

Characterizing Effects of Microbial Biostimulants and Whole-Soil Inoculums for Native Plant Revegetation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
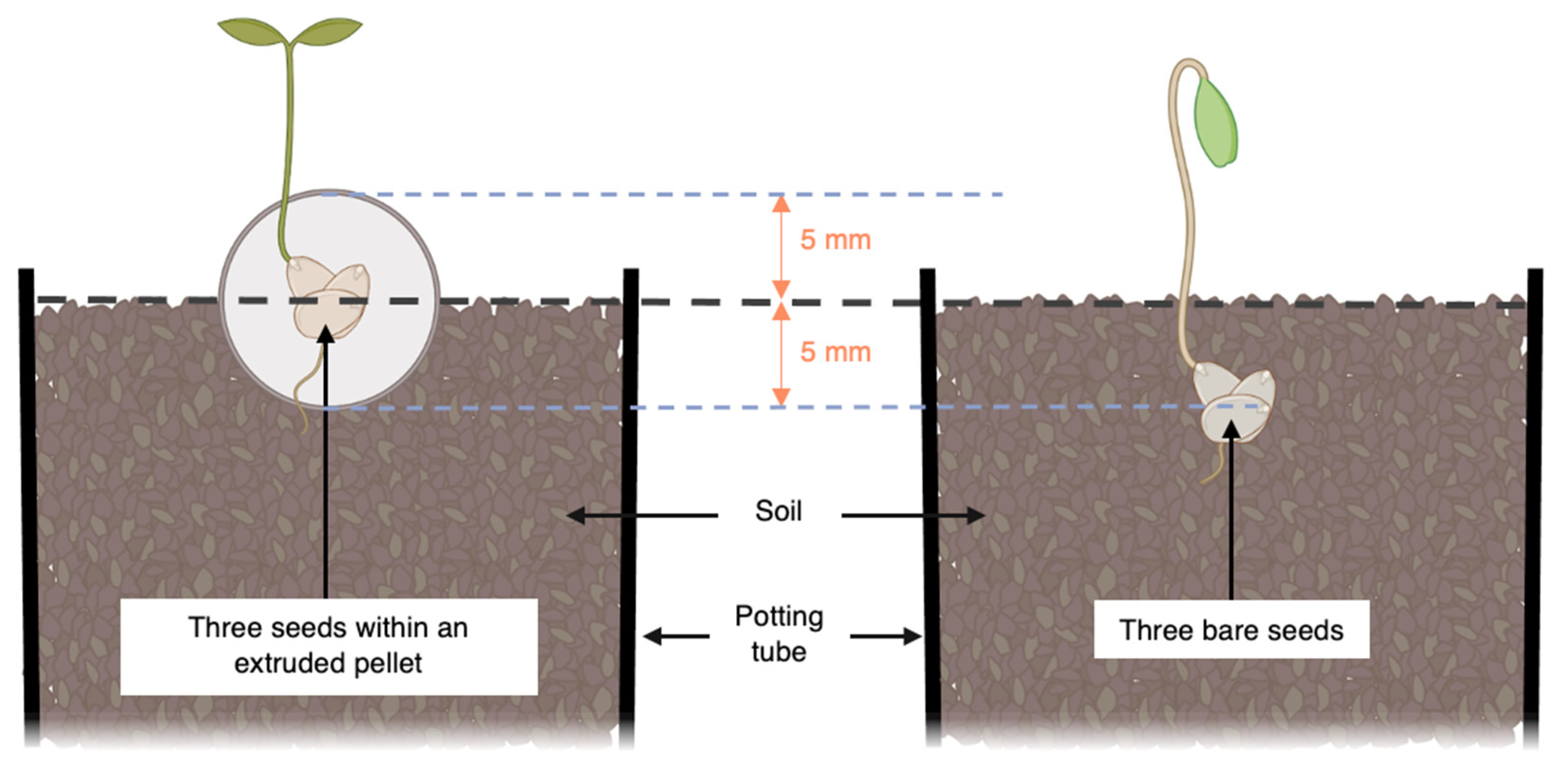
2.2. Growth Trials and Seed Treatments
2.3. Nutrient and Physicochemical Analyses Microbial Community Diversity Quantifications
2.4. Microbial Community Diversity Quantifications
2.5. Seedling Emergence, Survival, and Growth Data Collection
2.6. Statistical Analyses
3. Results
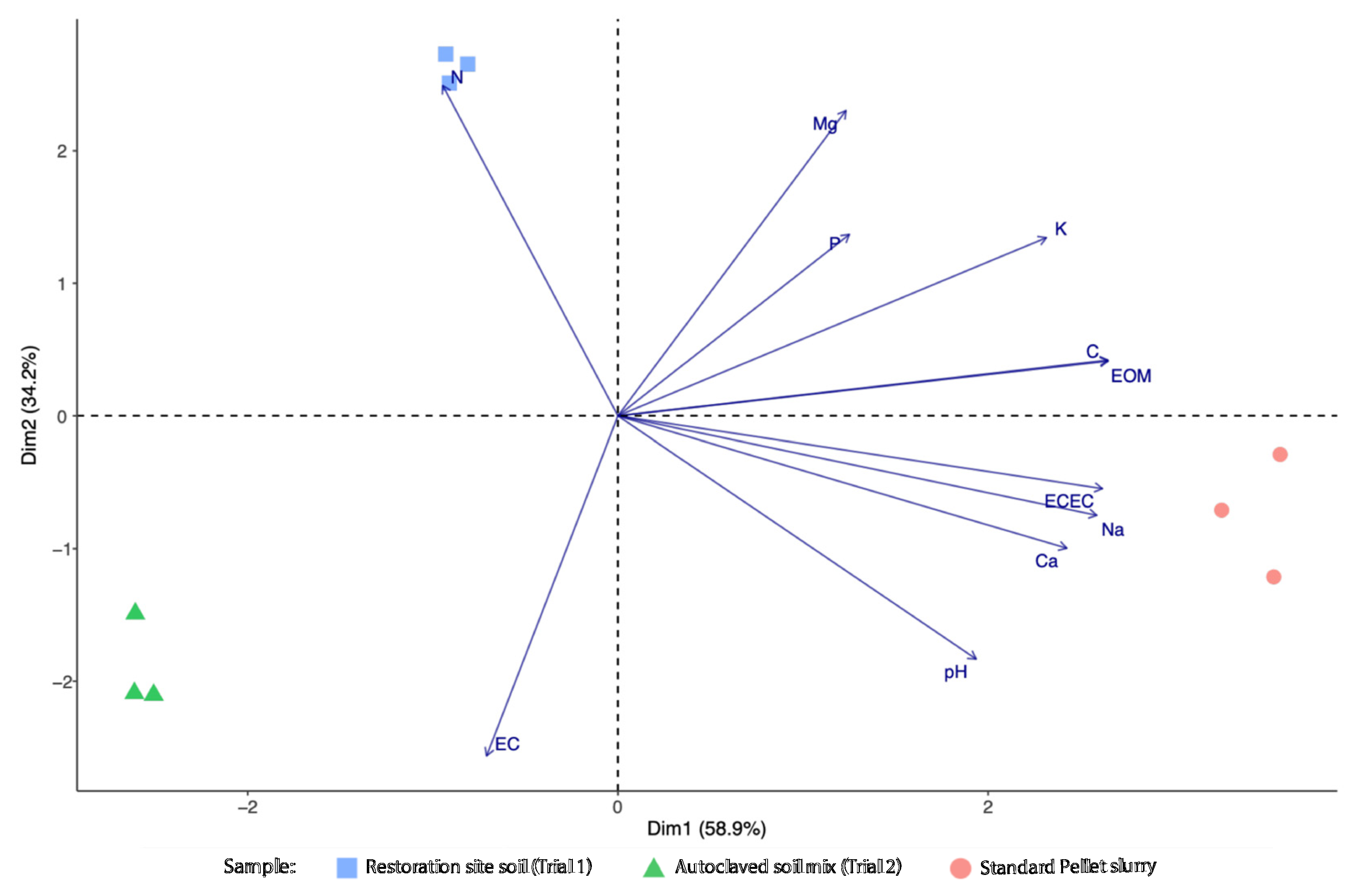
3.1. Nutrient and Physicochemical Properties of Extruded Seed Pellets and Soils
3.2. Microbial Community Diversity
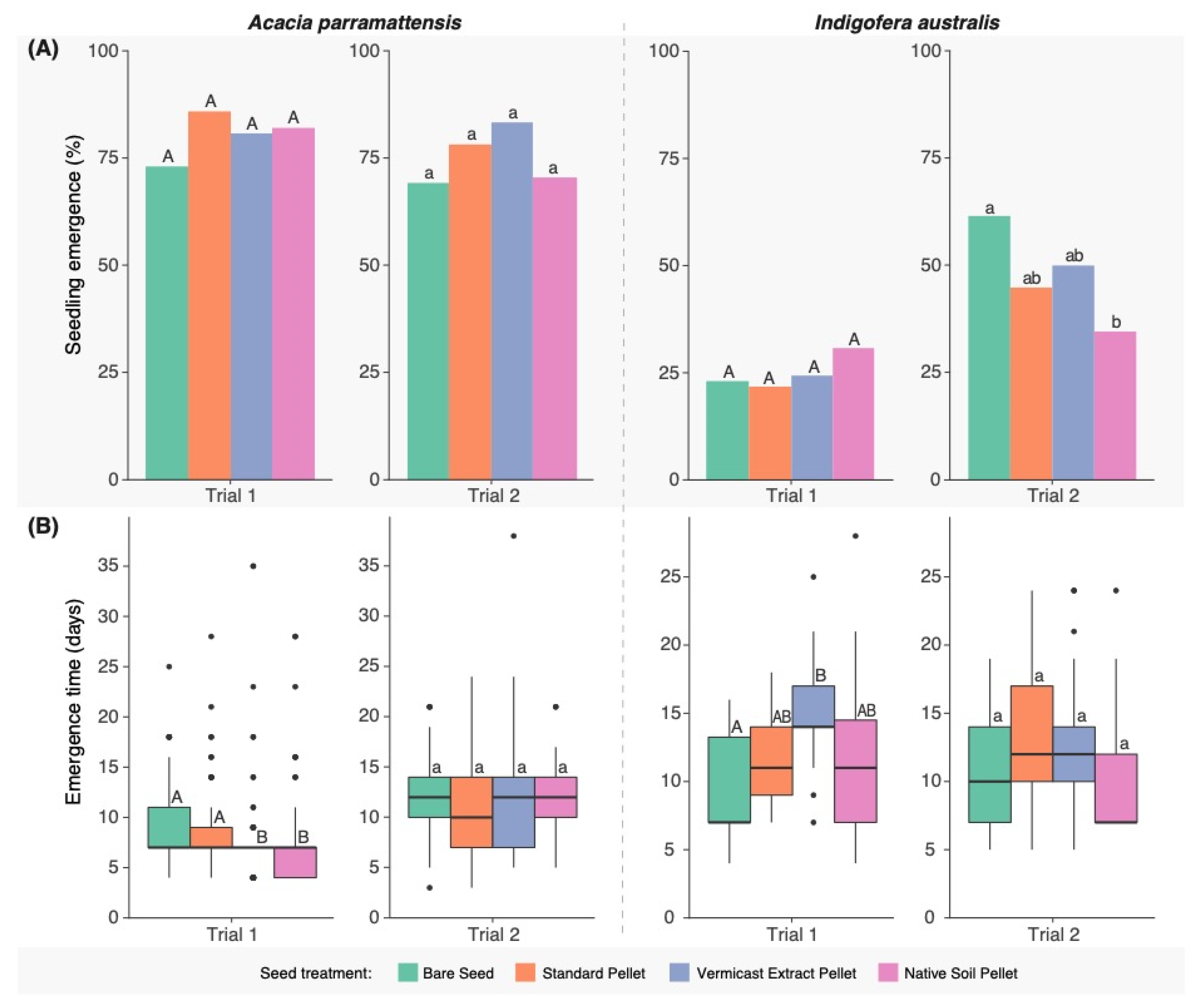
3.3. Seedling Emergence
3.4. Seedling Survival
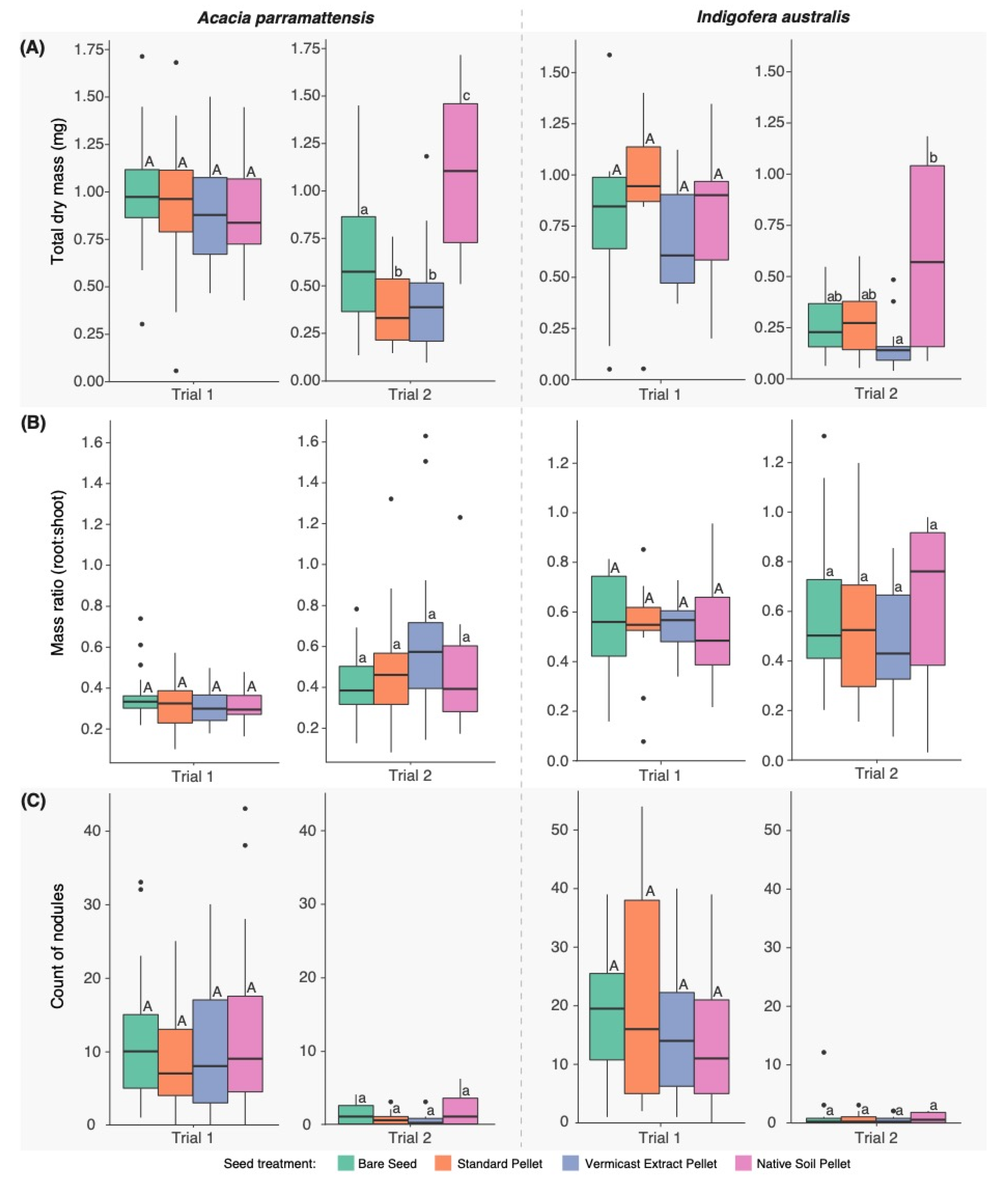
3.5. Post-Harvest Plant Growth Metrics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poore, J.A. Call for conservation: Abandoned pasture. Science 2016, 351, 132. [Google Scholar] [CrossRef] [PubMed]
- Verdone, M.; Seidl, A. Time, space, place, and the Bonn Challenge global forest restoration target. Restor. Ecol. 2017, 25, 903–911. [Google Scholar] [CrossRef]
- Löf, M.; Madsen, P.; Metslaid, M.; Witzell, J.; Jacobs, D.F. Restoring forests: Regeneration and ecosystem function for the future. New For. 2019, 50, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Parkhurst, T.; Prober, S.M.; Hobbs, R.J.; Standish, R.J. Global meta-analysis reveals incomplete recovery of soil conditions and invertebrate assemblages after ecological restoration in agricultural landscapes. J. Appl. Ecol. 2021, 59, 358–372. [Google Scholar] [CrossRef]
- Grossnickle, S.; Ivetić, V. Direct seeding in reforestation—A field performance review. Reforesta 2017, 4, 94–142. [Google Scholar] [CrossRef]
- Hancock, N.; Gibson-Roy, P.; Driver, M.; Broadhurst, L. The Australian Native Seed Survey Report; Australian Network for Plant Conservation: Canberra, Australia, 2020. [Google Scholar]
- Madsen, M.D.; Davies, K.W.; Boyd, C.S.; Kerby, J.D.; Svejcar, T.J. Emerging seed enhancement technologies for overcoming barriers to restoration. Restor. Ecol. 2016, 24, S77–S84. [Google Scholar] [CrossRef]
