
ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset Compilation
2.2. New ITS Barcodes
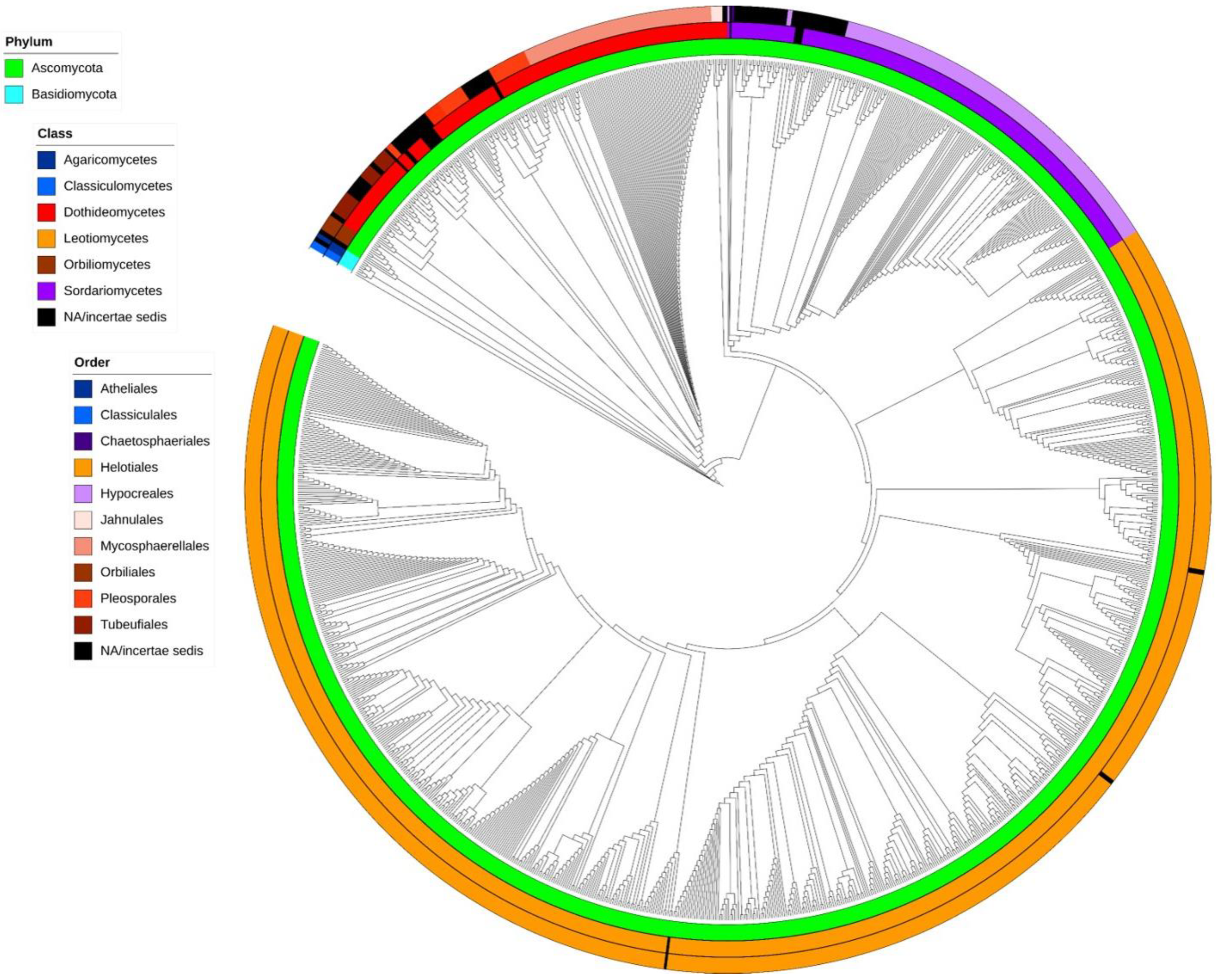
2.3. Phylogenetic Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pascoal, C.; Cássio, F. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river. Appl. Environ. Microbiol. 2004, 70, 5266–5273. [Google Scholar] [CrossRef] [Green Version]
- Gessner, M.; Gulis, V.; Kuehn, K.; Chauvet, E.K.S. Fungal decomposers of plant litter in aquatic ecosystems. In The Mycota: Environmental and Microbial Relationships; Kubicek, C., Druzhinina, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 301–321. [Google Scholar]
- Graça, M.A.S.; Canhoto, C. Leaf litter processing in low order streams. Limnetica 2006, 25, 1–10. [Google Scholar] [CrossRef]
- Descals, E. Ingoldian Fungi: Some field and laboratory techniques. Bolletí Soc. Història Nat. Balear. 1997, 40, 169–222. [Google Scholar]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Duarte, S.; Bärlocher, F.; Pascoal, C.; Cássio, F. Biogeography of aquatic hyphomycetes: Current knowledge and future perspectives. Fungal Ecol. 2016, 19, 169–181. [Google Scholar] [CrossRef]
- Chauvet, E. Aquatic Hyphomycete Distribution in South-Western France. J. Biogeogr. 1991, 18, 699. [Google Scholar] [CrossRef]
- Jabiol, J.; Bruder, A.; Gessner, M.O.; Makkonen, M.; McKie, B.G.; Peeters, E.T.H.M.; Vos, V.C.A.; Chauvet, E. Diversity patterns of leaf-associated aquatic hyphomycetes along a broad latitudinal gradient. Fungal Ecol. 2013, 6, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Seena, S.; Bärlocher, F.; Sobral, O.; Gessner, M.O.; Dudgeon, D.; McKie, B.G.; Chauvet, E.; Boyero, L.; Ferreira, V.; Frainer, A.; et al. Biodiversity of leaf litter fungi in streams along a latitudinal gradient. Sci. Total Environ. 2019, 661, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Cássio, F.; Pascoal, C. Environmental drivers are more important for structuring fungal decomposer communities than the geographic distance between streams. Limnetica 2017, 36, 491–506. [Google Scholar] [CrossRef]
