
A Review on Zoonotic Pathogens Associated with Non-Human Primates: Understanding the Potential Threats to Humans

Abstract
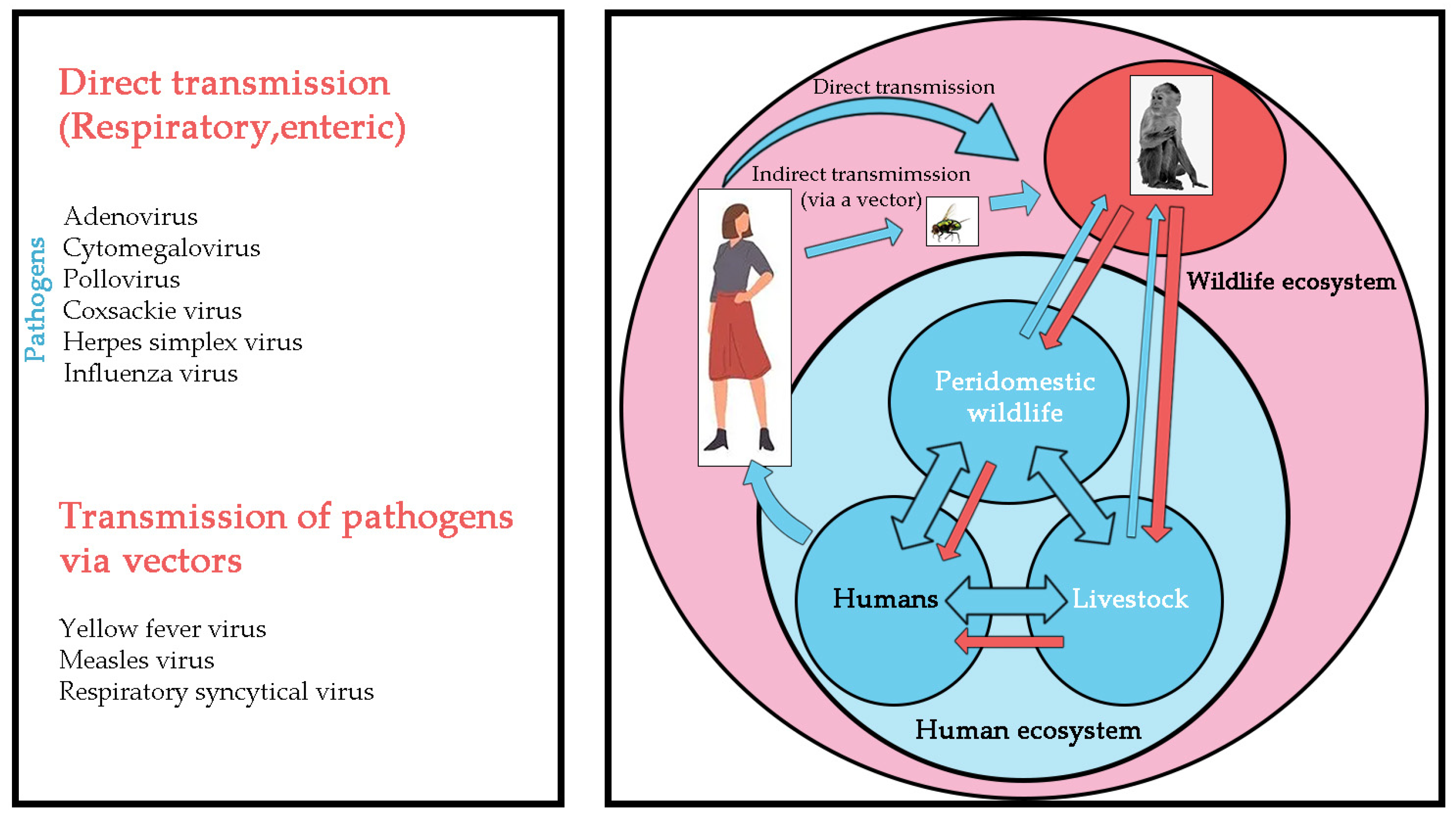
:1. Introduction
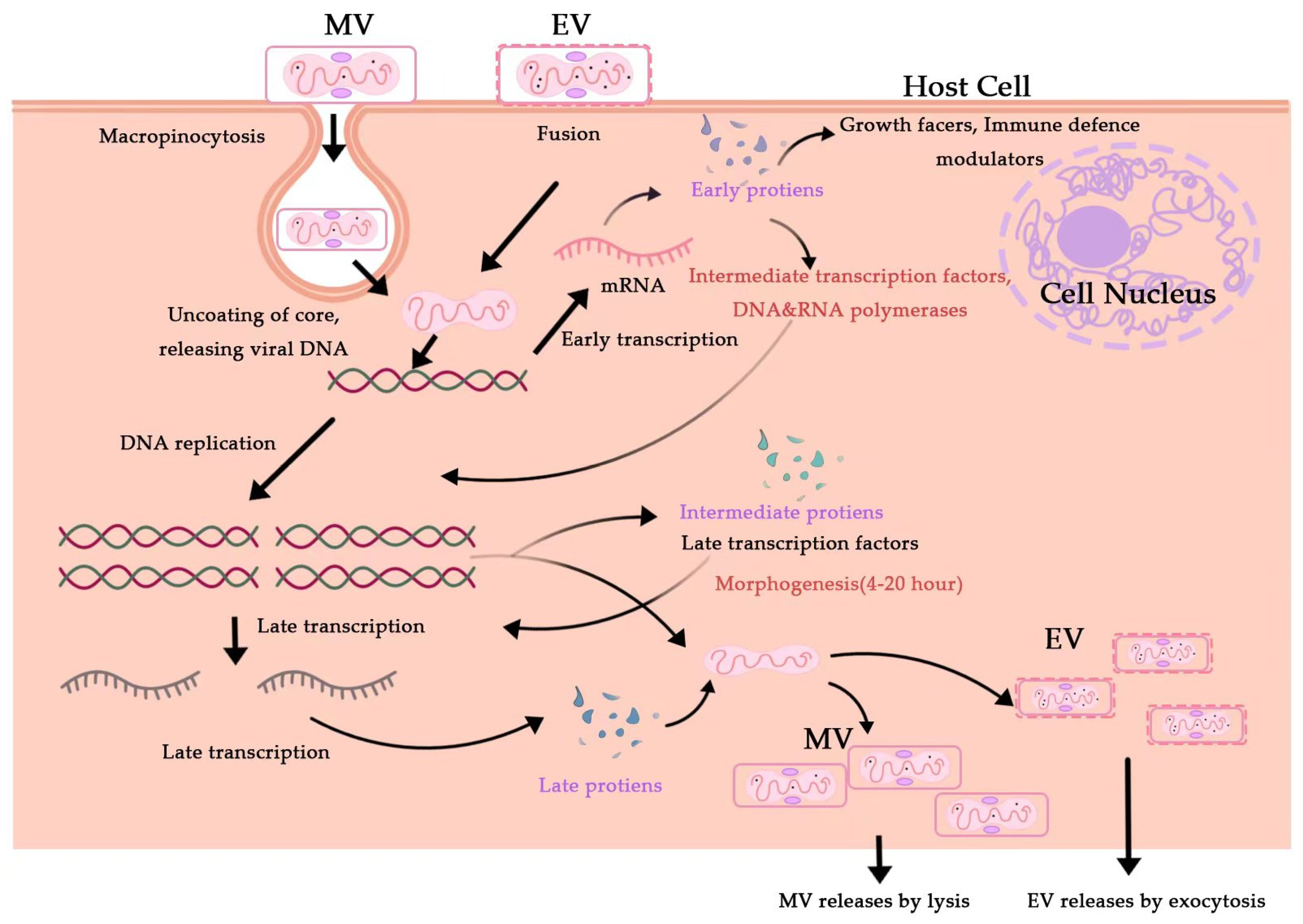
2. Monkeypox Virus (MPXV)
3. Rhesus Herpesviruses
4. Simian Retroviruses
5. Ebola Virus (EBOV)
6. Simian Hemorrhagic Fever Virus (SHFV)
7. Simian Adenovirus (SADV)
8. Simian Parvovirus (SPV)
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Devaux, C.A.; Mediannikov, O.; Medkour, H.; Raoult, D. Infectious Disease Risk Across the Growing Human-Non Human Primate Interface: A Review of the Evidence. Front. Public Health 2019, 7, 305. [Google Scholar] [CrossRef] [Green Version]
- Brashares, J.S.; Golden, C.D.; Weinbaum, K.Z.; Barrett, C.B.; Okello, G.V. Economic and geographic drivers of wildlife consumption in rural Africa. Proc. Natl. Acad. Sci. USA 2011, 108, 13931–13936. [Google Scholar] [CrossRef] [Green Version]
- Shanee, N.; Mendoza, A.P.; Shanee, S. Diagnostic overview of the illegal trade in primates and law enforcement in Peru. Am. J. Primatol. 2017, 79, e22516. [Google Scholar] [CrossRef]
- Estrada, A.; Garber, P.A.; Rylands, A.B.; Roos, C.; Fernandez-Duque, E.; Di Fiore, A.; Nekaris, K.A.; Nijman, V.; Heymann, E.W.; Lambert, J.E.; et al. Impending extinction crisis of the world’s primates: Why primates matter. Sci. Adv. 2017, 3, e1600946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabuga, A.I.; El Zowalaty, M.E. A review of the monkeypox virus and a recent outbreak of skin rash disease in Nigeria. J. Med. Virol. 2019, 91, 533–540. [Google Scholar] [CrossRef]
- Espinosa, S.; Branch, L.C.; Cueva, R. Road development and the geography of hunting by an Amazonian indigenous group: Consequences for wildlife conservation. PLoS ONE 2014, 9, e114916. [Google Scholar] [CrossRef] [Green Version]
- Dunay, E.; Apakupakul, K.; Leard, S.; Palmer, J.L.; Deem, S.L. Pathogen Transmission from Humans to Great Apes is a Growing Threat to Primate Conservation. Ecohealth 2018, 15, 148–162. [Google Scholar] [CrossRef]
- Lai, C.C.; Hsu, C.K.; Yen, M.Y.; Lee, P.I.; Ko, W.C.; Hsueh, P.R. Monkeypox: An emerging global threat during the COVID-19 pandemic. J. Microbiol. Immunol. Infect. 2022, 55, 787–794. [Google Scholar] [CrossRef]
- Cohen-Gihon, I.; Israeli, O.; Shifman, O.; Erez, N.; Melamed, S.; Paran, N.; Beth-Din, A.; Zvi, A. Identification and Whole-Genome Sequencing of a Monkeypox Virus Strain Isolated in Israel. Microbiol. Resour. Announc. 2020, 9, e01524-19. [Google Scholar] [CrossRef] [Green Version]
- Ferdous, J.; Barek, M.A.; Hossen, M.S.; Bhowmik, K.K.; Islam, M.S. A review on monkeypox virus outbreak: New challenge for world. Health Sci. Rep. 2023, 6, e1007. [Google Scholar] [CrossRef]
- Weigler, B.J. Biology of B virus in macaque and human hosts: A review. Clin. Infect. Dis. 1992, 14, 555–567. [Google Scholar] [CrossRef]
