
Interaction between Porcine Alveolar Macrophage-Tang Cells and Streptococcus suis Strains of Different Virulence: Phagocytosis and Apoptosis


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Cell Culture
2.3. PAM-Tang Adhesion Assays
2.4. Cytotoxicity Assay
2.5. Transmission Electron Microscopy
2.6. Confocal Microscopy
2.7. Cytokine Analysis
2.8. Live-Cell Imaging
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
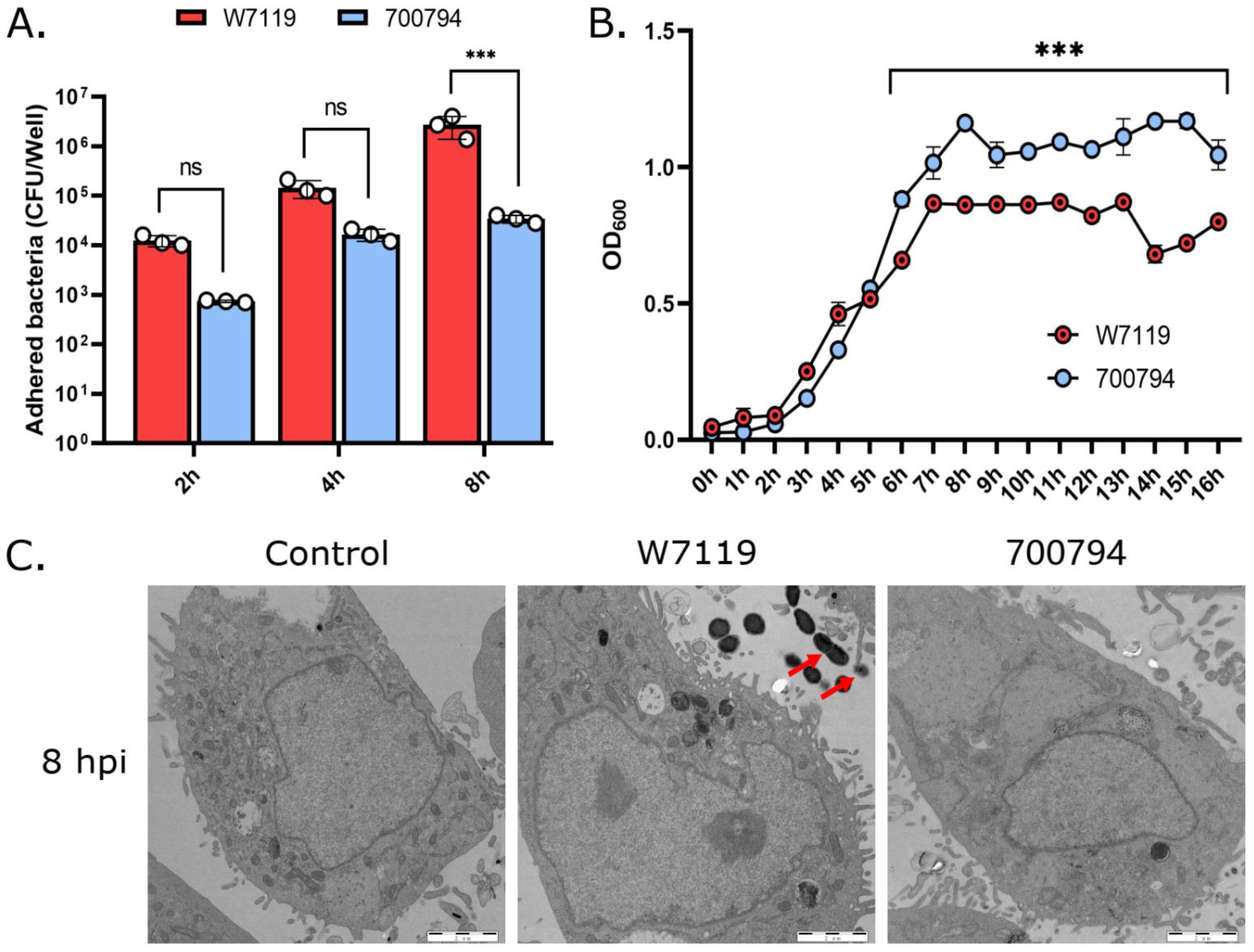
3.1. Avirulent S. suis Presented Higher Adhesion to PAM-Tang Cells
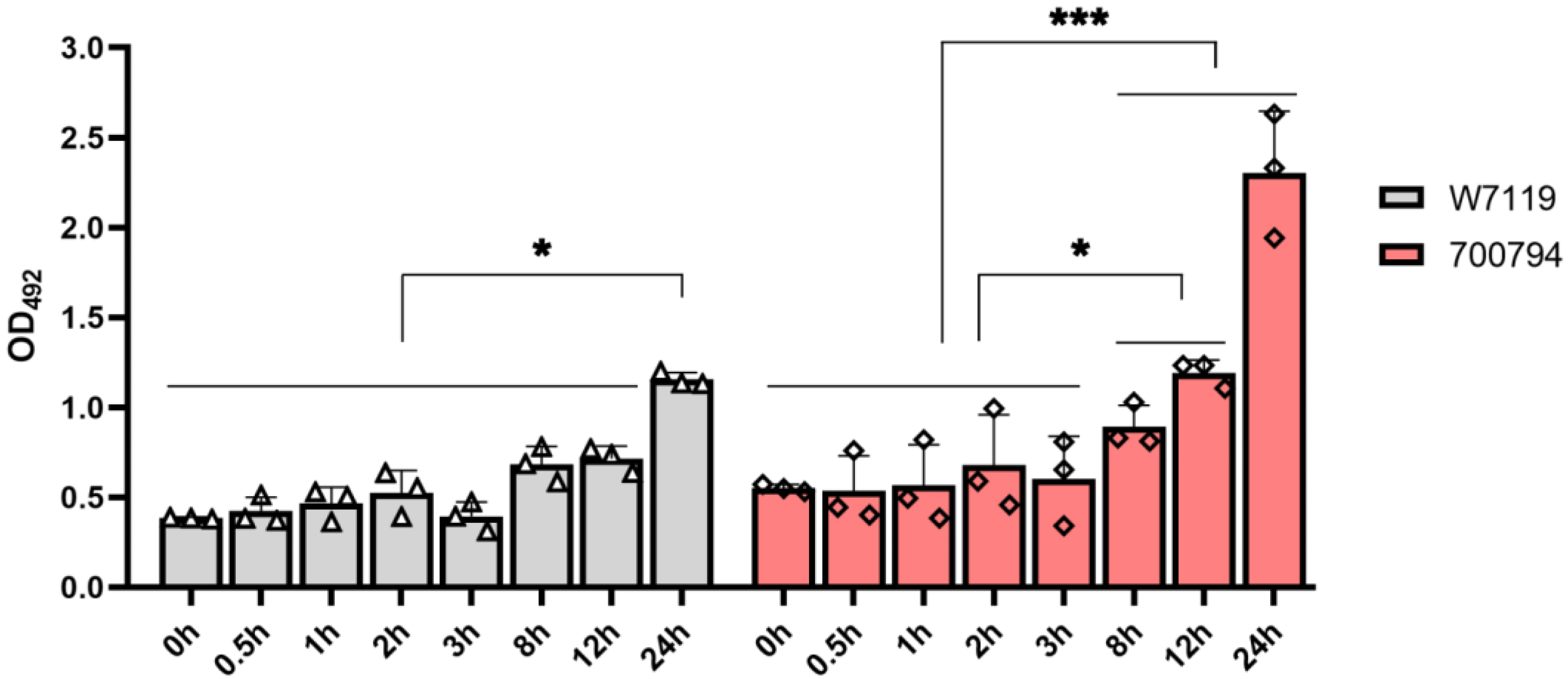
3.2. S. suis Virulent Strain Has Higher Cytotoxic to PAM-Tang Cells
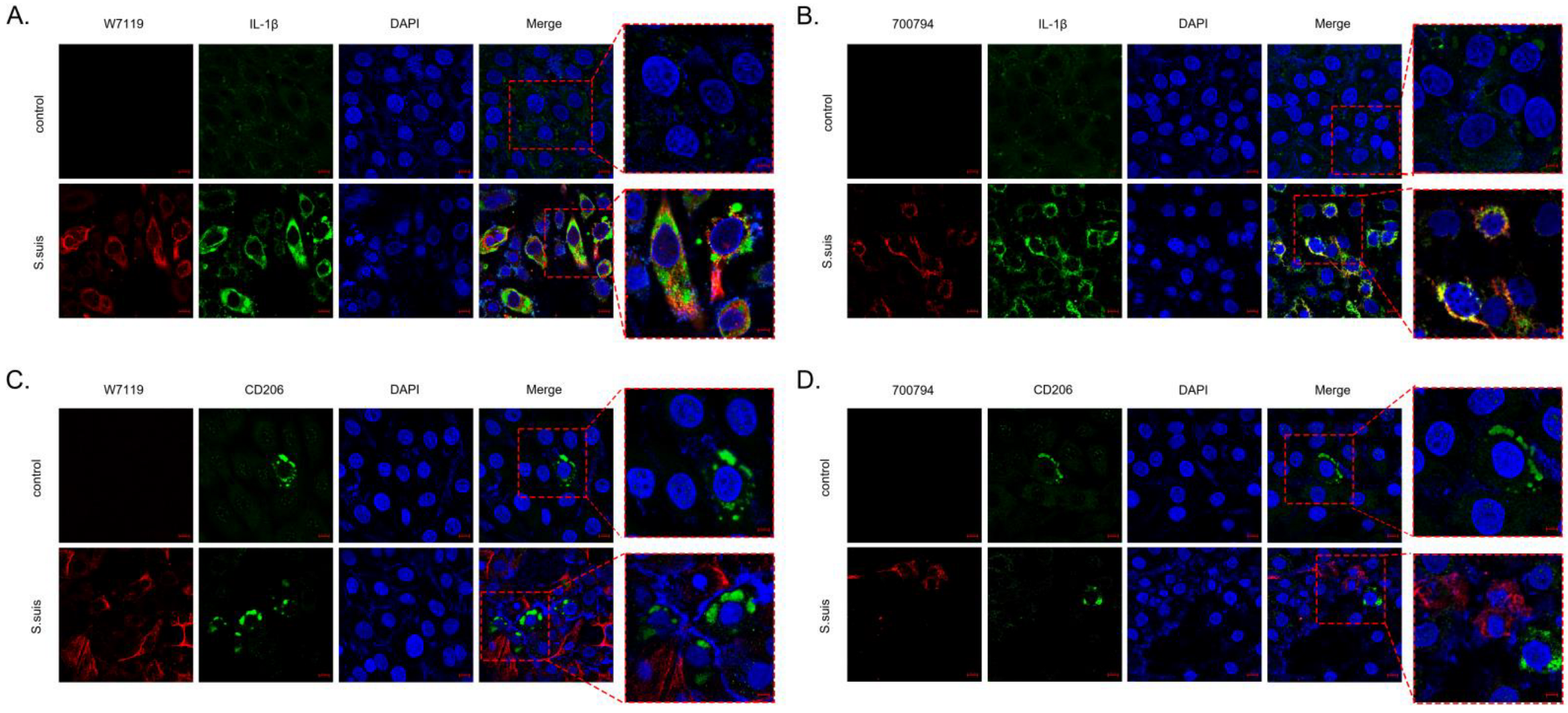
3.3. S. suis Induces PAM-Tang Polarization towards Pro-Inflammatory M1 Macrophages
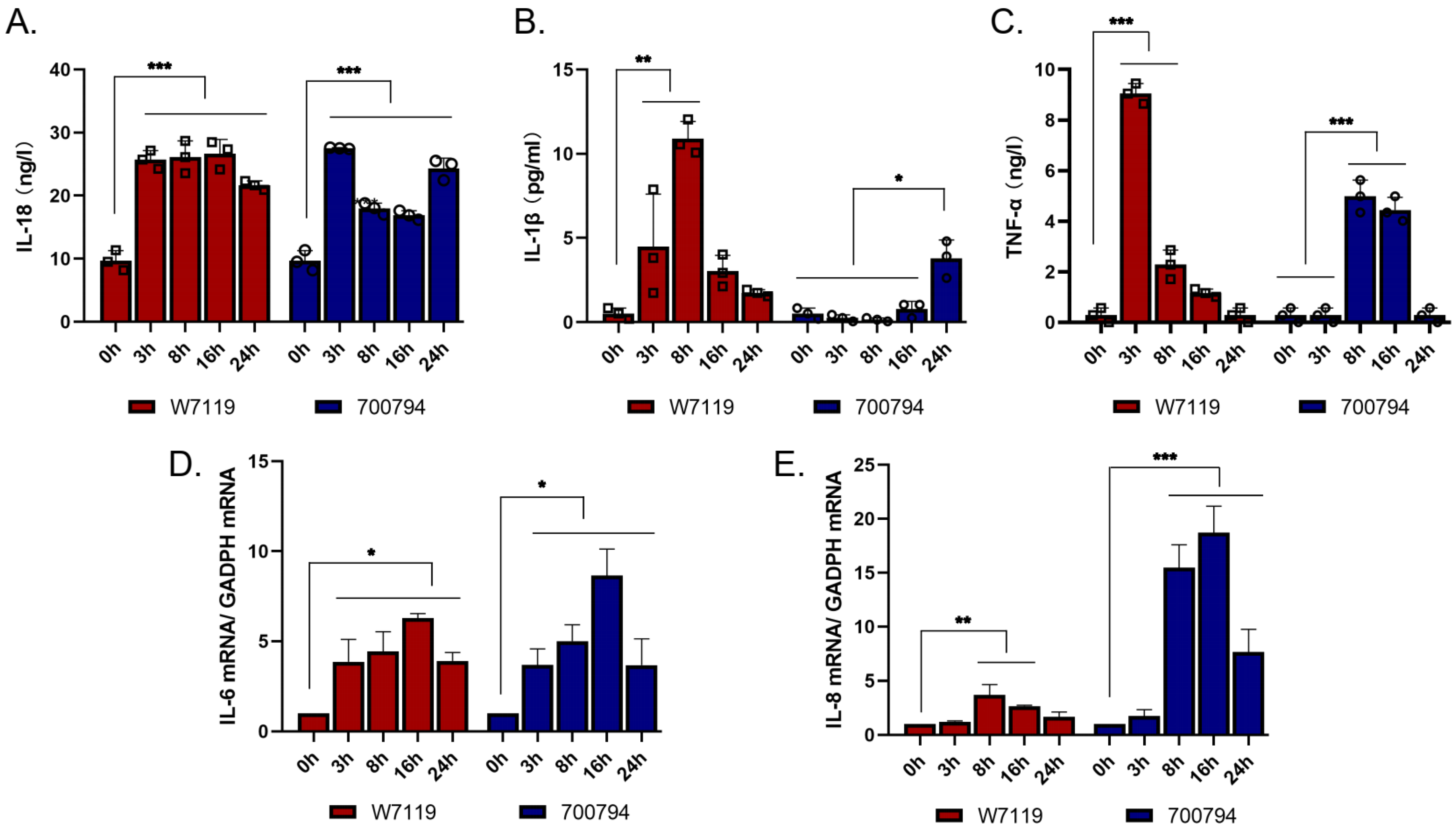
3.4. S. suis Induces PAM-Tang Cells to Secrete Pro-Inflammatory Cytokines
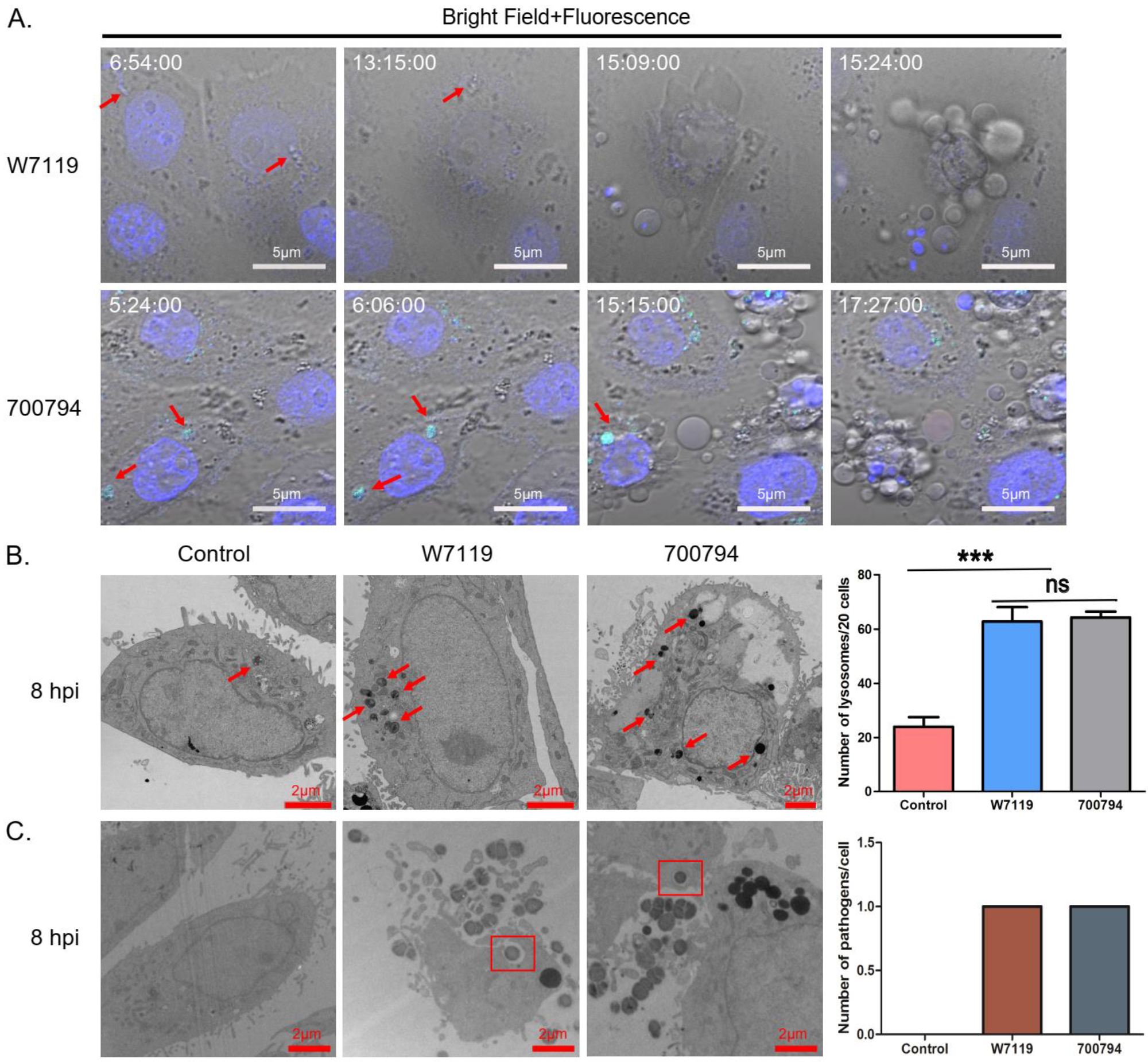
3.5. PAM-Tang Phagocytosis of S. suis
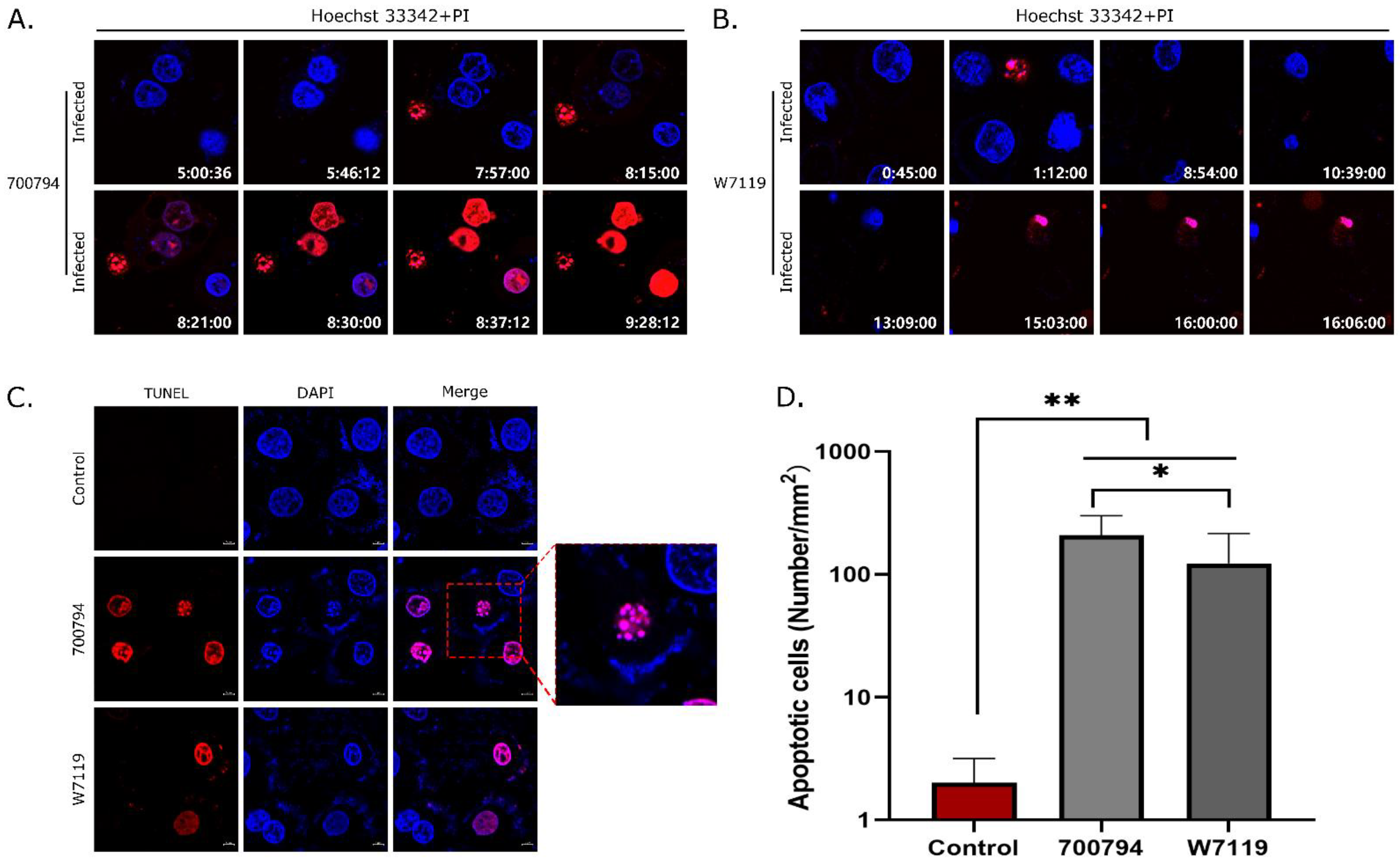
3.6. S. suis Induces Apoptosis in PAM-Tang Cells
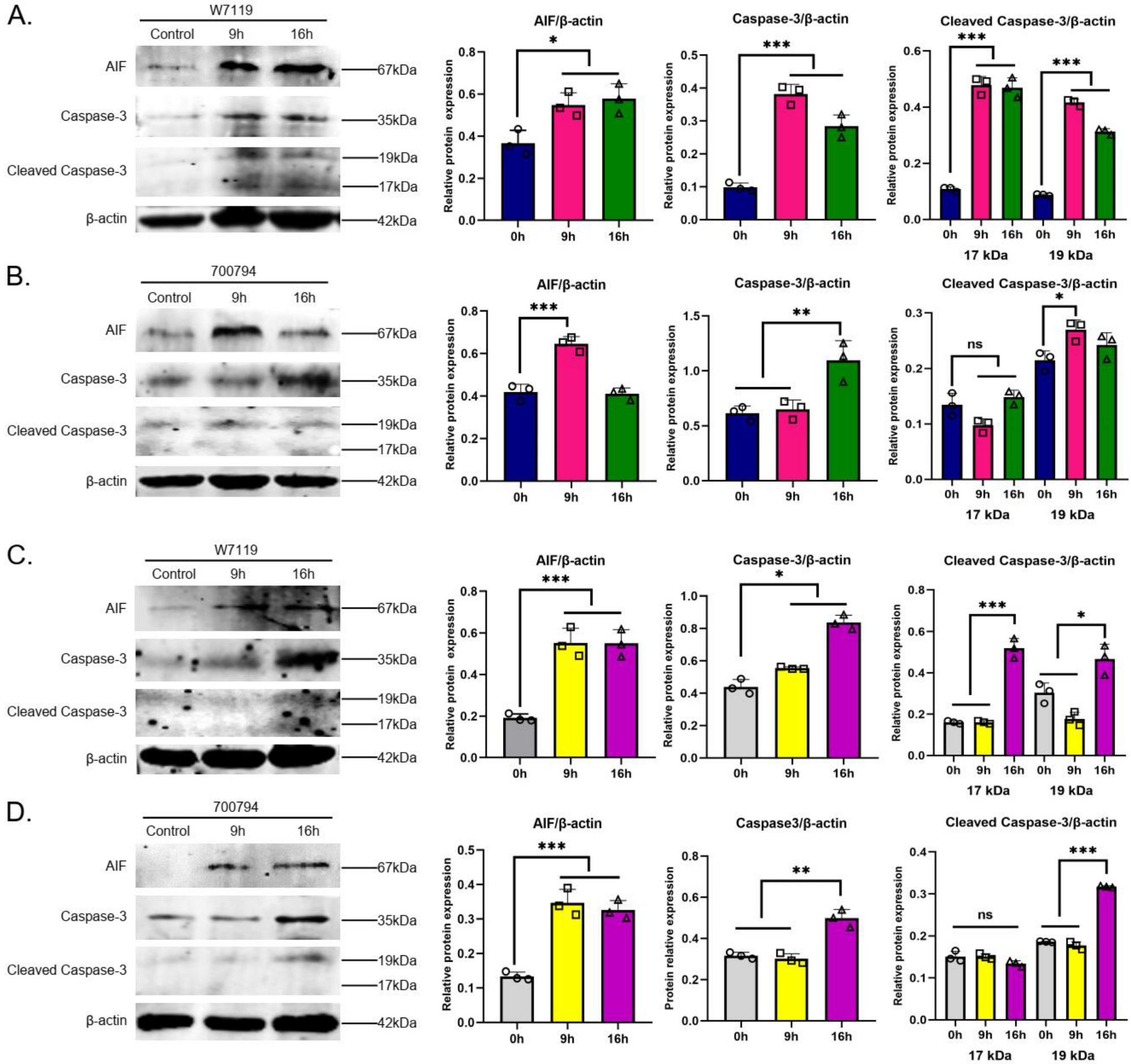
3.7. S. suis Induces Caspase- and p53-Dependent Apoptosis in PAM Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huang, W.; Wang, M.; Hao, H.; Yang, R.; Xie, J.; Su, J.; Lin, M.; Cui, Y.; Jiang, Y. Genomic epidemiological investigation of a Streptococcus suis outbreak in Guangxi, China, 