- Ferreira, V.; Gulis, V.; Pascoal, C.; Graça, M.A.S. Stream pollution and fungi. In Freshwater Fungi and Fungal-Like Organisms; Jones, E.B.G., Hyde, K.D., Pang, K.L., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 389–412. [Google Scholar]
- Gulis, V.; Su, R.; Kuehn, K.A. Fungal decomposers in freshwater environments. In Advances in Environmental Microbiology. Vol. 7: The Structure and Function of Aquatic Microbial Communities; Hurst, C.J., Ed.; Springer: Cham, Switzerland, 2019; pp. 121–155. [Google Scholar]
- Barlocher, F. The Ecology of Aquatic Hyphomycetes; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Belliveau, M.J.R.; Bärlocher, F. Molecular evidence confirms multiple origins of aquatic hyphomycetes. Mycol. Res. 2005, 109, 1407–1417. [Google Scholar] [CrossRef]
- Baschien; Marvanová, L.; Szewzyk, U. Phylogeny of selected aquatic hyphomycetes based on morphological and molecular data. Nov. Hedwigia 2006, 83, 311–352. [Google Scholar] [CrossRef]
- Campbell, J.; Shearer, C.; Marvanová, L. Evolutionary relationships among aquatic anamorphs and teleomorphs: Lemonniera, Margaritispora, and Goniopila. Mycol. Res. 2006, 110, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.; Marvanová, L.; Gulis, V. Evolutionary relationships between aquatic anamorphs and teleomorphs: Tricladium and Varicosporium. Mycol. Res. 2009, 113, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Baschien, C.; Tsui, C.K.M.; Gulis, V.; Szewzyk, U.; Marvanová, L. The molecular phylogeny of aquatic hyphomycetes with affinity to the Leotiomycetes. Fungal Biol. 2013, 117, 660–672. [Google Scholar] [CrossRef]
- Gulis, V.; Marvanová, L.; Descals, E. An illustrated key to the common temperate species of aquatic hyphomycetes. In Methods to Study Litter Decomposition: A Practical Guide; Graça, M.A.S., Bärlocher, F., Gessner, M.O., Eds.; Springer: Cham, Switzerland, 2020; pp. 223–239. [Google Scholar]
