
Progress of Molecular Display Technology Using Saccharomyces cerevisiae to Achieve Sustainable Development Goals
Abstract
:1. Introduction
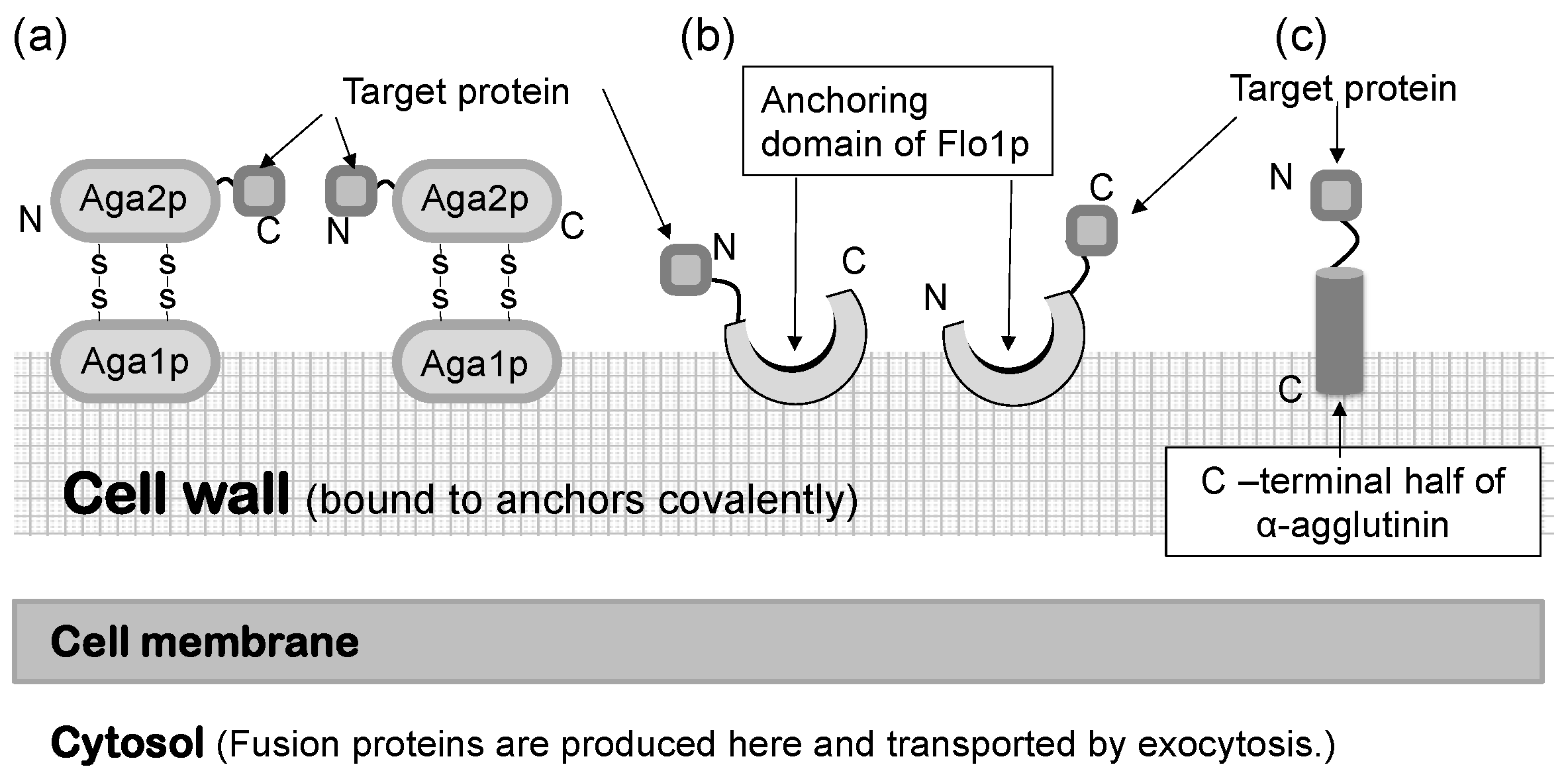
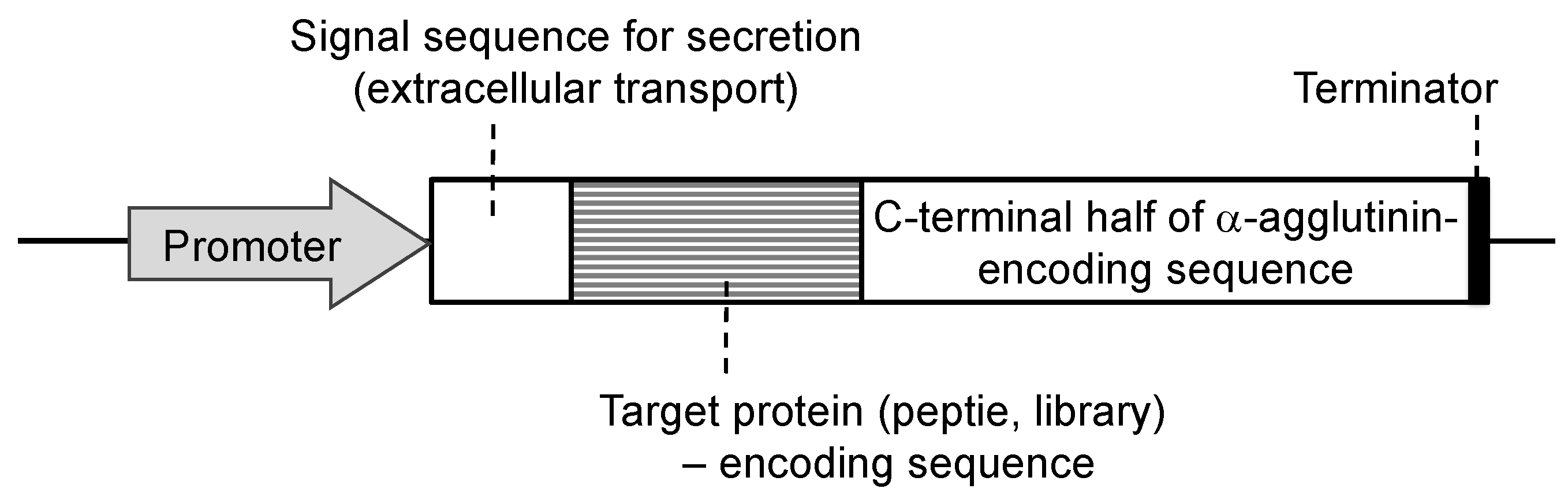
2. General Description of Molecular Display Technology
3. Principle of Molecular Display Technology Using Yeasts


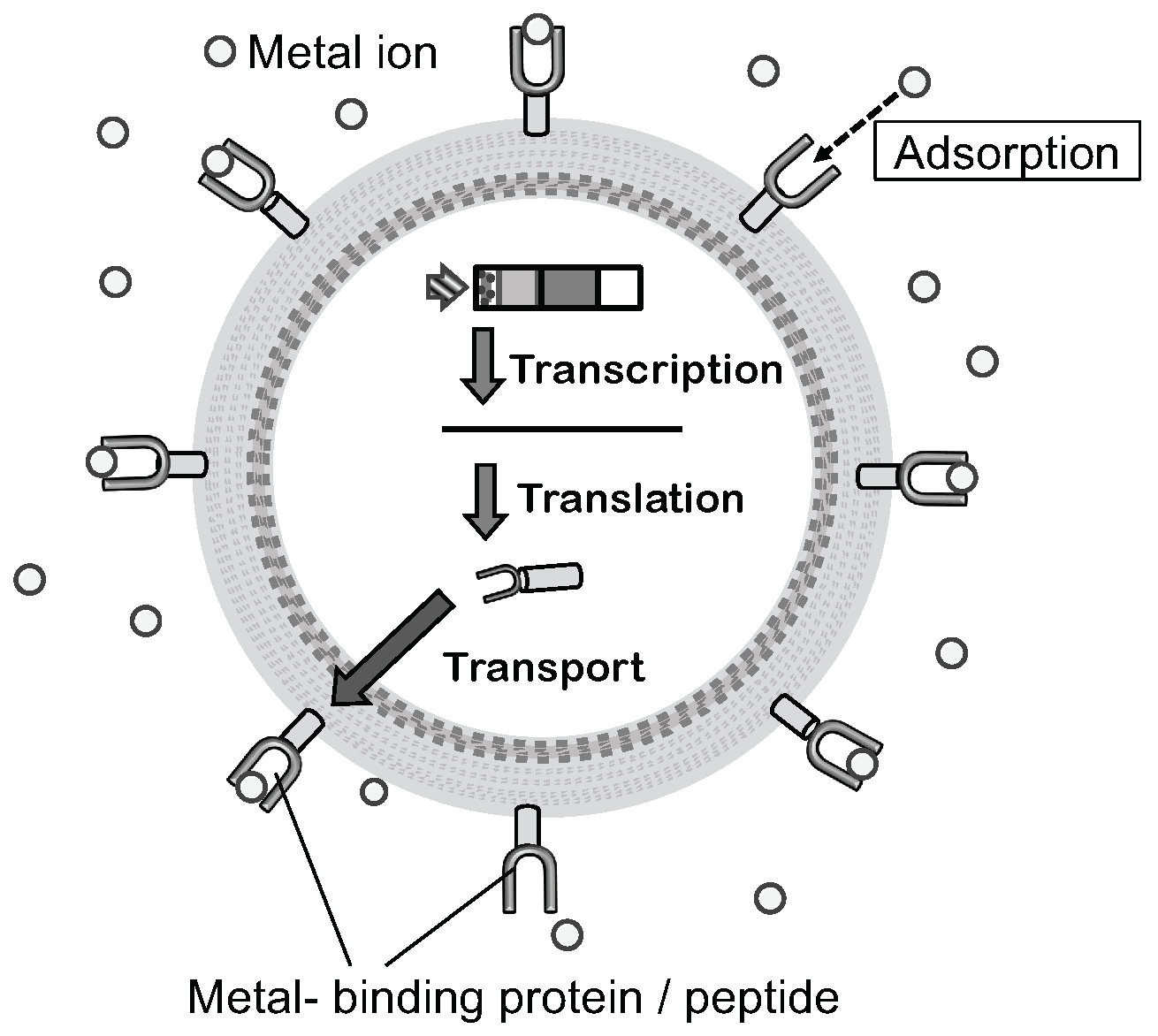
4. Applications in Environmental Processes
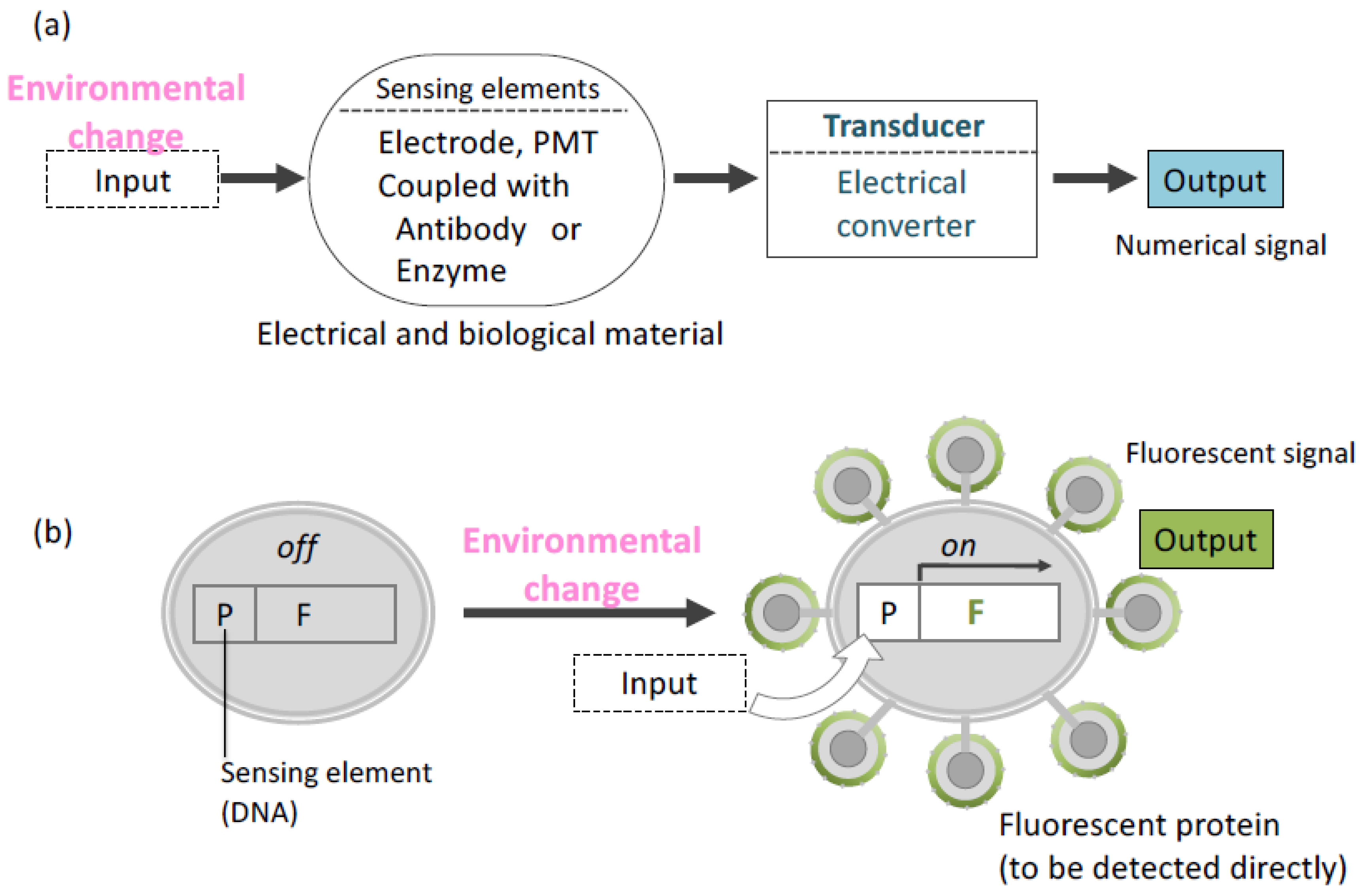
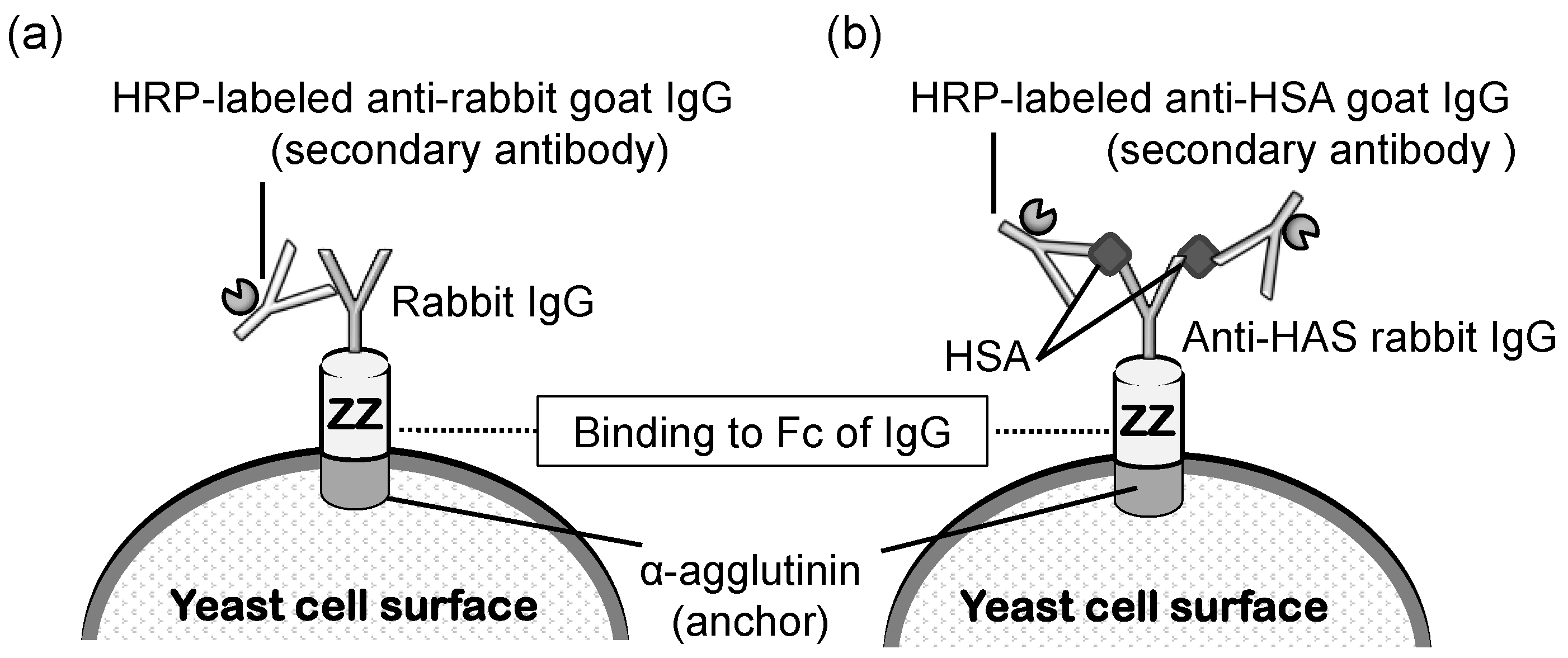
5. Applications as a Cell Sensor for Biosensing and Biomonitoring
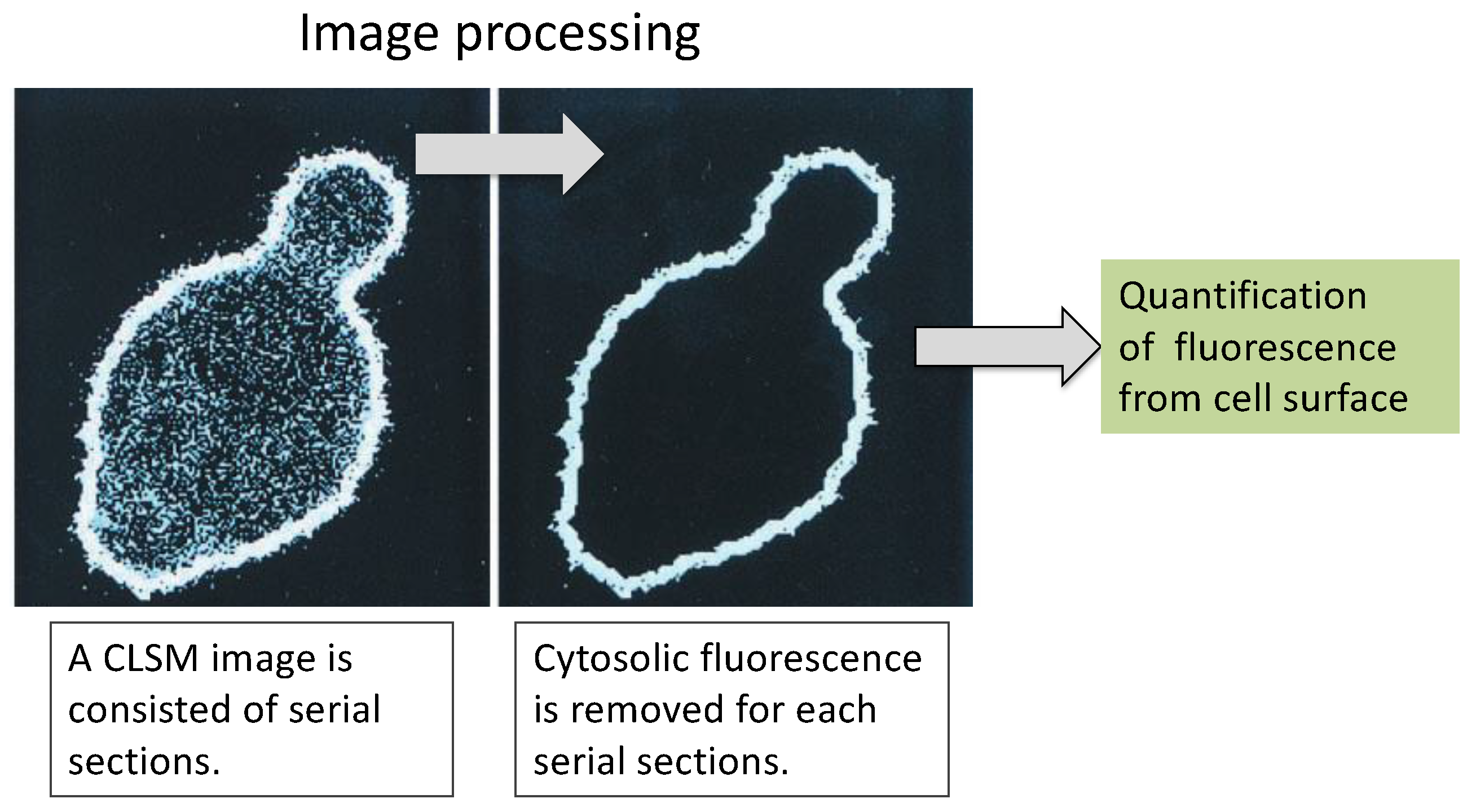
6. Application as a Tool in Biological Studies
7. Applications as a Whole-Cell Catalyst in Energy Production
8. Applications for Production of Useful Nutrients
9. Applications in the Production of Biochemicals and Pharmaceutics
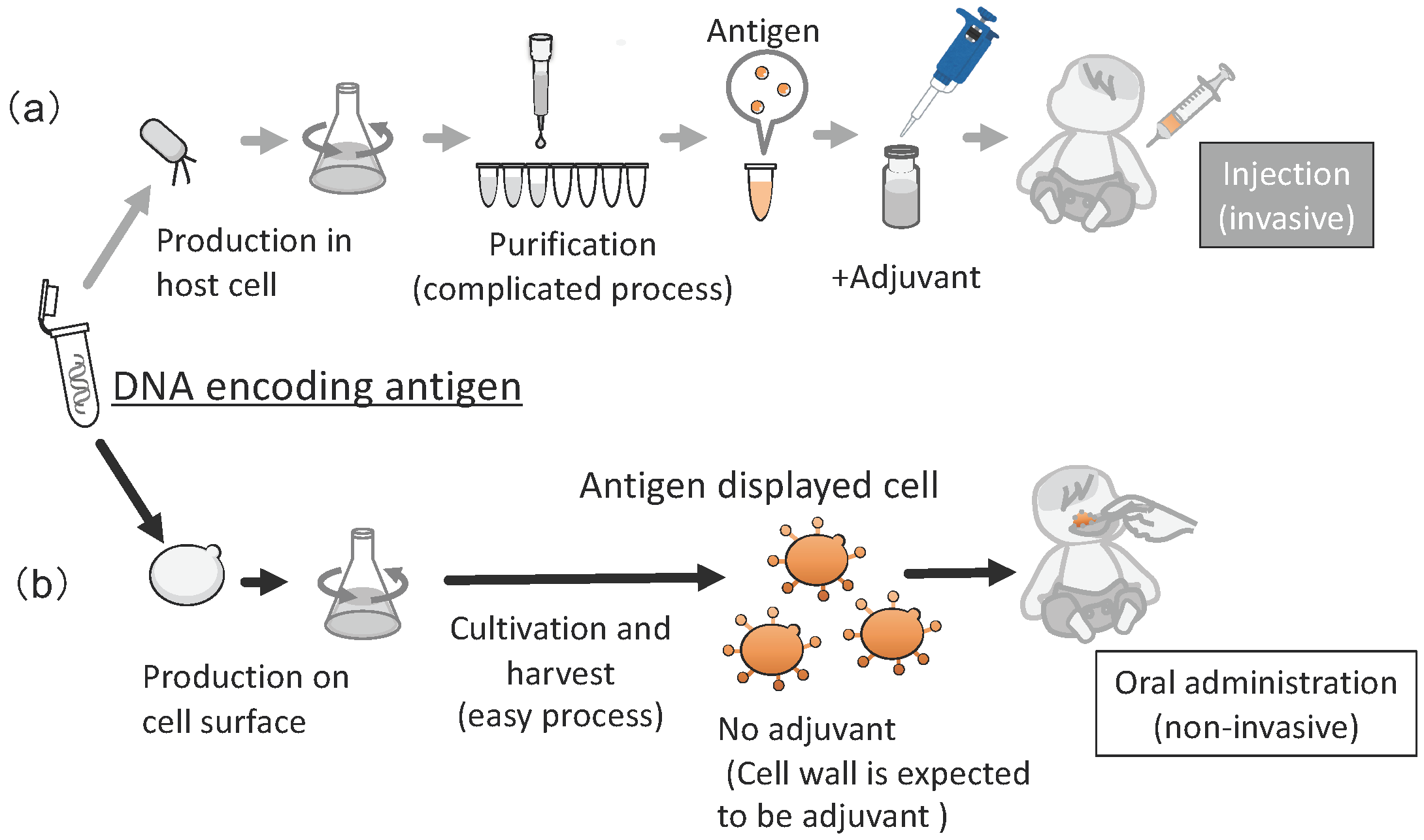
10. Application in Oral Vaccine Development
11. Summary
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baryshnikova, A.; Costanzo, M.; Dixon, S.; Vizeacoumar, F.J.; Myers, C.L.; Andrews, B.; Boone, C. Synthetic genetic array (SGA) analysis in Saccharomyces cerevisiae and Schizosaccharomyces pombe. Methods Enzymol. 2010, 470, 145–179. [Google Scholar] [PubMed]
- Nielsen, J. Yeast systems biology: Model organism and cell factory. Biotechnol. J. 2019, 14, e1800421. [Google Scholar] [CrossRef] [Green Version]
- Coronas-Serna, J.M.; Valenti, M.; Del Val, E.; Fernández-Acero, T.; Rodríguez-Escudero, I.; Mingo, J.; Luna, S.; Torices, L.; Pulido, R.; Molina, M.; et al. Modeling human disease in yeast: Recreating the PI3K-PTEN-Akt signaling pathway in Saccharomyces cerevisiae. Int. Microbiol. 2020, 23, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A.; Freitas, A.V.; Ralston, Z.A.; Tang, Z. Fission yeast Schizosaccharomyces pombe: A unicellular “micromammal” model organism. Curr. Protoc. 2021, 1, e151. [Google Scholar] [PubMed]
- Giga-Hama, Y.; Tohda, H.; Takegawa, K.; Kumagai, H. Schizosaccharomyces pombe minimum genome factory. Biotechnol. Appl. Biochem. 2007, 46, 147–155. [Google Scholar]
- Kitagaki, H.; Kitamoto, K. Breeding research on sake yeasts in Japan: History recent technological advances and future perspectives. Annu. Rev. Food Sci. Technol. 2013, 4, 215–235. [Google Scholar] [CrossRef]
- Akao, T.; Yashiro, I.; Hosoyama, A.; Kitagaki, H.; Horikawa, H.; Watanabe, D.; Akada, R.; Ando, Y.; Harashima, S.; Inoue, T.; et al. Whole-genome sequencing of sake yeast Saccharomyces cerevisiae Kyokai no. 7. DNA Res. 2011, 18, 423–434. [Google Scholar] [CrossRef] [Green Version]
- van Nierop, S.N.; Axcell, B.C.; Cantrell, I.C.; Rautenbach, M. Quality assessment of lager brewery yeast samples and strains using barley malt extracts with anti-yeast activity. Food Microbiol. 2009, 26, 192–196. [Google Scholar] [CrossRef]
- Lahue, C.; Madden, A.A.; Dunn, R.R.; Smukowski Heil, C. History and domestication of Saccharomyces cerevisiae in bread baking. Front. Genet. 2020, 1111, 584718. [Google Scholar] [CrossRef]
- Cobbold, C.A. The rise of alternative bread leavening technologies in the nineteenth century. Ann. Sci. 2018, 75, 21–39. [Google Scholar] [CrossRef]
- Hesseltine, C.W.; Shibasaki, K. Miso. III. Pure culture fermentation with Saccharomyces rouxii. Appl. Microbiol. 1961, 9, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Nakano, Y.; Satoh, H.; Ohtsubo, K. Improved palatability and bio-functionality of super-hard rice by soaking in a barley-koji miso suspension. Biosci. Biotechnol. Biochem. 2013, 77, 2419–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessou, P.; Keisam, S.; Tuikhar, N.; Gagara, M.; Farougou, S.; Jeyaram, K. High-Throughput Illumina MiSeq amplicon sequencing of yeast communities associated with indigenous dairy products from republics of Benin and Niger. Front. Microbiol. 2019, 310, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Hoek, S.A.; Darbani, B.; Zugaj, K.E.; Prabhala, B.K.; Biron, M.B.; Randelovic, M.; Medina, J.B.; Kell, D.B.; Borodina, I. Engineering the yeast Saccharomyces cerevisiae for the production of L-(+)-ergothioneine. Front. Bioeng. Biotechnol. 2019, 117, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anoop, V.; Rotaru, S.; Shwed, P.S.; Tayabali, A.F.; Arvanitakis, G. Review of current methods for characterizing virulence and pathogenicity potential of industrial Saccharomyces cerevisiae strains towards humans. FEMS Yeast Res. 2015, 15, fov057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Park, J.S.; Moon, J.Y.; Kim, K.Y.; Moon, H.M. The influence of glycosylation on secretion stability and immunogenicity of recombinant HBV pre-S antigen synthesized in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2003, 303, 427–432. [Google Scholar] [CrossRef]
