
Beneficial Shifts in Gut Microbiota by Lacticaseibacillus rhamnosus R0011 and Lactobacillus helveticus R0052 in Alcoholic Hepatitis

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
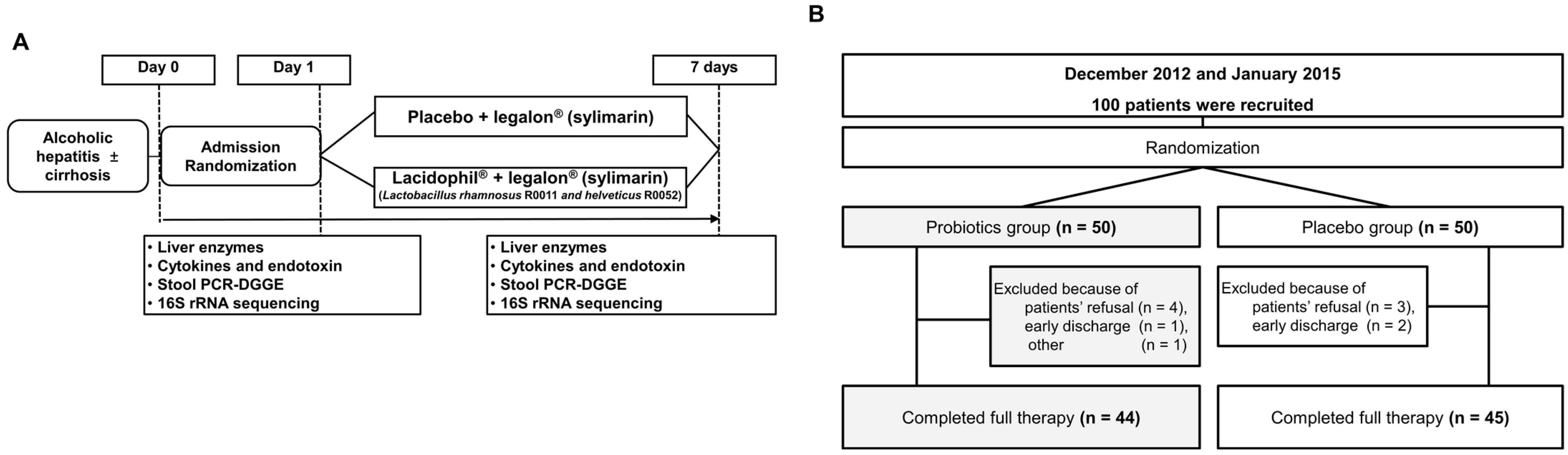
2.1. Study Design and Participants
2.2. Study Treatment
2.3. Randomization and Blinding
2.4. Quantitative Analysis of Cytokines and LPS
2.5. Sample Preparation and DNA Extraction
2.6. Polymerase Chain Reaction-Denaturing Gradient Gel Electrophoresis
2.7. Stool Analysis for the Metagenomics
2.8. Statistical Analysis
3. Results
3.1. Abstinence and Probiotics Improved Liver Function in Alcoholic Hepatitis
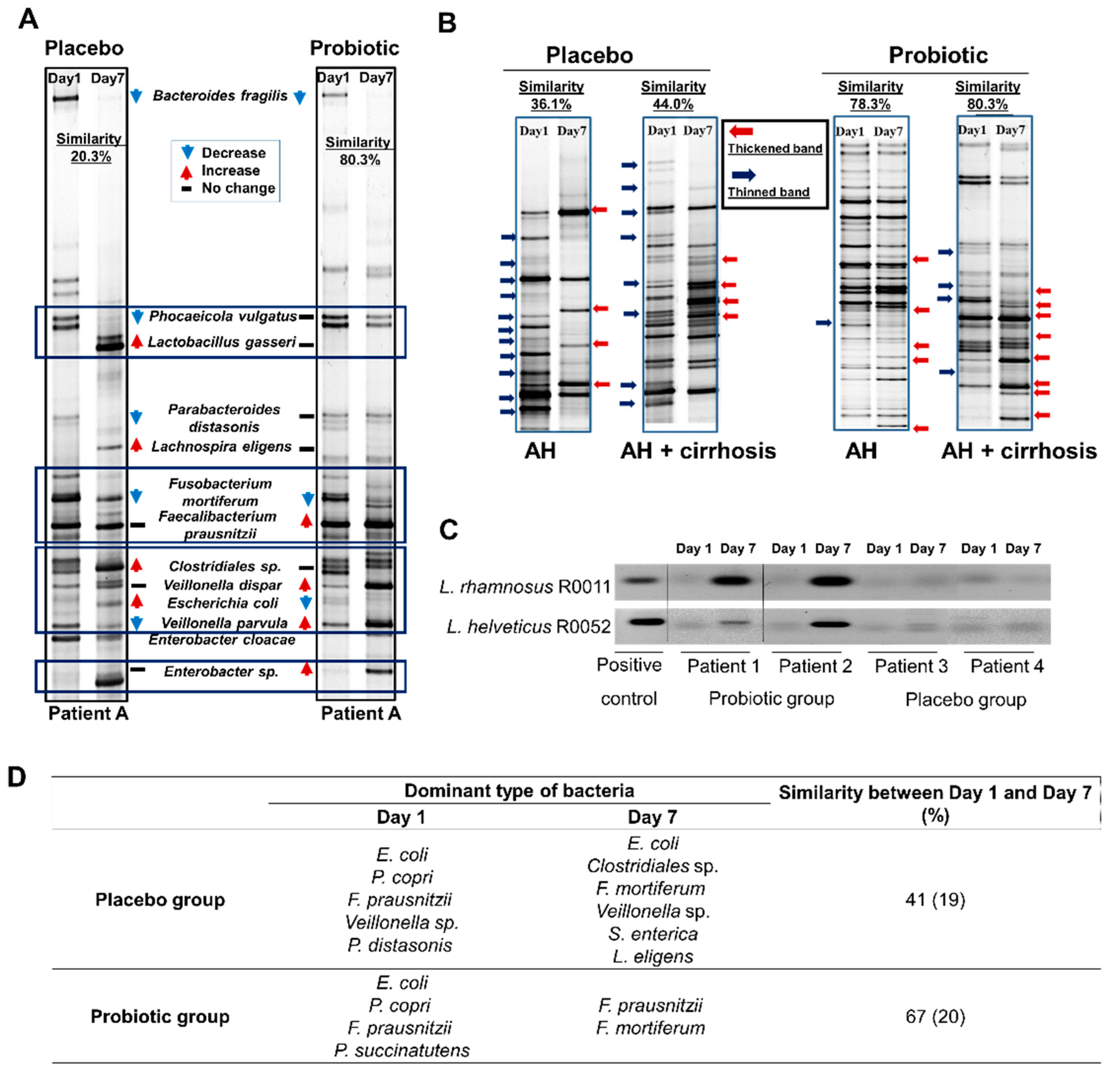
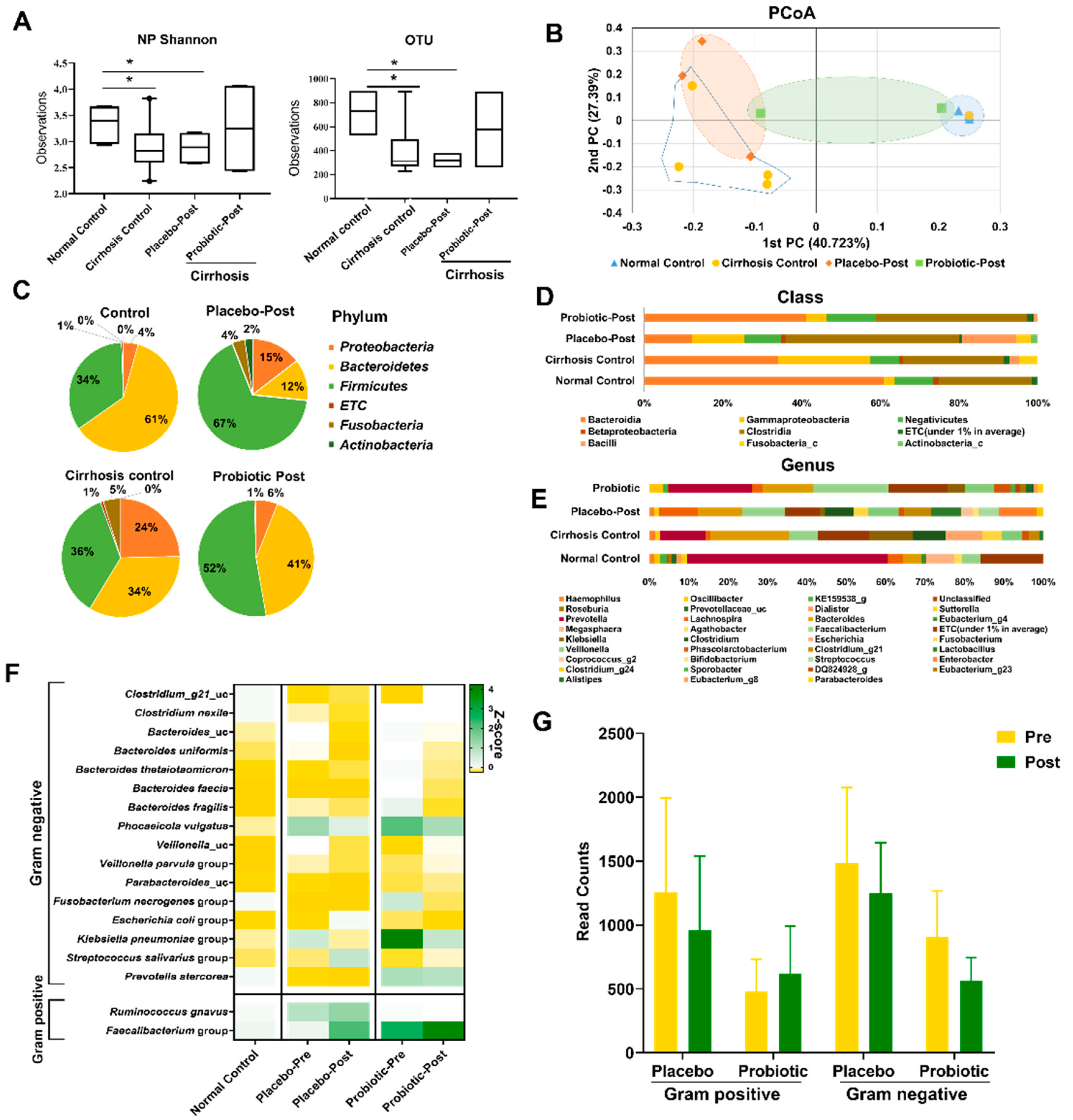
3.2. Probiotics Alter the Gut Microbiota in Patients with Alcohol Hepatitis
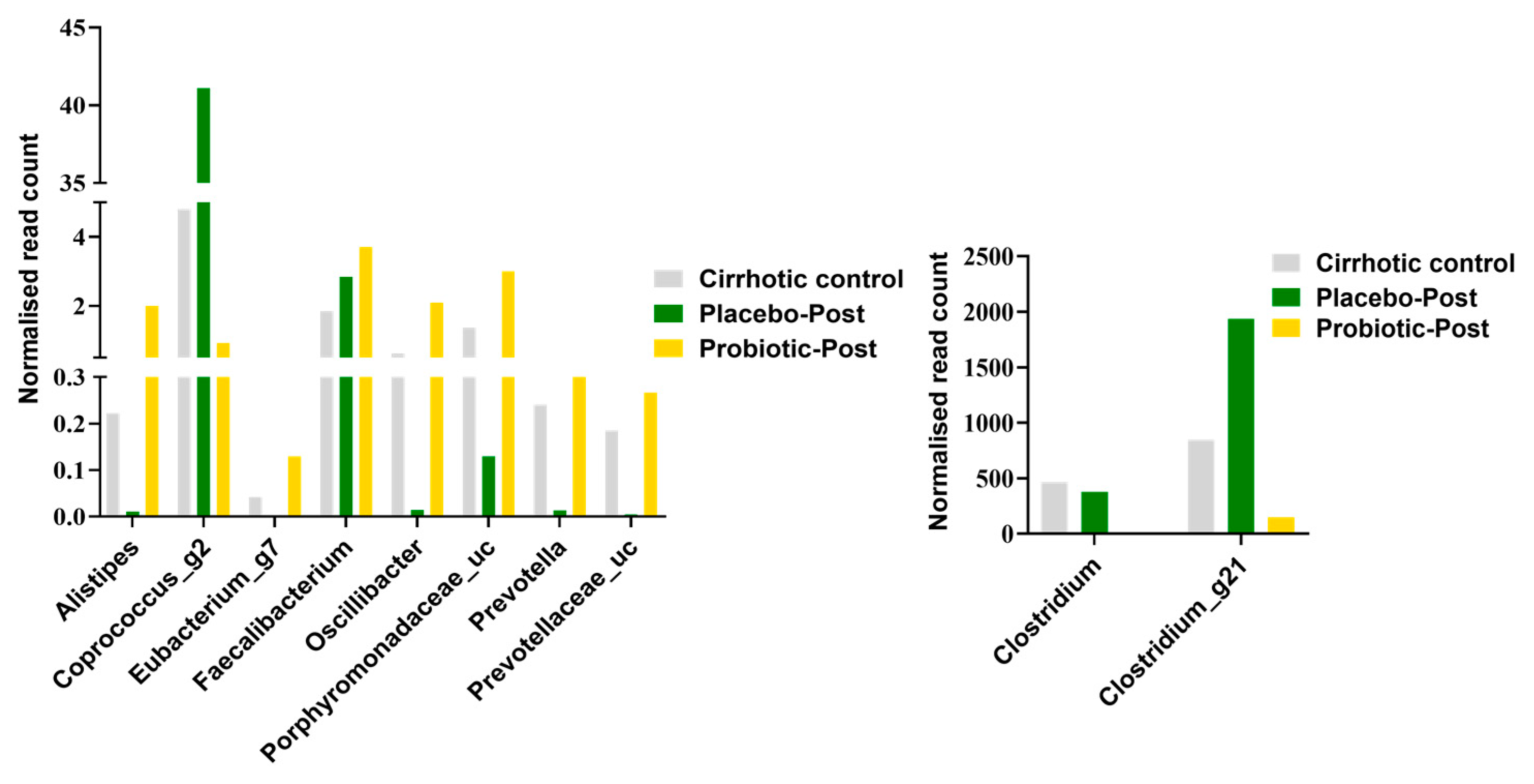
3.3. Lactobacillus Supplementation Modulates Alcohol-Induced Dysbiosis
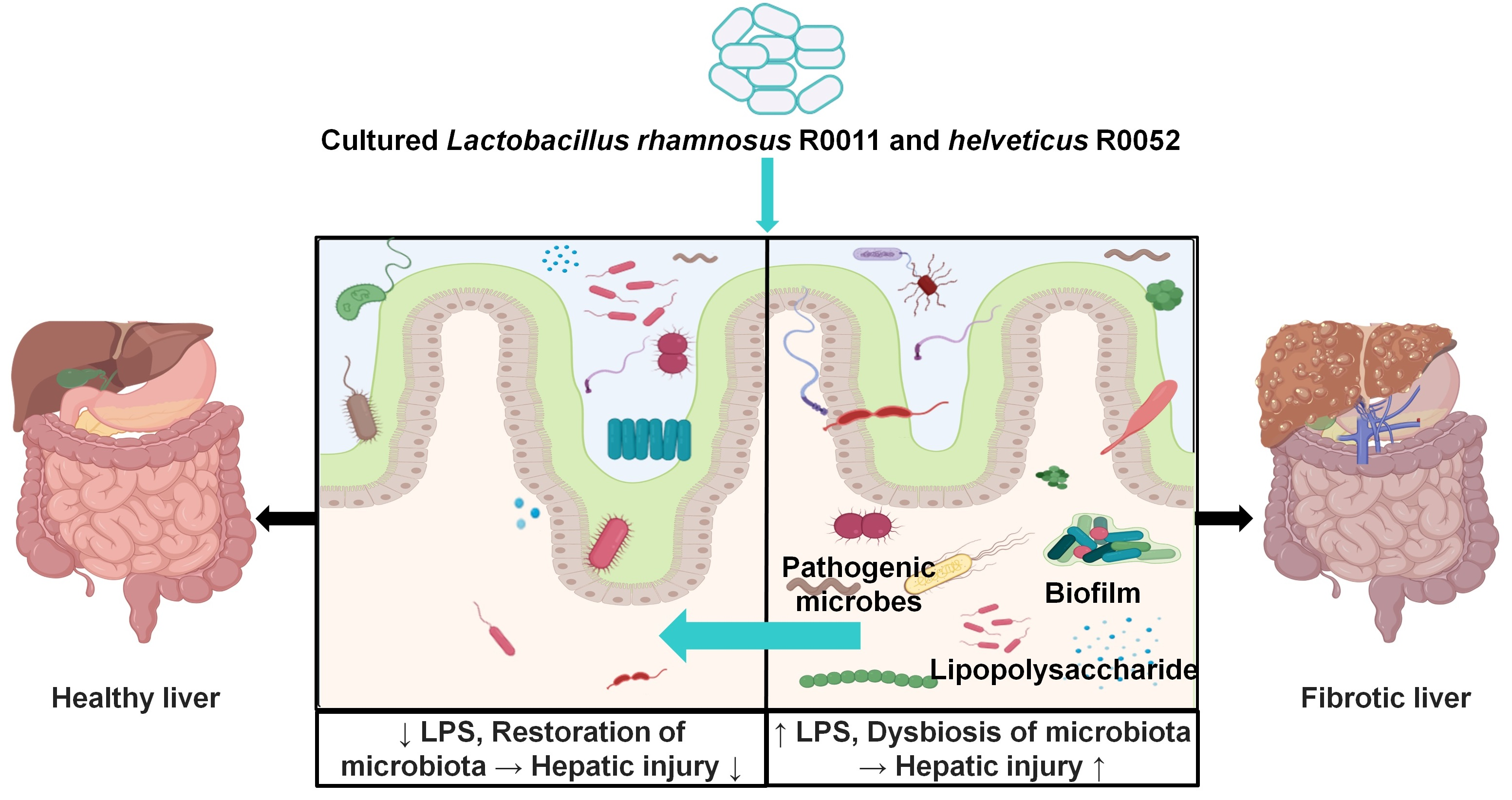
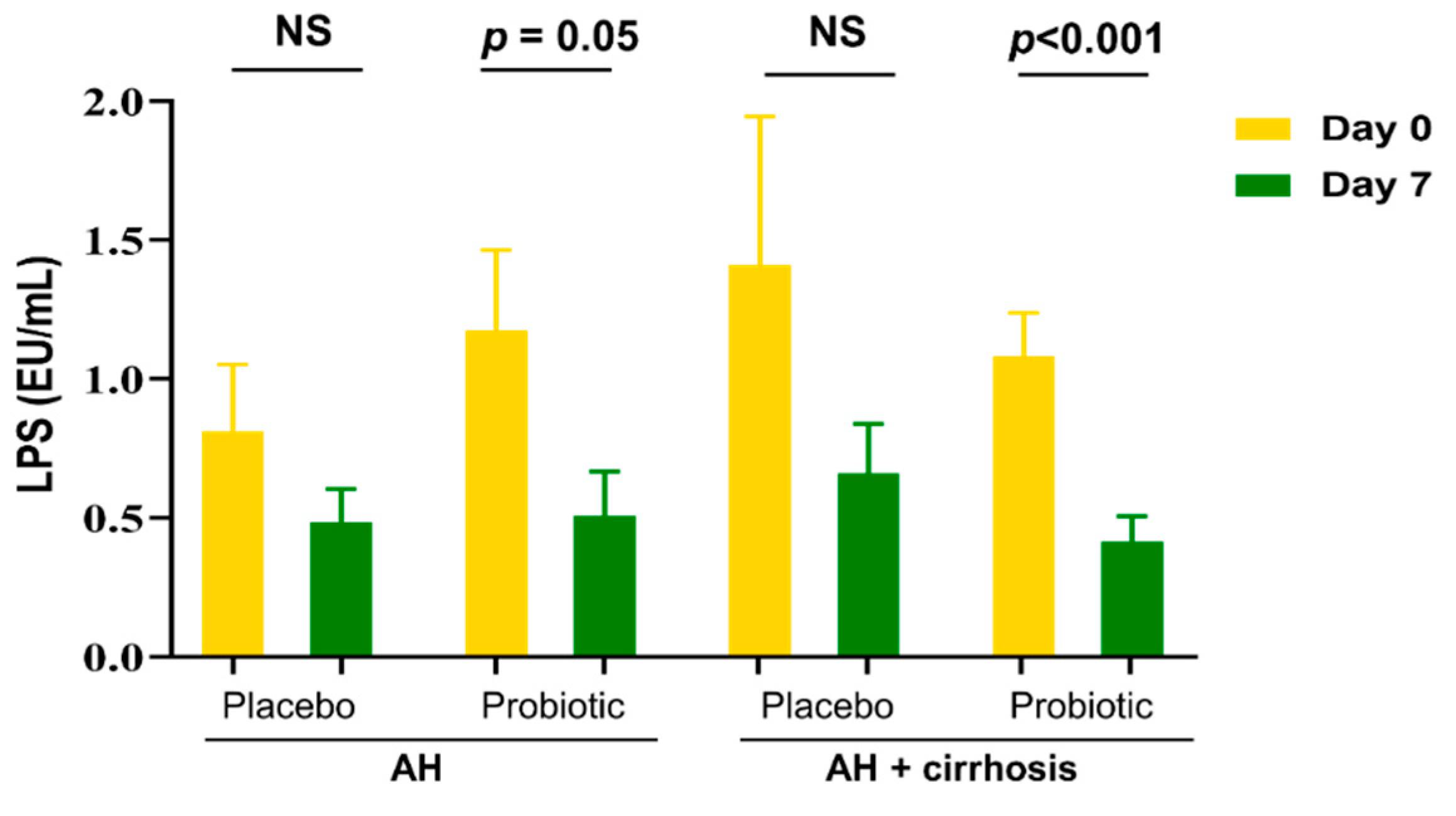
3.4. Probiotics Reduced Endotoxin Levels by Improving the Composition of Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lucey, M.R.; Mathurin, P.; Morgan, T.R. Alcoholic hepatitis. N. Engl. J. Med. 2009, 360, 2758–2769. [Google Scholar] [CrossRef] [PubMed]
- Chayanupatkul, M.; Liangpunsakul, S. Alcoholic hepatitis: A comprehensive review of pathogenesis and treatment. World J. Gastroenterol. 2014, 20, 6279–6286. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J.; Shield, K.D. Global alcohol-attributable deaths from cancer, liver cirrhosis, and injury in 2010. Alcohol Res. 2013, 35, 174–183. [Google Scholar] [PubMed]
- World Health Organization. Global Status Report on Alcohol and Health; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Jia, W.; Li, H.; Zhao, L.; Nicholson, J.K. Gut microbiota: A potential new territory for drug targeting. Nat. Rev. Drug Discov. 2008, 7, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, B.O.; Backhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Miele, L.; Marrone, G.; Lauritano, C.; Cefalo, C.; Gasbarrini, A.; Day, C.; Grieco, A. Gut-liver axis and microbiota in NAFLD: Insight pathophysiology for novel therapeutic target. Curr. Pharm. Des. 2013, 19, 5314–5324. [Google Scholar] [CrossRef]