- Alexopoulos, C.; Mims, C.; Blackwell, M. Introductory Mycology, 4th ed.; John Wiley and Sons: New York, NY, USA, 1996; ISBN 0-471-52229-5. [Google Scholar]
- Marvanová, L. Freshwater hyphomycetes: A survey with remarks on tropical taxa. In Tropical Mycology; Janardhanan, K., Rajendran, C., Natarajan, K., Hawksworth, D., Eds.; Science Publisher Inc.: Enfield, UK, 1997; pp. 169–226. [Google Scholar]
- Johnston, P.R.; Quijada, L.; Smith, C.A.; Baral, H.O.; Hosoya, T.; Baschien, C.; Pärtel, K.; Zhuang, W.Y.; Haelewaters, D.; Park, D.; et al. A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus 2019, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bruns, T.D.; White, T.J.; Taylor, J.W. Fungal molecular systematics. Annu. Rev. Ecol. Syst. 1991, 22, 525–564. [Google Scholar] [CrossRef]
- Berbee, M.; Taylor, J. Fungal molecular evolution: Gene trees and geologic time. In Systematics and Evolution; Springer: Berlin/Heidelberg, Germany, 2001; pp. 229–245. [Google Scholar]
- Taylor, J. A contemporary view of the holomorph: Nucleic acid sequence and computer databases are changing fungal classification. In The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, D., Taylor, J., Eds.; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Sati, S.C.; Pathak, R. Anamorph (asexual stage) Teleomorph (sexual stage) Connections in Aquatic hyphomycetes. Int. J. Plant Reprod. Biol. 2016, 8, 65–74. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Bärlocher, F.; Trabulo, J.; Cássio, F.; Pascoal, C. Stream-dwelling fungal decomposer communities along a gradient of eutrophication unraveled by 454 pyrosequencing. Fungal Divers. 2014, 70, 127–148. [Google Scholar] [CrossRef]
- Fernandes, I.; Pereira, A.; Trabulo, J.; Pascoal, C.; Cássio, F.; Duarte, S. Microscopy- or DNA-based analyses: Which methodology gives a truer picture of stream-dwelling decomposer fungal diversity? Fungal Ecol. 2015, 18, 130–134. [Google Scholar] [CrossRef]