- Bennett, R.S.; Logue, J.; Liu, D.X.; Reeder, R.J.; Janosko, K.B.; Perry, D.L.; Cooper, T.K.; Byrum, R.; Ragland, D.; St Claire, M.; et al. Kikwit Ebola Virus Disease Progression in the Rhesus Monkey Animal Model. Viruses 2020, 12, 753. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.J.; Burbelo, P.D.; Gill, R.B.; Sauri, M.A.; Schmitt, J.M.; Cohen, J.I. A seroprevalence study of primate workers for asymptomatic rhesus cytomegalovirus infection. J. Clin. Virol. 2014, 60, 411–413. [Google Scholar] [CrossRef] [Green Version]
- Burwitz, B.J.; Malouli, D.; Bimber, B.N.; Reed, J.S.; Ventura, A.B.; Hancock, M.H.; Uebelhoer, L.S.; Bhusari, A.; Hammond, K.B.; Espinosa Trethewy, R.G.; et al. Cross-Species Rhesus Cytomegalovirus Infection of Cynomolgus Macaques. PLoS Pathog. 2016, 12, e1006014. [Google Scholar] [CrossRef] [Green Version]
- Traina-Dorge, V.L.; Lorino, R.; Gormus, B.J.; Metzger, M.; Telfer, P.; Richardson, D.; Robertson, D.L.; Marx, P.A.; Apetrei, C. Molecular epidemiology of simian T-cell lymphotropic virus type 1 in wild and captive sooty mangabeys. J. Virol. 2005, 79, 2541–2548. [Google Scholar] [CrossRef] [Green Version]
- Montiel, N.A. An updated review of simian betaretrovirus (SRV) in macaque hosts. J. Med. Primatol. 2010, 39, 303–314. [Google Scholar] [CrossRef]
- Gardner, M.B.; Luciw, P.A.; Sawai, E.T.; Marthas, M.L.; Miller, C.J.; McChesney, M.B.; Lerche, N.W.; Pedersen, N.C. Simian retrovirus vaccines: Simian retrovirus and simian immunodeficiency lentivirus. AIDS Res. Hum. Retrovir. 1996, 12, 399–401. [Google Scholar] [CrossRef]
- Cai, Y.; Postnikova, E.N.; Bernbaum, J.G.; Yu, S.Q.; Mazur, S.; Deiuliis, N.M.; Radoshitzky, S.R.; Lackemeyer, M.G.; McCluskey, A.; Robinson, P.J.; et al. Simian hemorrhagic fever virus cell entry is dependent on CD163 and uses a clathrin-mediated endocytosis-like pathway. J. Virol. 2015, 89, 844–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornish, J.P.; Moore, I.N.; Perry, D.L.; Lara, A.; Minai, M.; Promeneur, D.; Hagen, K.R.; Virtaneva, K.; Paneru, M.; Buechler, C.R.; et al. Clinical Characterization of Host Response to Simian Hemorrhagic Fever Virus Infection in Permissive and Refractory Hosts: A Model for Determining Mechanisms of VHF Pathogenesis. Viruses 2019, 11, 67. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.E.; Liu, Z.; Gallinella, G.; Wong, S.; Mills, I.P.; O’Sullivan, M.G. Simian parvovirus infection: A potential zoonosis. J. Infect Dis. 2004, 190, 1900–1907. [Google Scholar] [CrossRef]
- Isidro, J.; Borges, V.; Pinto, M.; Sobral, D.; Santos, J.D.; Nunes, A.; Mixao, V.; Ferreira, R.; Santos, D.; Duarte, S.; et al. Phylogenomic characterization and signs of microevolution in the 2022 multi-country outbreak of monkeypox virus. Nat. Med. 2022, 28, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.H.; Price, J.T. The significant but understudied impact of pathogen transmission from humans to animals. Mt. Sinai J. Med. 2009, 76, 448–455. [Google Scholar] [CrossRef]