2016. Infect. Genet. Evol. 2019, 68, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, C.; Varaldo, P.E.; Facinelli, B. Streptococcus suis, an Emerging Drug-Resistant Animal and Human Pathogen. Front. Microbiol. 2011, 2, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Ji, S.; Liu, Z.; Lan, R.; Huang, Y.; Bai, X.; Gottschalk, M.; Xu, J. Eight Novel Capsular Polysaccharide Synthesis Gene Loci Identified in Nontypeable Streptococcus suis Isolates. Appl. Environ. Microbiol. 2015, 81, 4111–4119. [Google Scholar] [CrossRef] [Green Version]
- Bojarska, A.; Janas, K.; Pejsak, Z.; Otulak-Koziel, K.; Garbaczewska, G.; Hryniewicz, W.; Sadowy, E. Diversity of serotypes and new cps loci variants among Streptococcus suis isolates from pigs in Poland and Belarus. Vet. Microbiol. 2020, 240, 108534. [Google Scholar] [CrossRef]
- Huan, H.; Jiang, L.; Tang, L.; Wang, Y.; Guo, S. Isolation and characterization of Streptococcus suis strains from swine in Jiangsu province, China. J. Appl. Microbiol. 2020, 128, 1606–1612. [Google Scholar] [CrossRef]
- Tan, M.F.; Tan, J.; Zeng, Y.B.; Li, H.Q.; Yang, Q.; Zhou, R. Antimicrobial resistance phenotypes and genotypes of Streptococcus suis isolated from clinically healthy pigs from 2017 to 2019 in Jiangxi Province, China. J. Appl. Microbiol. 2020, 130, 797–806. [Google Scholar] [CrossRef]
