
Changes in Microbiota Profile in the Proximal Remnant Intestine in Infants Undergoing Surgery Requiring Enterostomy

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Clinical Data Collection
2.2. Enterostomy Effluent Samples Collection and DNA Extraction
2.3. Quantitative Analysis of Faecal Microbiota by RT-PCR
2.4. Microbial 16S rRNA Sequence Analysis
2.5. Bioinformatic Analysis
2.6. Statistical Analysis
3. Results
3.1. Clinical and Demographic Characteristics of Infants
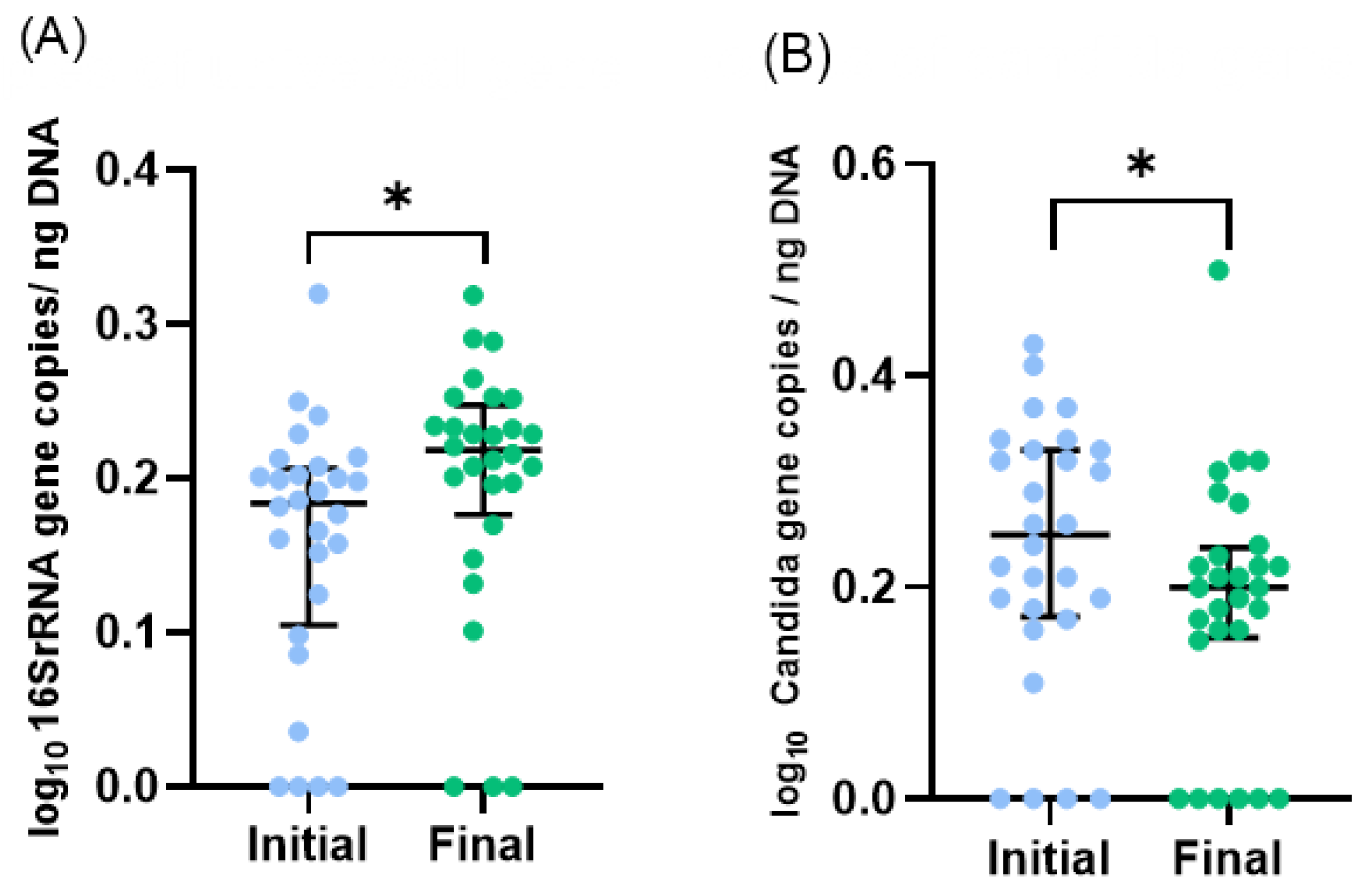
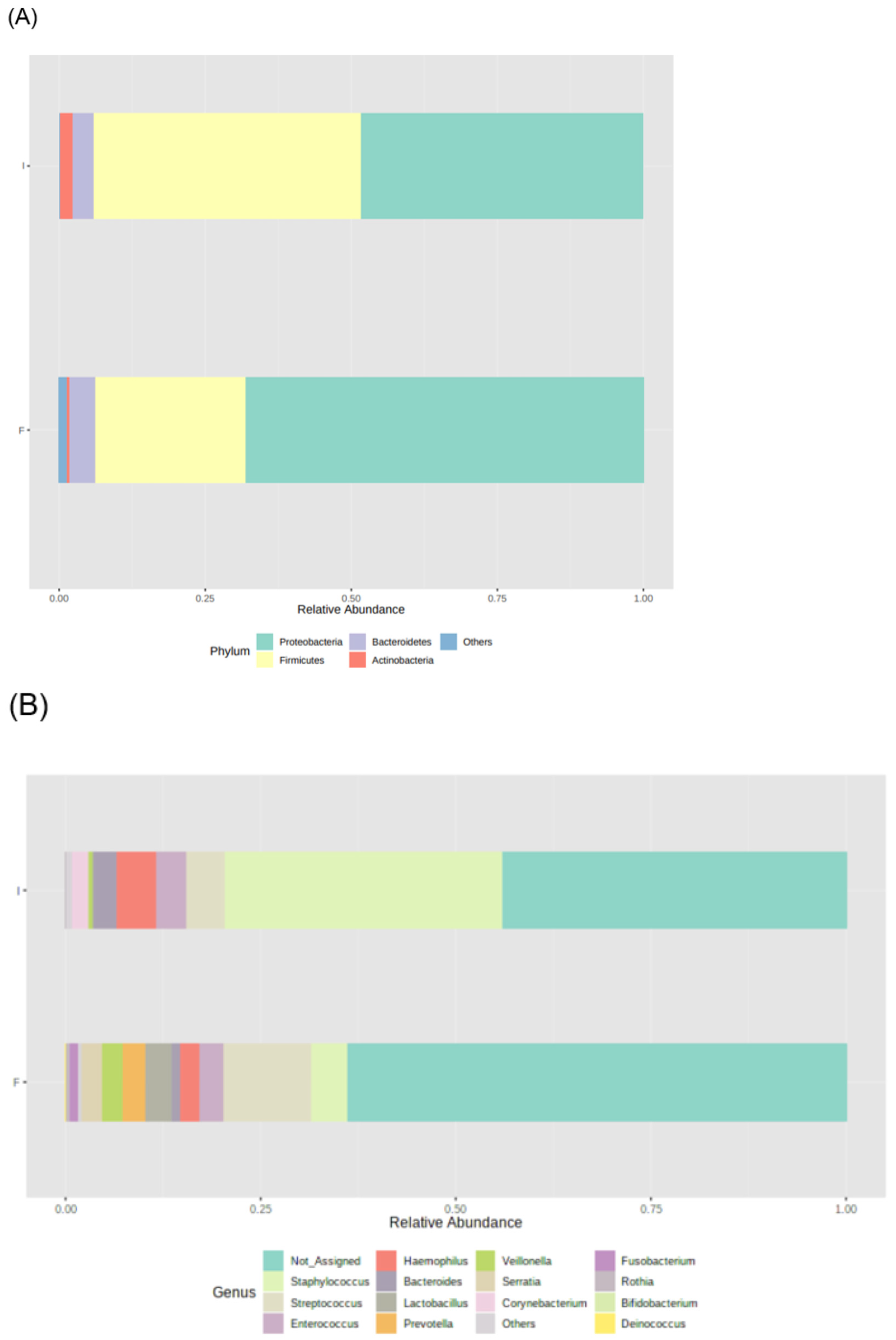
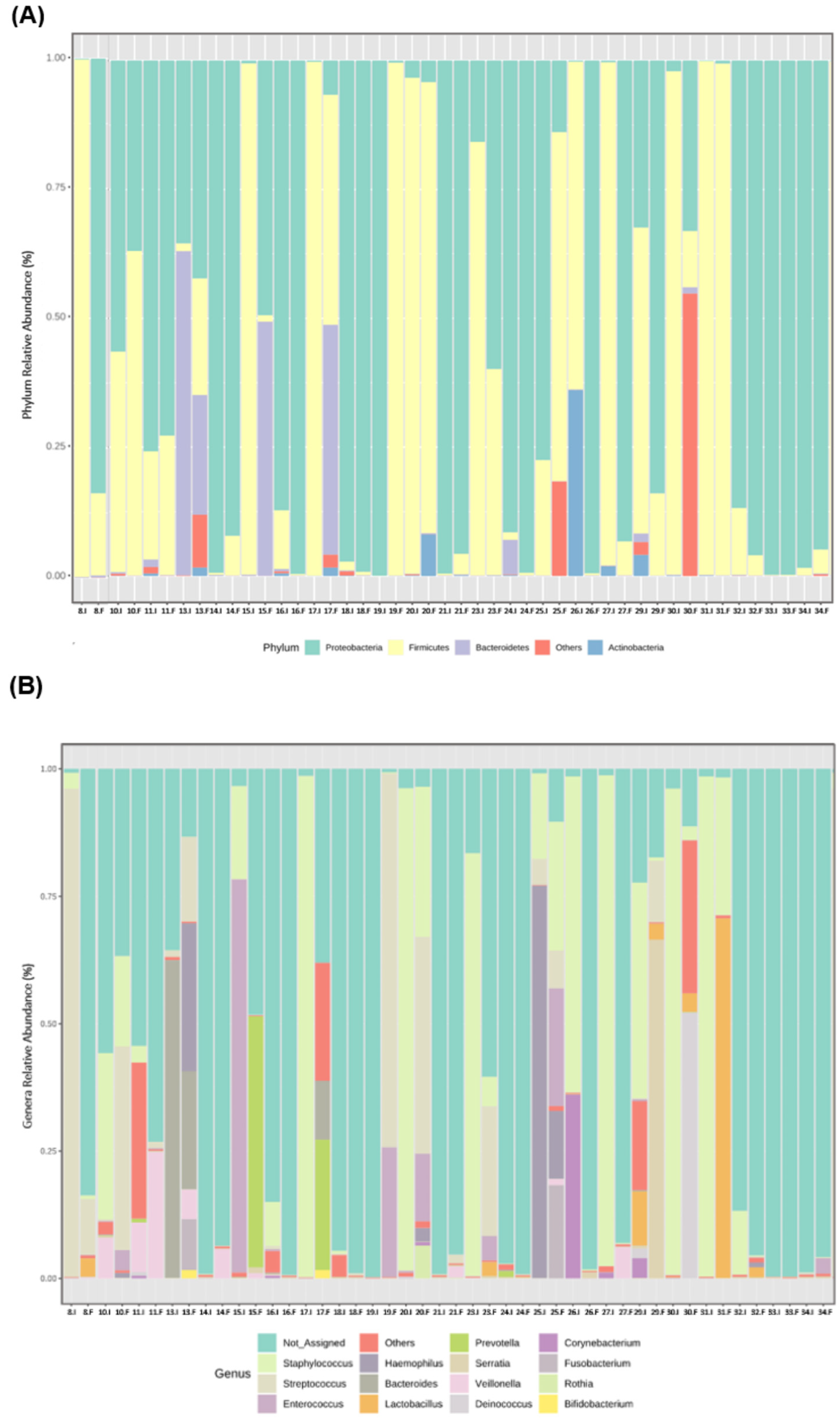
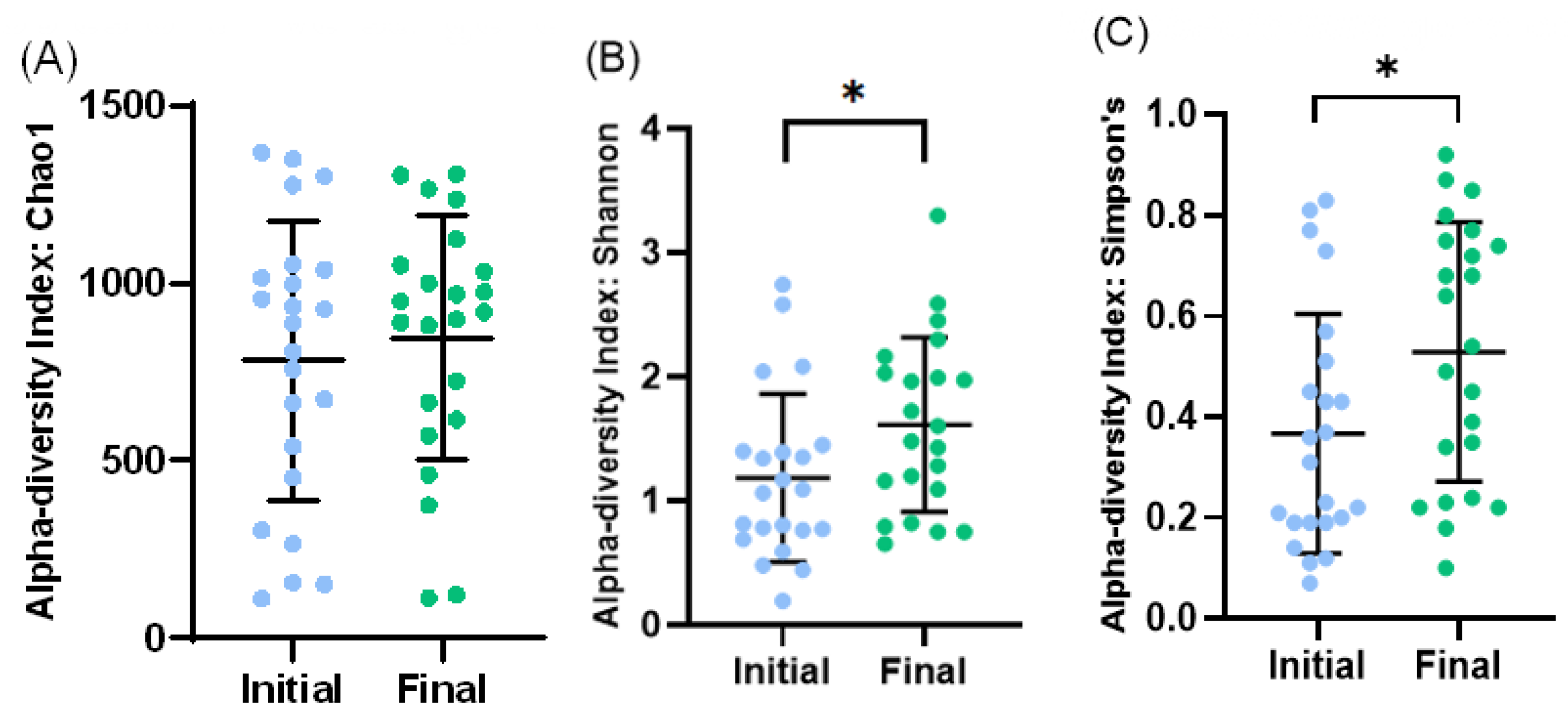
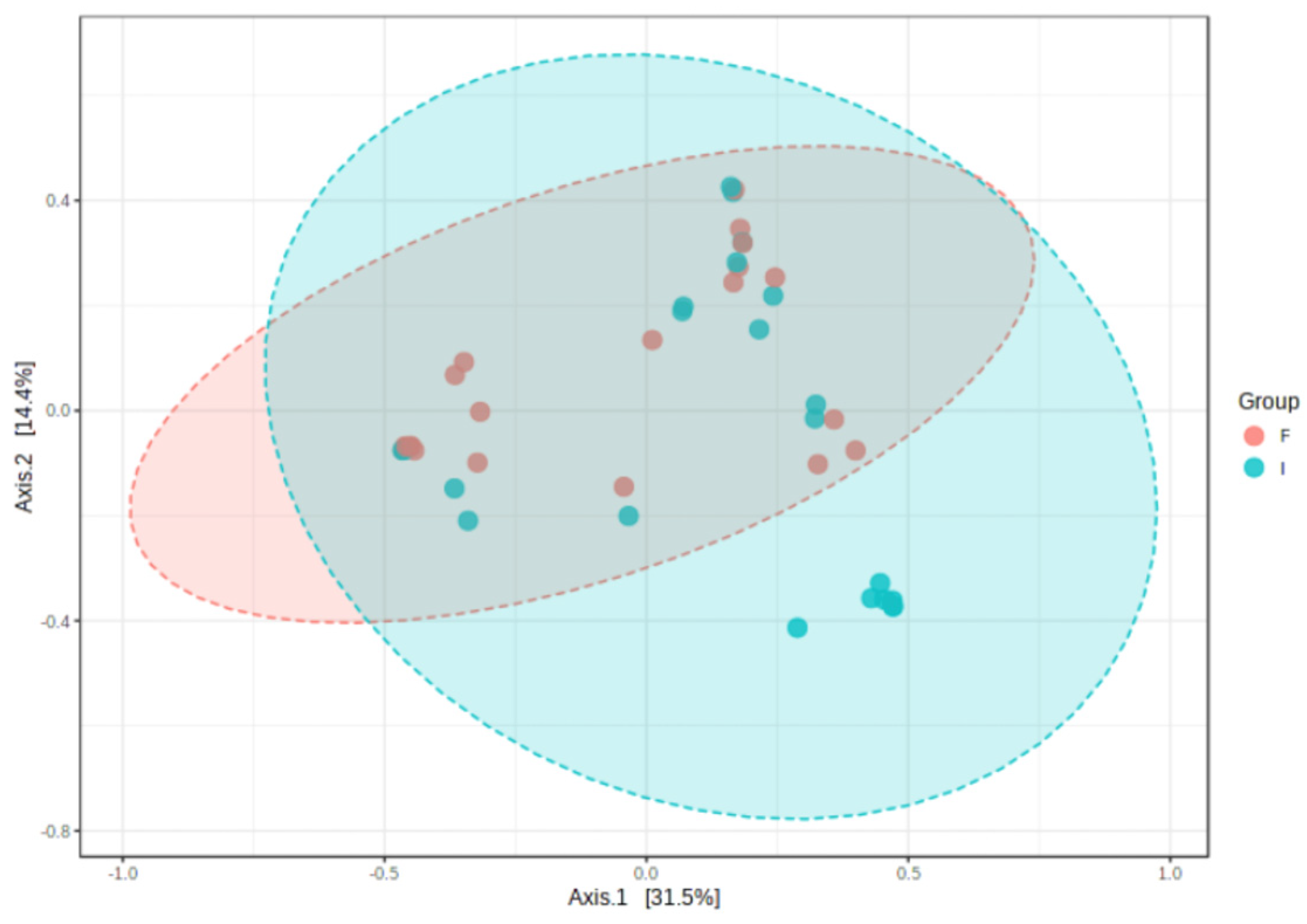
3.2. Microbiota Profile of Enterostomy Effluents