- Reynolds, M.G.; Doty, J.B.; McCollum, A.M.; Olson, V.A.; Nakazawa, Y. Monkeypox re-emergence in Africa: A call to expand the concept and practice of One Health. Expert Rev. Anti Infect. Ther. 2019, 17, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Durski, K.N.; McCollum, A.M.; Nakazawa, Y.; Petersen, B.W.; Reynolds, M.G.; Briand, S.; Djingarey, M.H.; Olson, V.; Damon, I.K.; Khalakdina, A. Emergence of Monkeypox—West and Central Africa, 1970–2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 306–310. [Google Scholar] [CrossRef] [PubMed]
- McCollum, A.M.; Damon, I.K. Human monkeypox. Clin. Infect. Dis. 2014, 58, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Sklenovska, N.; Van Ranst, M. Emergence of Monkeypox as the Most Important Orthopoxvirus Infection in Humans. Front. Public Health 2018, 6, 241. [Google Scholar] [CrossRef] [Green Version]
- Fine, P.E.; Jezek, Z.; Grab, B.; Dixon, H. The transmission potential of monkeypox virus in human populations. Int. J. Epidemiol. 1988, 17, 643–650. [Google Scholar] [CrossRef]
- Heymann, D.L.; Szczeniowski, M.; Esteves, K. Re-emergence of monkeypox in Africa: A review of the past six years. Br. Med. Bull. 1998, 54, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Doty, J.B.; Malekani, J.M.; Kalemba, L.N.; Stanley, W.T.; Monroe, B.P.; Nakazawa, Y.U.; Mauldin, M.R.; Bakambana, T.L.; Liyandja Dja Liyandja, T.; Braden, Z.H.; et al. Assessing Monkeypox Virus Prevalence in Small Mammals at the Human-Animal Interface in the Democratic Republic of the Congo. Viruses 2017, 9, 283. [Google Scholar] [CrossRef]
- Alakunle, E.; Moens, U.; Nchinda, G.; Okeke, M.I. Monkeypox Virus in Nigeria: Infection Biology, Epidemiology, and Evolution. Viruses 2020, 12, 1257. [Google Scholar] [CrossRef]
- Silva, N.I.O.; de Oliveira, J.S.; Kroon, E.G.; Trindade, G.S.; Drumond, B.P. Here, There, and Everywhere: The Wide Host Range and Geographic Distribution of Zoonotic Orthopoxviruses. Viruses 2020, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Dhama, K.; Chakraborty, C. Recently spreading human monkeypox virus infection and its transmission during COVID-19 pandemic period: A travelers’ prospective. Travel Med. Infect. Dis. 2022, 49, 102398. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Xie, Y.; Kuai, L.; Wang, H.; Qi, J.; Gao, G.F.; Shi, Y. Structure of monkeypox virus DNA polymerase holoenzyme. Science 2023, 379, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J.; Rathish, B.; Zahra, F. Monkeypox. In StatPearls; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Osadebe, L.; Hughes, C.M.; Shongo Lushima, R.; Kabamba, J.; Nguete, B.; Malekani, J.; Pukuta, E.; Karhemere, S.; Muyembe Tamfum, J.J.; Wemakoy Okitolonda, E.; et al. Enhancing case definitions for surveillance of human monkeypox in the Democratic Republic of Congo. PLoS Negl. Trop. Dis. 2017, 11, e0005857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radonic, A.; Metzger, S.; Dabrowski, P.W.; Couacy-Hymann, E.; Schuenadel, L.; Kurth, A.; Matz-Rensing, K.; Boesch, C.; Leendertz, F.H.; Nitsche, A. Fatal monkeypox in wild-living sooty mangabey, Cote d’Ivoire, 2012. Emerg. Infect. Dis. 2014, 20, 1009–1011. [Google Scholar] [CrossRef]