- Matajira, C.E.C.; Moreno, L.Z.; Poor, A.P.; Gomes, V.T.M.; Dalmutt, A.C.; Parra, B.M.; Oliveira, C.H.; Barbosa, M.R.F.; Sato, M.I.Z.; Calderaro, F.F.; et al. Streptococcus suis in Brazil: Genotypic, Virulence, and Resistance Profiling of Strains Isolated from Pigs between 2001 and 2016. Pathogens 2019, 9, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Lyu, C.; Duan, G.; Meng, F.; Yang, Y.; Yu, Y.; He, X.; Wang, Z.; Gottschalk, M.; Li, G.; et al. Streptococcus suis Serotype 2 Infection Causes Host Immunomodulation through Induction of Thymic Atrophy. Infect. Immun. 2020, 88, e00950-19. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M.; Ericsson, A.C. Function of Macrophages in Disease: Current Understanding on Molecular Mechanisms. Front. Immunol. 2021, 12, 620510. [Google Scholar] [CrossRef]
- Verdeguer, F.; Aouadi, M. Macrophage heterogeneity and energy metabolism. Exp. Cell Res. 2017, 360, 35–40. [Google Scholar] [CrossRef]
- Mathieu-Denoncourt, A.; Letendre, C.; Auger, J.P.; Segura, M.; Aragon, V.; Lacouture, S.; Gottschalk, M. Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies. Pathogens 2018, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Gottschalk, M. Streptococcus suis interactions with the murine macrophage cell line J774: Adhesion and cytotoxicity. Infect. Immun. 