
Comparative Genomic Analysis Reveals the Emergence of ST-231 and ST-395 Klebsiella pneumoniae Strains Associated with the High Transmissibility of blaKPC Plasmids


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
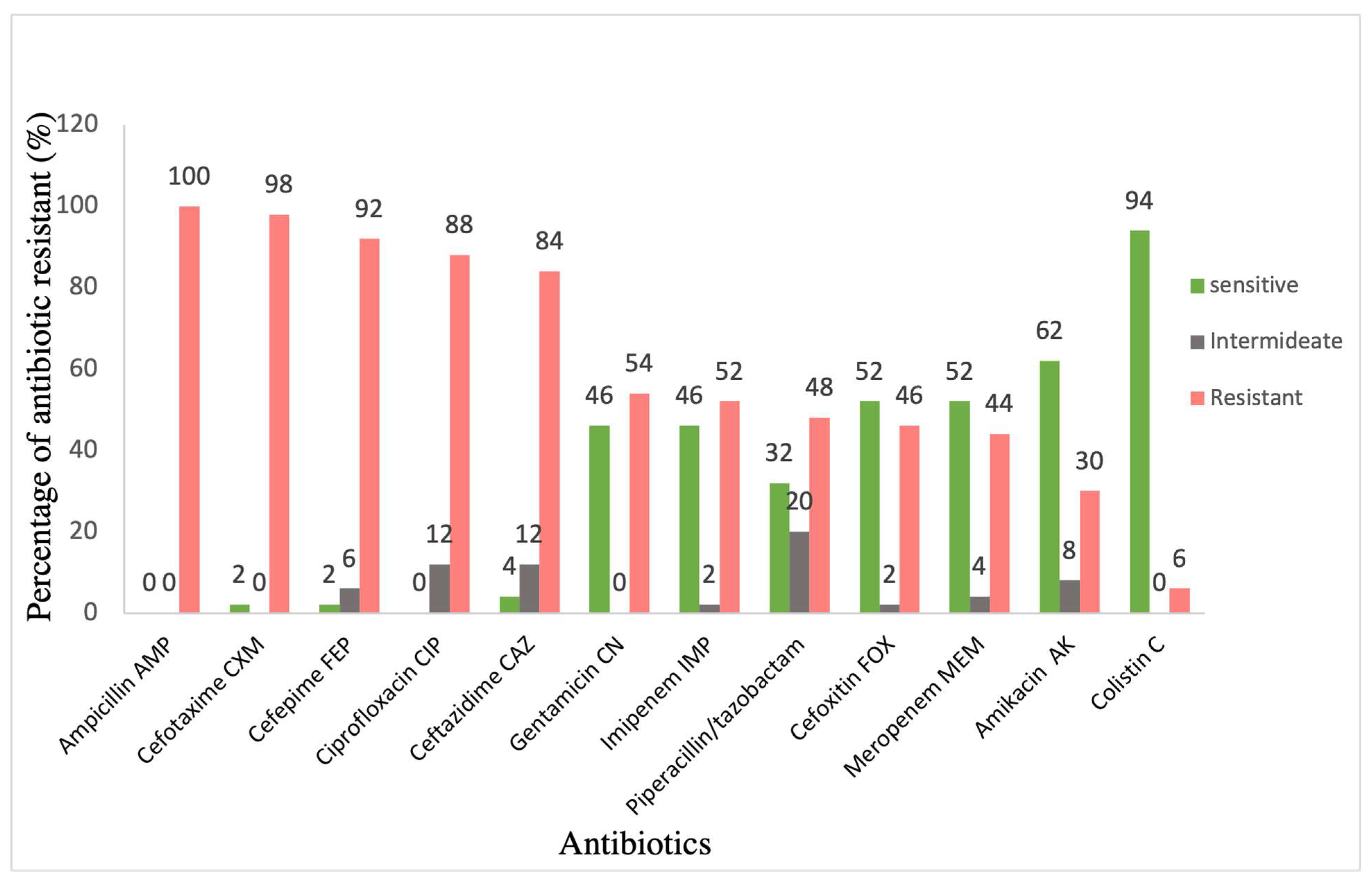
2.2. Antimicrobial Susceptibility
2.3. Genomic DNA Extraction and Purification
2.4. Polymerase Chain Reaction (PCR)
2.4.1. Detection and Characterization of Classes 1, 2, and 3 Integrons
2.4.2. PCR Purification
2.5. Gel Electrophoresis
2.6. Whole-Genome DNA Sequencing
2.7. Bioinformatics Analyses
2.8. Conjugation Experiment
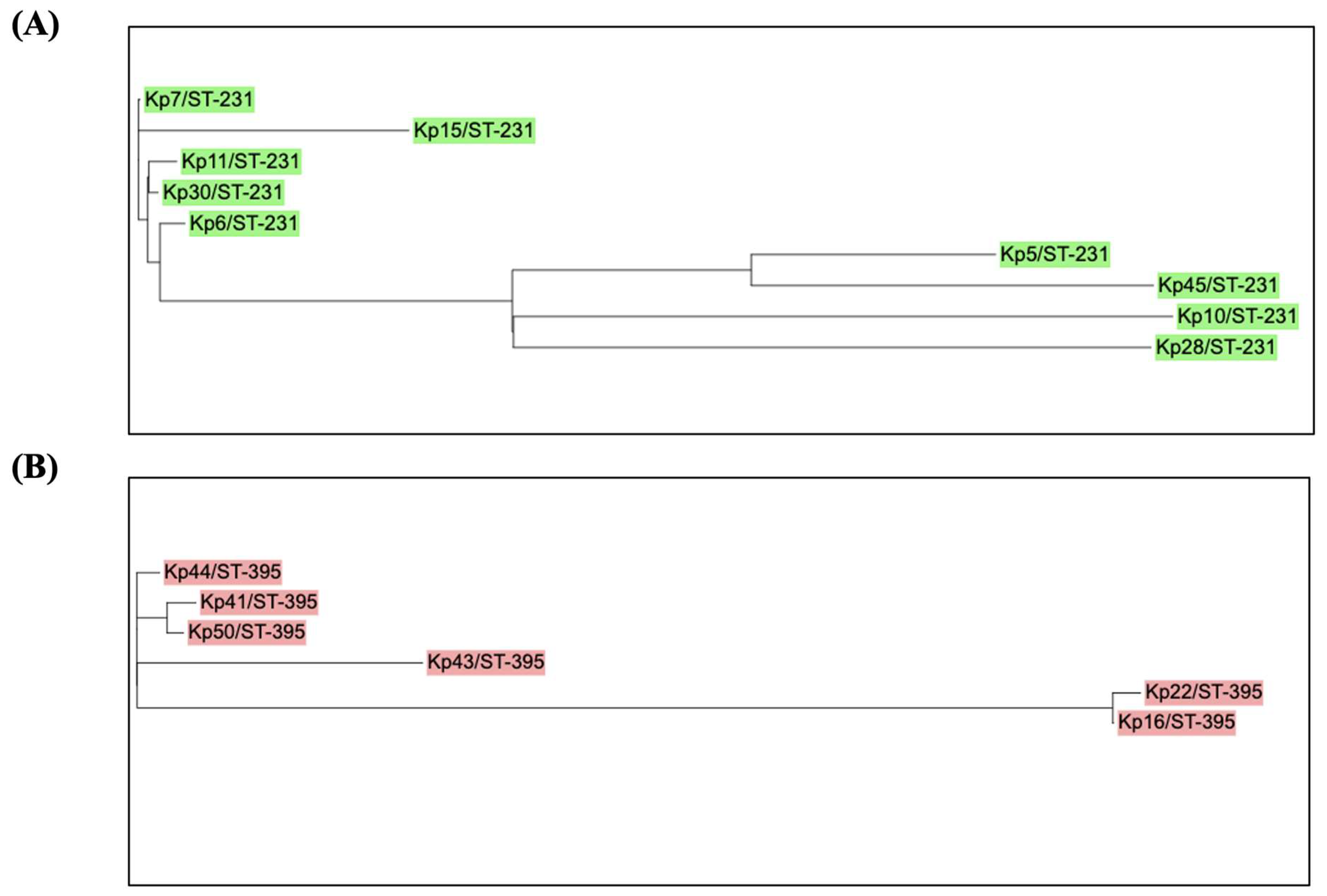
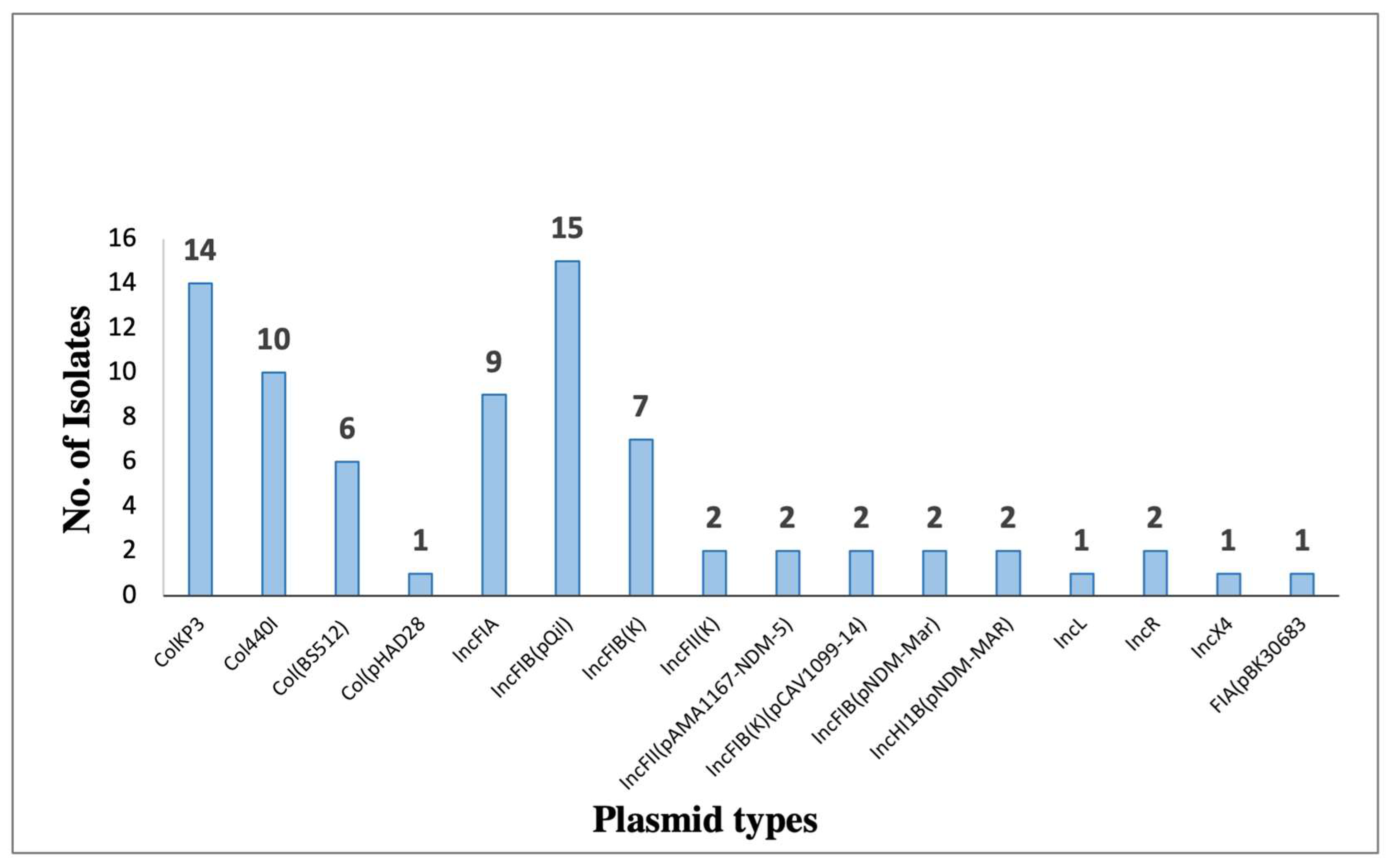
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Podschun, R.; Ullmann, U. Klebsiella Spp. as Nosocomial Pathogens: Epidemiology, Taxonomy, Typing Methods, and Pathogenicity Factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Ashurst, J.V.; Dawson, A. Klebsiella Pneumonia; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Aurilio, C.; Sansone, P.; Barbarisi, M.; Pota, V.; Giaccari, L.G.; Coppolino, F.; Barbarisi, A.; Passavanti, M.B.; Pace, M.C. Mechanisms of Action of Carbapenem Resistance. Antibiotics 2022, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Metwally, L.; Gomaa, N.; Attallah, M.; Kamel, N. High Prevalence of Klebsiella Pneumoniae Carbapenemase-Mediated Resistance in K. Pneumoniae Isolates from Egypt. East. Mediterr. Health J. 2013, 19, 947–952. [Google Scholar] [CrossRef]
- Ainoda, Y.; Aoki, K.; Ishii, Y.; Okuda, K.; Furukawa, H.; Manabe, R.; Sahara, T.; Nakamura-Uchiyama, F.; Kurosu, H.; Ando, Y.; et al. Klebsiella Pneumoniae Carbapenemase (KPC)-Producing Klebsiella Pneumoniae ST258 Isolated from a Japanese Patient without a History of Foreign Travel—A New Public Health Concern in Japan: A Case Report 21 History and Archaeology 2103 Historical Studies. BMC Infect. Dis. 2019, 19, 20. [Google Scholar] [CrossRef]
- Aires-De-Sousa, M.; De La Rosa, J.M.O.; Gonçalves, M.L.; Pereira, A.L.; Nordmann, P.; Poirel, L. Epidemiology of Carbapenemase-Producing Klebsiella Pneumoniae in a Hospital, Portugal. Emerg. Infect. Dis. 2019, 25, 1632–1638. [Google Scholar] [CrossRef]
- Arnold, R.S.; Thom, K.A.; Sharma, S.; Phillips, M.; Kristie Johnson, J.; Morgan, D.J. Emergence of Klebsiella Pneumoniae Carbapenemase-Producing Bacteria. South. Med. J. 2011, 104, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D. Integrons: Agents of Bacterial Evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Fluit, A.C.; Schmitz, F.J. Resistance Integrons and Super-Integrons. Clin. Microbiol. Infect. 2004, 10, 272–288. [Google Scholar] [CrossRef]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef]
- Tseng, C.H.; Huang, Y.T.; Mao, Y.C.; Lai, C.H.; Yeh, T.K.; Ho, C.M.; Liu, P.Y. Insight into the Mechanisms of Carbapenem Resistance in Klebsiella Pneumoniae: A Study on IS26 Integrons, Beta-Lactamases, Porin Modifications, and Plasmidome Analysis. Antibiotics 2023, 12, 749. [Google Scholar] [CrossRef]