- Yang, Z. Monkeypox: A potential global threat? J. Med. Virol. 2022, 94, 4034–4036. [Google Scholar] [CrossRef]
- Brown, K.; Leggat, P.A. Human Monkeypox: Current State of Knowledge and Implications for the Future. Trop. Med. Infect. Dis. 2016, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Petersen, B.W.; Kabamba, J.; McCollum, A.M.; Lushima, R.S.; Wemakoy, E.O.; Muyembe Tamfum, J.J.; Nguete, B.; Hughes, C.M.; Monroe, B.P.; Reynolds, M.G. Vaccinating against monkeypox in the Democratic Republic of the Congo. Antivir. Res. 2019, 162, 171–177. [Google Scholar] [CrossRef]
- Adalja, A.; Inglesby, T. A Novel International Monkeypox Outbreak. Ann. Intern. Med. 2022, 175, 1175–1176. [Google Scholar] [CrossRef]
- Yinka-Ogunleye, A.; Aruna, O.; Dalhat, M.; Ogoina, D.; McCollum, A.; Disu, Y.; Mamadu, I.; Akinpelu, A.; Ahmad, A.; Burga, J.; et al. Outbreak of human monkeypox in Nigeria in 2017-18: A clinical and epidemiological report. Lancet Infect. Dis. 2019, 19, 872–879. [Google Scholar] [CrossRef]
- Cabanillas, B.; Valdelvira, R.; Akdis, C.A. Monkeypox outbreak in Europe, UK, North America, and Australia: A changing trend of a zoonotic disease. Allergy 2022, 77, 2284–2286. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Wang, C.; Chuai, X.; Chiu, S. Monkeypox virus: A re-emergent threat to humans. Virol. Sin. 2022, 37, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Eberle, R.; Jones-Engel, L. Questioning the Extreme Neurovirulence of Monkey B Virus (Macacine alphaherpesvirus 1). Adv. Virol. 2018, 2018, 5248420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, L.K.; Black, D.H.; Wright, G.E.; Breshears, M.A.; Eberle, R. Effective Prophylactic Therapy for Exposure to Monkey B Virus (Macacine alphaherpesvirus 1). Comp. Med. 2020, 70, 56–66. [Google Scholar] [CrossRef]
- Johnston, W.F.; Yeh, J.; Nierenberg, R.; Procopio, G. Exposure to Macaque Monkey Bite. J. Emerg. Med. 2015, 49, 634–637. [Google Scholar] [CrossRef]
- Wang, W.; Qi, W.; Liu, J.; Du, H.; Zhao, L.; Zheng, Y.; Wang, G.; Pan, Y.; Huang, B.; Feng, Z.; et al. First Human Infection Case of Monkey B Virus Identified in China, 2021. China CDC Wkly. 2021, 3, 632–633. [Google Scholar] [CrossRef]
- Fan, Q.; Nelson, C.S.; Bialas, K.M.; Chiuppesi, F.; Amos, J.; Gurley, T.C.; Marshall, D.J.; Eudailey, J.; Heimsath, H.; Himes, J.; et al. Plasmablast Response to Primary Rhesus Cytomegalovirus (CMV) Infection in a Monkey Model of Congenital CMV Transmission. Clin. Vaccine Immunol. 2017, 24, e00510-16. [Google Scholar] [CrossRef] [Green Version]
