
Protocooperative Effect of Sphaerodes mycoparasitica Biocontrol and Crop Genotypes on FHB Mycotoxin Reduction in Bread and Durum Wheat Grains Intended for Human and Animal Consumption


Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Trials and Sampling Methods
2.2. Seed Sample Preparation
2.3. Analysis of Fusarium Profile Using High-Throughput Sequencing and Bioinformatics
2.4. Mycotoxin Analysis by Liquid Chromatography with Tandem Mass Spectroscopy
2.4.1. Sample Preparation
2.4.2. Mycotoxin Analysis by LC-MS/MS
2.5. Statistical Analysis
3. Results
3.1. Mycotoxins Results
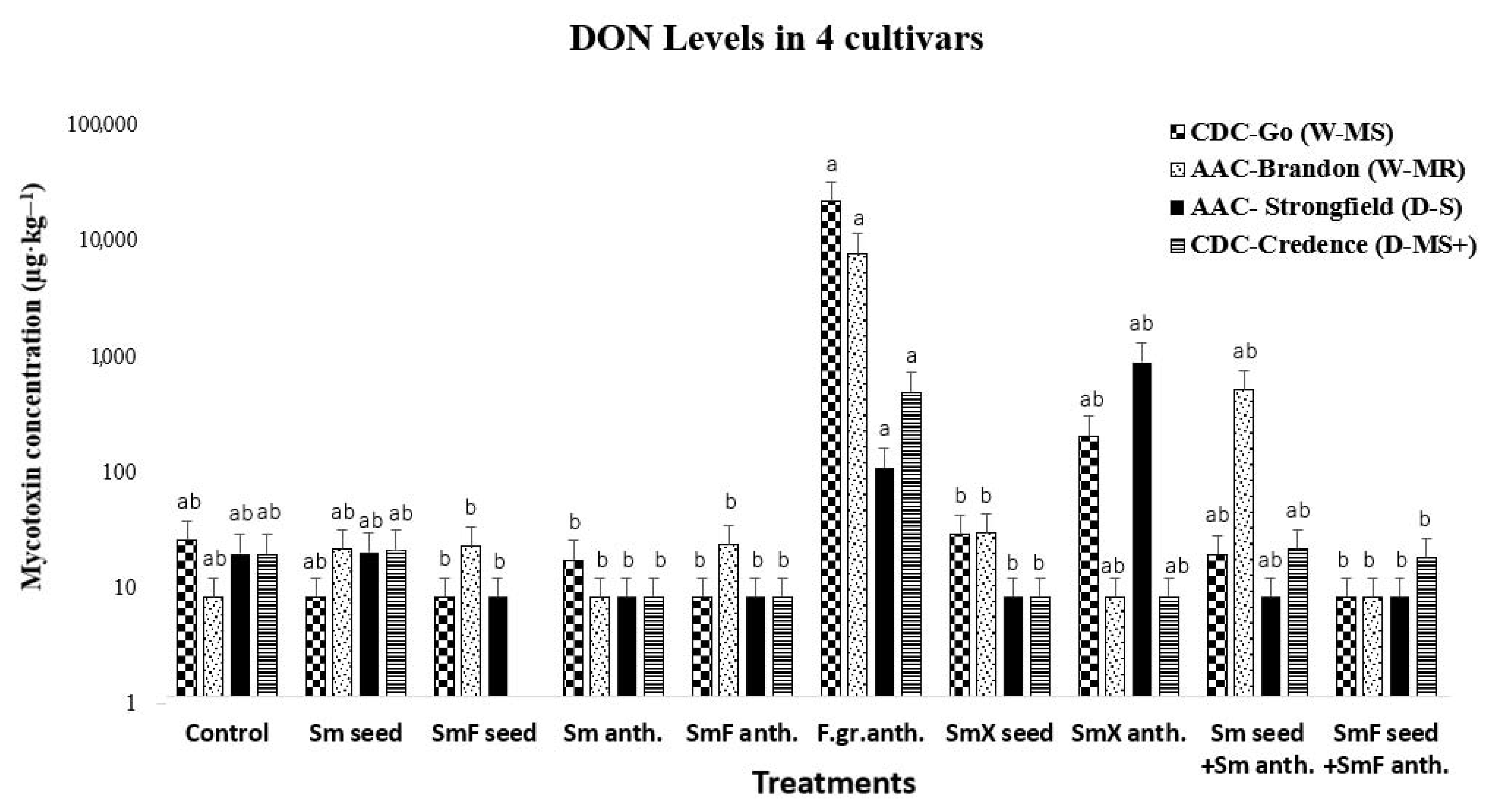
3.2. Deoxynivalenol (DON) Concentration in Common Wheat and Durum Wheat Cultivars
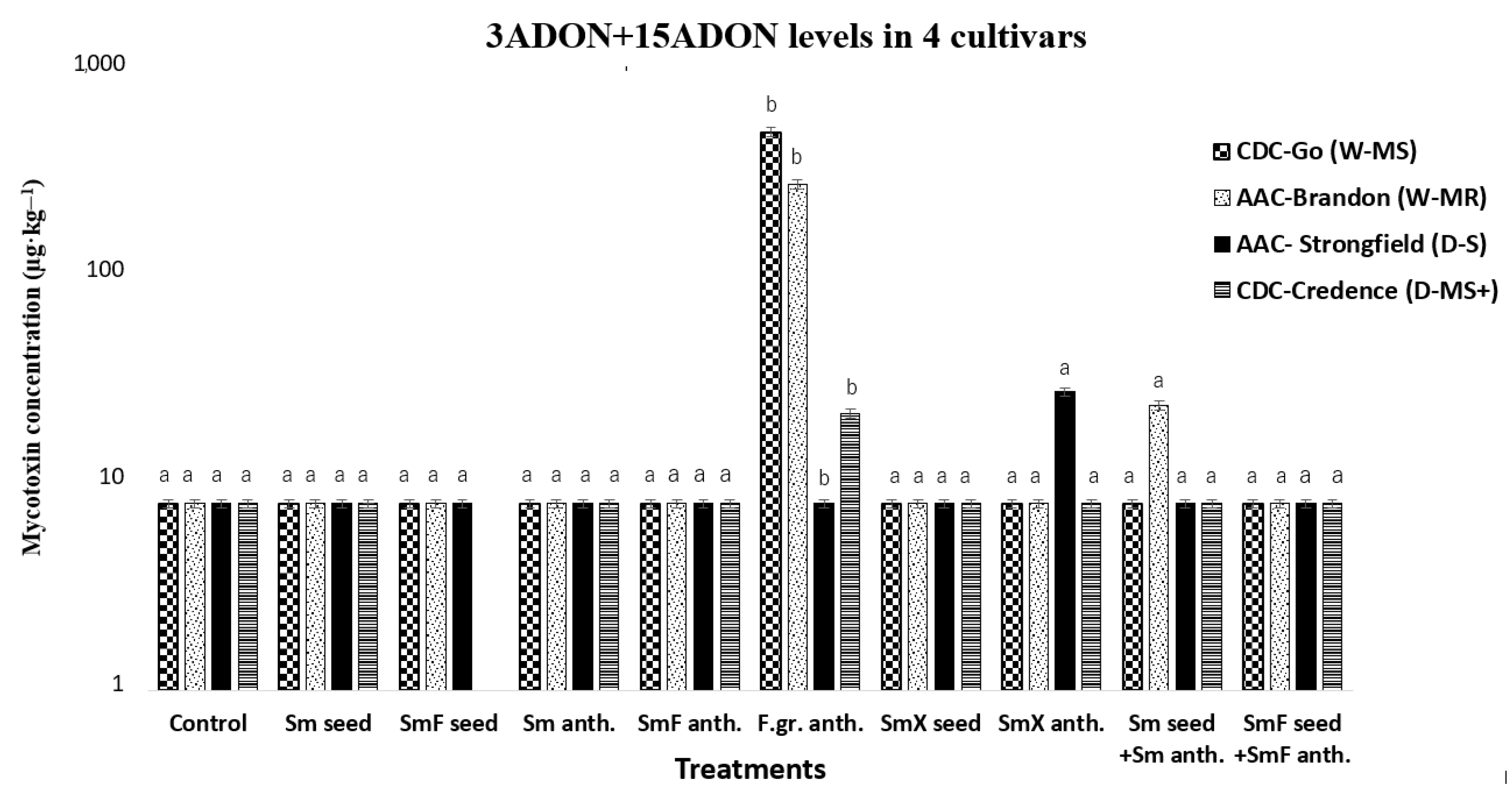
3.3. Acetylated 3ADON+15ADON in Common Wheat and Durum Wheat Cultivars
3.4. Fumonisin B1 (FB1) and Fumonisin B2 (FB2) in Common Wheat and Durum Wheat Cultivars
3.5. Treatment Effect on Mycotoxin Levels in Common Wheat and Durum Wheat Cultivars
3.5.1. Treatment Effect on DON Level
3.5.2. Treatment Effect on 3ADON +15ADON
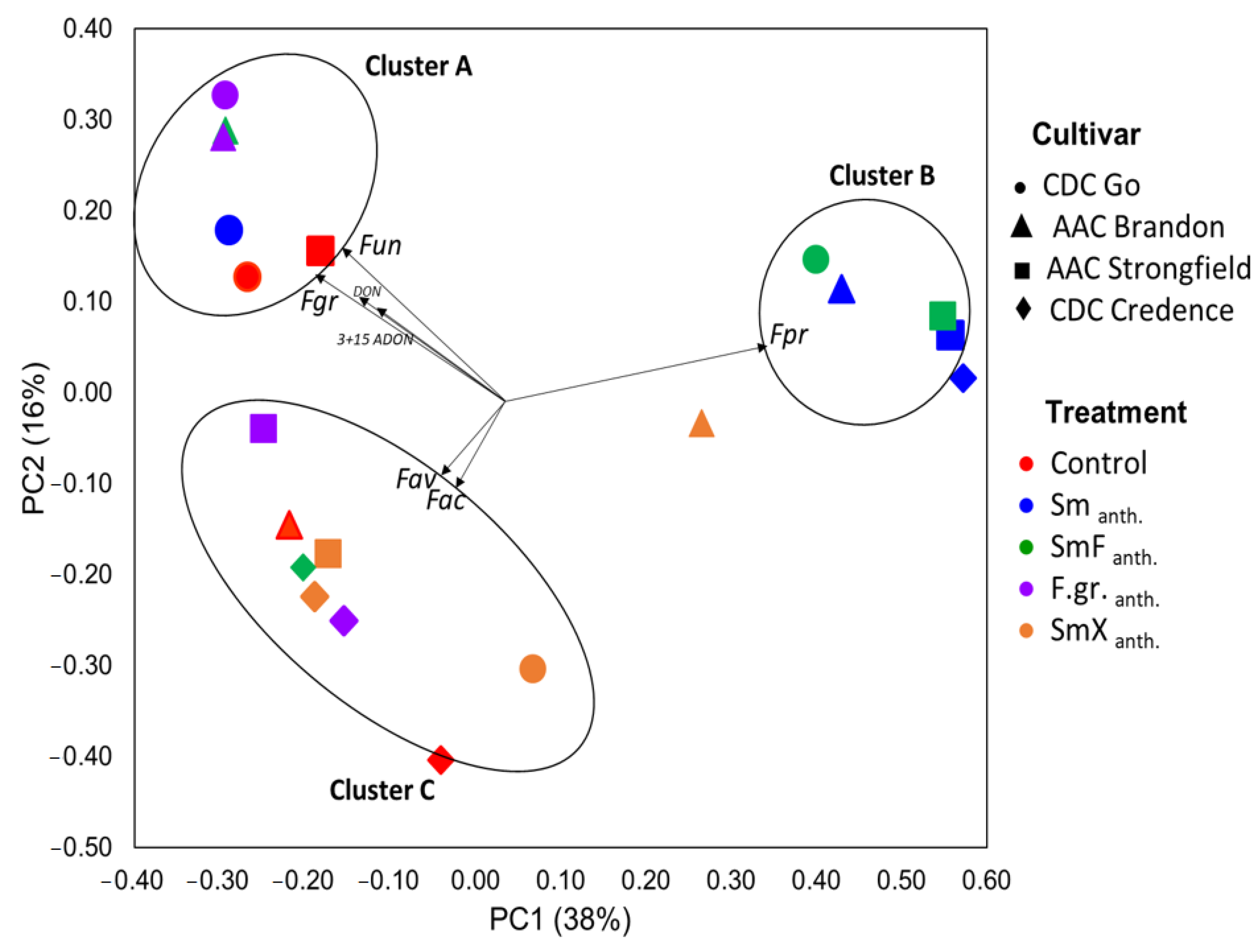
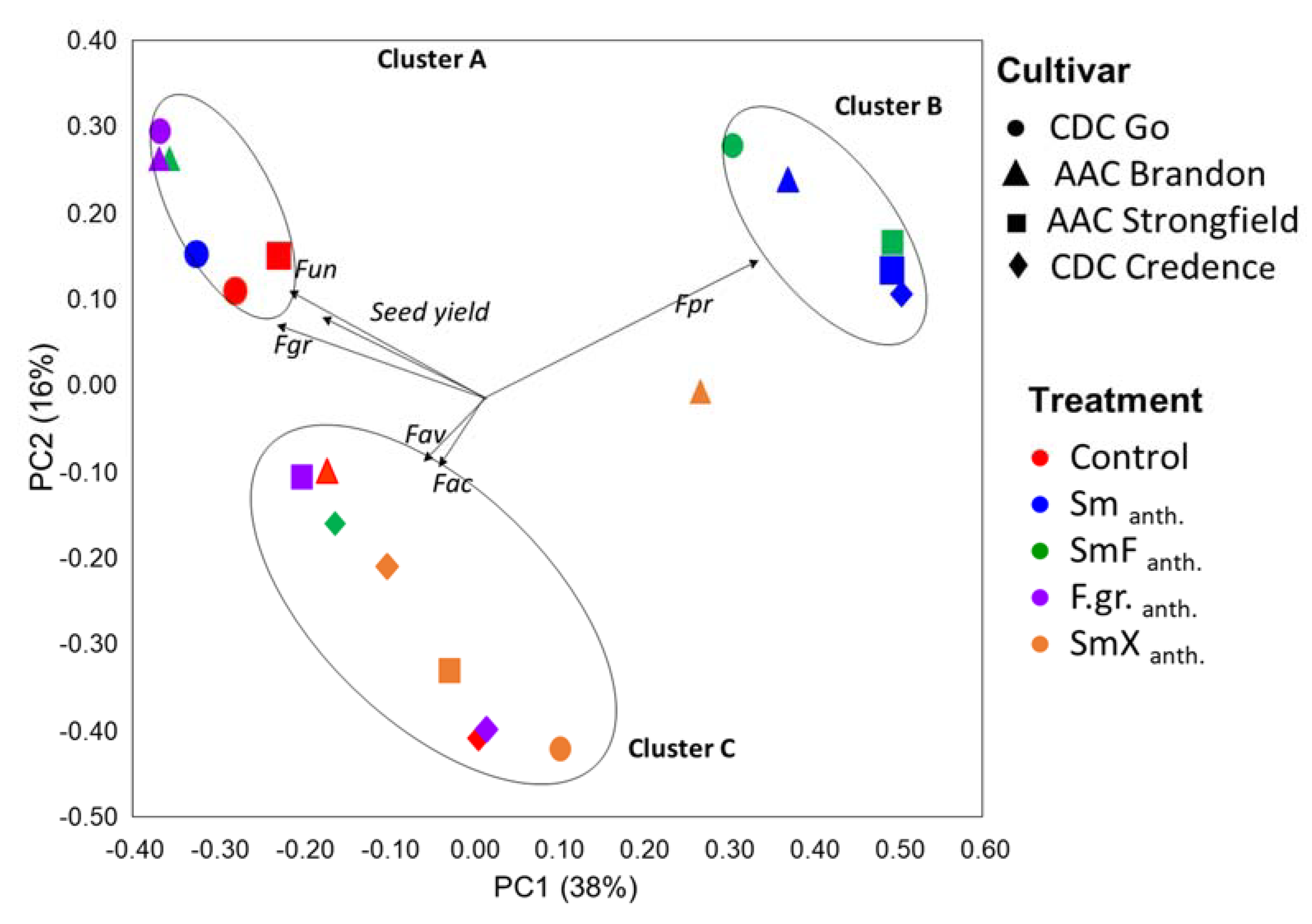
3.6. Fusarium Diversity and Associated Accumulation of Mycotoxins in Seeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stack, R.W. Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; APS Press: St Paul, MN, USA, 2003. [Google Scholar]