- Pedrini, S.; Balestrazzi, A.; Madsen, M.D.; Bhalsing, K.; Hardegree, S.P.; Dixon, K.W.; Kildisheva, O.A. Seed enhancement: Getting seeds restoration-ready. Restor. Ecol. 2020, 28, S266–S275. [Google Scholar] [CrossRef]
- Perring, M.P.; Standish, R.J.; Price, J.N.; Craig, M.D.; Erickson, T.E.; Ruthrof, K.X.; Whiteley, A.S.; Valentine, L.E.; Hobbs, R.J. Advances in restoration ecology: Rising to the challenges of the coming decades. Ecosphere 2015, 6, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Bennett, J.A.; Klironomos, J. Mechanisms of plant-soil feedback: Interactions among biotic and abiotic drivers. New Phytol. 2019, 222, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contos, P.; Wood, J.L.; Murphy, N.P.; Gibb, H. Rewilding with invertebrates and microbes to restore ecosystems: Present trends and future directions. Ecol. Evol. 2021, 11, 7187–7200. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Hautier, Y.; Pellkofer, S.; Banerjee, S.; Schmid, B.; van der Heijden, M.G. Diversity and asynchrony in soil microbial communities stabilizes ecosystem functioning. Elife 2021, 10, e62813. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica (Cairo) 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.C. Perspectives and challenges of microbial application for crop improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Ansari, R.A.; Mahmood, I. Plant Health under Biotic Stress: Volume 1: Organic Strategies; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Coban, O.; De Deyn, G.B.; van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, abe0725. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Lehmann, A.; Zheng, W.; Ryo, M.; Soutschek, K.; Roy, J.; Rongstock, R.; Maass, S.; Rillig, M.C. Fungal traits important for soil aggregation. Front. Microbiol. 2019, 10, 2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneller, T.; Harris, R.J.; Bateman, A.; Munoz-Rojas, M. Native-plant amendments and topsoil addition enhance soil function in post-mining arid grasslands. Sci. Total Environ. 2018, 621, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Grez, B.; Munoz-Rojas, M.; Kariman, K.; Storer, P.; O’Donnell, A.G.; Kumaresan, D.; Whiteley, A.S. Reconditioning degraded mine site soils with exogenous soil microbes: Plant fitness and soil microbiome outcomes. Front. Microbiol. 2019, 10, 1617. [Google Scholar] [CrossRef] [Green Version]