- Deere, J.D.; Chang, W.L.W.; Villalobos, A.; Schmidt, K.A.; Deshpande, A.; Castillo, L.D.; Fike, J.; Walter, M.R.; Barry, P.A.; Hartigan-O’Connor, D.J. Neutralization of rhesus cytomegalovirus IL-10 reduces horizontal transmission and alters long-term immunity. Proc. Natl. Acad. Sci. USA 2019, 116, 13036–13041. [Google Scholar] [CrossRef] [Green Version]
- Abel, K.; Martinez, J.; Yue, Y.; Lacey, S.F.; Wang, Z.; Strelow, L.; Dasgupta, A.; Li, Z.; Schmidt, K.A.; Oxford, K.L.; et al. Vaccine-induced control of viral shedding following rhesus cytomegalovirus challenge in rhesus macaques. J. Virol. 2011, 85, 2878–2890. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Itell, H.L.; Ehlinger, E.P.; Varner, V.; Gantt, S.; Permar, S.R. Natural history of postnatal rhesus cytomegalovirus shedding by dams and acquisition by infant rhesus monkeys. PLoS ONE 2018, 13, e0206330. [Google Scholar] [CrossRef] [Green Version]
- Truitt, L.L.; Yang, D.; Espinoza, D.A.; Fan, X.; Ram, D.R.; Mostrom, M.J.; Tran, D.; Sprehe, L.M.; Reeves, R.K.; Donahue, R.E.; et al. Impact of CMV Infection on Natural Killer Cell Clonal Repertoire in CMV-Naive Rhesus Macaques. Front. Immunol. 2019, 10, 2381. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.G.; Strelow, L.I.; Franchi, D.C.; Anders, D.G.; Wong, S.W. Complete sequence and genomic analysis of rhesus cytomegalovirus. J. Virol. 2003, 77, 6620–6636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malouli, D.; Nakayasu, E.S.; Viswanathan, K.; Camp, D.G., 2nd; Chang, W.L.; Barry, P.A.; Smith, R.D.; Fruh, K. Reevaluation of the coding potential and proteomic analysis of the BAC-derived rhesus cytomegalovirus strain 68-1. J. Virol. 2012, 86, 8959–8973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norley, S.; Kurth, R. The role of the immune response during SIVagm infection of the African green monkey natural host. Front. Biosci. 2004, 9, 550–564. [Google Scholar] [CrossRef] [Green Version]
- Ziani, W.; Shao, J.; Wang, X.; Russell-Lodrigue, K.; Liu, Y.Z.; Montaner, L.J.; Veazey, R.S.; Xu, H. Increased Proviral DNA in Circulating Cells Correlates with Plasma Viral Rebound in Simian Immunodeficiency Virus-Infected Rhesus Macaques after Antiretroviral Therapy Interruption. J. Virol. 2021, 95, e02064-20. [Google Scholar] [CrossRef]
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 1980, 77, 7415–7419. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.; Yasunaga, J.I.; Washizaki, A.; Seki, Y.; Kuramitsu, M.; Tan, W.K.; Hu, A.; Okuma, K.; Hamaguchi, I.; Mizukami, T.; et al. Frequent horizontal and mother-to-child transmission may contribute to high prevalence of STLV-1 infection in Japanese macaques. Retrovirology 2020, 17, 15. [Google Scholar] [CrossRef]