- Bai, G.; Shaner, G. Management and Resistance in Wheat and Barley to Fusarium Head Blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium Head Blight of Wheat and Barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Parry, D.W. Review-Fusarium Ear Blight (Scab) in Small Grain Cereals—A Review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Kant, P.; Reinprecht, Y.; Martin, C.J.; Islam, R.; Pauls, K.P. Disease Resistance. In Comprehensive Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 789–805. ISBN 978-0-444-64047-5. [Google Scholar]
- Abbas, A.; Yli-Mattila, T. Biocontrol of Fusarium graminearum, a Causal Agent of Fusarium Head Blight of Wheat, and Deoxynivalenol Accumulation: From In Vitro to In Planta. Toxins 2022, 14, 299. [Google Scholar] [CrossRef] [PubMed]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium Species and Mycotoxins Associated with Head Blight in Small-Grain Cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Proctor, R.H.; Desjardins, A.E.; Mccormick, S.P.; Plattner, R.D.; Alexander, N.J.; Brown, D.W. Genetic Analysis of the Role of Trichothecene and Fumonisin Mycotoxins in the Virulence of Fusarium. Eur. J. Plant Pathol. 2002, 108, 691–698. [Google Scholar] [CrossRef]
- Soyer, J.L.; Hamiot, A.; Ollivier, B.; Balesdent, M.H.; Rouxel, T.; Fudal, I. The APSES Transcription Factor LmStuA Is Required for Sporulation, Pathogenic Development and Effector Gene Expression in Leptosphaeria maculans. Mol. Plant Pathol. 2015, 16, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evolume 2019, 3, 430–439. [Google Scholar] [CrossRef]
- David Miller, J.; Richardson, S.N. Mycotoxins in Canada: A Perspective for 2013. Regul. Gov. Initiat. 2013. Available online: https://scabusa.org/pdfs/Mycotoxins-in-Canada_Sept-13.pdf (accessed on 14 December 2021).
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dweba, C.C.; Figlan, S.; Shimelis, H.A.; Motaung, T.E.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium Head Blight of Wheat: Pathogenesis and Control Strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Mycotoxins. World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/mycotoxins (accessed on 21 November 2022).
- Chen, Y.; Kistler, H.C.; Ma, Z. Annual Review of Phytopathology Fusarium graminearum Trichothecene Mycotoxins: Biosynthesis, Regulation, and Management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef] [Green Version]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium Head Blight, Mycotoxins and Strategies for Their Reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Liu, X.; Li, J. Updating Techniques on Controlling Mycotoxins—A Review. Food Control 2018, 89, 123–132. [Google Scholar] [CrossRef]