- Koziol, L.; Bauer, J.T.; Duell, E.B.; Hickman, K.; House, G.L.; Schultz, P.A.; Tipton, A.G.; Wilson, G.W.T.; Bever, J.D. Manipulating plant microbiomes in the field: Native mycorrhizae advance plant succession and improve native plant restoration. J. Appl. Ecol. 2021, 59, 1976–1985. [Google Scholar] [CrossRef]
- Bizuti, D.T.G.; Robin, A.; Soares, T.M.; Moreno¹, V.S.; Almeida, D.R.A.; Andreote, F.D.; Casagrande, J.C.; Guillemot, J.; Herrmann, L.; Melis, J.; et al. Multifunctional soil recovery during the restoration of Brazil’s Atlantic Forest after bauxite mining. J. Appl. Ecol. 2022, 59, 2262–2273. [Google Scholar] [CrossRef]
- Albornoz, F.E.; Prober, S.M.; Ryan, M.H.; Standish, R.J. Ecological interactions among microbial functional guilds in the plant-soil system and implications for ecosystem function. Plant Soil 2022, 476, 301–313. [Google Scholar] [CrossRef]
- House, G.L.; Bever, J.D. Disturbance reduces the differentiation of mycorrhizal fungal communities in grasslands along a precipitation gradient. Ecol. Appl. 2018, 28, 736–748. [Google Scholar] [CrossRef] [Green Version]
- Kiesewetter, K.N.; Afkhami, M.E. Microbiome-mediated effects of habitat fragmentation on native plant performance. New Phytol. 2021, 232, 1823–1838. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; Erickson, T.E.; Dixon, K.W.; Merritt, D.J. Soil quality indicators to assess functionality of restored soils in degraded semiarid ecosystems. Restor. Ecol. 2016, 24, S43–S52. [Google Scholar] [CrossRef]
- van der Heyde, M.; Bunce, M.; Dixon, K.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Changes in soil microbial communities in post mine ecological restoration: Implications for monitoring using high throughput DNA sequencing. Sci. Total Environ. 2020, 749, 142262. [Google Scholar] [CrossRef]