3.3. Correlations between Microbiota and Clinical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguilar-lopez, M.; Dinsmoor, A.M.; Donovan, S.M. A systematic review of the factors influencing microbial colonization of the preterm infant gut. Gut Microbes 2021, 13, 1884514. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Ye, Y.; Wang, F.; Chen, F.; Zheng, C. The role of microbiota in infant health: From early life to adulthood. Front. Immunol. 2021, 12, 708472. [Google Scholar] [CrossRef]
- Cerdó, T.; García-Santos, J.A.; Rodríguez-Pöhnlein, A.; García-Ricobaraza, M.; Nieto-Ruíz, A.; G Bermúdez, M.; Campoy, C. Impact of total parenteral nutrition on gut microbiota in pediatric population suffering intestinal disorders. Nutrients 2022, 14, 4691. [Google Scholar] [CrossRef] [PubMed]
- Neelis, E.G.; de Koning, B.A.E.; Hulst, J.M.; Papadopoulou, R.; Kerbiriou, C.; Rings, E.H.H.M.; Wijnen, R.M.H.; Nichols, B.; Gerasimidis, K. Gut microbiota and its diet-related activity in children with intestinal failure receiving long-term parenteral nutrition. JPEN J. Parenter. Enter. Nutr. 2022, 46, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Saeed, N.K.; Al-Beltagi, M.; Bediwy, A.S.; El-Sawaf, Y.; Toema, O. Gut microbiota in various childhood disorders: Implication and indications. World J. Gastroenterol. 2022, 28, 1875–1901. [Google Scholar] [CrossRef]
- Korpela, K.; Blakstad, E.W.; Moltu, S.J.; Strømmen, K.; Nakstad, B.; Rønnestad, A.E.; Brække, K.; Iversen, P.O.; Drevon, C.A.; de Vos, W. Intestinal microbiota development and gestational age in preterm neonates. Sci. Rep. 2018, 8, 2453. [Google Scholar] [CrossRef]