- Firoozeh, F.; Mahluji, Z.; Khorshidi, A.; Zibaei, M. Molecular Characterization of Class 1, 2 and 3 Integrons in Clinical Multi-Drug Resistant Klebsiella Pneumoniae Isolates. Antimicrob. Resist. Infect. Control 2019, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Joshi, P.; Trivedi, P.; Akinwotu, O.; Gajjar, D. Genomic Islands in Klebsiella Pneumoniae. In Microbial Genomic Islands in Adaptation and Pathogenicity; Springer Nature Singapore: Singapore, 2023. [Google Scholar] [CrossRef]
- Di Pilato, V.; Principe, L.; Andriani, L.; Aiezza, N.; Coppi, M.; Ricci, S.; Giani, T.; Luzzaro, F.; Rossolini, G.M. Deciphering Variable Resistance to Novel Carbapenem-Based β-Lactamase Inhibitor Combinations in a Multi-Clonal Outbreak Caused by Klebsiella Pneumoniae Carbapenemase (KPC)-Producing Klebsiella Pneumoniae Resistant to Ceftazidime/Avibactam. Clin. Microbiol. Infect. 2023, 29, 537.e1–537.e8. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Polo, J.A.; Hernández-García, M.; Morosini, M.I.; Pérez-Viso, B.; Soriano, C.; De Pablo, R.; Cantón, R.; Ruiz-Garbajosa, P. Outbreak by KPC-62-Producing ST307 Klebsiella Pneumoniae Isolates Resistant to Ceftazidime/Avibactam and Cefiderocol in a University Hospital in Madrid, Spain. J. Antimicrob. Chemother. 2023, 78, 1259–1264. [Google Scholar] [CrossRef]
- Jahanbin, F.; Marashifard, M.; Jamshidi, S.; Zamanzadeh, M.; Dehshiri, M.; Hosseini, S.A.A.M.; Khoramrooz, S.S. Investigation of Integron-Associated Resistance Gene Cassettes in Urinary Isolates of Klebsiella Pneumoniae in Yasuj, Southwestern Iran during 2015-16. Avicenna J. Med. Biotechnol. 2020, 12, 124–131. [Google Scholar]
- Derakhshan, S.; Peerayeh, S.N.; Bakhshi, B. Association between Presence of Virulence Genes and Antibiotic Resistance in Clinical Klebsiella Pneumoniae Isolates. Lab. Med. 2016, 47, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, X.; Luo, M.; Liu, P.; Su, K.; Qing, Y.; Chen, S.; Qiu, J.; Li, Y. Molecular Characterisations of Integrons in Clinical Isolates of Klebsiella Pneumoniae in a Chinese Tertiary Hospital. Microb. Pathog. 2017, 104, 164–170. [Google Scholar] [CrossRef]
- Solberg, O.D.; Ajiboye, R.M.; Riley, L.W. Origin of Class 1 and 2 Integrons and Gene Cassettes in a Population-Based Sample of Uropathogenic Escherichia Coli. J. Clin. Microbiol. 2006, 44, 1347–1351. [Google Scholar] [CrossRef]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance Integrons: Class 1, 2 and 3 Integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef]
- Drouin, F.; Mélançon, J.; Roy, P.H. The IntI-like Tyrosine Recombinase of Shewanella Oneidensis is Active as an Integron Integrase. J. Bacteriol. 2002, 184, 1811–1815. [Google Scholar] [CrossRef]
- Al Balushi, M.; Kumar, R.; Al-Rashdi, A.; Ratna, A.; Al-Jabri, A.; Al-Shekaili, N.; Rani, R.; Al Sumri, S.; Al-Ghabshi, L.; Al-Abri, S.; et al. Genomic Analysis of the Emerging Carbapenem-Resistant Klebsiella Pneumoniae Sequence Type 11 Harbouring Klebsiella Pneumoniae Carbapenemase (KPC) in Oman. J. Infect. Public Health 2022, 15, 1089–1096. [Google Scholar] [CrossRef]