- Jansa, J.; Mozafar, A.; Anken, T.; Ruh, R.; Sanders, I.R.; Frossard, E. Diversity and structure of AMF communities as affected by tillage in a temperate soil. Mycorrhiza 2002, 12, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Zuber, S.M.; Villamil, M.B. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biol. Biochem. 2016, 97, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Oehl, F.; Sieverding, E.; Ineichen, K.; Mader, P.; Boller, T.; Wiemken, A. Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of Central Europe. Appl. Environ. Microbiol. 2003, 69, 2816–2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayyab, M.; Yang, Z.; Zhang, C.; Islam, W.; Lin, W.; Zhang, H. Sugarcane monoculture drives microbial community composition, activity and abundance of agricultural-related microorganisms. Env. Sci Pollut Res Int 2021, 28, 48080–48096. [Google Scholar] [CrossRef] [PubMed]
- Säle, V.; Aguilera, P.; Laczko, E.; Mäder, P.; Berner, A.; Zihlmann, U.; van der Heijden, M.G.A.; Oehl, F. Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2015, 84, 38–52. [Google Scholar] [CrossRef]
- Smith, M.E.; Delean, S.; Cavagnaro, T.R.; Facelli, J.M. Evidence for species-specific plant responses to soil microbial communities from remnant and degraded land provides promise for restoration. Austral Ecol. 2018, 43, 301–308. [Google Scholar] [CrossRef]
- Grman, E.; Allen, J.; Galloway, E.; McBride, J.; Bauer, J.T.; Price, P.A. Inoculation with remnant prairie soils increased the growth of three native prairie legumes but not necessarily their associations with beneficial soil microbes. Restor. Ecol. 2020, 28, S393–S399. [Google Scholar] [CrossRef]
- Remke, M.J.; Hoang, T.; Kolb, T.; Gehring, C.; Johnson, N.C.; Bowker, M.A. Familiar soil conditions help Pinus ponderosa seedlings cope with warming and drying climate. Restor. Ecol. 2020, 28, S344–S354. [Google Scholar] [CrossRef]
- Román, J.R.; Roncero-Ramos, B.; Chamizo, S.; Rodríguez-Caballero, E.; Cantón, Y. Restoring soil functions by means of cyanobacteria inoculation: Importance of soil conditions and species selection. Land Degrad. Dev. 2018, 29, 3184–3193. [Google Scholar] [CrossRef]
- Lance, A.C.; Burke, D.J.; Hausman, C.E.; Burns, J.H. High-throughput sequencing provides insight into manipulated soil fungal community structure and diversity during temperate forest restoration. Restor. Ecol. 2020, 28. [Google Scholar] [CrossRef]