- Johnston, P.R.; Baschien, C. Tricladiaceae fam. nov. (Helotiales, Leotiomycetes). Fungal Syst. Evol. 2020, 6, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Duarte, S.; Barlocher, F.; Cássio, F.; Pascoal, C. Current status of DNA barcoding of aquatic hyphomycetes. Sydowia 2014, 66, 191–202. [Google Scholar]
- Duarte, S.; Batista, D.; Bärlocher, F.; Cássio, F.; Pascoal, C. Some new DNA barcodes of aquatic hyphomycete species. Mycoscience 2015, 56, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Seena, S.; Marvanová, L.; Letourneau, A.; Bärlocher, F. Articulospora—Phylogeny vs morphology. Fungal Biol. 2018, 122, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, C.; Marvanová, L.; Cássio, F. Aquatic hyphomycete diversity in streams of Northwest Portugal. Fungal Divers. 2005, 19, 109–128. [Google Scholar]
- Descals, E. Techniques for handling Ingoldian fungi. In Methods to Study Litter Decomposition—A Practical Guide; Springer: Berlin/Heidelberg, Germany, 2005; pp. 129–141. [Google Scholar] [CrossRef]
- Duarte, S.; Seena, S.; Bärlocher, F.; Cássio, F.; Pascoal, C. Preliminary Insights into the Phylogeography of Six Aquatic Hyphomycete Species. PLoS ONE 2012, 7, e45289. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Inorg. Chem. Front. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Seena, S.; Pascoal, C.; Marvanová, L.; Cássio, F. DNA barcoding of fungi: A case study using ITS sequences for identifying aquatic hyphomycete species. Fungal Divers 2010, 44, 77–87. [Google Scholar] [CrossRef]
- Letourneau, A.; Seena, S.; Marvanová, L.; Bärlocher, F. Potential use of barcoding to identify aquatic hyphomycetes. Fungal Divers 2010, 40, 51–64. [Google Scholar] [CrossRef]
- Barlocher, F. Molecular approaches promise a deeper and broader understanding of the evolutionary ecology of aquatic hyphomycetes. J. N. Am. Benthol. Soc. 2010, 29, 1027–1041. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Nikolcheva, L.G.; Bärlocher, F. Phylogeny of Tetracladium based on 18S rDNA. Czech Mycol. 2002, 53, 285–295. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, X.; Wu, W.; Hao, Y.; Su, Y.; Cai, L.; Xiang, M.; Liu, X. Psychrophilic fungi from the world’s roof. Persoonia Mol. Phylogeny Evol. Fungi 2015, 34, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Pratibha, J.; Bhat, D.J.; Prabhugaonkar, A. Molecular phylogeny of Speiropsis pedatospora. Mycosphere 2016, 7, 679–686. [Google Scholar] [CrossRef]
- Prihatini, R.; Boonyuen, N.; Sivichai, S. Phylogenetic Evidence that Two Submerged-Habitat Fungal Species, Speiropsis pedatospora and Xylomyces chlamydosporus, Belong to the Order Jahnulales Insertae Sedis Dothideomycetes. Microbiol. Indones. 2008, 2, 136–140. [Google Scholar] [CrossRef]
- Suetrong, S.; Boonyuen, N.; Pang, K.L.; Ueapattanakit, J.; Klaysuban, A.; Sri-Indrasutdhi, V.; Sivichai, S.; Jones, E.B.G. A taxonomic revision and phylogenetic reconstruction of the Jahnulales (Dothideomycetes), and the new family Manglicolaceae. Fungal Divers 2011, 51, 163–188. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes; Springer: Dordrecht, The Netherlands, 2020; Volume 105, ISBN 1322502000463. [Google Scholar]
- Suetrong, S.; Rungjindamai, N.; Sommai, S.; Rung-Areerate, P.; Sommrithipol, S.; Gareth Jones, E.B. Wiesneriomyces a new lineage of Dothideomycetes (Ascomycota) basal to Tubeufiales. Phytotaxa 2014, 176, 283–297. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. Barcoding: Bold: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekpinar, A.D.; Kalmer, A. Utility of various molecular markers in fungal identification and phylogeny. Nov. Hedwigia 2019, 109, 187–224. [Google Scholar] [CrossRef]
- Harrower, E.; Ammirati, J.F.; Cappuccino, A.A.; Ceska, O.; Kranabetter, J.M.; Kroeger, P.; Lim, S.R.; Taylor, T.; Berbee, M.L. Cortinarius species diversity in British Columbia and molecular phylogenetic comparison with European specimen sequences. Botany 2011, 89, 799–810. [Google Scholar] [CrossRef]