- Pirgozliev, S.R.; Edwards, S.G.; Hare, M.C.; Jenkinson, P. Strategies for the Control of Fusarium Head Blight in Cereals. Eur. J. Plant Pathol. 2003, 109, 731–742. [Google Scholar] [CrossRef]
- Shah, L.; Ali, A.; Yahya, M.; Zhu, Y.; Wang, S.; Si, H.; Rahman, H.; Ma, C. Integrated Control of Fusarium Head Blight and Deoxynivalenol Mycotoxin in Wheat. Plant Pathol. 2018, 67, 532–548. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of Action, Human Exposure, and Toxicological Relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Zhang, G.L.; Feng, Y.L.; Song, J.L.; Zhou, X.S. Zearalenone: A Mycotoxin With Different Toxic Effect in Domestic and Laboratory Animals’ Granulosa Cells. Front. Genet. 2018, 9, 667. [Google Scholar] [CrossRef] [Green Version]
- European Union Setting Maximum Levels for Certain Contaminants in Foodstuffs (Text with EEA Relevance). Off. J. Eur. Union 2006, 364, 1–24.
- Guidance for Industry and FDA: Advisory Levels for Deoxynivalenol (DON) in Finished Wheat Products for Human Consumption and Grains and Grain By-Products Used for Animal Feed. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-fda-advisory-levels-deoxynivalenol-don-finished-wheat-products-human (accessed on 21 November 2022).
- Health Canada’s Maximum Levels for Chemical Contaminants in Foods. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/food-safety/chemical-contaminants/maximum-levels-chemical-contaminants-foods.html (accessed on 21 November 2022).