- Schnabl, B.; Brenner, D.A. Interactions between the intestinal microbiome and liver diseases. Gastroenterology 2014, 146, 1513–1524. [Google Scholar] [CrossRef] [Green Version]
- Haque, T.R.; Barritt, A.S.t. Intestinal microbiota in liver disease. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 133–142. [Google Scholar] [CrossRef]
- Bajaj, J.S. The role of microbiota in hepatic encephalopathy. Gut Microbes 2014, 5, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Mathurin, P.; Deng, Q.G.; Keshavarzian, A.; Choudhary, S.; Holmes, E.W.; Tsukamoto, H. Exacerbation of alcoholic liver injury by enteral endotoxin in rats. Hepatology 2000, 32, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Parlesak, A.; Schafer, C.; Schutz, T.; Bode, J.C.; Bode, C. Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease. J. Hepatol. 2000, 32, 742–747. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Y.; Wang, Y.; Li, Y.; Zhang, X.; Zheng, H.; Ma, F.; Ma, C.; Lu, B.; Xie, Z.; et al. Beneficial changes of gut microbiota and metabolism in weaned rats with Lactobacillus acidophilus NCFM and Bifidobacterium lactis Bi-07 supplementation. J. Funct. Foods 2018, 48, 252–265. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhong, W. Targeting the gut barrier for the treatment of alcoholic liver disease. Liver Res. 2017, 1, 197–207. [Google Scholar] [CrossRef]
- Tenorio-Jimenez, C.; Martinez-Ramirez, M.J.; Tercero-Lozano, M.; Arraiza-Irigoyen, C.; Del Castillo-Codes, I.; Olza, J.; Plaza-Diaz, J.; Fontana, L.; Migueles, J.H.; Olivares, M.; et al. Evaluation of the effect of Lactobacillus reuteri V3401 on biomarkers of inflammation, cardiovascular risk and liver steatosis in obese adults with metabolic syndrome: A randomized clinical trial (PROSIR). BMC Complement. Altern Med. 2018, 18, 306. [Google Scholar] [CrossRef]