- Schiller, J.T.; Müller, M. Next generation prophylactic human papillomavirus vaccines. Lancet Oncol. 2015, 16, e217–e225. [Google Scholar] [CrossRef]
- López-Gálvez, F.; Gómez, P.A.; Artés, F.; Artés-Hernández, F.; Aguayo, E. Interactions between microbial food safety and environmental sustainability in the fresh produce supply chain. Foods 2021, 10, 1655. [Google Scholar] [CrossRef]
- Lokko, Y.; Heijde, M.; Schebesta, K.; Scholtès, P.; Van Montagu, M.; Giacca, M. Biotechnology and the bioeconomy–Towards inclusive and sustainable industrial development. New Biotechnol. 2018, 40, 5–10. [Google Scholar] [CrossRef]
- Timmis, K.; de Lorenzo, V.; Verstraete, W.; Ramos, J.L.; Danchin, A.; Brüssow, H.; Singh, B.K.; Timmis, J.K. The contribution of microbial biotechnology to economic growth and employment creation. Microb. Biotechnol. 2017, 10, 1137–1144. [Google Scholar] [CrossRef]
- Fagunwa, O.E.; Olanbiwoninu, A.A. Accelerating the sustainable development goals through microbiology: Some efforts and opportunities. Access Microbiol. 2020, 2, acmi000112. [Google Scholar] [PubMed]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Grimm, S.; Lundberg, E.; Yu, F.; Shibasaki, S.; Vernet, E.; Skogs, M.; Nygren, P.Å.; Gräslund, T. Selection and characterisation of affibody molecules inhibiting the interaction between Ras and Raf in vitro. New Biotechnol. 2010, 27, 766–773. [Google Scholar] [CrossRef]
- von Witting, E.; Lindbo, S.; Lundqvist, M.; Möller, M.; Wisniewski, A.; Kanje, S.; Rockberg, J.; Tegel, H.; Åstrand, M.; Uhlén, M.; et al. Small bispecific affinity proteins for simultaneous target binding and albumin-associated half-life extension. Mol. Pharm. 2021, 18, 328–337. [Google Scholar] [CrossRef]
- Okano, K.; Zhang, Q.; Kimura, S.; Narita, J.; Tanaka, T.; Fukuda, H.; Kondo, A. System using tandem repeats of the cA peptidoglycan-binding domain from Lactococcus lactis for display of both N- and C-terminal fusions on cell surfaces of lactic acid bacteria. Appl. Environ. Microbiol. 2008, 74, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Wernérus, H.; Lehtiö, J.; Samuelson, P.; Ståhl, S. Engineering of staphylococcal surfaces for biotechnological applications. J. Biotechnol. 2002, 96, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salema, V.; Fernández, L.Á. Escherichia coli surface display for the selection of nanobodies. Microb. Biotechnol. 2017, 10, 1468–1484. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Khera, E.; Wen, F. Engineering novel and improved biocatalysts by cell surface display. Ind. Eng. Chem. Res. 2015, 54, 4021–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, M.; Tanaka, A. Genetic immobilization of proteins on the yeast cell surface. Biotechnol. Adv. 2000, 18, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Maeda, H.; Ueda, M. Molecular display technology using yeast-arming technology. Anal. Sci. 2009, 25, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, S.; Ueda, M. Development of yeast molecular display systems focused on therapeutic proteins, enzymes, and foods: Functional analysis of proteins and its application to bioconversion. Recent Pat. Biotechnol. 2010, 4, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Kosugi, A.; Tanaka, T.; Kuroda, K.; Shibasaki, S.; Ogino, C.; Ueda, M.; Fukuda, H.; Doi, R.H.; Kondo, A. Regulation of the display ratio of enzymes on the Saccharomyces cerevisiae cell surface by the immunoglobulin G and cellulosomal enzyme binding domains. Appl. Environ. Microbiol. 2009, 75, 4149–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanAntwerp, J.J.; Wittrup, K.D. Fine affinity discrimination by yeast surface display and flow cytometry. Biotechnol. Prog. 2000, 16, 31–37. [Google Scholar] [CrossRef]