- Berg, G.; Kusstatscher, P.; Abdelfattah, A.; Cernava, T.; Smalla, K. Microbiome Modulation-Toward a Better Understanding of Plant Microbiome Response to Microbial Inoculants. Front. Microbiol. 2021, 12, 650610. [Google Scholar] [CrossRef] [PubMed]
- Tejada, M.; García-Martínez, A.M.; Parrado, J. Effects of a vermicompost composted with beet vinasse on soil properties, soil losses and soil restoration. Catena 2009, 77, 238–247. [Google Scholar] [CrossRef]
- Fehmi, J.S.; Rasmussen, C.; Gallery, R.E. Biochar and woodchip amendments alter restoration outcomes, microbial processes, and soil moisture in a simulated semi-arid ecosystem. Restor. Ecol. 2020, 28, S355–S364. [Google Scholar] [CrossRef]
- Halmer, P. Seed technology and seed enhancement. Acta Hortic. 2008, 771, 17–26. [Google Scholar] [CrossRef]
- Brown, V.S.; Erickson, T.E.; Merritt, D.J.; Madsen, M.D.; Hobbs, R.J.; Ritchie, A.L. A global review of seed enhancement technology use to inform improved applications in restoration. Sci. Total Environ. 2021, 798, 149096. [Google Scholar] [CrossRef]
- Stock, E.; Standish, R.J.; Muñoz-Rojas, M.; Bell, R.W.; Erickson, T.E. Field-deployed extruded seed pellets show promise for perennial grass establishment in arid zone mine rehabilitation. Front. Ecol. Evol. 2020, 8, 576125. [Google Scholar] [CrossRef]
- Madsen, M.D.; Svejcar, L.; Radke, J.; Hulet, A. Inducing rapid seed germination of native cool season grasses with solid matrix priming and seed extrusion technology. PLoS ONE 2018, 13, e0204380. [Google Scholar] [CrossRef] [Green Version]
- Pedrini, S. Seed Enhancement Research for Improving Ecological Restoration; Curtin University: Bently, Australia, 2018. [Google Scholar]
- Müller, H.; Berg, G. Impact of formulation procedures on the effect of the biocontrol agent Serratia plymuthica HRO-C48 on Verticillium wilt in oilseed rape. BioControl 2007, 53, 905–916. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and, Evolution of Dormancy and Germination; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods—Australasia; CSIRO Publishing: Clayton, Australia, 2010. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. Available online: https://www.R-project.org/ (accessed on 1 October 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.6.2-1. 2021. Available online: https://github.com/rvlenth/emmeans (accessed on 1 October 2022).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Farrell, H.L.; Léger, A.; Breed, M.F.; Gornish, E.S. Restoration, soil organisms, and soil processes: Emerging approaches. Restor. Ecol. 2020, 28, S307–S310. [Google Scholar] [CrossRef]
- Yang, B.; Balazs, K.R.; Butterfield, B.J.; Laushman, K.M.; Munson, S.M.; Gornish, E.S.; Barberán, A. Does restoration of plant diversity trigger concomitant soil microbiome changes in dryland ecosystems? J. Appl. Ecol. 2021, 59, 560–573. [Google Scholar] [CrossRef]
- Deng, S.; Wipf, H.M.; Pierroz, G.; Raab, T.K.; Khanna, R.; Coleman-Derr, D. A plant growth-promoting microbial soil amendment dynamically alters the strawberry root bacterial microbiome. Sci. Rep. 2019, 9, 17677. [Google Scholar] [CrossRef] [Green Version]
- Roman, J.R.; Chilton, A.M.; Canton, Y.; Munoz-Rojas, M. Assessing the viability of cyanobacteria pellets for application in arid land restoration. J. Environ. Manag. 2020, 270, 110795. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.; Bougoure, J.J.; Southam, G.; Chan, T.S.; Lu, Y.R.; Haw, S.C.; Nguyen, T.A.H.; You, F.; Huang, L. Organic matter amendment and plant colonization drive mineral weathering, organic carbon sequestration, and water-stable aggregation in magnetite Fe ore tailings. Environ. Sci. Technol. 2019, 53, 13720–13731. [Google Scholar] [CrossRef] [PubMed]
- Sarma, B.K.; Singh, P.; Pandey, S.K.; Singh, H.B. Vermicompost as modulator of plant growth and disease suppression. Dyn. Soil Dyn. Plant 2010, 4, 58–66. [Google Scholar]
- Piya, S.; Gauchan, D.; Lamichhane, J. Efficacy of vermicompost on seedling growth of Solanum melongena var. Mayalu and Arka Keshav. Int. J. Biosci. 2019, 13, 177–191. [Google Scholar]
- Alfonzetti, M.; Tetu, S.; Mills, C.H.; Gallagher, R.V. Assessing the efficacy of extruded seed pellets and microbial 511 amendments for native revegetation. Restoration Ecology 2022. in review. [Google Scholar] [CrossRef]
