - |
| Chicken | 9 | - | - | - | - | - | - | - | - | - | - |
| Cheese | 9 | - | - | - | - | - | - | - | - | - | - |
| Raw shrimp | 8 | 3 | 3 | - | - | 2 | 2 | - | - | - | - |
| Fish | 7 | - | - | - | - | - | - | - | - | - | - |
| Codfish | 6 | - | - | - | - | - | - | - | - | - | - |
| Broccoli | 5 | - | - | - | - | - | - | - | - | - | - |
| Fruit juice | 5 | - | - | - | - | - | - | - | - | - | - |
| Total | 80 | 3 | 3 | - | - | 2 | 2 | 1 | 1 | - | - |
| Positive Detection rate | / | 3.75% | 3.75% | 0% | 0% | 2.50% | 2.50% | 1.25% | 1.25% | 0% | 0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, B.; Ma, B.; Li, J.; Hong, Y.; Zhang, M. Simultaneous Detection of Five Foodborne Pathogens Using a Mini Automatic Nucleic Acid Extractor Combined with Recombinase Polymerase Amplification and Lateral Flow Immunoassay. Microorganisms 2022, 10, 1352. https://doi.org/10.3390/microorganisms10071352
Jin B, Ma B, Li J, Hong Y, Zhang M. Simultaneous Detection of Five Foodborne Pathogens Using a Mini Automatic Nucleic Acid Extractor Combined with Recombinase Polymerase Amplification and Lateral Flow Immunoassay. Microorganisms. 2022; 10(7):1352. https://doi.org/10.3390/microorganisms10071352
Chicago/Turabian StyleJin, Bei, Biao Ma, Jiali Li, Yi Hong, and Mingzhou Zhang. 2022. "Simultaneous Detection of Five Foodborne Pathogens Using a Mini Automatic Nucleic Acid Extractor Combined with Recombinase Polymerase Amplification and Lateral Flow Immunoassay" Microorganisms 10, no. 7: 1352. https://doi.org/10.3390/microorganisms10071352
APA StyleJin, B., Ma, B., Li, J., Hong, Y., & Zhang, M. (2022). Simultaneous Detection of Five Foodborne Pathogens Using a Mini Automatic Nucleic Acid Extractor Combined with Recombinase Polymerase Amplification and Lateral Flow Immunoassay. Microorganisms, 10(7), 1352. https://doi.org/10.3390/microorganisms10071352