- Balkhair, A.; Al-Muharrmi, Z.; Al’Adawi, B.; Al Busaidi, I.; Taher, H.B.; Al-Siyabi, T.; Al Amin, M.; Hassan, K.S. Prevalence and 30-Day All-Cause Mortality of Carbapenem-and Colistin-Resistant Bacteraemia Caused by Acinetobacter Baumannii, Pseudomonas Aeruginosa, and Klebsiella Pneumoniae: Description of a Decade-Long Trend. Int. J. Infect. Dis. 2019, 85, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Balkhair, A.; Al-Farsi, Y.M.; Al-Muharrmi, Z.; Al-Rashdi, R.; Al-Jabri, M.; Neilson, F.; Al-Adawi, S.S.; El-Beeli, M.; Al-Adawi, S. Epidemiology of Multi-Drug Resistant Organisms in a Teaching Hospital in Oman: A One-Year Hospital-Based Study. Sci. World J. 2014, 2014, 157102. [Google Scholar] [CrossRef] [PubMed]
- CLSI. M100 Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; Clinical Laboratory Standard Institute: Wayne, NY, USA, 2021. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters; Version 10.0; The European Committee on Antimicrobial Susceptibility Testing: Brussels, Belgium, 2020; Available online: http://www.eucast.org (accessed on 23 December 2020).
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing A CLSI Supplement for Global Application; Clinical and Laboratory Standards Institute: Wayne, NY, USA, 2020. [Google Scholar]
- Queipo-Ortuño, M.I.; De Dios Colmenero, J.; Macias, M.; Bravo, M.J.; Morata, P. Preparation of Bacterial DNA Template by Boiling and Effect of Immunoglobulin g as an Inhibitor in Real-Time PCR for Serum Samples from Patients with Brucellosis. Clin. Vaccine Immunol. 2008, 15, 293–296. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Larsen, J.; Enright, M.C.; Godoy, D.; Spratt, B.G.; Larsen, A.R.; Skov, R.L. Multilocus Sequence Typing Scheme for Staphylococcus Aureus: Revision of the Gmk Locus. J. Clin. Microbiol. 2012, 50, 2538–2539. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. PlasmidFinder and PMLST: In Silico Detection and Typing of Plasmid. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Velhner, M.; Todorović, D.; Novović, K.; Jovčić, B.; Lazić, G.; Kojić, M.; Kehrenberg, C. Characterization of Antibiotic Resistance in Escherichia Coli Isolates from Black-Headed Gulls (Larus Ridibundus) Present in the City of Novi Sad, Serbia. Vet. Res. Commun. 2021, 45, 199–209. [Google Scholar] [CrossRef]
- Filipić, B.; Malešević, M.; Vasiljević, Z.; Novović, K.; Kojić, M.; Jovčić, B. Comparative Genomics of Trimethoprim-Sulfamethoxazole-Resistant Achromobacter Xylosoxidans Clinical Isolates from Serbia Reveals Shortened Variant of Class 1 Integron Integrase Gene. Folia Microbiol. 2023, 68, 431–440. [Google Scholar] [CrossRef]
- Malek, M.M.; Amer, F.A.; Allam, A.A.; El-Sokkary, R.H.; Gheith, T.; Arafa, M.A. Occurrence of Classes I and II Integrons in Enterobacteriaceae Collected from Zagazig University Hospitals, Egypt. Front. Microbiol. 2015, 6, 601. [Google Scholar] [CrossRef] [PubMed]
- Al-Jabri, Z.; Zamudio, R.; Horvath-Papp, E.; Ralph, J.D.; Al-Muharrami, Z.; Rajakumar, K.; Oggioni, M.R. Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter Baumannii. Genes 2018, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Al-Jabri, Z.J.; Busaidi, B.A.; Muzahmi, M.A.; Shabibi, Z.A.; Rizvi, M.; Rashdi, A.A.; Al-Jardani, A.; Farzand, R. Diversity of the Sequence Type Determines the Unique Genetic Arrangement of K-Loci in Hypervirulent Klebsiella Pneumoniae Clinical Strains in Review. Available online: www.frontiersin.org (accessed on 2 August 2023).