- Haile, J.K.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Clarke, J.M.; Kutcher, H.R.; Steiner, B.; Buerstmayr, H.; Pozniak, C.J. Fusarium Head Blight in Durumwheat: Recent Status, Breeding Directions, and Future Research Prospects. Phytopathology 2019, 109, 1664–1675. [Google Scholar] [CrossRef] [PubMed]
- Thielecke, F.; Nugent, A.P. Contaminants in Grain—A Major Risk for Whole Grain Safety? Nutrients 2018, 10, 1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed Raghda, A.; Jebur Ali, B.; Kang, W.; El-Demerdash, F.M. An Overview of the Major Mycotoxin in Food Products. J. Future Foods 2022, 2, 91–102. [Google Scholar] [CrossRef]
- Gilbert, J.; Tekauz, A. Review: Recent Developments in Research on Fusarium Head Blight of Wheat in Canada. Can. J. Plant Pathol. 2000, 22, 6. [Google Scholar] [CrossRef]
- Ji, F.; He, D.; Olaniran, A.O.; Mokoena, M.P.; Xu, J.; Shi, J. Occurrence, Toxicity, Production and Detection of Fusarium Mycotoxin: A Review. Food Prod. Process. Nutr. 2019, 1, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of Mycotoxins: Tales from Known and Unexplored Worlds. Front. Microbiol. 2016, 7, 561. [Google Scholar] [CrossRef] [Green Version]
- Worth the Wheat: New Durum Wheat Line Shows Intermediate Resistance to Fusarium Head Blight, a First for Canada and the World. 2021. Available online: https://agriculture.canada.ca/en/news-agriculture-and-agri-food-canada/scientific-achievements-agriculture/worth-wheat-new-durum-wheat-line-shows-intermediate-resistance-fusarium-head-blight-first-canada-and (accessed on 15 July 2022).
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A Unified Effort to Fight an Enemy of Wheat and Barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Xiao, X.; Li, T.; Chen, W.; Wang, J.; Fraaije, B.A.; Zhou, M. Impact of Epoxiconazole on Fusarium Head Blight Control, Grain Yield and Deoxynivalenol Accumulation in Wheat. Pestic. Biochem. Physiol. 2018, 152, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, X.; Jiang, J.; Hamada, M.S.; Yin, Y.; Ma, Z. Detection and Dynamics of Different Carbendazim-Resistance Conferring β-Tubulin Variants of Gibberella zeae Collected from Infected Wheat Heads and Rice Stubble in China. Pest Manag. Sci. 2014, 70, 1228–1236. [Google Scholar] [CrossRef]
- Zhou, Z.; Duan, Y.; Zhou, M. Carbendazim-Resistance Associated β 2-Tubulin Substitutions Increase β 2-Tubulin and IDH3 in Fusarium graminearum. Environ. Biol. 2019, 22, 598–614. [Google Scholar] [CrossRef]
- Canada, H. Proposed Maximum Residue Limit PMRL2020-44, Pydiflumetofen. Available online: https://www.canada.ca/en/health-canada/services/consumer-product-safety/pesticides-pest-management/public/consultations/proposed-maximum-residue-limit/2020/pydiflumetofen/document.html (accessed on 28 November 2022).
- Hou, Y.P.; Mao, X.W.; Wang, J.X.; Zhan, S.W.; Zhou, M.G. Sensitivity of Fusarium asiaticum to a Novel Succinate Dehydrogenase Inhibitor Fungicide Pydiflumetofen. Crop Prot. 2017, 96, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Mesterházy, A. Types and Components of Resistance to Fusarium Head Blight of Wheat. Plant Breed. 1995, 114. [Google Scholar] [CrossRef]
- Schroeder, H.W.; Christensen, J.J. Factors Affecting Resistance of Wheat to Scab Caused by Gibberella zeae. Phytopathology 1963, 53, 831–838. [Google Scholar]
- Miller, J.D.; Young, J.C.; Sampson, D.R. Deoxynivalenol and Fusarium Head Blight Resistance in Spring Cereals. J. Phytopathol. 1985, 113, 359–367. [Google Scholar] [CrossRef]
- Snijders, C.H.A. Fusarium Head Blight and Mycotoxin Contamination of Wheat, a Review. Neth. J. Plant Pathol. 1990, 96, 187–198. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium Head Blight From a Microbiome Perspective. Front. Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef]
- Vujanovic, V. Tremellomycetes Yeasts in Kernel Ecological Niche: Early Indicators of Enhanced Competitiveness of Endophytic and Mycoparasitic Symbionts against Wheat Pathobiota. Plants 2021, 10, 905. [Google Scholar] [CrossRef]