- Sun, Y.; Ban, B.; Bradbury, A.; Ansari, G.A.; Blake, D.A. Combining yeast display and competitive FACS to select rare hapten-specific clones from recombinant antibody libraries. Anal. Chem. 2016, 88, 9181–9189. [Google Scholar] [CrossRef] [PubMed]
- Borodina, I.; Jensen, B.M.; Søndergaard, I.; Poulsen, L.K. Display of wasp venom allergens on the cell surface of Saccharomyces cerevisiae. Microb. Cell Fact. 2010, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anonymous. Arming yeast with cell-surface catalysts. Chem. Eng. News 1997, 75, 32. [Google Scholar]
- Ueda, M.; Tanaka, A. Cell surface engineering of yeast: Construction of arming yeast with biocatalyst. J. Biosci. Bioeng. 2000, 90, 125–136. [Google Scholar] [CrossRef]
- Kondo, A.; Ueda, M. Yeast cell-surface display–Applications of molecular display. Appl. Microbiol. Biotechnol. 2004, 64, 28–40. [Google Scholar] [CrossRef]
- Seong, K.T.; Katakura, Y.; Ninomiya, K.; Bito, Y.; Katahira, S.; Kondo, A.; Ueda, M.; Shioya, S. Effect of flocculation on performance of arming yeast in direct ethanol fermentation. Appl. Microbiol. Biotechnol. 2006, 73, 60–66. [Google Scholar] [CrossRef]
- Andreu, C.; Del Olmo, M.L. Yeast arming systems: Pros and cons of different protein anchors and other elements required for display. Appl. Microbiol. Biotechnol. 2018, 102, 2543–2561. [Google Scholar] [CrossRef]
- Nicchi, S.; Giuliani, M.; Giusti, F.; Pancotto, L.; Maione, D.; Delany, I.; Galeotti, C.L.; Brettoni, C. Decorating the surface of Escherichia coli with bacterial lipoproteins: A comparative analysis of different display systems. Microb. Cell Fact. 2021, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Vahed, M.; Ramezani, F.; Tafakori, V.; Mirbagheri, V.S.; Najafi, A.; Ahmadian, G. Molecular dynamics simulation and experimental study of the surface-display of SPA protein via Lpp-OmpA system for screening of IgG. AMB Express. 2020, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Narita, J.; Okano, K.; Kitao, T.; Ishida, S.; Sewaki, T.; Sung, M.H.; Fukuda, H.; Kondo, A. Display of alpha-amylase on the surface of Lactobacillus casei cells by use of the PgsA anchor protein and production of lactic acid from starch. Appl. Environ. Microbiol. 2006, 72, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage, G.; Bussey, H. Cell wall assembly in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cid, V.J.; Durán, A.; del Rey, F.; Snyder, M.P.; Nombela, C.; Sánchez, M. Molecular basis of cell integrity and morphogenesis in Saccharomyces cerevisiae. Microbiol. Rev. 1995, 59, 345–386. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Ueda, M. Bioadsorption strategies with yeast molecular display technology. Biocontrol Sci. 2014, 19, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M. Establishment of cell surface engineering and its development. Biosci. Biotechnol. Biochem. 2016, 80, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Kipnis, P.; Thomas, N.; Ovalle, R.; Lipke, P.N. The ER-Golgi v-SNARE Bet1p is required for cross-linking alpha-agglutinin to the cell wall in yeast. Microbiology 2004, 150, 3219–3228. [Google Scholar] [CrossRef] [Green Version]
- Neogi, S.; Sharma, V.; Khan, N.; Chaurasia, D.; Ahmad, A.; Chauhan, S.; Singh, A.; You, S.; Pandey, A.; Bhargava, P.C. Sustainable biochar: A facile strategy for soil and environmental restoration energy generation mitigation of global climate change and circular bioeconomy. Chemosphere 2022, 293, 133474. [Google Scholar] [CrossRef]
- Bellis, D.J.; Satake, K.; Tsunoda, K.; McLeod, C.W. Environmental monitoring of historical change in arsenic deposition with tree bark pockets. J. Environ. Monit. 2003, 5, 671–674. [Google Scholar] [CrossRef]
- Perlatti, F.; Martins, E.P.; de Oliveira, D.P.; Ruiz, F.; Asensio, V.; Rezende, C.F.; Otero, X.L.; Ferreira, T.O. Copper release from waste rocks in an abandoned mine (NE, Brazil) and its impacts on ecosystem environmental quality. Chemosphere 2021, 262, 127843. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, J.K.; Paliwal, A.; Saraf, P. Effects of heavy metals on reproduction owing to infertility. J. Biochem. Mol. Toxicol. 2021, 35, e22823. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.R.; Schoenung, J.M. Human health and ecological toxicity potentials due to heavy metal content in waste electronic devices with flat panel displays. J. Hazard. Mater. 2010, 177, 251–259. [Google Scholar] [CrossRef]