- Ge, Z.W.; Jacobs, A.; Vellinga, E.C.; Sysouphanthong, P.; van der Walt, R.; Lavorato, C.; An, Y.F.; Yang, Z.L. A multi-gene phylogeny of Chlorophyllum (Agaricaceae, Basidiomycota): New species, new combination and infrageneric classification. MycoKeys 2018, 32, 65–90. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-K.; Hyde, K.D.; Bhat, D.J.; Wen, T.-C. Novel Taxa within Nectriaceae: Cosmosporella gen. nov. and Aquanectria sp. nov. from Freshwater Habitats in China. Cryptogam. Mycol. 2018, 39, 169–192. [Google Scholar] [CrossRef]
- Gordillo, A.; Decock, C. Multigene phylogenetic and morphological evidence for seven new species of Aquanectria and Gliocladiopsis (Ascomycota, Hypocreales) from tropical areas. Mycologia 2019, 111, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Descals, E.; Marvanová, L.; Webster, J. New taxa and combinations of aquatic hyphomycetes. Can. J. Bot. 1998, 76, 1647–1659. [Google Scholar] [CrossRef]
- Wirtz, N.; Printzen, C.; Lumbsch, H.T. Using haplotype networks, estimation of gene flow and phenotypic characters to understand species delimitation in fungi of a predominantly Antarctic Usnea group (Ascomycota, Parmeliaceae). Org. Divers. Evol. 2012, 12, 17–37. [Google Scholar] [CrossRef]
- Chiva, S.; Garrido-Benavent, I.; Moya, P.; Molins, A.; Barreno, E. How did terricolous fungi originate in the Mediterranean region? A case study with a gypsicolous lichenized species. J. Biogeogr. 2019, 46, 515–525. [Google Scholar] [CrossRef]
- Li, Y.; Steenwyk, J.L.; Chang, Y.; Wang, Y.; James, T.Y.; Stajich, J.E.; Spatafora, J.W.; Groenewald, M.; Dunn, C.W.; Hittinger, C.T.; et al. A genome-scale phylogeny of the kingdom Fungi. Curr. Biol. 2021, 31, 1653–1665. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Seena, S. Fungi in Freshwaters: Prioritising Aquatic Hyphomycetes in Conservation Goals. Water 2022, 14, 605. [Google Scholar] [CrossRef]


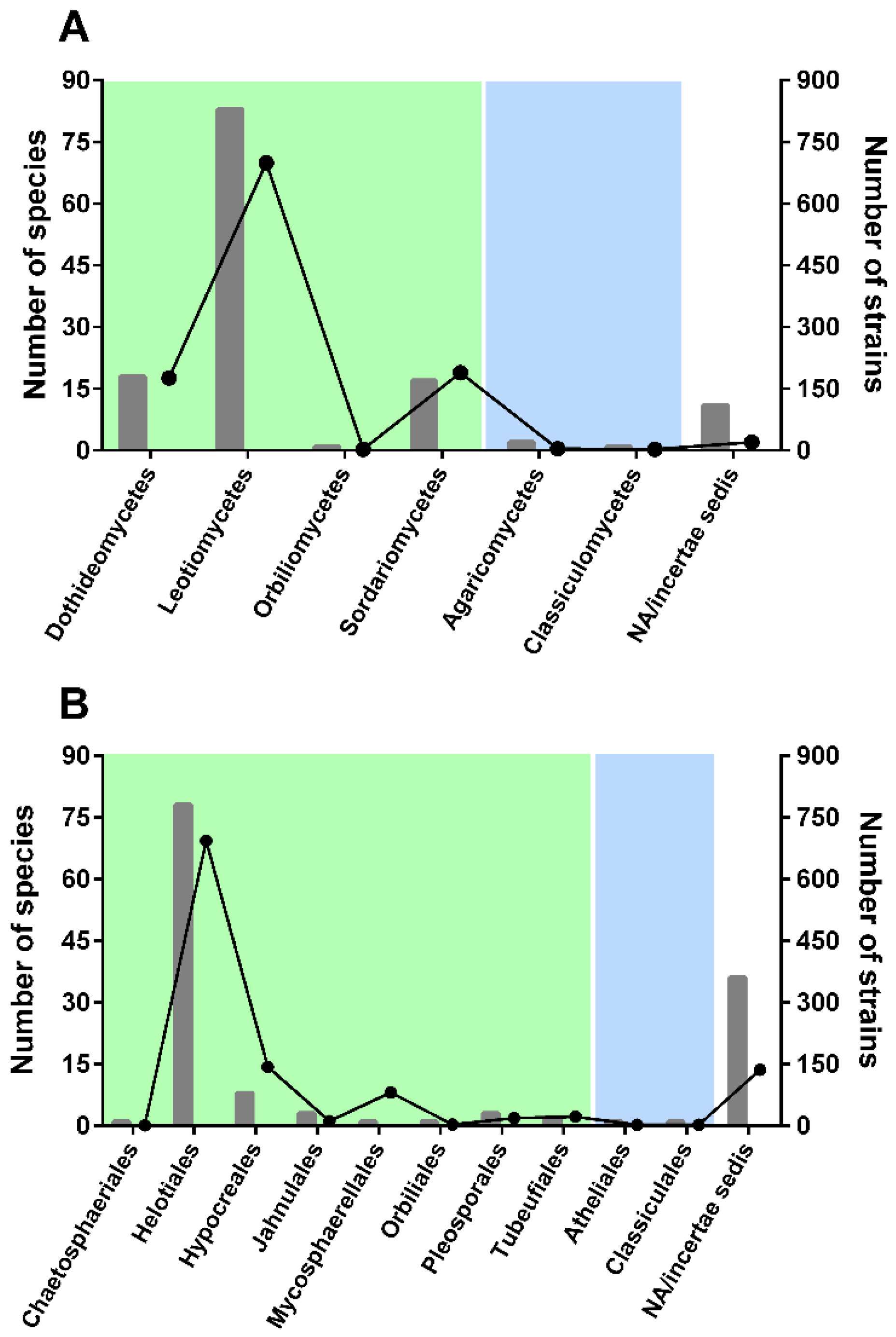
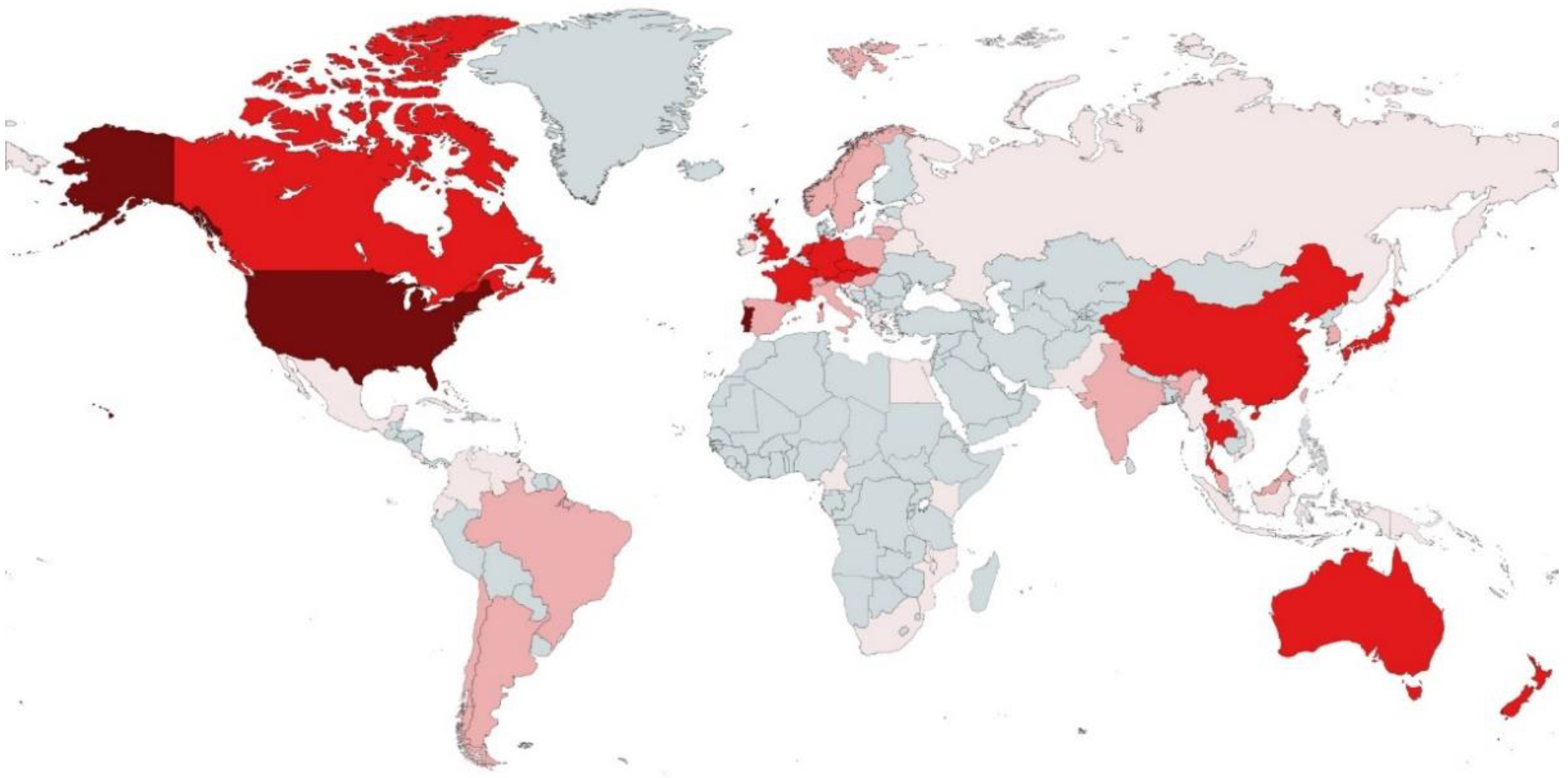
 —more than 30;
—more than 30;  —from 11 to 29;
—from 11 to 29;  —from 3 to 10;
—from 3 to 10;  —less than 3 species;
—less than 3 species;  —zero species.