- Dandachi, I.; Chaddad, A.; Hanna, J.; Matta, J.; Daoud, Z. Understanding the Epidemiology of Multi-Drug Resistant Gram-Negative Bacilli in the Middle East Using a One Health Approach. Front. Microbiol. 2019, 10, 1941. [Google Scholar] [CrossRef]
- AL Quraini, M.; AL Jabri, Z.; Sami, H.; Mahindroo, J.; Taneja, N.; AL Muharrmi, Z.; AL Busaidi, I.; Rizvi, M. Exploring Synergistic Combinations in Extended and Pan-Drug Resistant (XDR and PDR) Whole Genome Sequenced Acinetobacter Baumannii. Microorganisms 2023, 11, 1409. [Google Scholar] [CrossRef] [PubMed]
- Al-Quraini, M.; Rizvi, M.; Al-Jabri, Z.; Sami, H.; Al-Muzahmi, M.; Al-Muharrmi, Z.; Taneja, N.; Al-Busaidi, I.; Soman, R. Assessment of In-Vitro Synergy of Fosfomycin with Meropenem, Amikacin and Tigecycline in Whole Genome Sequenced Extended and Pan Drug Resistant Klebsiella Pneumoniae: Exploring A Colistin Sparing Protocol. Antibiotics 2022, 11, 153. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.I.A.; Ahmed, A.M.; Sato, M.; Shimamoto, T. Characterization of Integrons and Antimicrobial Resistance Genes in Clinical Isolates of Gram-Negative Bacteria from Palestinian Hospitals. Microbiol. Immunol. 2009, 53, 595–602. [Google Scholar] [CrossRef]
- Derakhshan, S.; Peerayeh, S.N.; Fallah, F.; Bakhshi, B.; Rahbar, M.; Ashrafi, A. Detection of Class 1, 2, and 3 Integrons among Klebsiella Pneumoniae Isolated from Children in Tehran Hospitals. Arch. Pediatr. Infect. Dis. 2014, 1, 164–168. [Google Scholar] [CrossRef]
- Mohammadi, R.; Haddadi, A.; Harzandi, N. Prevalence of Integrons as the Carrier of Multidrug Resistance Genes among Clinical Isolates of Klebsiella. J. Med. Bacteriol. 2019, 8, 23–30. [Google Scholar]
- Xu, H.; Davies, J.; Miao, V. Molecular Characterization of Class 3 Integrons from Delftia spp. J. Bacteriol. 2007, 189, 6276–6283. [Google Scholar] [CrossRef]
- Hobson, C.A.; Pierrat, G.; Tenaillon, O.; Bonacorsi, S.; Bercot, B.; Jaouen, E.; Jacquier, H.; Birgy, A. Klebsiella Pneumoniae Carbapenemase Variants Resistant to Ceftazidime-Avibactam: An Evolutionary Overview. Antimicrob. Agents Chemother. 2022, 66, e0044722. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, G.; Zhao, Q.; Guo, L.; Yang, J. Molecular Characteristics of Clinical IMP-Producing Klebsiella Pneumoniae Isolates: Novel IMP-90 and Integron In2147. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 38. [Google Scholar] [CrossRef] [PubMed]
- Omar, F.H.; Ibrahim, A.H. The Prevalence of Integron Class I and II Among Multi-Drug Resistance Producing Klebsiella Pneumonia. Iraqi J. Agric. Sci. 2023, 54, 619–629. [Google Scholar] [CrossRef]
- Collis, C.M.; Kim, M.J.; Partridge, S.R.; Stokes, H.W.; Hall, R.M. Characterization of the Class 3 Integron and the Site-Specific Recombination System It Determines. J. Bacteriol. 2002, 184, 3017–3026. [Google Scholar] [CrossRef]
- Li, B.; Hu, Y.; Wang, Q.; Yi, Y.; Woo, P.C.Y.; Jing, H.; Zhu, B.; Liu, C.H. Structural Diversity of Class 1 Integrons and Their Associated Gene Cassettes in Klebsiella Pneumoniae Isolates from a Hospital in China. PLoS ONE 2013, 8, e75805. [Google Scholar] [CrossRef] [PubMed]
- da Lima, A.M.S.; de Melo, M.E.S.; Alves, L.C.; Brayner, F.A.; Lopes, A.C.S. Investigation of Class 1 Integrons in Klebsiella Pneumoniae Clinical and Microbiota Isolates Belonging to Different Phylogenetic Groups in Recife, State of Pernambuco. Rev. Soc. Bras. Med. Trop. 2014, 47, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Levesque, C.; Piche, L.; Larose, C.; Roy, P.H. PCR Mapping of Integrons Reveals Several Novel Combinations of Resistance Genes. Antimicrob. Agents Chemother. 1995, 39, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Antunes, P.; Machado, J.; Sousa, J.C.; Peixe, L. Dissemination of Sulfonamide Resistance Genes (Sul1, Sul2, and Sul3) in Portuguese Salmonella Enterica Strains and Relation with Integrons. Antimicrob. Agents Chemother. 2005, 49, 836–839. [Google Scholar] [CrossRef]
- Domínguez, M.; Miranda, C.D.; Fuentes, O.; De La Fuente, M.; Godoy, F.A.; Bello-Toledo, H.; González-Rocha, G. Occurrence of Transferable Integrons and Suland Dfrgenes among Sulfonamide-and/or Trimethoprim-Resistant Bacteria Isolated from Chilean Salmonid Farms. Front. Microbiol. 2019, 10, 748. [Google Scholar] [CrossRef]
- Chuanchuen, R.; Koowatananukul, C.; Khemtong, S. Characterization of Class 1 Integrons with Unusual 3′ Conserved Region from Salmonella Enterica Isolates. Southeast Asian J. Trop. Med. Public Health 2008, 39, 419–424. [Google Scholar]
- Evershed, N.J.; Levings, R.S.; Wilson, N.L.; Djordjevic, S.P.; Hall, R.M. Unusual Class 1 Integron-Associated Gene Cassette Configuration Found in IncA/C Plasmids from Salmonella Enterica. Antimicrob. Agents Chemother. 2009, 53, 2640–2642. [Google Scholar] [CrossRef]
- Domingues, S.; da Silva, G.J.; Nielsen, K.M. Integrons: Vehicles and Pathways for Horizontal Dissemination in Bacteria. Mob. Genet. Elements 2012, 2, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Lan, F.; Lu, Y.; Li, B. Characterization of Integrons and Antimicrobial Resistance in Escherichia Coli Sequence Type 131 Isolates. Can. J. Infect. Dis. Med. Microbiol. 2020, 2020, 3826186. [Google Scholar] [CrossRef] [PubMed]