- Cowan, A.R.; Costanzo, C.M.; Benham, R.; Loveridge, E.J.; Moody, S.C. Fungal bioremediation of polyethylene: Challenges and perspectives. J. Appl. Microbiol. 2022, 132, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Fomina, M.; Gadd, G.M. Biosorption: Current perspectives on concept definition and application. Bioresour. Technol. 2014, 160, 3–14. [Google Scholar] [CrossRef]
- Salama, A.K.; Osman, K.A.; Gouda, N.A. Remediation of lead and cadmium-contaminated soils. Int. J. Phytoremediation 2016, 18, 364–367. [Google Scholar] [CrossRef]
- Usman, K.; Abu-Dieyeh, M.H.; Zouari, N.; Al-Ghouti, M.A. Lead (Pb) bioaccumulation and antioxidative responses in Tetraena qataranse. Sci. Rep. 2020, 10, 17070. [Google Scholar] [CrossRef]
- Kuroda, K.; Ueda, M.; Shibasaki, S.; Tanaka, A. Cell surface-engineered yeast with ability to bind and self-aggregate in response to copper ion. Appl. Microbiol. Biotechnol. 2002, 59, 259–264. [Google Scholar]
- Kuroda, K.; Ueda, M. Effective display of metallothionein tandem repeats on the bioadsorption of cadmium ion. Appl. Microbiol. Biotechnol. 2006, 70, 458–463. [Google Scholar] [CrossRef]
- Ferreira, C.R.; Gahl, W.A. Disorders of metal metabolism. Transl. Sci. Rare Dis. 2017, 2, 101–139. [Google Scholar] [CrossRef] [Green Version]
- Filler, G.; McIntyre, C. Chromium: Rise and shine in peritoneal dialysis patients? Perit. Dial. Int. 2019, 39, 320–322. [Google Scholar] [CrossRef]
- Arya, S.; Kumar, S. Bioleaching: Urban mining option to curb the menace of E-waste challenge. Bioengineered 2020, 11, 640–660. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Candeago, R.; Rim, G.; Raymond, D.; Park, A.A.; Su, X. Electrochemical approaches for selective recovery of critical elements in hydrometallurgical processes of complex feedstocks. iScience 2021, 24, 102374. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.C.; Ali, K.; Sonnenschein, M.; Robrahn, L.; Strauss, D.; Narberhaus, F.; Masepohl, B. Molybdate uptake by Agrobacterium tumefaciens correlates with the cellular molybdenum cofactor status. Mol. Microbiol. 2016, 101, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, T.; Shimada, M.; Kuroda, K.; Ueda, M. Molecular design of yeast cell surface for adsorption and recovery of molybdenum one of rare metals. Appl. Microbiol. Biotechnol. 2010, 86, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Nishitani, T.; Ueda, M. Specific adsorption of tungstate by cell surface display of the newly designed ModE mutant. Appl. Microbiol. Biotechnol. 2012, 96, 153–159. [Google Scholar] [CrossRef]
- Kuroda, K.; Shibasaki, S.; Ueda, M.; Tanaka, A. Cell surface-engineered yeast displaying a histidine oligopeptide (hexa-His) has enhanced adsorption of and tolerance to heavy metal ions. Appl. Microbiol. Biotechnol. 2001, 57, 697–701. [Google Scholar] [CrossRef]
- Beszterda, M.; Frański, R. Endocrine disruptor compounds in environment: As a danger for children health. Pediatr. Endocrinol. Diabetes Metab. 2018, 24, 88–95. [Google Scholar] [CrossRef]
- Viguié, C.; Chaillou, E.; Gayrard, V.; Picard-Hagen, N.; Fowler, P.A. Toward a better understanding of the effects of endocrine disrupting compounds on health: Human-relevant case studies from sheep models. Mol. Cell Endocrinol. 2020, 505, 110711. [Google Scholar] [CrossRef]
- Balabanič, D.; Rupnik, M.; Klemenčič, A.K. Negative impact of endocrine-disrupting compounds on human reproductive health. Reprod. Fertil. Dev. 2011, 23, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Bradley, P.M.; Battaglin, W.A.; Iwanowicz, L.R.; Clark, J.M.; Journey, C.A. Aerobic biodegradation potential of endocrine-disrupting chemicals in surface-water sediment at Rocky Mountain National Park USA. Environ. Toxicol. Chem. 2016, 35, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Gea, M.; Toso, A.; Schilirò, T. Estrogenic activity of biological samples as a biomarker. Sci. Total Environ. 2020, 740, 140050. [Google Scholar] [CrossRef] [PubMed]