- Dadzie, F.A.; Moles, A.T.; Erickson, T.E.; Slavich, E.; Muñoz-Rojas, M. Native bacteria and cyanobacteria can influence seedling emergence and growth of native plants used in dryland restoration. J. Appl. Ecol. 2022, 59, 2983–2992. [Google Scholar] [CrossRef]
- Maltz, M.R.; Treseder, K.K. Sources of inocula influence mycorrhizal colonization of plants in restoration projects: A meta-analysis. Restor. Ecol. 2015, 23, 625–634. [Google Scholar] [CrossRef]
- Herrera Paredes, S.; Gao, T.; Law, T.F.; Finkel, O.M.; Mucyn, T.; Teixeira, P.; Salas Gonzalez, I.; Feltcher, M.E.; Powers, M.J.; Shank, E.A.; et al. Design of synthetic bacterial communities for predictable plant phenotypes. PLoS Biol. 2018, 16, e2003962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Látr, A.; Rocha, I.; Freitas, H.; Vosátka, M.; Oliveira, R. Delivery of inoculum of Rhizophagus irregularis via seed coating in combination with Pseudomonas libanensis for cowpea production. Agronomy 2019, 9, 33. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Env. Microbiol 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, M.; Germanier, F.; Vuille, N.; Weisskopf, L. Combining different potato-associated pseudomonas strains for improved biocontrol of Phytophthora infestans. Front. Microbiol. 2018, 9, 2573. [Google Scholar] [CrossRef]
- Wong, W.S.; Morald, T.K.; Whiteley, A.S.; Nevill, P.G.; Trengove, R.D.; Yong, J.W.H.; Dixon, K.W.; Valliere, J.M.; Stevens, J.C.; Veneklaas, E.J. Microbial inoculation to improve plant performance in mine waste substrates—A test using pigeon pea (Cajanus cajan). Land Degrad. Dev. 2021, 33, 497–511. [Google Scholar] [CrossRef]
- Rajendran, G.; Sing, F.; Desai, A.J.; Archana, G. Enhanced growth and nodulation of pigeon pea by co-inoculation of Bacillus strains with Rhizobium spp. Bioresour. Technol. 2008, 99, 4544–4550. [Google Scholar] [CrossRef]
- John, R.P.; Tyagi, R.D.; Brar, S.K.; Prévost, D. Development of emulsion from rhizobial fermented starch industry wastewater for application as Medicago sativa seed coat. Eng. Life Sci. 2010, 10, 248–256. [Google Scholar] [CrossRef]