- Gamallat, Y.; Ren, X.; Meyiah, A.; Li, M.; Ren, X.; Jamalat, Y.; Song, S.; Xie, L.; Ahmad, B.; Shopit, A.; et al. The immune-modulation and gut microbiome structure modification associated with long-term dietary supplementation of Lactobacillus rhamnosus using 16S rRNA sequencing analysis. J. Funct. Foods 2019, 53, 227–236. [Google Scholar] [CrossRef]
- Hong, M.; Kim, S.W.; Han, S.H.; Kim, D.J.; Suk, K.T.; Kim, Y.S.; Kim, M.J.; Kim, M.Y.; Baik, S.K.; Ham, Y.L. Probiotics (Lactobacillus rhamnosus R0011 and acidophilus R0052) Reduce the Expression of Toll-Like Receptor 4 in Mice with Alcoholic Liver Disease. PLoS ONE 2015, 10, e0117451. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Gabbard, S.L.; Lacy, B.E.; Levine, G.M.; Crowell, M.D. The impact of alcohol consumption and cholecystectomy on small intestinal bacterial overgrowth. Dig. Dis. Sci. 2014, 59, 638–644. [Google Scholar] [CrossRef]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Lowe, P.P.; Gyongyosi, B.; Satishchandran, A.; Iracheta-Vellve, A.; Cho, Y.; Ambade, A.; Szabo, G. Reduced gut microbiome protects from alcohol-induced neuroinflammation and alters intestinal and brain inflammasome expression. J. Neuroinflammation 2018, 15, 298. [Google Scholar] [CrossRef]
- Chen, R.-C.; Xu, L.-M.; Du, S.-J.; Huang, S.-S.; Wu, H.; Dong, J.-J.; Huang, J.-R.; Wang, X.-D.; Feng, W.-K.; Chen, Y.-P. Lactobacillus rhamnosus GG supernatant promotes intestinal barrier function, balances Treg and TH17 cells and ameliorates hepatic injury in a mouse model of chronic-binge alcohol feeding. Toxicol. Lett. 2016, 241, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Kirpich, I.; Ma, Z.; Wang, C.; Zhang, M.; Suttles, J.; McClain, C.; Feng, W. Lactobacillus rhamnosus GG reduces hepatic TNFα production and inflammation in chronic alcohol-induced liver injury. J. Nutr. Biochem. 