- Yasui, M.; Shibasaki, S.; Kuroda, K.; Ueda, M.; Kawada, N.; Nishikawa, J.; Nishihara, T.; Tanaka, A. An arming yeast with the ability to entrap fluorescent 17beta-estradiol on the cell surface. Appl. Microbiol. Biotechnol. 2002, 59, 329–331. [Google Scholar] [PubMed]
- Nakamura, H.; Shimomura-Shimizu, M.; Karube, I. Development of microbial sensors and their application. Adv. Biochem. Eng. Biotechnol. 2008, 109, 351–394. [Google Scholar]
- Mulla, M.Y.; Torsi, L.; Manoli, K. Electronic biosensors based on EGOFETs. Methods Enzymol. 2020, 642, 403–433. [Google Scholar] [PubMed]
- Liu, W.; Wang, J.; Mitsui, K.; Shen, H.; Tsurugi, K. Interaction of the GTS1 gene product with glyceraldehyde- 3-phosphate dehydrogenase 1 required for the maintenance of the metabolic oscillations of the yeast Saccharomyces cerevisiae. Eur. J. Biochem. 2002, 269, 3560–3569. [Google Scholar] [CrossRef] [PubMed]
- Chalfie, M. Green fluorescent protein. Photochem. Photobiol. 1995, 62, 651–656. [Google Scholar] [CrossRef]
- Ye, K.; Shibasaki, S.; Ueda, M.; Murai, T.; Kamasawa, N.; Osumi, M.; Shimizu, K.; Tanaka, A. Construction of an engineered yeast with glucose-inducible emission of green fluorescence from the cell surface. Appl. Microbiol. Biotechnol. 2000, 54, 90–96. [Google Scholar] [CrossRef]
- Umemura, K.; Atomi, H.; Kanai, T.; Takeshita, S.; Kanayama, N.; Ueda, M.; Tanaka, A. Derepression of gene expression mediated by the 5’ upstream region of the isocitrate lyase gene of Candida tropicalis is controlled by two distinct regulatory pathways in Saccharomyces cerevisiae. Eur. J. Biochem. 1997, 243, 748–752. [Google Scholar] [CrossRef]
- Shibasaki, S.; Ueda, M.; Ye, K.; Shimizu, K.; Kamasawa, N.; Osumi, M.; Tanaka, A. Creation of cell surface-engineered yeast that display different fluorescent proteins in response to the glucose concentration. Appl. Microbiol. Biotechnol. 2001, 57, 528–533. [Google Scholar]
- Krasnenko, V.; Tkaczyk, A.H.; Tkaczyk, E.R.; Mauring, K. Physicochemical properties of blue fluorescent protein determined via molecular dynamics simulation. Biopolymers 2008, 89, 1136–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; El Masoudi, A.; Pronk, J.T.; van Gulik, W.M. Quantitative physiology of non-energy-limited retentostat cultures of Saccharomyces cerevisiae at near-zero specific growth rates. Appl. Environ. Microbiol. 2019, 85, e01161-19. [Google Scholar] [CrossRef] [PubMed]
- Kliewe, F.; Kumme, J.; Grigat, M.; Hintze, S.; Schüller, H.J. Opi1 mediates repression of phospholipid biosynthesis by phosphate limitation in the yeast Saccharomyces cerevisiae. Yeast 2017, 34, 67–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Berg, B.; Chembath, A.; Jefferies, D.; Basle, A.; Khalid, S.; Rutherford, J.C. Structural basis for Mep2 ammonium transceptor activation by phosphorylation. Nat. Commun. 2016, 7, 11337. [Google Scholar] [CrossRef] [Green Version]
- Marini, A.M.; Soussi-Boudekou, S.; Vissers, S.; Andre, B. A family of ammonium transporters in Saccharomyces cerevisiae. Mol. Cell Biol. 1997, 17, 4282–4293. [Google Scholar] [CrossRef]
- Tate, J.J.; Rai, R.; Cooper, T.G. Ammonia-specific regulation of Gln3 localization in Saccharomyces cerevisiae by protein kinase Npr1. J. Biol. Chem. 2006, 281, 28460–28469. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, S.; Ninomiya, Y.; Ueda, M.; Iwahashi, M.; Katsuragi, T.; Tani, Y.; Harashima, S.; Tanaka, A. Intelligent yeast strains with the ability to self-monitor the concentrations of intra- and extracellular phosphate or ammonium ion by emission of fluorescence from the cell surface. Appl. Microbiol. Biotechnol. 2001, 57, 702–707. [Google Scholar]
- Mirisola, M.G.; Gallo, A.; De Leo, G. Ras-pathway has a dual role in yeast galactose metabolism. FEBS Lett. 2007, 581, 2009–2016. [Google Scholar] [CrossRef] [Green Version]
- Kremers, G.J.; Goedhart, J.; van den Heuvel, D.J.; Gerritsen, H.C.; Gadella, T.W., Jr. Improved green and blue fluorescent proteins for expression in bacteria and mammalian cells. Biochemistry 2007, 46, 3775–3783. [Google Scholar] [CrossRef]