- Ploy, M.C.; Chainier, D.; Thi, N.H.T.; Poilane, I.; Cruaud, P.; Denis, F.; Collignon, A.; Lambert, T. Integron-Associated Antibiotic Resistance in Salmonella Enterica Serovar Typhi from Asia. Antimicrob. Agents Chemother. 2003, 47, 1427–1429. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.; Hansen, L.H.; Sørensen, S.J. Conjugative Plasmids: Vessels of the Communal Gene Pool. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2275–2289. [Google Scholar] [CrossRef]
- Herron, P.R.; Evans, M.C.; Dyson, P.J. Low Target Site Specificity of an IS6100-Based Mini-Transposon, Tn1792, Developed for Transposon Mutagenesis of Antibiotic-Producing Streptomyces. FEMS Microbiol. Lett. 1999, 171, 215–221. [Google Scholar] [CrossRef]
- Wang, D.; Hou, W.; Chen, J.; Yang, L.; Liu, Z.; Yin, Z.; Feng, J.; Zhou, D. Characterization of Novel Integrons, In1085 and In1086, and the Surrounding Genes in Plasmids from Enterobacteriaceae, and the Role for AttCaadA16 Structural Features during Atti1 × Attc Integration. Front. Microbiol. 2017, 8, 1003. [Google Scholar] [CrossRef]
- Martínez-García, S.; Rodríguez-Martínez, S.; Cancino-Diaz, M.E.; Cancino-Diaz, J.C. Extracellular Proteases of Staphylococcus Epidermidis: Roles as Virulence Factors and Their Participation in Biofilm. APMIS 2018, 126, 177–185. [Google Scholar] [CrossRef]
- Oliveira, P.H.; Touchon, M.; Rocha, E.P.C. The Interplay of Restriction-Modification Systems with Mobile Genetic Elements and Their Prokaryotic Hosts. Nucleic Acids Res. 2014, 42, 10618–10631. [Google Scholar] [CrossRef]
- Greene, P.J.; Gupta, M.; Boyer, H.W.; Brown, W.E.; Rosenberg, J.M. Sequence Analysis of the DNA Encoding the Eco RI Endonuclease and Methylase. J. Biol. Chem. 1981, 256, 2143–2153. [Google Scholar] [CrossRef]
- Shetty, V.P.; Akshay, S.D.; Rai, P.; Deekshit, V.K. Integrons as the Potential Targets for Combating Multidrug Resistance in Enterobacteriaceae Using CRISPR- Cas9 Technique. J. Appl. Microbiol. 2023, 134, lxad137. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, M.; Yan, C.; Zhang, Y.; He, X.; Wu, L.; Xu, J.; Lu, J.; Bao, Q.; Hu, Y.; et al. Class 1 Integrons and Multiple Mobile Genetic Elements in Clinical Isolates of the Klebsiella Pneumoniae Complex from a Tertiary Hospital in Eastern China. Front. Microbiol. 2023, 14, 985102. [Google Scholar] [CrossRef] [PubMed]
- Gai, Z.; Wang, X.; Liu, X.; Tai, C.; Tang, H.; He, X.; Wu, G.; Deng, Z.; Xu, P. The Genes Coding for the Conversion of Carbazole to Catechol Are Flanked by IS6100 Elements in Sphingomonas sp. Strain XLDN2-5. PLoS ONE 2010, 5, e10018. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Qi, Y.; Zhang, X.X.; Ma, L. Xenogenetic Evolutionary of Integrons Promotes the Environmental Pollution of Antibiotic Resistance Genes—Challenges, Progress and Prospects. Water Res. 2023, 231, 119629. [Google Scholar] [CrossRef]
- Abdul Momin, M.H.F.; Liakopoulos, A.; Phee, L.M.; Wareham, D.W. Emergence and Nosocomial Spread of Carbapenem-Resistant OXA-232-Producing Klebsiella Pneumoniae in Brunei Darussalam. J. Glob. Antimicrob. Resist. 2017, 9, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Teo, J.W.P.; Kurup, A.; Lin, R.T.P.; Hsien, K.T. Emergence of Clinical Klebsiella Pneumoniae Producing OXA-232 Carbapenemase in Singapore. New Microbes New Infect. 2013, 1, 13–15. [Google Scholar] [CrossRef]
- Shankar, C.; Mathur, P.; Venkatesan, M.; Pragasam, A.K.; Anandan, S.; Khurana, S.; Veeraraghavan, B. Rapidly Disseminating Bla OXA-232 Carrying Klebsiella Pneumoniae Belonging to ST231 in India: Multiple and Varied Mobile Genetic Elements. BMC Microbiol. 2019, 19, 137. [Google Scholar] [CrossRef]
- Mancini, S.; Poirel, L.; Tritten, M.L.; Lienhard, R.; Bassi, C.; Nordmann, P. Emergence of an MDR Klebsiella Pneumoniae ST231 Producing OXA-232 and RmtF in Switzerland. J. Antimicrob. Chemother. 2018, 73, 821–823. [Google Scholar] [CrossRef]
- Muggeo, A.; Guillard, T.; Klein, F.; Reffuveille, F.; François, C.; Babosan, A.; Bajolet, O.; Bertrand, X.; de Champs, C. Spread of Klebsiella Pneumoniae ST395 Non-Susceptible to Carbapenems and Resistant to Fluoroquinolones in North-Eastern France. J. Glob. Antimicrob. Resist. 2018, 13, 98–103. [Google Scholar] [CrossRef]
- Maida, C.M.; Bonura, C.; Geraci, D.M.; Graziano, G.; Carattoli, A.; Rizzo, A.; Torregrossa, M.V.; Vecchio, D.; Giuffrè, M. Outbreak of ST395 KPC-Producing Klebsiella Pneumoniae in a Neonatal Intensive Care Unit in Palermo, Italy. Infect. Control Hosp. Epidemiol. 2018, 39, 496–498. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Ghazawi, A.A.; Hashmey, R.; Jamal, W.; Rotimi, V.O.; Shibl, A.M.; Al-Jardani, A.; Al-Abri, S.S.; Tariq, W.U.Z.; Weber, S.; et al. Characterization of Carbapenem-Resistant Enterobacteriaceae with High Rate of Autochthonous Transmission in the Arabian Peninsula. PLoS ONE 2015, 10, e0131372. [Google Scholar] [CrossRef]
- Dortet, L.; Poirel, L.; Al Yaqoubi, F.; Nordmann, P. NDM-1, OXA-48 and OXA-181 Carbapenemase-Producing Enterobacteriaceae in Sultanate of Oman. Clin. Microbiol. Infect. 2012, 18, E144–E148. [Google Scholar] [CrossRef] [PubMed]
- uz Zaman, T.; Alrodayyan, M.; AlBladi, M.; AlDrees, M.; Siddique, M.I.; Aljohani, S.; Balkhy, H.H. Clonal Diversity and Genetic Profiling of Antibiotic Resistance among Multidrug/Carbapenem-Resistant Klebsiella Pneumoniae Isolates from a Tertiary Care Hospital in Saudi Arabia. BMC Infect. Dis. 2018, 18, 205. [Google Scholar] [CrossRef] [PubMed]