- McIntyre, H.J.; Davies, H.; Hore, T.A.; Miller, S.H.; Dufour, J.P.; Ronson, C.W. Trehalose biosynthesis in Rhizobium leguminosarum bv. trifolii and its role in desiccation tolerance. Appl. Environ. Microbiol. 2007, 73, 3984–3992. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Restoration Site Soil | Autoclaved Soil Mix | Standard Pellet Slurry |
|---|---|---|---|
| Estimated organic matter (%OM) | 9.36 ± 0.29 | 4.03 ± 0.09 | 16.79 ± 0.80 |
| Total carbon (%) | 5.36 ± 0.18 | 2.30 ± 0.06 | 9.59 ± 0.46 |
| Exchangeable magnesium (mg/kg) | 413.33 ± 2.73 | 207.00 ± 4.16 | 347.14 ± 16.17 |
| Exchangeable calcium (mg/kg) | 2562.67 ± 58.87 | 2708.00 ± 103.91 | 4167.55 ± 238.95 |
| Exchangeable potassium (mg/kg) | 556.00 ± 5.20 | 268.33 ± 4.67 | 656.21 ± 22.27 |
| Exchangeable sodium (mg/kg) | 47.67 ± 13.68 | 61.00 ± 1.00 | 1840.53 ± 26.34 |
| Phosphorus (mg/kg P) | 6.47 ± 0.12 | 4.00 ± 0.06 | 6.57 ± 1.79 |
| Total nitrogen (%) | 0.36 ± 0.01 | 0.12 ± 0.00 | 0.09 ± 0.01 |
| pH | 6.80 ± 0.00 | 7.60 ± 0.00 | 8.50 ± 0.05 |
| Electrical Conductivity (dS/m) | 0.16 ± 0.00 | 0.88 ± 0.07 | 0.52 ± 0.02 |
| Effective Cation Exchange Capacity (cmol + /kg) | 17.67 ± 0.33 | 16.00 ± 0.58 | 33.34 ± 1.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfonzetti, M.; Doleac, S.; Mills, C.H.; Gallagher, R.V.; Tetu, S. Characterizing Effects of Microbial Biostimulants and Whole-Soil Inoculums for Native Plant Revegetation. Microorganisms 2023, 11, 55. https://doi.org/10.3390/microorganisms11010055
Alfonzetti M, Doleac S, Mills CH, Gallagher RV, Tetu S. Characterizing Effects of Microbial Biostimulants and Whole-Soil Inoculums for Native Plant Revegetation. Microorganisms. 2023; 11(1):55. https://doi.org/10.3390/microorganisms11010055
Chicago/Turabian StyleAlfonzetti, Matthew, Sebastien Doleac, Charlotte H. Mills, Rachael V. Gallagher, and Sasha Tetu. 2023. "Characterizing Effects of Microbial Biostimulants and Whole-Soil Inoculums for Native Plant Revegetation" Microorganisms 11, no. 1: 55. https://doi.org/10.3390/microorganisms11010055
APA StyleAlfonzetti, M., Doleac, S., Mills, C. H., Gallagher, R. V., & Tetu, S. (2023). Characterizing Effects of Microbial Biostimulants and Whole-Soil Inoculums for Native Plant Revegetation. Microorganisms, 11(1), 55. https://doi.org/10.3390/microorganisms11010055