- Sen, T.; Mamontova, A.V.; Titelmayer, A.V.; Shakhov, A.M.; Astafiev, A.A.; Acharya, A.; Lukyanov, K.A.; Krylov, A.I.; Bogdanov, A.M. Influence of the first chromophore-forming residue on photobleaching and oxidative photoconversion of EGFP and EYFP. Int. J. Mol. Sci. 2019, 20, 5229. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, S.; Tanaka, A.; Ueda, M. Development of combinatorial bioengineering using yeast cell surface display-order-made design of cell and protein for bio-monitoring. Biosens. Bioelectron. 2003, 19, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, J.; Jovanovic, G.; Kotta-Loizou, I.; Buck, M. Single-step method for β-galactosidase assays in Escherichia coli using a 96-well microplate reader. Anal. Biochem. 2016, 503, 56–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, R.S.; Hartmann, K. Interferon therapies in small animals. Vet. J. 2021, 271, 105648. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.E.; Bartlett, A.W.; Tangye, S.G. Severe COVID-19 represents an undiagnosed primary immunodeficiency in a high proportion of infected individuals. Clin. Transl. Immunol. 2022, 11, e1365. [Google Scholar] [CrossRef]
- Takagi, T.; Osumi, M.; Shinohara, A. Ultrastructural analysis in yeast reveals a meiosis-specific actin-containing nuclear bundle. Commun. Biol. 2021, 254, 1009. [Google Scholar] [CrossRef]
- Shibasaki, Y.; Kamasawa, N.; Shibasaki, S.; Zou, W.; Murai, T.; Ueda, M.; Tanaka, A.; Osumi, M. Cytochemical evaluation of localization and secretion of a heterologous enzyme displayed on yeast cell surface. FEMS Microbiol. Lett. 2000, 192, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.T.; Cheng, L.; Kain, S.R. Optimized codon usage and chromophore mutations provide enhanced sensitivity with the green fluorescent protein. Nucleic Acids Res. 1996, 24, 4592–4593. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, S.; Ueda, M.; Iizuka, T.; Hirayama, M.; Ikeda, Y.; Kamasawa, N.; Osumi, M.; Tanaka, A. Quantitative evaluation of the enhanced green fluorescent protein displayed on the cell surface of Saccharomyces cerevisiae by fluorometric and confocal laser scanning microscopic analyses. Appl. Microbiol. Biotechnol. 2001, 55, 471–475. [Google Scholar] [CrossRef]
- Eswaran, N.; Parameswaran, S.; Johnson, T.S. Biofuels and sustainability. Methods Mol. Biol. 2021, 2290, 317–342. [Google Scholar]
- Ramamurthy, P.C.; Singh, S.; Kapoor, D.; Parihar, P.; Samuel, J.; Prasad, R.; Kumar, A.; Singh, J. Microbial biotechnological approaches: Renewable bioprocessing for the future energy systems. Microb. Cell Fact. 2021, 20, 55. [Google Scholar] [CrossRef]
- Hamaamin, H.N.; Hameed, H.A.; Jamalis, J.; Shakya, S.; Chander, S.; Kharkwal, H.; Murugesan, S.; Ajit, B.V.; Pyarelal, G.P. Potential inhibitory activity of phytoconstituents against black fungus: In silico ADMET molecular docking and MD simulation studies. Comput Toxicol. 2022, 24, 100247. [Google Scholar] [CrossRef] [PubMed]
- Murai, T.; Ueda, M.; Yamamura, M.; Atomi, H.; Shibasaki, Y.; Kamasawa, N.; Osumi, M.; Amachi, T.; Tanaka, A. Construction of a starch-utilizing yeast by cell surface engineering. Appl. Environ. Microbiol. 1997, 63, 1362–1366. [Google Scholar] [CrossRef] [Green Version]
- Murai, T.; Ueda, M.; Shibasaki, Y.; Kamasawa, N.; Osumi, M.; Imanaka, T.; Tanaka, A. Development of an arming yeast strain for efficient utilization of starch by co-display of sequential amylolytic enzymes on the cell surface. Appl. Microbiol. Biotechnol. 1999, 51, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Murai, T.; Shibasaki, Y.; Kamasawa, N.; Osumi, M.; Tanaka, A. Molecular breeding of polysaccharide-utilizing yeast cells by cell surface engineering. Ann. N. Y. Acad. Sci. 1998, 13864, 528–537. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Shetti, N.P.; Reddy, K.R.; Kwon, E.E.; Nadagouda, M.N.; Aminabhavi, T.M. Biomass utilization and production of biofuels from carbon neutral materials. Environ. Pollut. 2021, 276, 116731. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.R.; Natarajan, V.; Krishnan, C. Marine enzymes and microorganisms for bioethanol production. Adv. Food Nutr. Res. 2017, 80, 181–197. [Google Scholar] [PubMed]