- Mataseje, L.F.; Boyd, D.A.; Fuller, J.; Haldane, D.; Hoang, L.; Lefebvre, B.; Melano, R.G.; Poutanen, S.; Van Caeseele, P.; Mulvey, M.R. Characterization of OXA-48-like Carbapenemase Producers in Canada, 2011–2014. J. Antimicrob. Chemother. 2018, 73, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Hopkins, K.L.; Loy, R.; Doumith, M.; Meunier, D.; Hill, R.; Pike, R.; Mustafa, N.; Livermore, D.M.; Woodford, N. OXA-48-like Carbapenemases in the UK: An Analysis of Isolates and Cases from 2007 to 2014. J. Antimicrob. Chemother. 2017, 72, 1340–1349. [Google Scholar] [CrossRef]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like Carbapenemases: The Phantom Menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef]
- Li, X.; Ma, W.; Qin, Q.; Liu, S.; Ye, L.; Yang, J.; Li, B. Nosocomial Spread of OXA-232-Producing Klebsiella Pneumoniae ST15 in a Teaching Hospital, Shanghai, China. BMC Microbiol. 2019, 19, 235. [Google Scholar] [CrossRef]
- Lutgring, J.D.; Zhu, W.; De Man, T.J.B.; Avillan, J.J.; Anderson, K.F.; Lonsway, D.R.; Rowe, L.A.; Batra, D.; Rasheed, J.K.; Limbago, B.M. Phenotypic and Genotypic Characterization of Enterobacteriaceae Producing Oxacillinase-48-like Carbapenemases, United States. Emerg. Infect. Dis. 2018, 24, 700–709. [Google Scholar] [CrossRef]
- Zarfel, G.; Hoenigl, M.; Leitner, E.; Salzer, H.J.F.; Feierl, G.; Masoud, L.; Valentin, T.; Krause, R.; Grisold, A.J. Emergence of New Delhi Metallo-β-Lactamase, Austria. Emerg. Infect. Dis. 2011, 17, 129–130. [Google Scholar] [CrossRef]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The Role of Epidemic Resistance Plasmids and International High-Risk Clones in the Spread of Multidrug-Resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef]
- Poirel, L.; Naas, T.; Nordmann, P. Class D Ss-Lactamases: Diversity, Epidemiology and Genetics. Antimicrob. Agents Chemother. 2009, 54, 24–38. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular Mechanisms Related to Colistin Resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Jayol, A.; Nordmann, P.; Brink, A.; Poirel, L. Heteroresistance to Colistin in Klebsiella Pneumoniae Associated with Alterations in the PhoPQ Regulatory System. Antimicrob. Agents Chemother. 2015, 59, 2780–2784. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.S.; Suzuki, Y.; Jones, M.B.; Marshall, S.H.; Rudin, S.D.; Van Duin, D.; Kaye, K.; Jacobs, M.R.; Bonomo, R.A.; Adamsa, M.D. Genomic and Transcriptomic Analyses of Colistin-Resistant Clinical Isolates of Klebsiella Pneumoniae Reveal Multiple Pathways of Resistance. Antimicrob. Agents Chemother. 2015, 59, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Lin, T.L.; Pan, Y.J.; Wang, Y.P.; Lin, Y.T.; Wang, J.T. Colistin Resistance Mechanisms in Klebsiella Pneumoniae Strains from Taiwan. Antimicrob. Agents Chemother. 2015, 59, 2909–2913. [Google Scholar] [CrossRef]
- Kuhn, J.M.; Di, Y.P. Determination of Mutational Timing of Colistin-Resistance Genes through Klebsiella Pneumoniae Evolution. Pharmaceutics 2023, 15, 270. [Google Scholar] [CrossRef]
- Song, Y.; Hu, L.; Shu, Q.; Ye, J.; Liang, J.; Chen, X.; Tan, L. Tigecycline Salvage Therapy for Critically Ill Children with Multidrug-Resistant/Extensively Drug-Resistant Infections after Surgery. Int. J. Infect. Dis. 2018, 75, 82–88. [Google Scholar] [CrossRef]
- Al-Jabri, Z.; Al-Shabibi, Z.; Al-Bimani, A.; Al-Hinai, A.; Al-Shabibi, A.; Rizvi, M. Whole Genome Sequencing of Methicillin-Resistant Staphylococcus Epidermidis Clinical Isolates Reveals Variable Composite SCCmec ACME among Different STs in a Tertiary Care Hospital in Oman. Microorganisms 2021, 9, 1824. [Google Scholar] [CrossRef]
- Pournaras, S.; Koumaki, V.; Spanakis, N.; Gennimata, V.; Tsakris, A. Current Perspectives on Tigecycline Resistance in Enterobacteriaceae: Susceptibility Testing Issues and Mechanisms of Resistance. Int. J. Antimicrob. Agents 2016, 48, 11–18. [Google Scholar] [CrossRef]
- Wang, X.; Xie, Y.; Li, G.; Liu, J.; Li, X.; Tian, L.; Sun, J.; Ou, H.Y.; Qu, H. Whole-Genome-Sequencing Characterization of Bloodstream Infection-Causing Hypervirulent Klebsiella Pneumoniae of Capsular Serotype K2 and ST374. Virulence 2018, 9, 510–521. [Google Scholar] [CrossRef]
- Veleba, M.; Schneiders, T. Tigecycline Resistance Can Occur Independently of the RamA Gene in Klebsiella Pneumoniae. Antimicrob. Agents Chemother. 2012, 56, 4466–4467. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; He, F.; Shi, Q.; Zhao, F.; Xu, J.; Fu, Y.; Yu, Y. The Rapid Emergence of Tigecycline Resistance in BlaKPC-2 Harboring Klebsiella Pneumoniae, as Mediated in Vivo by Mutation in TetA during Tigecycline Treatment. Front. Microbiol. 2018, 9, 648. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, P.; Galimand, M.; Bauraing, C.; Deplano, A.; Vanhoof, R.; De Mendonca, R.; Rodriguez-Villalobos, H.; Struelens, M.; Glupczynski, Y. Emergence of ArmA and RmtB Aminoglycoside Resistance 16S RRNA Methylases in Belgium. J. Antimicrob. Chemother. 2007, 59, 459–464. [Google Scholar] [CrossRef]
- Ayad, A.; Drissi, M.; de Curraize, C.; Dupont, C.; Hartmann, A.; Solanas, S.; Siebor, E.; Amoureux, L.; Neuwirth, C. Occurence of ArmA and RmtB Aminoglycoside Resistance 16S RRNA Methylases in Extended-Spectrum β-Lactamases Producing Escherichia Coli in Algerian Hospitals. Front. Microbiol. 2016, 7, 1409. [Google Scholar] [CrossRef] [PubMed]