2013, 24, 1609–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, M.L.Y.; Turner, P.C.; Allen, K.J.; El-Nezami, H. Lactobacillus rhamnosus GG modulates intestinal mucosal barrier and inflammation in mice following combined dietary exposure to deoxynivalenol and zearalenone. J. Funct. Foods 2016, 22, 34–43. [Google Scholar] [CrossRef]
- Han, S.H.; Suk, K.T.; Kim, D.J.; Kim, M.Y.; Baik, S.K.; Kim, Y.D.; Cheon, G.J.; Choi, D.H.; Ham, Y.L.; Shin, D.H.; et al. Effects of probiotics (cultured Lactobacillus subtilis/Streptococcus faecium) in the treatment of alcoholic hepatitis: Randomized-controlled multicenter study. Eur. J. Gastroenterol. Hepatol. 2015, 27, 1300–1306. [Google Scholar] [CrossRef]
- Pessione, F.; Ramond, M.J.; Peters, L.; Pham, B.N.; Batel, P.; Rueff, B.; Valla, D.C. Five-year survival predictive factors in patients with excessive alcohol intake and cirrhosis. Effect of alcoholic hepatitis, smoking and abstinence. Liver Int. Off. J. Int. Assoc. Study Liver 2003, 23, 45–53. [Google Scholar] [CrossRef]
- Borowsky, S.A.; Strome, S.; Lott, E. Continued heavy drinking and survival in alcoholic cirrhotics. Gastroenterology 1981, 80, 1405–1409. [Google Scholar] [CrossRef]
- Lata, J.; Novotny, I.; Pribramska, V.; Jurankova, J.; Fric, P.; Kroupa, R.; Stiburek, O. The effect of probiotics on gut flora, level of endotoxin and Child-Pugh score in cirrhotic patients: Results of a double-blind randomized study. Eur. J. Gastroenterol. Hepatol. 2007, 19, 1111–1113. [Google Scholar] [CrossRef]
- Wick, E.C.; Sears, C.L. Bacteroides spp. and diarrhea. Curr. Opin. Infect. Dis. 2010, 23, 470–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, Z.; Hu, S.; Zhang, J.; Wu, J.; Shao, N.; Bo, X.; Ni, M.; Ying, X. Gut metagenomes of type 2 diabetic patients have characteristic single-nucleotide polymorphism distribution in Bacteroides coprocola. Microbiome 2017, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubinkina, V.B.; Tyakht, A.V.; Odintsova, V.Y.; Yarygin, K.S.; Kovarsky, B.A.; Pavlenko, A.V.; Ischenko, D.S.; Popenko, A.S.; Alexeev, D.G.; Taraskina, A.Y.; et al. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease. Microbiome 2017, 5, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrario, C.; Taverniti, V.; Milani, C.; Fiore, W.; Laureati, M.; De Noni, I.; Stuknyte, M.; Chouaia, B.; Riso, P.; Guglielmetti, S. Modulation of fecal Clostridiales bacteria and butyrate by probiotic intervention with Lactobacillus paracasei DG varies among healthy adults. J. Nutr. 2014, 144, 1787–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Liu, M.C.; Yang, J.; Wang, J.F.; Zhu, Y.H. Lactobacillus rhamnosus GR-1 Ameliorates Escherichia coli-Induced Inflammation and Cell Damage via Attenuation of ASC-Independent NLRP3 Inflammasome Activation. Appl. Environ. Microbiol. 2016, 82, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Medellin-Peña, M.J.; Griffiths, M.W. Effect of Molecules Secreted by Lactobacillus acidophilus Strain La-5 on Escherichia coli O157:H7 Colonization. Appl. Environ. Microbiol. 2009, 75, 1165–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Puri, P.; Liangpunsakul, S.; Christensen, J.E.; Shah, V.H.; Kamath, P.S.; Gores, G.J.; Walker, S.; Comerford, M.; Katz, B.; Borst, A.; et al. The circulating microbiome signature and inferred functional metagenomics in alcoholic hepatitis. Hepatology 2018, 67, 1284–1302. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, F.; Lu, H.; Wang, B.; Chen, Y.; Lei, D.; Wang, Y.; Zhu, B.; Li, L. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 2011, 54, 562–572. [Google Scholar] [CrossRef]
- Korpela, K.; Salonen, A.; Virta, L.J.; Kumpu, M.; Kekkonen, R.A.; de Vos, W.M. Lactobacillus rhamnosus GG Intake Modifies Preschool Children’s Intestinal Microbiota, Alleviates Penicillin-Associated Changes, and Reduces Antibiotic Use. PLoS ONE 2016, 11, e0154012. [Google Scholar] [CrossRef]