- Kim, S.H.; Vujanovic, V. Relationship between Mycoparasites Lifestyles and Biocontrol Behaviors against Fusarium spp. and Mycotoxins Production. Appl. Microbiol. Biotechnol. 2016, 100, 5257–5272. [Google Scholar] [CrossRef]
- Karuppiah, V.; Sun, J.; Li, T.; Vallikkannu, M.; Chen, J. Co-Cultivation of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 Causes Differential Gene Expression and Improvement in the Wheat Growth and Biocontrol Activity. Front. Microbiol. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Harman, G.E. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matarese, F.; Sarrocco, S.; Gruber, S.; Seidl-Seiboth, V.; Vannacci, G. Biocontrol of Fusarium Head Blight: Interactions between Trichoderma and Mycotoxigenic Fusarium. Microbiology 2012, 158, 98–106. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Horwitz, B.A.; Herrera-Estrella, A.; Schmoll, M.; Kenerley, C.M. Trichoderma Research in the Genome Era. Annu. Rev. Phytopathol. 2013, 51, 105–129. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, V.; Goh, Y.K. Sphaerodes mycoparasitica sp. Nov., a New Biotrophic Mycoparasite on Fusarium avenaceum, F. graminearum and F. oxysporum. Mycol. Res. 2009, 113, 1172–1180. [Google Scholar] [CrossRef]
- Goh, Y.K.; Vujanovic, V. Ascospore Germination Patterns Revealed Ascomycetous Biotrophic Mycoparasite Specificity to Fusarium Hosts. Botany 2010, 88, 1033–1043. [Google Scholar] [CrossRef]
- Vujanovic, V.; Goh, Y.K. Qpcr Quantification of Sphaerodes mycoparasitica Biotrophic Mycoparasite Interaction with Fusarium Graminearum: In Vitro and in Planta Assays. Arch. Microbiol. 2012, 194, 707–717. [Google Scholar] [CrossRef]
- Vujanovic, V.; Kim, S.H. Adaptability of Mitosporic Stage in Sphaerodes mycoparasitica towards Its Mycoparasitic-Polyphagous Lifestyle. Mycologia 2017, 109, 701–709. [Google Scholar] [CrossRef]
- Powell, A.J.; Vujanovic, V. Evolution of Fusarium Head Blight Management in Wheat: Scientific Perspectives on Biological Control Agents and Crop Genotypes Protocooperation. Appl. Sci. 2021, 11, 8960. [Google Scholar] [CrossRef]
- Beeck, M.O.D.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and Validation of Some ITS Primer Pairs Useful for Fungal Metabarcoding Studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [Green Version]
- de la Cuesta-Zuluaga, J.; Escobar, J.S. Considerations For Optimizing Microbiome Analysis Using a Marker Gene. Front. Nutr. 2016, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pẽa, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, Version 7 for Windows; Wild Bluenerry Media: Corvallis, OR, USA, 2016. [Google Scholar]
- Nualkaw, K.; Poapolathep, S.; Zhang, Z.; Zhang, Q.; Giorgi, M.; Li, P.; Logrieco, A.F.; Poapolathep, A. Simultaneous Determination of Multiple Mycotoxins in Swine, Poultry and Dairy Feeds Using Ultra High Performance Liquid Chromatography-Tandem Mass Spectrometry. Toxins 2020, 12, 253. [Google Scholar] [CrossRef]
- Kim, S.H.; Vujanovic, V. Biodegradation and Biodetoxification of Fusarium Mycotoxins by Sphaerodes mycoparasitica. AMB Express 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.; Tekauz, A. Insects and Diseases PS&C Prairie Soils & Crops Journal Strategies for Management of Fusarium Head Blight (FHB) in Cereals. Prairie Soils Crops J. 2011, 4, 97–104. [Google Scholar]
- Tittlemier, S.A.; Blagden, R.; Chan, J.; Gaba, D.; Mckendry, T.; Pleskach, K.; Roscoe, M. Fusarium and Alternaria Mycotoxins Present in Canadian Wheat and Durum Harvest Samples. Can. J. Plant Pathol. 2019, 41, 403–414. [Google Scholar] [CrossRef]
- Podgórska-Kryszczuk, I.; Solarska, E.; Kordowska-Wiater, M. Reduction of the Fusarium Mycotoxins: Deoxynivalenol, Nivalenol and Zearalenone by Selected Non-Conventional Yeast Strains in Wheat Grains and Bread. Molecules 2022, 27, 1578. [Google Scholar] [CrossRef] [PubMed]
- CRC. Falling Number and Deoxynivalenol (DON) as Potential Official Grain Grading Factors; Canadian Grain Commission: Winnipeg, MB, Canada, 2019; pp. 1–15. Available online: https://grainscanada.gc.ca/en/about-us/consultations/2019/falling-number-don.html (accessed on 4 February 2022).