- Ragupathi, N.K.D.; Bakthavatchalam, Y.D.; Mathur, P.; Pragasam, A.K.; Walia, K.; Ohri, V.C.; Veeraraghavan, B. Plasmid Profiles among Some ESKAPE Pathogens in a Tertiary Care Centre in South India. Indian J. Med. Res. 2019, 149, 222–231. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Guerout, A.M.; Ploncard, P.; Dychinco, B.; Davies, J.; Mazel, D. The Evolutionary History of Chromosomal Super-Integrons Provides an Ancestry for Multiresistant Integrons. Proc. Natl. Acad. Sci. USA 2001, 98, 652–657. [Google Scholar] [CrossRef]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal Transfer of Antibiotic Resistance Genes in Clinical Environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Month of Isolation | Type of Resistance | Specimen Type | Hospital Unit |
|---|---|---|---|---|
| Kp1 | July | XDR | Tracheal aspirate | Emergency |
| Kp 2 | July | XDR | Urine | Emergency |
| Kp 3 | July | ESBL | Urine | Oncology |
| Kp 4 | July | ESBL | Urine | Pediatrics |
| Kp 5 | July | XDR | Urine | Male Medical |
| Kp 6 | July | PAN | Tracheal aspirate | ICU |
| Kp 7 | July | XDR | Wound | Male Medical |
| Kp 8 | July | ESBL | Urine | Day Care |
| Kp 9 | July | ESBL | Biopsy | Surgery |
| Kp 10 | July | ESBL | Urine | Oncology |
| Kp 11 | August | PAN | Tracheal aspirate | Male Medical |
| Kp 12 | August | ESBL | Pus | Surgery |
| Kp 13 | August | XDR | Blood culture | Emergency |
| Kp 14 | August | ESBL | Wound | Male Medical |
| Kp 15 | August | XDR | Catheter urine | Male Medical |
| Kp 16 | August | XDR | Sputum | ICU |
| Kp 17 | August | ESBL | Catheter urine | Pediatrics |
| Kp 18 | August | ESBL | Urine | Emergency |
| Kp 19 | August | ESBL | Urine | Pediatrics |
| Kp 20 | August | ESBL | Urine | Pediatrics |
| Kp 21 | August | ESBL | urine | Emergency |
| Kp 22 | September | XDR | Wound | ICU |
| Kp 23 | September | ESBL | Blood culture | Emergency |
| Kp 24 | September | XDR | Skin | Emergency |
| Kp 25 | September | ESBL | Peritoneal fluid | Male Medical |
| Kp 26 | September | ESBL | wound | Surgery |
| Kp 27 | September | ESBL | Urine | Emergency |
| Kp 28 | September | XDR | Urine | Male Medical |
| Kp 29 | September | XDR | Urine | Day Care |
| Kp 30 | September | XDR | Wound | Male Medical |
| Kp 31 | September | XDR | Sputum | ICU |
| Kp 32 | September | XDR | bronchial wash | Emergency |
| Kp 33 | September | ESBL | wound | Female Medical |
| Kp 34 | September | ESBL | Urine | Pediatrics |
| Kp 35 | September | ESBL | Blood culture | Female Medical |
| Kp 36 | September | ESBL | Urine | Emergency |
| Kp 37 | September | ESBL | Urine | Urology |
| Kp 38 | October | ESBL | Urine | Emergency |
| Kp 39 | October | ESBL | Urine | Emergency |
| Kp 40 | October | ESBL | Tracheal aspirate | Neonatal unit |
| Kp 41 | October | XDR | Wound | Male Medical |
| Kp 42 | October | ESBL | Blood culture | Neonatal unit |
| Kp 43 | October | XDR | Tracheal aspirate | Male Medical |
| Kp 44 | October | XDR | Urine | Male Medical |
| Kp 45 | October | XDR | Tracheal aspirate | Male Medical |
| Kp 46 | October | ESBL | Tracheal aspirate | Neonatal unit |
| Kp 47 | October | ESBL | Urine | Female Medical |
| Kp 48 | October | XDR | Urine | Surgery |
| Kp 49 | October | XDR | Urine | Male Medical |
| Kp 50 | October | XDR | Urine | ICU |
| Zone Diameter Breakpoints (mm) | ||||
|---|---|---|---|---|
| Antibiotic | Disk Content | Susceptible | Intermediate | Resistant |
| Ampicillin | AMP 10 µg | ≥17 | 14–16 | ≤13 |
| Piperacillin–tazobactam | TZP 110 µg | ≥21 | 18–20 | ≤17 |
| Cefepime | FEP 30 µg | ≥25 | 19–24 | ≤18 |
| Cefotaxime | CTX 30 µg | ≥26 | 23–25 | ≤22 |
| Cefoxitin | FOX 30 µg | ≥18 | 15–17 | ≤14 |
| Ceftazidime | CAZ 30 µg | ≥21 | 18–20 | ≤17 |
| Imipenem | IMP 10 µg | ≥23 | 20–22 | ≤19 |
| Meropenem | MEM 10 µg | ≥23 | 20–22 | ≤18 |
| Gentamicin | CN 30 µg | ≥15 | 13–14 | ≤12 |
| Amikacin | AK 10 µg | ≥17 | 15–16 | ≤14 |
| Ciprofloxacin | CIP 5 µg | ≥31 | 21–30 | ≤20 |
| Step | Temperature | Time |
|---|---|---|
| Initial denaturation | 95 °C | 2 min |
| Denaturation | 95 °C | 30 s |
| Annealing | 30 s | |
| Extension | 72 °C | 1 min/kb |
| Final extension | 72 °C | 10 min |
| Hold | 15 °C |
| Gene | Annealing Temperature | Nucleotide Sequence (5′-3′) | Expected Size | |
|---|---|---|---|---|
| Class 1 integrase gene | Intl1 | 56 °C | IntI1F (ACGAGCGCAAGGTTTCGGT) IntI1R (GAAAGGTCTGGTCATACATG) | 565 |
| Class 2 integrase gene | Intl2 | 52 °C | IntI2F (GTGCAACGCATTTTGCAGG) IntI2R (CAACGGAGTCATGCAGATG) | 403 |
| Class 3 integrase gene | Intl3 | 57 °C | IntI3F (CATTTGTGTTGTGGACGGC) IntI3R (GACAGATACGTGTTTGGCAA) | 717 |
| Variable regions | 52 °C | 5′-CS (GGCATCCAAGCAGCAAG) 3′-CS (AAGCAGACTTGACCTGAT) | Uncertain |
| Isolate | AMP | CTX | FEP | CIP | CAZ | TZP | FOX | IPM | MEM | CN | AK | CL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kp 1 | R | R | R | R | R | R | R | R | R | R | I | S |
| Kp 2 | R | R | R | R | R | R | R | R | R | S | I | R |
| Kp 5 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 6 PDR | R | R | R | R | R | R | R | R | R | R | R | R |
| Kp 7 | R | R | R | R | R | I | R | S | S | R | R | S |
| Kp 11 PDR | R | R | R | R | R | R | R | R | R | R | R | R |
| Kp 13 | R | R | R | R | R | R | R | I | R | R | R | S |