- Kwak, D.S.; Jun, D.W.; Seo, J.G.; Chung, W.S.; Park, S.E.; Lee, K.N.; Khalid-Saeed, W.; Lee, H.L.; Lee, O.Y.; Yoon, B.C.; et al. Short-term probiotic therapy alleviates small intestinal bacterial overgrowth, but does not improve intestinal permeability in chronic liver disease. Eur. J. Gastroenterol. Hepatol. 2014, 26, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Hylemon, P.B.; Ridlon, J.M.; Heuman, D.M.; Daita, K.; White, M.B.; Monteith, P.; Noble, N.A.; Sikaroodi, M.; Gillevet, P.M. Colonic mucosal microbiome differs from stool microbiome in cirrhosis and hepatic encephalopathy and is linked to cognition and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G675–G685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fang, Z.; Zhang, C.; Xia, H.; Jie, Z.; Han, X.; Chen, Y.; Ji, L. Effects of Acarbose on the Gut Microbiota of Prediabetic Patients: A Randomized, Double-blind, Controlled Crossover Trial. Diabetes Ther. 2017, 8, 293–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Values |
|---|---|
| Male (n (%)) | 83 (93) |
| Age (years) | 50.8 ± 9.4 |
| BMI (kg/m2) | 22.9 ± 3.7 |
| Presence of LC (n (%)) | 62 (52) |
| Total protein (g/dL) | 6.7 ± 0.8 |
| Albumin (g/dL) | 3.8 ± 0.7 |
| AST (IU/l) | 159.2 ± 250.7 |
| ALT (IU/l) | 101.6 ± 176.4 |
| ALP (IU/l) | 118.0 ± 53.7 |
| γGT (IU/l) | 436.8 ± 466.0 |
| TB (mg/dL) | 2.3 ± 3.3 |
| Cholesterol (mg/dL) | 158.8 ± 46.1 |
| PT (s) | 12.8 ± 4.2 |
| CP score in patients with LC | 7.8 ± 2.5 |
| Variable (Mean ± SD) | Placebo (n = 45) | Probiotic (n = 45) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre | Post | p Value | Pre | Post | p Value | |||||
| AST (IU/L) | 126.2 | ±144.3 | 70.4 | ±73.3 * | 0.010 | 192.3 | ±322.3 | 64.0 | ±48.5 * | 0.008 |
| ALT (IU/L) | 91.3 | ±160.3 | 40.2 | ±40.5 * | 0.010 | 111.9 | ±192.2 | 39.9 | ±37.2 * | 0.004 |
| γ-GT (IU/L) | 434.7 | ±379.8 | 332.9 | ±314.5 * | 0.002 | 439.0 | ±542.7 | 323.4 | ±399.7 * | 0.008 |