- Xue, A.G.; Chen, Y.; Voldeng, H.D.; Fedak, G.; Savard, M.E.; Längle, T.; Zhang, J.; Harman, G.E. Concentration and Cultivar Effects on Efficacy of CLO-1 Biofungicide in Controlling Fusarium Head Blight of Wheat. Biol. Control 2014, 73. [Google Scholar] [CrossRef]
- Okorski, A.; Milewska, A.; Pszczółkowska, A.; Karpiesiuk, K.; Kozera, W.; Dąbrowska, J.A.; Radwińska, J. Prevalence of Fusarium Fungi and Deoxynivalenol Levels in Winter Wheat Grain in Different Climatic Regions of Poland. Toxins 2022, 14, 102. [Google Scholar] [CrossRef]
- Ruan, Y.; Babonich, R.; Clarke, J.M.; Hucl, P.J.; Clarke, F.R.; Knox, R.E.; Bokore, F.E.; Pozniak, C.J. Differential Reaction of Hexaploid and Tetraploid Wheat to Fusarium graminearum Chemotypes in a Controlled Environment. Can. J. Plant Pathol. 2021, 43, 760–768. [Google Scholar] [CrossRef]
- Walkowiak, S.; Rowland, O.; Rodrigue, N.; Subramaniam, R. Whole Genome Sequencing and Comparative Genomics of Closely Related Fusarium Head Blight Fungi: Fusarium graminearum, F. meridionale and F. asiaticum. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walkowiak, S.; Bonner, C.T.; Wang, L.; Blackwell, B.; Rowland, O.; Subramaniam, R. Intraspecies Interaction of Fusarium graminearum Contributes to Reduced Toxin Production and Virulence. Mol. Plant. Microbe Interact. 2015, 28, 1256–1267. [Google Scholar] [CrossRef] [Green Version]
- Alexander, N.J.; McCormick, S.P.; Waalwijk, C.; van der Lee, T.; Proctor, R.H. The Genetic Basis for 3-ADON and 15-ADON Trichothecene Chemotypes in Fusarium. Fungal Genet. Biol. 2011, 48, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, P.; Liu, Z.; Liu, S.; Yao, L.; Liu, Y.; Wu, Y.; Gong, Z. Natural Occurrence of Deoxynivalenol and Its Acetylated Derivatives in Chinese Maize and Wheat Collected in 2017. Toxins 2020, 12, 200. [Google Scholar] [CrossRef] [Green Version]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-clauw, M.; Oswald, I.P. Toxicity of Deoxynivalenol and Its Acetylated Derivatives on the Intestine: Differential Effects on Morphology, Barrier Function, Tight Junction Proteins, and Mitogen-Activated Protein Kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef]
- Schmale, D.G.; Munkvold, G.P. Mycotoxins in Crops: A Threat to Human and Domestic Animal Health. Plant Health Instr. 2014. [Google Scholar] [CrossRef]
- Marin, S.; Sanchis, V.; Magan, N. Water Activity, Temperature, and PH Effects on Growth of Fusarium Moniliforme and Fusarium proliferatum Isolates from Maize. Can. J. Microbiol. 1995, 41, 12. [Google Scholar] [CrossRef] [PubMed]
- Gdanetz, K.; Trail, F. The Wheat Microbiome under Four Management Strategies, and Potential for Endophytes in Disease Protection. Phytobiomes J. 2017, 1, 158–1683. [Google Scholar] [CrossRef] [Green Version]
- Cendoya, E.; Chiotta, M.L.; Zachetti, V.; Chulze, S.N.; Ramirez, M.L. Fumonisins and Fumonisin-Producing Fusarium Occurrence in Wheat and Wheat by Products: A Review. J. Cereal Sci. 2018, 80, 158–166. [Google Scholar] [CrossRef]
- Guo, Z.; Pfohl, K.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Fumonisin B1 and Beauvericin Accumulation in Wheat Kernels after Seed-Borne Infection with Fusarium proliferatum. Agric. Food Sci. 2016, 25, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Cendoya, E.; Farnochi, M.C.; Chulze, S.N.; Ramirez, M.L. Two-Dimensional Environmental Profiles of Growth and Fumonisin Production by Fusarium proliferatum on a Wheat-Based Substrate. Int. J. Food Microbiol. 2014, 182–183, 9–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Description |
|---|---|
| 1. Control | No biocontrol agents (BCAs) or any other treatment |
| 2. Smseed | BCA- S. mycoparasitica 2220-01 (main BCA strain used in all studies) applied on seed |
| 3. SmFseed | S. mycoparasitica 2220-01 + Fungicide on seed (1/2 of the effective dose of fungicide- Prosaro was used) |
| 4. Smanth. | S. mycoparasitica 2220-01 applied at anthesis |
| 5. SmFanth. | S. mycoparasitica 2220-01+ Fungicide at anthesis |
| 6. F.granth | A mixture of F. graminearum 3ADON strains applied at anthesis |
| 7. SmXseed | S. mycoparasitica SMCD 2220-01+ SMCD 2220-02(5) mixture of BCA beneficial strains, applied on seed |
| 8. SmXanth. | S. mycoparasitica SMCD 2220-01 + SMCD 2220-02(5) applied at anthesis |