- Sanidad, K.Z.; Zeng, M.Y. Neonatal gut microbiome and immunity. Curr. Opin. Microbiol. 2020, 56, 30–37. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The first microbial colonizers of the human gut: Composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef]
- Yang, M.; Du, J.; Yang, Q.; Dou, W.; Jiang, M.; Hei, M. Influence of family integrated care on the intestinal microbiome of preterm infants with necrotizing enterocolitis and enterostomy: A preliminary study. Front. Pediatr. 2021, 9, 678254. [Google Scholar] [CrossRef]
- Younge, N.; Yang, Q.; Seed, P.C. Enteral high fat-polyunsaturated fatty acid blend alters the pathogen composition of the intestinal microbiome in premature infants with an enterostomy. J. Pediatr. 2017, 181, 93–101.e6. [Google Scholar] [CrossRef]
- James, S.A.; Phillips, S.; Telatin, A.; Baker, D.; Ansorge, R.; Clarke, P.J.; Hall, L.R.; Carding, S. Preterm infants harbour a rapidly changing mycobiota that includes candida pathobionts. J. Fungi 2020, 6, 273. [Google Scholar] [CrossRef]
- Barreiros-Mota, I.; Marques, C.; Faria, A.; Neto, M.T.; Cordeiro-Ferreira, G.; Virella, D.; Pita, A.; Pereira-da-Silva, L.; Calhau, C. Colonisation of the proximal intestinal remnant in newborn infants with enterostomy: A longitudinal study protocol. BMJ Open 2019, 9, e028916. [Google Scholar] [CrossRef]
- Kaiser, A.M.; Israelit, S.; Klaristenfeld, D.; Selvindoss, P.; Vukasin, P.; Ault, G.; Beart, R.W. Morbidity of ostomy takedown. J. Gastrointest. Surg. 2008, 12, 437–441. [Google Scholar] [CrossRef]
- Qian, T.; Zhu, H.; Zhu, L.; Chen, C.; Shen, C.; Zhang, R. Small intestinal microbiota composition and the prognosis of infants with ileostomy resulting from distinct primary diseases. BMC Gastroenterol. 2020, 20, 224. [Google Scholar] [CrossRef]
- Ambe, P.C.; Kurz, N.R.; Nitschke, C.; Odeh, S.F.; Möslein, G.; Zirngibl, H. Intestinal ostomy. Dtsch. Arztebl. Int. 2018, 115, 182–188. [Google Scholar] [CrossRef]
- Rao, S.C.; Esvaran, M.; Patole, S.K.; Simmer, K.N.; Gollow, I.; Keil, A.; Wemheuer, B.; Chen, L.; Conway, P.L. Gut microbiota in neonates with congenital gastrointestinal surgical conditions: A prospective study. Pediatr. Res. 2020, 88, 878–886. [Google Scholar] [CrossRef]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved bacterial 16S rRNA Gene (V4 and V4-5) and fungal internal transcribed spacer marker gene primers for microbial community surveys. mSystems 2015, 1, e0009-15. [Google Scholar] [CrossRef] [PubMed]
- Greengenes Database. Available online: https://greengenes.lbl.gov/Download/ (accessed on 7 July 2020).