- Castro, I.; Giret, T.M.; Magnani, D.M.; Maxwell, H.S.; Umland, O.; Perry, J.K.; Pecotte, J.K.; Brasky, K.M.; Barber, G.N.; Desrosiers, R.C.; et al. Cellular Immune Responses against Simian T-Lymphotropic Virus Type 1 Target Tax in Infected Baboons. J. Virol. 2016, 90, 5280–5291. [Google Scholar] [CrossRef] [Green Version]
- Termini, J.M.; Magnani, D.M.; Maxwell, H.S.; Lauer, W.; Castro, I.; Pecotte, J.; Barber, G.N.; Watkins, D.I.; Desrosiers, R.C. Simian T Lymphotropic Virus 1 Infection of Papio anubis: Tax Sequence Heterogeneity and T Cell Recognition. J. Virol. 2017, 91, e00950-17. [Google Scholar] [CrossRef] [Green Version]
- Afonso, P.V.; Fagrouch, Z.; Deijs, M.; Niphuis, H.; Bogers, W.; Gessain, A.; van der Hoek, L.; Verschoor, E.J. Absence of accessory genes in a divergent simian T-lymphotropic virus type 1 isolated from a bonnet macaque (Macaca radiata). PLoS Negl. Trop. Dis. 2019, 13, e0007521. [Google Scholar] [CrossRef]
- Alais, S.; Pasquier, A.; Jegado, B.; Journo, C.; Rua, R.; Gessain, A.; Tobaly-Tapiero, J.; Lacoste, R.; Turpin, J.; Mahieux, R. STLV-1 co-infection is correlated with an increased SFV proviral load in the peripheral blood of SFV/STLV-1 naturally infected non-human primates. PLoS Negl. Trop. Dis. 2018, 12, e0006812. [Google Scholar] [CrossRef] [PubMed]
- Liegeois, F.; Boue, V.; Mouacha, F.; Butel, C.; Ondo, B.M.; Pourrut, X.; Leroy, E.; Peeters, M.; Rouet, F. New STLV-3 strains and a divergent SIVmus strain identified in non-human primate bushmeat in Gabon. Retrovirology 2012, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.E.; Torres, J.V. Simian retrovirus receptor and neutralization mechanism by antibodies to the envelope glycoprotein. Viral Immunol. 1999, 12, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Sotir, M.; Switzer, W.; Schable, C.; Schmitt, J.; Vitek, C.; Khabbaz, R.F. Risk of occupational exposure to potentially infectious nonhuman primate materials and to simian immunodeficiency virus. J. Med. Primatol. 1997, 26, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Apakupakul, K.; Deem, S.L.; Maqsood, R.; Sithiyopasakul, P.; Wang, D.; Lim, E.S. Endogenization of a Prosimian Retrovirus during Lemur Evolution. Viruses 2021, 13, 383. [Google Scholar] [CrossRef]
- Grant, R.; Keele, B.; Kuller, L.; Watanabe, R.; Perret, A.; Smedley, J. Identification of novel simian endogenous retroviruses that are indistinguishable from simian retrovirus (SRV) on current SRV diagnostic assays. J. Med. Primatol. 2017, 46, 158–161. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, L.; Zhang, Q.; Meng, J.; Lu, Z.L.; Rong, R. Autophagy Induced by Simian Retrovirus Infection Controls Viral Replication and Apoptosis of Jurkat T Lymphocytes. Viruses 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Togami, H.; Shimura, K.; Okamoto, M.; Yoshikawa, R.; Miyazawa, T.; Matsuoka, M. Comprehensive in vitro analysis of simian retrovirus type 4 susceptibility to antiretroviral agents. J. Virol. 2013, 87, 4322–4329. [Google Scholar] [CrossRef] [Green Version]