- Toyosawa, Y.; Ikeo, M.; Taneda, D.; Okino, S. Quantitative analysis of adsorption and desorption behavior of individual cellulase components during the hydrolysis of lignocellulosic biomass with the addition of lysozyme. Bioresour. Technol. 2017, 234, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yamada, R.; Ogino, C.; Kondo, A. Recent developments in yeast cell surface display toward extended applications in biotechnology. Appl. Microbiol. Biotechnol. 2012, 95, 577–591. [Google Scholar] [CrossRef]
- Liu, Z.; Inokuma, K.; Ho, S.H.; Haan, R.d.; Hasunuma, T.; Zyl, W.H.; Kondo, A. Combined cell-surface display- and secretion-based strategies for production of cellulosic ethanol with Saccharomyces cerevisiae. Biotechnol. Biofuels 2015, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Smeriglio, A.; Calderaro, A.; Denaro, M.; Laganà, G.; Bellocco, E. Effects of isolated isoflavones intake on health. Curr. Med. Chem. 2019, 26, 5094–5107. [Google Scholar] [CrossRef]
- Kaya, M.; Ito, J.; Kotaka, A.; Matsumura, K.; Bando, H.; Sahara, H.; Ogino, C.; Shibasaki, S.; Kuroda, K.; Ueda, M.; et al. Isoflavone aglycones production from isoflavone glycosides by display of beta-glucosidase from Aspergillus oryzae on yeast cell surface. Appl. Microbiol. Biotechnol. 2008, 79, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Xu, P.; Gallagher, S. Immunoblotting and Immunodetection. Curr. Protoc. Cell Biol. 2017, 74, 6.2.1–6.2.37. [Google Scholar] [CrossRef] [PubMed]
- Olender, L.; Thapa, R.; Gazit, R. Isolation of murine myeloid progenitor populations by CD34/CD150 surface markers. Cells 2022, 11, 350. [Google Scholar] [CrossRef] [PubMed]
- Takei, J.; Asano, T.; Tanaka, T.; Sano, M.; Hosono, H.; Nanamiya, R.; Tateyama, N.; Saito, M.; Suzuki, H.; Harada, H.; et al. Development of a novel anti-HER2 monoclonal antibody H2Mab-181 for gastric cancer. Monoclon. Antib. Immunodiagn. Immunother. 2021, 40, 168–176. [Google Scholar] [CrossRef]
- Kang, S.; Tanaka, T.; Narazaki, M.; Kishimoto, T. Targeting interleukin-6 signaling in clinic. Immunity. 2019, 50, 1007–1023. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Iwasaki, T.; Kitano, S.; Satake, A.; Nomura, S.; Furukawa, T.; Matsui, K.; Sano, H. IL-2-anti-IL-2 monoclonal antibody immune complexes inhibit collagen-induced arthritis by augmenting regulatory T cell functions. J. Immunol. 2018, 201, 1899–1906. [Google Scholar] [CrossRef] [Green Version]
- Coluccio, M.L.; Grillo, F.; Onesto, V.; Garo, V.; Scala, C.; Cuzzola, P.; Calfa, M.; Candeloro, P.; Gentile, F.; Piletsky, S.; et al. Enhancing antibodies’ binding capacity through oriented functionalization of plasmonic surfaces. Nanomaterials 2021, 11, 2620. [Google Scholar] [CrossRef]
- Lando, P.; Gabriel, J.; Berzins, K.; Perlmann, P. Determination of the immunoglobulin class of complement-dependent cytotoxic antibodies in serum of D23 hepatoma-bearing rats. Scand. J. Immunol. 1980, 11, 253–260. [Google Scholar] [CrossRef]
- Nakamura, Y.; Shibasaki, S.; Ueda, M.; Tanaka, A.; Fukuda, H.; Kondo, A. Development of novel whole-cell immunoadsorbents by yeast surface display of the IgG-binding domain. Appl. Microbiol. Biotechnol. 2001, 57, 500–505. [Google Scholar]
- Shibasaki, S.; Kawabata, A.; Ishii, J.; Yagi, S.; Kadonosono, T.; Kato, M.; Fukuda, N.; Kondo, A.; Ueda, M. Construction of a novel synergistic system for production and recovery of secreted recombinant proteins by the cell surface engineering. Appl. Microbiol. Biotechnol. 2007, 75, 821–828. [Google Scholar] [CrossRef]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. More than 50 long-term effects of COVID-19: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 16144. [Google Scholar] [CrossRef] [PubMed]
- Alene, K.A.; Elagali, A.; Barth, D.D.; Rumisha, S.F.; Amratia, P.; Weiss, D.J.; Atalell, K.A.; Erena, A.K.; Gething, P.W.; Clements, A.C.A. Spatial codistribution of HIV tuberculosis and malaria in Ethiopia. BMJ Glob. Health 2022, 7, e007599. [Google Scholar] [CrossRef] [PubMed]