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Jiwon, S.; Guevarra, R.; Lee, J.; Kim, D.; Seol, K.H.; Lee, J.H.; Kim, H.B.; Isaacson, R. Deciphering diversity indices for a better understanding of microbial communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed]
- Schober, P.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Zeng, S.; Ying, J.; Li, S.; Qu, Y.; Mu, D.; Wang, S. First 1000 days and beyond after birth: Gut microbiota and necrotizing enterocolitis in preterm infants. Front. Microbiol. 2022, 13, 905380. [Google Scholar] [CrossRef]
- Mills, S.; Lane, J.A.; Smith, G.J.; Grimaldi, K.A.; Ross, R.P.; Stanton, C. Precision nutrition and the microbiome part ii: Potential opportunities and pathways to commercialisation. Nutrients 2019, 11, 1468. [Google Scholar] [CrossRef]
- Barrett, E.; Guinane, C.M.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.P.; O’Toole, P.W.; Fitzgerald, G.F.; Cotter, P.D.; Ross, R.P.; Stanton, C. Microbiota diversity and stability of the preterm neonatal ileum and colon of two infants. Microbiologyopen 2013, 2, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.A.; Sohn, K. The microbiota of the extremely preterm infant. Clin. Perinatol. 2017, 44, 407–427. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.; Fernandes, I.; Meireles, M.; Faria, A.; Spencer, J.P.E.; Mateus, N.; Calhau, C. Gut microbiota modulation accounts for the neuroprotective properties of anthocyanins. Sci. Rep. 2018, 8, 11341. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.; Meireles, M.; Norberto, S.; Leite, J.; Freitas, J.; Pestana, D.; Faria, A.; Calhau, C. High-fat diet-induced obesity Rat model: A comparison between Wistar and Sprague-Dawley Rat. Adipocyte 2015, 5, 11–21. [Google Scholar] [CrossRef]
- Gosiewski, T.; Salamon, D.; Szopa, M.; Sroka, A.; Malecki, M.T.; Bulanda, M. Quantitative evaluation of fungi of the genus Candida in the feces of adult patients with type 1 and 2 diabetes—A pilot study. Gut Pathog. 2014, 6, 43. [Google Scholar] [CrossRef]
- Moles, L.; Gómez, M.; Moroder, E.; Bustos, G.; Melgar, A.; del Campo, R.; Rodríguez, J.M. Staphylococcus epidermidis in feedings and feces of preterm neonates. PLoS ONE 2020, 15, e0227823. [Google Scholar] [CrossRef]
- Thongaram, T.; Hoeflinger, J.L.; Chow, J.M.; Miller, M.J. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J. Dairy Sci. 2017, 100, 7825–7833. [Google Scholar] [CrossRef]
- Gupta, R.W.; Tran, L.; Norori, J.; Ferris, M.J.; Eren, A.M.; Taylor, C.M.; Dowd, S.E.; Penn, D. Histamine-2 receptor blockers alter the fecal microbiota in premature infants. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 397–400. [Google Scholar] [CrossRef]
- Fu, X.; Li, S.; Jiang, Y.; Hu, X.; Wu, H. Necrotizing enterocolitis and intestinal microbiota: The timing of disease and combined effects of multiple species. Front. Pediatr. 2021, 9, 657349. [Google Scholar] [CrossRef]
- Imhann, F.; Bonder, M.J.; Vila, A.V.; Fu, J.; Mujagic, Z.; Vork, L.; Tigchelaar, E.F.; Jankipersadsing, S.A.; Cenit, M.C.; Harmsen, H.J.; et al. Proton pump inhibitors affect the gut microbiome. Gut 2016, 65, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, Y.; Iwanaka, T.; Sugiyama, M.; Komura, M.; Takahashi, T.; Yuki, N.; Morotomi, M.; Tanaka, R. Early use of probiotics is important therapy in infants with severe congenital anomaly. Pediatr. Int. 2010, 5, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Hussey, S.G.; Ryan, C.A.; O’Neill, M.; Fitzgerald, G.; Stanton, C.; Ross, R.P. Presence of two Lactobacillus and Bifidobacterium probiotic strains in the neonatal ileum. ISME J. 2008, 2, 83–91. [Google Scholar] [CrossRef]