- Lerche, N.W.; Heneine, W.; Kaplan, J.E.; Spira, T.; Yee, J.L.; Khabbaz, R.F. An expanded search for human infection with simian type D retrovirus. AIDS Res. Hum. Retrovir. 1995, 11, 527–529. [Google Scholar] [CrossRef] [Green Version]
- Sandstrom, P.A.; Phan, K.O.; Switzer, W.M.; Fredeking, T.; Chapman, L.; Heneine, W.; Folks, T.M. Simian foamy virus infection among zoo keepers. Lancet 2000, 355, 551–552. [Google Scholar] [CrossRef]
- Lambert, C.; Couteaudier, M.; Gouzil, J.; Richard, L.; Montange, T.; Betsem, E.; Rua, R.; Tobaly-Tapiero, J.; Lindemann, D.; Njouom, R.; et al. Potent neutralizing antibodies in humans infected with zoonotic simian foamy viruses target conserved epitopes located in the dimorphic domain of the surface envelope protein. PLoS Pathog. 2018, 14, e1007293. [Google Scholar] [CrossRef] [PubMed]
- Switzer, W.M.; Bhullar, V.; Shanmugam, V.; Cong, M.E.; Parekh, B.; Lerche, N.W.; Yee, J.L.; Ely, J.J.; Boneva, R.; Chapman, L.E.; et al. Frequent simian foamy virus infection in persons occupationally exposed to nonhuman primates. J. Virol. 2004, 78, 2780–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Switzer, W.M.; Salemi, M.; Shanmugam, V.; Gao, F.; Cong, M.E.; Kuiken, C.; Bhullar, V.; Beer, B.E.; Vallet, D.; Gautier-Hion, A.; et al. Ancient co-speciation of simian foamy viruses and primates. Nature 2005, 434, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcone, V.; Schweizer, M.; Neumann-Haefelin, D. Replication of primate foamy viruses in natural and experimental hosts. Curr. Top. Microbiol. Immunol. 2003, 277, 161–180. [Google Scholar] [CrossRef]
- Rua, R.; Betsem, E.; Montange, T.; Buseyne, F.; Gessain, A. In vivo cellular tropism of gorilla simian foamy virus in blood of infected humans. J. Virol. 2014, 88, 13429–13435. [Google Scholar] [CrossRef] [Green Version]
- Weingartl, H.M.; Embury-Hyatt, C.; Nfon, C.; Leung, A.; Smith, G.; Kobinger, G. Transmission of Ebola virus from pigs to non-human primates. Sci. Rep. 2012, 2, 811. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Blancett, C.D.; Koistinen, K.A.; Schellhase, C.W.; Bearss, J.J.; Radoshitzky, S.R.; Honnold, S.P.; Chance, T.B.; Warren, T.K.; Froude, J.W.; et al. Identification and pathological characterization of persistent asymptomatic Ebola virus infection in rhesus monkeys. Nat. Microbiol. 2017, 2, 17113. [Google Scholar] [CrossRef]
- Warren, T.K.; Kane, C.D.; Wells, J.; Stuthman, K.S.; Van Tongeren, S.A.; Garza, N.L.; Donnelly, G.; Steffens, J.; Gomba, L.; Weidner, J.M.; et al. Remdesivir is efficacious in rhesus monkeys exposed to aerosolized Ebola virus. Sci. Rep. 2021, 11, 19458. [Google Scholar] [CrossRef]
- Brinton, M.A.; Di, H.; Vatter, H.A. Simian hemorrhagic fever virus: Recent advances. Virus Res. 2015, 202, 112–119. [Google Scholar] [CrossRef]