| 9.Smseed + Smanth. | S. mycoparasitica 2220-01 seed + S.mycoparasitica 2220-01 at anthesis |
| 10. SmFseed + SmFanth. | S. mycoparasitica 2220-01 + Fungicide seed and S. mycoparasitica 2220-01+ Fungicide at anthesis. |
| Treatments | CDC Go W-MS | AAC Brandon W-MR | AAC Strongfield D-S | CDC Credence D-MS+ | |
|---|---|---|---|---|---|
| 1 | Control | 26 | <16 | 19.8 | 19.5 |
| 2 | Sm seed | <16 | 21.7 | 20.2 | 21.4 |
| 3 | SmF seed | <16 | 23.1 | <16 | NR |
| 4 | Sm anth. | 17.2 | <16 | <16 | <16 |
| 5 | SmF anth. | <16 | 23.9 | <16 | <16 |
| 6 | F.gr.anth. | 29720 | 10150 | 1052 | 564.9 |
| 7 | SmX seed | 29.4 | 30 | <16 | <16 |
| 8 | SmX anth. | 227.5 | <16 | 116.7 | <16 |
| 9 | Sm seed + Sm anth. | 19.3 | 592.3 | <16 | 21.7 |
| 10 | SmF seed + SmF anth. | <16 | <16 | <16 | 17.9 |
| Total DON conc. in each cultivar | 30071.4 | 10873 | 1256.7 | 677.4 |
| Treatments | CDC Go W-MS | AAC Brandon W-MR | AAC Strongfield D-S | CDC Credence D-MS+ | |
|---|---|---|---|---|---|
| 1 | Control | <16 | <16 | <16 | <16 |
| 2 | Sm seed | <16 | <16 | <16 | <16 |
| 3 | SmF seed | <16 | <16 | <16 | NR |
| 4 | Sm anth. | <16 | <16 | <16 | <16 |
| 5 | SmF anth. | <16 | <16 | <16 | <16 |
| 6 | F.gr. anth. | 499.4 | 278.8 | <16 | 21.9 |
| 7 | SmX seed | <16 | <16 | <16 | <16 |
| 8 | SmX anth. | <16 | <16 | 27.7 | <16 |
| 9 | Sm seed + Sm anth. | <16 | 23.8 | <16 | <16 |
| 10 | SmF seed + SmF anth. | <16 | <16 | <16 | <16 |
| Total 3 + 15ADON conc. in each cultivar | 571.4 | 366.6 | 99.7 | 85.9 |
| Treatments | CDC Go W-MS | AAC Brandon W-MR | AAC Strongfield D-S | CDC Credence D-MS+ | |
|---|---|---|---|---|---|
| 1 | Control | <1 | <1 | 2.9 | <1 |
| 2 | Sm seed | 2.03 | 4.8 | <1 | 3.9 |
| 3 | SmF seed | 22.8 | <1 | 163.5 | NR |
| 4 | Sm anth. | <1 | <1 | 2.4 | <1 |
| 5 | SmF anth. | 4.13 | 25.3 | <1 | <1 |
| 6 | F.gr.anth. | 2.1 | <1 | <1 | 23.2 |
| 7 | SmX seed | 338 | <1 | 7.79 | <1 |
| 8 | SmX anth. | 2.5 | 11.2 | <1 | 8.11 |
| 9 | Sm seed + Sm anth. | 5.4 | 268.1 | <1 | <1 |
| 10 | SmF seed + SmF anth. | <1 | 163.6 | 42.1 | <1 |
| Total FB1 conc. in each cultivar | 378.5 | 475.5 | 224.2 | 38.21 |
| Treatments | CDC Go W-MS | AAC Brandon W-MR | AAC Strongfield D-S | CDC Credence D-S+ | |
|---|---|---|---|---|---|
| 1 | Control | <1 | <1 | <1 | <1 |
| 2 | Sm seed | <1 | <1 | <1 | <1 |
| 3 | SmF seed | 2.3 | <1 | 11.8 | NR |
| 4 | Sm anth. | <1 | <1 | <1 | <1 |
| 5 | SmF anth. | <1 | 4.04 | <1 | <1 |
| 6 | F. gr.anth. | <1 | <1 | <1 | 4.1 |
| 7 | SmX seed | 137.7 | <1 | <1 | 1 |
| 8 | SmX anth. | <1 | 2.08 | <1 | <1 |
| 9 | Sm seed + Sm anth. | <1 | 72.8 | <1 | <1 |
| 10 | SmFseed + SmF anth. | <1 | 26.6 | <1 | <1 |
| Total FB2 conc. in each cultivar | 144 | 108.5 | 16.3 | 8.1 |
| Fusarium | |||
|---|---|---|---|
| Source | d.f | F | p |
| Cultivar | 3 | 0.95 | 0.51 |
| Treatment | 4 | 0.19 | 0.08 |
| Fusarium | |||
|---|---|---|---|
| Source | d.f | F | p |
| Cultivar | 3 | 0.95 | 0.51 |
| Treatment | 4 | 1.5 | 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Powell, A.J.; Kim, S.H.; Cordero, J.; Vujanovic, V. Protocooperative Effect of Sphaerodes mycoparasitica Biocontrol and Crop Genotypes on FHB Mycotoxin Reduction in Bread and Durum Wheat Grains Intended for Human and Animal Consumption. Microorganisms 2023, 11, 159. https://doi.org/10.3390/microorganisms11010159
Powell AJ, Kim SH, Cordero J, Vujanovic V. Protocooperative Effect of Sphaerodes mycoparasitica Biocontrol and Crop Genotypes on FHB Mycotoxin Reduction in Bread and Durum Wheat Grains Intended for Human and Animal Consumption. Microorganisms. 2023; 11(1):159. https://doi.org/10.3390/microorganisms11010159
Chicago/Turabian StylePowell, Antonia J., Seon Hwa Kim, Jorge Cordero, and Vladimir Vujanovic. 2023. "Protocooperative Effect of Sphaerodes mycoparasitica Biocontrol and Crop Genotypes on FHB Mycotoxin Reduction in Bread and Durum Wheat Grains Intended for Human and Animal Consumption" Microorganisms 11, no. 1: 159. https://doi.org/10.3390/microorganisms11010159
APA StylePowell, A. J., Kim, S. H., Cordero, J., & Vujanovic, V. (2023). Protocooperative Effect of Sphaerodes mycoparasitica Biocontrol and Crop Genotypes on FHB Mycotoxin Reduction in Bread and Durum Wheat Grains Intended for Human and Animal Consumption. Microorganisms, 11(1), 159. https://doi.org/10.3390/microorganisms11010159