2002, 70, 4312–4322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Greeff, A.; Benga, L.; Wichgers Schreur, P.J.; Valentin-Weigand, P.; Rebel, J.M.; Smith, H.E. Involvement of NF-kappaB and MAP-kinases in the transcriptional response of alveolar macrophages to Streptococcus suis. Vet. Microbiol. 2010, 141, 59–67. [Google Scholar] [CrossRef]
- Tanabe, S.; Gottschalk, M.; Grenier, D. Hemoglobin and Streptococcus suis cell wall act in synergy to potentiate the inflammatory response of monocyte-derived macrophages. Innate Immun. 2008, 14, 357–363. [Google Scholar] [CrossRef]
- Segura, M.; Gottschalk, M.; Olivier, M. Encapsulated Streptococcus suis inhibits activation of signaling pathways involved in phagocytosis. Infect. Immun. 2004, 72, 5322–5330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.Y.; Liu, Y.G.; Li, L.; Wang, G.; Wang, H.M.; Zhang, H.L.; Zhao, S.F.; Gao, J.C.; An, T.Q.; Tian, Z.J.; et al. Porcine alveolar macrophage CD163 abundance is a pivotal switch for porcine reproductive and respiratory syndrome virus infection. Oncotarget 2018, 9, 12174–12185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- Vanier, G.; Segura, M.; Friedl, P.; Lacouture, S.; Gottschalk, M. Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infect. Immun. 2004, 72, 1441–1449. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gagnon, C.A.; Savard, C.; Music, N.; Srednik, M.; Segura, M.; Lachance, C.; Bellehumeur, C.; Gottschalk, M. Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells. Infect. Immun. 2013, 81, 4498–4508. [Google Scholar] [CrossRef] [Green Version]