| Kp 15 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 16 | R | R | R | R | R | R | R | R | R | S | S | S |
| Kp 22 | R | R | R | R | R | S | R | R | R | S | S | R |
| Kp 24 | R | R | R | R | R | R | R | R | R | S | S | S |
| Kp 28 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 29 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 30 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 31 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 32 | R | R | R | R | I | R | R | R | R | R | R | S |
| Kp 41 | R | R | R | R | R | R | R | R | R | R | S | S |
| Kp 43 | R | R | R | R | R | R | R | R | R | R | S | S |
| Kp 44 | R | R | R | R | R | R | R | R | R | R | I | S |
| Kp 45 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 48 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 49 | R | R | R | R | R | R | R | R | R | R | R | S |
| Kp 50 | R | R | R | R | R | R | R | R | R | R | S | R |
| Demographic and Clinical Characteristics | Case Patients, n = 13 |
|---|---|
| Male, n (%) | 12 (92%) |
| Age | |
| Adults ≤ 24 years, n (%) | 1 (7.7%) |
| From 25–50 years, n (%) | 3 (23.1%) |
| More than 50 years, n (%) | 9 (69.2%) |
| Age at first positive culture | |
| Mean age of adults, years (range) | 59.7 (20–86) |
| Length of stay | |
| Median length of stay after a first positive culture, days (range) | 48 (1–134) |
| Hospital location | |
| Intensive Care Unit (ICU), n (%) | 8 (61.5%) |
| Intermediate care ward, n (%) | 5 (83.5%) |
| Isolate (Kp) | PhoP | PhoQ |
|---|---|---|
| Kp 6 and Kp 11 | 3 SNPs | −Ve |
| (Gln147His) | ||
| (Gln131Glu) | ||
| (Pro129Leu) | ||
| Kp 22 and Kp 50 | 3 SNPs | −Ve |
| (Val130Glu) (Gln147His) | ||
| (Gln131Glu) |
| K. pneumoniae (n = 23) | Phenotypic Resistance | Positive Genes | Level of Agreement Genotype with Phenotypic Expression % | |
|---|---|---|---|---|
| Aminoglycosides | Amikacin | 9 | 21 | 42.90% |
| Gentamicin | 15 | 21 | 71.40% | |
| Quinolones | 23 | 23 | 100% | |
| Carbapenems | 14 | 14 | 100% | |
| Cephalosporins | 23 | 23 | 100% | |
| Isolate | ST Type | Gene Cassette |
|---|---|---|
| Kp 40 | 37 | dfrA12, ant1 |
| Kp 49 | 147 | Arr3, ereA2, aadA, cmlA1 |
| Kp 21 | 45 | dfra12, ant1 |
| Kp 41 | 395 | dfrA14, |
| Kp 43 | 395 | dfrA12, APH(3″)-Ia |
| Kp 44 | 395 | dfrA14 |
| Kp 50 | 395 | dfrA14 |
| Kp 16 | 395 | In0 |
| Kp 22 | 395 | In0 |
| Kp 5 | 231 | aac(6′)-Ib, arr2 |
| Kp 6 | 231 | dfrA12, emrE, ant1 |
| Kp 7 | 231 | dfrA12, emrE, ant1 |
| Kp 10 | 231 | dfrA5 |
| Kp 11 | 231 | aacA4, cat1, ant1 |
| Kp 15 | 231 | aacA4, emrE, ant1 |
| Kp 28 | 231 | Ant1, erm, cat1 |
| Kp 30 | 231 | aacA4 |
| Kp 45 | 231 | dfrA14 |
| Kp 42 | 405 | dfrA14 |
| Kp 46 | 405 | dfrA14 |
| Kp 27 | 280 | dfrA14 |
| Kp 25 | 1741 | In0 |
| Kp 37 | 1710 | dfrA14 |
| Isolate (KP) | Plasmid | Size (bp) | Replicon | Resistance Genes |
|---|---|---|---|---|
| 5, 6, 7, 10, 11, 15, 16, 22, 28, 30, 41, 43, 44, 45, 50 | pKPQIL-IT | 115,300 | IncFIB (QIL) | blaTEM-1, blaKPC-3 |
| 5, 6, 11, 15, 16, 22, 28, 30, 41, 43, 44, 45, 49, 50 | pKP3-A | 7605 | ColKP3 | BlaOXA-181 |
| 5, 6, 7, 15, 28, 30, 45 | pAMA1167-NDM-5 | 11,310 | IncFII (pAMA1167-NDM-5) | aadA5, aadA2, aac(3)-IId, aph(6)-Id, aph(3″)-Ib, aac(6′)-Ib-cr5, blaNDM-5, blaOXA-1, blaCTX-M-15-1, blaTEM-1 dfrA17, dfrA12, Mph(A), Sul1, Sul2, emrE, tet(b), tet(C), cat |
| 10, 25, 27, 37, 42, 46 | pKPN-IT | 208,191 | IncFIB (K) | aadA2, cat, Mph(A), Sul1, dfrA12 |
| 21, 40 | pCAV1099-14 | 113,992 | IncFIB(K) (pCAV1099-14) | dfrA19, APH(3″)-Ib, APH(3′)-Ia, QnrB52 |
| 41, 50 | pNDM-MAR | 267,242 | IncFIB (pNDM-Mar) IncHI1B (pNDM-Mar) | aac(6′)-Ib, blaOXA-1, blaNDM-1, cat, QnrB1 |
| 27, 49 | pK245 | 98,264 | IncR | aacC2, strA, strB, dfrA14, catA2, Qnrs, blaSH2A, blaTEM |
| 49 | pC15-1a | 92,353 | IncFII | aac(6′)-Ib, aac(3)-II, blaTEM-1, blaOXA-1, blaCTX-M-15-1, tet(A) |
| 37 | pBK30683 | 139,941 | FIA (pBK30683) | dfrA14, StrA, StrB, blaTEM-1, blaOXA-9, blaKPC-3 Sul2,ant(3″)-Ia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AL-Muzahmi, M.; Rizvi, M.; AL-Quraini, M.; AL-Muharrmi, Z.; AL-Jabri, Z. Comparative Genomic Analysis Reveals the Emergence of ST-231 and ST-395 Klebsiella pneumoniae Strains Associated with the High Transmissibility of blaKPC Plasmids. Microorganisms 2023, 11, 2411. https://doi.org/10.3390/microorganisms11102411
AL-Muzahmi M, Rizvi M, AL-Quraini M, AL-Muharrmi Z, AL-Jabri Z. Comparative Genomic Analysis Reveals the Emergence of ST-231 and ST-395 Klebsiella pneumoniae Strains Associated with the High Transmissibility of blaKPC Plasmids. Microorganisms. 2023; 11(10):2411. https://doi.org/10.3390/microorganisms11102411
Chicago/Turabian StyleAL-Muzahmi, Muna, Meher Rizvi, Munawr AL-Quraini, Zakariya AL-Muharrmi, and Zaaima AL-Jabri. 2023. "Comparative Genomic Analysis Reveals the Emergence of ST-231 and ST-395 Klebsiella pneumoniae Strains Associated with the High Transmissibility of blaKPC Plasmids" Microorganisms 11, no. 10: 2411. https://doi.org/10.3390/microorganisms11102411
APA StyleAL-Muzahmi, M., Rizvi, M., AL-Quraini, M., AL-Muharrmi, Z., & AL-Jabri, Z. (2023). Comparative Genomic Analysis Reveals the Emergence of ST-231 and ST-395 Klebsiella pneumoniae Strains Associated with the High Transmissibility of blaKPC Plasmids. Microorganisms, 11(10), 2411. https://doi.org/10.3390/microorganisms11102411