- Childs, L.M.; Abuelezam, N.N.; Dye, C.; Gupta, S.; Murray, M.B.; Williams, B.G.; Buckee, C.O. Modelling challenges in context: Lessons from malaria HIV and tuberculosis. Epidemics 2015, 10, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and mechanisms of antifungal resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Saunus, J.M.; Kazoullis, A.; Farah, C.S. Cellular and molecular mechanisms of resistance to oral Candida albicans infections. Front Biosci. 2008, 13, 5345–5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibasaki, S.; Aoki, W.; Nomura, T.; Miyoshi, A.; Tafuku, S.; Sewaki, T.; Ueda, M. An oral vaccine against candidiasis generated by a yeast molecular display system. Pathog. Dis. 2013, 69, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Leu, S.J.; Lee, Y.C.; Lee, C.H.; Liao, P.Y.; Chiang, C.W.; Yang, C.M.; Su, C.H.; Ou, T.Y.; Liu, K.J.; Lo, H.J.; et al. Generation and characterization of single chain variable fragment against alpha-enolase of Candida albicans. Int. J. Mol Sci. 2020, 21, 2903. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, S.; Karasaki, M.; Tafuku, S.; Aoki, W.; Sewaki, T.; Ueda, M. Oral immunization against candidiasis using Lactobacillus casei displaying enolase 1 from Candida albicans. Sci. Pharm. 2014, 82, 697–708. [Google Scholar] [CrossRef]
- Shibasaki, S.; Ueda, M. Preparation of an oral vaccine by proteome analysis and molecular display technology. Methods Mol. Biol. 2017, 1625, 237–245. [Google Scholar]
- Vendramin, N.; Alencar, A.L.F.; Iburg, T.M.; Dahle, M.K.; Wessel, Ø.; Olsen, A.B.; Rimstad, E.; Olesen, N.J. Piscine orthoreovirus infection in Atlantic salmon (Salmo salar) protects against subsequent challenge with infectious hematopoietic necrosis virus (IHNV). Vet. Res. 2018, 1349, 30. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, T.; Kinoshita, S.; Kim, W.S.; Higashi, S.; Yoshimizu, M. Nucleotide diversity of Japanese isolates of infectious hematopoietic necrosis virus (IHNV) based on the glycoprotein gene. Dis. Aquat. Organ. 2006, 71, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S. Novel bioconversion systems using a yeast molecular display system. Yakugaku Zasshi 2010, 130, 1437–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaru, Y.; Ohtsuka, M.; Kato, K.; Manabe, S.; Kuroda, K.; Sanada, M.; Ueda, M. Application of the arming system for the expression of the 380R antigen from red sea bream iridovirus (RSIV) on the surface of yeast cells: A first step for the development of an oral vaccine. Biotechnol. Prog. 2006, 22, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Houston-Cummings, N.R.; Prinz, B.; Moore, R.; Bobrowicz, B.; Davidson, R.C.; Wildt, S.; Stadheim, T.A.; Zha, D. A novel fragment of antigen binding (Fab) surface display platform using glycoengineered Pichia pastoris. J. Immunol. Methods 2012, 375, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Gätjen, D.; Tomszak, F.; Dettmann, J.C.; Droste, M.; Nölle, V.; Wieczorek, M. Design of a novel switchable antibody display system in Pichia pastoris. Appl. Microbiol. Biotechnol. 2022, 106, 6209–6224. [Google Scholar] [CrossRef]
- Yuzbasheva, E.Y.; Yuzbashev, T.V.; Perkovskaya, N.I.; Mostova, E.B.; Vybornaya, T.V.; Sukhozhenko, A.V.; Toropygin, I.Y.; Sineoky, S.P. Cell surface display of Yarrowia lipolytica lipase Lip2p using the cell wall protein YlPir1p its characterization and application as a whole-cell biocatalyst. Appl. Biochem. Biotechnol. 2015, 175, 3888–3900. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibasaki, S.; Ueda, M. Progress of Molecular Display Technology Using Saccharomyces cerevisiae to Achieve Sustainable Development Goals. Microorganisms 2023, 11, 125. https://doi.org/10.3390/microorganisms11010125
Shibasaki S, Ueda M. Progress of Molecular Display Technology Using Saccharomyces cerevisiae to Achieve Sustainable Development Goals. Microorganisms. 2023; 11(1):125. https://doi.org/10.3390/microorganisms11010125
Chicago/Turabian StyleShibasaki, Seiji, and Mitsuyoshi Ueda. 2023. "Progress of Molecular Display Technology Using Saccharomyces cerevisiae to Achieve Sustainable Development Goals" Microorganisms 11, no. 1: 125. https://doi.org/10.3390/microorganisms11010125
APA StyleShibasaki, S., & Ueda, M. (2023). Progress of Molecular Display Technology Using Saccharomyces cerevisiae to Achieve Sustainable Development Goals. Microorganisms, 11(1), 125. https://doi.org/10.3390/microorganisms11010125