- Rromano-Keeler, J.; Moore, D.J.; Wang, C.; Brucker, R.M.; Fonnesbeck, C.; Slaughter, J.C.; Li, H.; Curran, D.P.; Meng, S.; Correa, H.; et al. Early life establishment of site-specific microbial communities in the gut. Gut Microbes 2014, 5, 192–201. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | |
|---|---|
| Male sex, n (%) | 17 (59%) |
| C-section delivery, n (%) | 16 (55%) |
| Gestational age, weeks, median (IQR) | 33 (27–37) |
| Birth weight, g, median (IQR) | 1779 (884–2485) |
| Apgar score at 5 min, median (IQR) | 9 (8–10) |
| Peripartum antibiotics, n (%) | 6 (23%) * |
| Surgical condition - CMGIT, n (%) - NEC, n (%) - SIP, n (%) | 12 (41%) 13 (45%) 4 (13.8%) |
| Level of enterostomy - Small intestine **, n (%) - Colon, n (%) | 26 (90%) 3 (10%) |
| Age at enterostomy, postnatal days, median (IQR) | 2 (1–8) |
| Body weight gain during the study period, g, median (IQR) | 332 (192–466) |
| Antibiotic therapy, n (%) | 26 (90%) |
| Antifungal therapy, n (%) | 9 (31%) |
| H2-blockers and/or proton pump inhibitors, n (%) | 8 (28%) |
| Sepsis confirmed by blood culture, n (%) | 11 (38%) |
| Type of nutrition post enterostomy
- Human milk, n (%) - Formula, n (%) - Exclusive parenteral nutrition, n (%) | 14 (48%) 13 (45%) 2 (7%) |
| Staphylococcus | Shannon’s Index | Simpson’s Index | Total Bacteria Number | |
|---|---|---|---|---|
| NEC | −0.436 * | 0.468 * | 0.483 * | 0.004 |
| H2-blockers and/or proton pump inhibitors | 0.043 | 0.198 | 0.200 | 0.385 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barreiros-Mota, I.; R. Araújo, J.; Marques, C.; Sousa, L.; Morais, J.; Castela, I.; Faria, A.; Neto, M.T.; Cordeiro-Ferreira, G.; Virella, D.; et al. Changes in Microbiota Profile in the Proximal Remnant Intestine in Infants Undergoing Surgery Requiring Enterostomy. Microorganisms 2023, 11, 2482. https://doi.org/10.3390/microorganisms11102482
Barreiros-Mota I, R. Araújo J, Marques C, Sousa L, Morais J, Castela I, Faria A, Neto MT, Cordeiro-Ferreira G, Virella D, et al. Changes in Microbiota Profile in the Proximal Remnant Intestine in Infants Undergoing Surgery Requiring Enterostomy. Microorganisms. 2023; 11(10):2482. https://doi.org/10.3390/microorganisms11102482
Chicago/Turabian StyleBarreiros-Mota, Inês, João R. Araújo, Cláudia Marques, Laura Sousa, Juliana Morais, Inês Castela, Ana Faria, Maria Teresa Neto, Gonçalo Cordeiro-Ferreira, Daniel Virella, and et al. 2023. "Changes in Microbiota Profile in the Proximal Remnant Intestine in Infants Undergoing Surgery Requiring Enterostomy" Microorganisms 11, no. 10: 2482. https://doi.org/10.3390/microorganisms11102482
APA StyleBarreiros-Mota, I., R. Araújo, J., Marques, C., Sousa, L., Morais, J., Castela, I., Faria, A., Neto, M. T., Cordeiro-Ferreira, G., Virella, D., Pita, A., Pereira-da-Silva, L., & Calhau, C. (2023). Changes in Microbiota Profile in the Proximal Remnant Intestine in Infants Undergoing Surgery Requiring Enterostomy. Microorganisms, 11(10), 2482. https://doi.org/10.3390/microorganisms11102482