- Imori, P.F.; Passaglia, J.; Souza, R.A.; Rocha, L.B.; Falcao, J.P. Virulence-related genes, adhesion and invasion of some Yersinia enterocolitica-like strains suggests its pathogenic potential. Microb. Pathog. 2017, 104, 72–77. [Google Scholar] [CrossRef]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- Hartenstein, V.; Martinez, P. Phagocytosis in cellular defense and nutrition: A food-centered approach to the evolution of macrophages. Cell Tissue Res. 2019, 377, 527–547. [Google Scholar] [CrossRef]
- Krantic, S.; Mechawar, N.; Reix, S.; Quirion, R. Apoptosis-inducing factor: A matter of neuron life and death. Prog. Neurobiol. 2007, 81, 179–196. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.M.; Norton, P.; Haverson, K.; Leigh, J.; Bailey, M. Interactions between Streptococcus suis serotype 2 and cells of the myeloid lineage in the palatine tonsil of the pig. Vet. Immunol. Immunopathol. 2007, 117, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Sagong, M.; Park, C.K.; Kim, S.H.; Lee, K.K.; Lee, O.S.; Lee du, S.; Cha, S.Y.; Lee, C. Human telomerase reverse transcriptase-immortalized porcine monomyeloid cell lines for the production of porcine reproductive and respiratory syndrome virus. J. Virol. Methods 2012, 179, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Weingartl, H.M.; Sabara, M.; Pasick, J.; van