| Na (mEq/L) | 139.0 | ±3.1 | 139.5 | ±4.2 | 0.480 | 138.3 | ±3.3 | 133.7 | ±29.9 | 0.330 |
| TB (mg/dL) | 2.5 | ±3.9 | 1.6 | ±3.0 * | <0.001 | 2.1 | ±2.6 | 1.4 | ±2.0 * | <0.001 |
| Chol (mg/dL) | 159.2 | ±46.1 | 168.8 | ±47.7 | 0.115 | 158.3 | ±46.6 | 164.8 | ±40.6 | 0.207 |
| ALP (IU/L) | 121.3 | ±55.0 | 113.4 | ±53.6 | 0.101 | 114.8 | ±52.8 | 104.0 | ±43.4 * | 0.011 |
| Albumin (g/dL) | 3.8 | ±0.7 | 3.8 | ±0.6 | 0.615 | 3.7 | ±0.7 | 3.8 | ±0.6 | 0.258 |
| TP (g/dL) | 6.7 | ±0.8 | 6.7 | ±0.7 | 0.939 | 6.7 | ±0.8 | 6.8 | ±0.7 | 0.229 |
| Glucose (mg/dL) | 147.5 | ±79.1 | 136.2 | ±63.3 | 0.272 | 145.6 | ±69.4 | 144.8 | ±71.8 | 0.933 |
| INR | 1.2 | ±0.3 | 1.2 | ±0.3 | 0.737 | 1.2 | ±0.4 | 1.1 | ±0.4 | 0.146 |
| PT (s) | 12.8 | ±4.0 | 12.5 | ±3.3 | 0.447 | 12.9 | ±4.5 | 12.4 | ±3.9 | 0.239 |
| Child-Pugh score a | 7.7 | ±2.5 | 7.2 | ±2.0 | 0.053 | 7.9 | ±2.5 | 7.2 | ±2.1 * | <0.001 |
| Taxon Name | Taxon Rank | LDA | p-Value | Normal Control | Cirrhosis Control | Placebo-Post | Probiotic-Post |
|---|---|---|---|---|---|---|---|
| Clostridium_g21 | Genus | 3.520 | 0.023 | 0.004 | 2.568 | 6.983 | 0.666 |
| Ruminococcus gnavus | Species | 3.505 | 0.029 | 0.003 | 2.517 | 6.817 | 0.644 |
| Eubacterium_g7 | Genus | 2.036 | 0.037 | 0.558 | 0.022 | 0.000 | 0.080 |
| FM873843_s | Species | 2.010 | 0.037 | 0.520 | 0.020 | 0.000 | 0.079 |
| Prevotellaceae_uc | Genus | 2.895 | 0.044 | 1.187 | 0.211 | 0.004 | 0.367 |
| Prevotella copri | Species | 2.010 | 0.045 | 0.167 | 0.021 | 0.000 | 0.054 |
| Taxon Name | p-Value | Normal Control | Probiotic | Placebo | Change | |
|---|---|---|---|---|---|---|
| Placebo | Probiotic | |||||
| Prevotella | 0.009 | 1.187 | 0.367 | 0.004 | ↓ | ↑ |
| Oscillibacter | 0.016 | 1.490 | 2.713 | 0.042 | ↓ | ↑ |
| Prevotellaceae_uc | 0.009 | 1.186 | 0.432 | 0.011 | ↓ | ↑ |
| Alistipes | 0.010 | 0.952 | 1.244 | 0.008 | ↓ | ↑ |
| Eubacterium_g7 | 0.014 | 0.558 | 0.066 | 0.000 | ↓ | ↑ |
| Sporobacter | 0.035 | 0.409 | 0.852 | 0.010 | ↓ | ↑ |
| Subdoligranulum | 0.045 | 0.330 | 0.273 | 0.003 | ↓ | ↑ |