—more than 30; —from 11 to 29; —from 3 to 10; —less than 3 species; —zero species.
—zero species.
—more than 30; —from 11 to 29; —from 3 to 10; —less than 3 species; —zero species.

{kind=link}
{kind=link}
{kind=link}
| Species Name | Strain | Isolation Country | Isolation Substrate | GenBank Accession Number |
|---|---|---|---|---|
| Alatospora acuminata Ingold | UMB-223 | Portugal | foam | OM273714 |
| Alatospora acuminata Ingold | UMB-741 | Portugal | leaves | MZ773535 |
| Alatospora acuminata Ingold | UMB-902 | Portugal | oak leaves | OM273715 |
| Alatospora pulchella Marvanová | UMB-1115 | Portugal | oak leaves | MZ773536 |
| Anguillospora crassa Ingold | UMB-217 | Portugal | foam | OM273716 |
| Anguillospora crassa Ingold | UMB-1150 | Portugal | foam | MZ773539 |
| Anguillospora crassa Ingold | VG33-1 | USA | dead submerged tree roots | OM907724 |
| Anguillospora curvula S.H. Iqbal | VG69-4 | USA | grass blades | OM907725 |
| Anguillospora filiformis Greath. | UMB-016 | Portugal | leaves | OM273717 |
| Anguillospora filiformis Greath. | UMB-225 | Portugal | leaves | MZ773533 |
| Aquanectria penicillioides (Ingold) L. Lombard and Crous | VG205-1-2 | USA | wood | OM907726 |
| Arbusculina irregularis (R.H. Petersen) Marvanová and Descals | CCM F-23687 | Canada | unknown | OM273718 |
| Arbusculina irregularis (R.H. Petersen) Marvanová and Descals | VG76-8 | USA | foam | OM906795 |
| Articulospora atra Descals | VG233-6 | USA | wood | OM907727 |
| Articulospora proliferata A. Roldán and W.J.J. van der Merwe | VG229-6 | USA | grasses | OM907728 |
| Articulospora tetracladia Ingold | UMB-712 | Portugal | foam | OK605572 |
| Articulospora tetracladia Ingold | UMB-1144 | Portugal | foam | OK605573 |
| Casaresia sphagnorum Gonz. Frag. | VG7-1 | USA | Quercus prinus leaves | OM907729 |
| Clavariana aquatica Nawawi | VG75-4 | USA | foam | OM907730 |
| Clavatospora longibrachiata (Ingold) Sv. Nilsson ex Marvanová and Sv. Nilsson | VG80-6 | USA | Tilia sp. leaves | OM907731 |
| Culicidospora gravida R.H. Petersen | VG39-4 | USA | foam | OM907732 |
| Dendrosporomyces prolifer Nawawi, J. Webster and R.A. Davey | VG258-1 | USA | foam | OM906797 |
| Dendrosporomyces prolifer Nawawi, J. Webster and R.A. Davey | VG98-3 | USA | foam | OM906796 |
| Dimorphospora foliicola Tubaki | UMB-215 | Portugal | leaves | OM273719 |
| Dimorphospora foliicola Tubaki | UMB-1119 | Portugal | oak leaves | MZ773538 |
| Filosporella exilis Gulis and Marvanová | VG211-1 | USA | grasses | OM907733 |
| Filosporella fistucella Marvanová and P.J. Fisher | UMB-007 | Portugal | water | OM273720 |
| Fontanospora alternibrachiata Dyko | VG8-4 | USA | Rhododendron maximum leaves | OM907734 |
| Geniculospora inflata (Ingold) Sv. Nilsson ex Marvanová and Sv. Nilsson | VG79-1 | USA | twigs | OM907735 |
| Heliscella stellata (Ingold and V.J. Cox) Marvanová | VG254-5 | S. Korea | Betula sp. leaves | OM907736 |