Moorlehem, E.; Babiuk, L. Continuous porcine cell lines developed from alveolar macrophages: Partial characterization and virus susceptibility. J. Virol. Methods 2002, 104, 203–216. [Google Scholar] [CrossRef]
- Olvera, A.; Ballester, M.; Nofrarías, M.; Sibila, M.; Aragon, V. Differences in phagocytosis susceptibility in Haemophilus parasuis strains. Vet. Res. 2009, 40, 24. [Google Scholar] [CrossRef] [Green Version]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef]
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis Virulence Factors: Are They All Really Critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.G.; Lacouture, S.; Dubreuil, J.D. Characterization of Streptococcus suis capsular type 2 haemolysin. Microbiology 1995, 141 Pt 1, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, A.A.; Loeffen, P.L.; van den Berg, A.J.; Storm, P.K. Identification, purification, and characterization of a thiol-activated hemolysin (suilysin) of Streptococcus suis. Infect. Immun. 1994, 62, 1742–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Wang, H.; Weng, C.; Jiang, H.; Chen, J. Caspase 3/GSDME-dependent pyroptosis contributes to chemotherapy drug-induced nephrotoxicity. Cell Death Dis. 2021, 12, 186. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Vadeboncoeur, N.; Gottschalk, M. CD14-dependent and -independent cytokine and chemokine production by human THP-1 monocytes stimulated by Streptococcus suis capsular type 2. Clin. Exp. Immunol. 2002, 127, 243–254. [Google Scholar] [CrossRef]