| Bacteroidaceae_uc | 0.049 | 0.316 | 0.258 | 0.105 | ↓ | ↑ |
| Parabacteroides | 0.047 | 0.240 | 1.265 | 0.333 | ↓ | ↑ |
| Ruminococcaceae_uc | 0.018 | 0.146 | 0.254 | 0.055 | ↓ | ↑ |
| Porphyromonadaceae_uc | 0.007 | 0.028 | 0.079 | 0.007 | ↓ | ↑ |
| Eisenbergiella | 0.024 | 0.200 | 0.181 | 0.020 | ↓ | ↑ |
| Clostridium_g21 | 0.010 | 0.004 | 0.655 | 5.416 | ↑ | ↓ |
| Lactobacillus | 0.023 | 0.072 | 0.039 | 4.644 | ↑ | ↓ |
| Fusobacterium | 0.035 | 0.001 | 2.039 | 4.508 | ↑ | ↓ |
| Clostridium | 0.023 | 0.021 | 0.051 | 10.535 | ↑ | ↓ |
| Clostridiaceae_uc | 0.035 | 0.001 | 0.008 | 0.062 | ↑ | ↓ |
| Enterococcaceae_uc | 0.048 | 0.000 | 0.005 | 0.006 | ↑ | ↓ |
| Eubacterium_g20 | 0.043 | 0.076 | 0.004 | 0.005 | ↑ | ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, H.; Kim, S.H.; Kim, S.K.; Han, S.H.; Kwon, H.C.; Suk, K.T. Beneficial Shifts in Gut Microbiota by Lacticaseibacillus rhamnosus R0011 and Lactobacillus helveticus R0052 in Alcoholic Hepatitis. Microorganisms 2022, 10, 1474. https://doi.org/10.3390/microorganisms10071474
Gupta H, Kim SH, Kim SK, Han SH, Kwon HC, Suk KT. Beneficial Shifts in Gut Microbiota by Lacticaseibacillus rhamnosus R0011 and Lactobacillus helveticus R0052 in Alcoholic Hepatitis. Microorganisms. 2022; 10(7):1474. https://doi.org/10.3390/microorganisms10071474
Chicago/Turabian StyleGupta, Haripriya, Sung Hun Kim, Seul Ki Kim, Sang Hak Han, Hak Cheol Kwon, and Ki Tae Suk. 2022. "Beneficial Shifts in Gut Microbiota by Lacticaseibacillus rhamnosus R0011 and Lactobacillus helveticus R0052 in Alcoholic Hepatitis" Microorganisms 10, no. 7: 1474. https://doi.org/10.3390/microorganisms10071474
APA StyleGupta, H., Kim, S. H., Kim, S. K., Han, S. H., Kwon, H. C., & Suk, K. T. (2022). Beneficial Shifts in Gut Microbiota by Lacticaseibacillus rhamnosus R0011 and Lactobacillus helveticus R0052 in Alcoholic Hepatitis. Microorganisms, 10(7), 1474. https://doi.org/10.3390/microorganisms10071474