| Heliscina antennata Marvanová | VG50-2 | USA | artificial foam | OM907737 |
| Hydrocina chaetocladia Scheuer | UMB-1116 | Portugal | oak leaves | MZ773531 |
| Isthmotricladia gombakiensis Nawawi | VG113-5 | USA | foam | OM907738 |
| Lateriramulosa uni-inflata Matsush. | VG80-7 | USA | unident. dicot leaves | OM907739 |
| Lemonniera alabamensis R.C. Sinclair and Morgan-Jones | UMB-594 | Portugal | leaves | MZ773530 |
| Lemonniera aquatica De Wild. | VG66-7 | USA | sedges | OM907740 |
| Lemonniera cornuta Ranzoni | VG77-4 | USA | foam | OM907741 |
| Lemonniera pseudofloscula Dyko | VG30-2 | USA | Acer rubrum leaves | OM907742 |
| Lemonniera terrestris Tubaki | VG209-3 | USA | leaves | OM907743 |
| Mycofalcella calcarata Marvanová, Khattab and J. Webster | VG44-4 | USA | decorticated branch | OM907744 |
| Neonectria lugdunensis (Sacc. and Therry) L. Lombard and Crous | UMB-161 | Portugal | Twigs | OK605576 |
| Pleuropedium multiseptatum Marvanová and Descals | CCM F-46594 | Canada | Unknown | OM273721 |
| Pyramidospora constricta N. Singh | VG116-5 | USA | Platanus sp. leaves | OM907745 |
| Pyramidospora ramificata Miura | VG54-1 | USA | unident. dicot leaves | OM907746 |
| Tricladium curvisporum Descals | VG69-3 | USA | Grasses | OM907747 |
| Tricladium curvisporum Descals | VG242-1 | USA | Grasses | OM907748 |
| Tricladium splendens Ingold | UMB-414 | Portugal | Foam | OK605580 |
| Tricladium splendens Ingold | UMB-1117 | Portugal | oak leaves | MZ773537 |
| Tumularia tuberculata (Gönczöl) Descals and Marvanová | VG262-4 | S. Korea | Quercus sp. leaves | OM907749 |
| Tumularia tuberculata (Gönczöl) Descals and Marvanová | VG264-4 | S. Korea | Quercus sp. leaves | OM907750 |
| Varicosporium elodeae W. Kegel | UMB-878 | Portugal | Foam | OK605582 |
| Variocladium giganteum (S.H. Iqbal) Descals and Marvanová | VG43-4 | USA | Quercus sp. leaves | OM907751 |
| Variocladium rangiferinum (Descals) Descals and Marvanová | VG71-1 | USA | Sedges | OM907752 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franco-Duarte, R.; Fernandes, I.; Gulis, V.; Cássio, F.; Pascoal, C. ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes. Microorganisms 2022, 10, 1569. https://doi.org/10.3390/microorganisms10081569
Franco-Duarte R, Fernandes I, Gulis V, Cássio F, Pascoal C. ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes. Microorganisms. 2022; 10(8):1569. https://doi.org/10.3390/microorganisms10081569
Chicago/Turabian StyleFranco-Duarte, Ricardo, Isabel Fernandes, Vladislav Gulis, Fernanda Cássio, and Cláudia Pascoal. 2022. "ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes" Microorganisms 10, no. 8: 1569. https://doi.org/10.3390/microorganisms10081569
APA StyleFranco-Duarte, R., Fernandes, I., Gulis, V., Cássio, F., & Pascoal, C. (2022). ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes. Microorganisms, 10(8), 1569. https://doi.org/10.3390/microorganisms10081569