- Torretta, S.; Scagliola, A.; Ricci, L.; Mainini, F.; Di Marco, S.; Cuccovillo, I.; Kajaste-Rudnitski, A.; Sumpton, D.; Ryan, K.M.; Cardaci, S. D-mannose suppresses macrophage IL-1beta production. Nat. Commun. 2020, 11, 6343. [Google Scholar] [CrossRef]
- Liu, H.; Wu, X.; Gang, N.; Wang, S.; Deng, W.; Zan, L.; Yu, S. Macrophage functional phenotype can be consecutively and reversibly shifted to adapt to microenvironmental changes. Int. J. Clin. Exp. Med. 2015, 8, 3044–3053. [Google Scholar]
- Italiani, P.; Mazza, E.M.; Lucchesi, D.; Cifola, I.; Gemelli, C.; Grande, A.; Battaglia, C.; Bicciato, S.; Boraschi, D. Transcriptomic profiling of the development of the inflammatory response in human monocytes in vitro. PLoS ONE 2014, 9, e87680. [Google Scholar] [CrossRef]
- Wang, S.; Wang, G.; Tang, Y.D.; Li, S.; Qin, L.; Wang, M.; Yang, Y.B.; Gottschalk, M.; Cai, X. Streptococcus suis Serotype 2 Infection Induces Splenomegaly with Splenocyte Apoptosis. Microbiol. Spectr. 2022, 10, e0321022. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wong, Y.S. Selenocystine induces caspase-independent apoptosis in MCF-7 human breast carcinoma cells with involvement of p53 phosphorylation and reactive oxygen species generation. Int. J. Biochem. Cell Biol. 2009, 41, 666–676. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, C.; Tang, Y.-D.; Qin, L.; Chen, T.; Wang, S.; Bai, Y.; Cai, X.; Wang, S. Interaction between Porcine Alveolar Macrophage-Tang Cells and Streptococcus suis Strains of Different Virulence: Phagocytosis and Apoptosis. Microorganisms 2023, 11, 160. https://doi.org/10.3390/microorganisms11010160
Li S, Wang C, Tang Y-D, Qin L, Chen T, Wang S, Bai Y, Cai X, Wang S. Interaction between Porcine Alveolar Macrophage-Tang Cells and Streptococcus suis Strains of Different Virulence: Phagocytosis and Apoptosis. Microorganisms. 2023; 11(1):160. https://doi.org/10.3390/microorganisms11010160
Chicago/Turabian StyleLi, Siqi, Chunsheng Wang, Yan-Dong Tang, Lei Qin, Tianfeng Chen, Shanghui Wang, Yuanzhe Bai, Xuehui Cai, and Shujie Wang. 2023. "Interaction between Porcine Alveolar Macrophage-Tang Cells and Streptococcus suis Strains of Different Virulence: Phagocytosis and Apoptosis" Microorganisms 11, no. 1: 160. https://doi.org/10.3390/microorganisms11010160
APA StyleLi, S., Wang, C., Tang, Y. -D., Qin, L., Chen, T., Wang, S., Bai, Y., Cai, X., & Wang, S. (2023). Interaction between Porcine Alveolar Macrophage-Tang Cells and Streptococcus suis Strains of Different Virulence: Phagocytosis and Apoptosis. Microorganisms, 11(1), 160. https://doi.org/10.3390/microorganisms11010160