- Lapin, B.A.; Shevtsova, Z.V. [To the 50th anniversary of the discovery of the simian hemorrhagic fever and SHF virus]. Vopr. Virusol. 2015, 60, 5–11. [Google Scholar]
- Vatter, H.A.; Donaldson, E.F.; Huynh, J.; Rawlings, S.; Manoharan, M.; Legasse, A.; Planer, S.; Dickerson, M.F.; Lewis, A.D.; Colgin, L.M.; et al. A simian hemorrhagic fever virus isolate from persistently infected baboons efficiently induces hemorrhagic fever disease in Japanese macaques. Virology 2015, 474, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Lauck, M.; Sibley, S.D.; Hyeroba, D.; Tumukunde, A.; Weny, G.; Chapman, C.A.; Ting, N.; Switzer, W.M.; Kuhn, J.H.; Friedrich, T.C.; et al. Exceptional simian hemorrhagic fever virus diversity in a wild African primate community. J. Virol. 2013, 87, 688–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl-Jensen, V.; Johnson, J.C.; Lauck, M.; Weinfurter, J.T.; Moncla, L.H.; Weiler, A.M.; Charlier, O.; Rojas, O.; Byrum, R.; Ragland, D.R.; et al. Divergent Simian Arteriviruses Cause Simian Hemorrhagic Fever of Differing Severities in Macaques. Mbio 2016, 7, e02009-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, G.M.; Harrach, B.; Zakhartchouk, A.N.; Davison, A.J. Complete genome sequence of simian adenovirus 1: An Old World monkey adenovirus with two fiber genes. J. Gen. Virol. 2005, 86, 1681–1686. [Google Scholar] [CrossRef] [PubMed]
- Kobler, H.; Regenfuss, P. [A simple technique for orotracheal intubation in the rabbit]. Z. Gesamte Exp. Med. Einschl. Exp. Chir. 1971, 154, 325–327. [Google Scholar]
- Morgan, T.J.; Glowaski, M.M. Teaching a new method of rabbit intubation. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 32–36. [Google Scholar]
- Han, J.W.; La, T.M.; Kim, J.H.; Choi, I.S.; Song, C.S.; Park, S.Y.; Lee, J.B.; Lee, S.W. The possible origin of human adenovirus type 3: Evidence of natural genetic recombination between human and simian adenovirus. Infect. Genet. Evol. 2018, 65, 380–384. [Google Scholar] [CrossRef]
- Wevers, D.; Metzger, S.; Babweteera, F.; Bieberbach, M.; Boesch, C.; Cameron, K.; Couacy-Hymann, E.; Cranfield, M.; Gray, M.; Harris, L.A.; et al. Novel adenoviruses in wild primates: A high level of genetic diversity and evidence of zoonotic transmissions. J. Virol. 2011, 85, 10774–10784. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.J.; Sebastian, S.; Spencer, A.J.; Gilbert, S.C. Simian adenoviruses as vaccine vectors. Future Virol. 2016, 11, 649–659. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, M.G.; Anderson, D.C.; Fikes, J.D.; Bain, F.T.; Carlson, C.S.; Green, S.W.; Young, N.S.; Brown, K.E. Identification of a novel simian parvovirus in cynomolgus monkeys with severe anemia. A paradigm of human B19 parvovirus infection. J. Clin. Investig. 1994, 93, 1571–1576. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogens | Symptoms | Diagnostic Methods | Vaccination | Antiviral Drugs | Reference |
|---|---|---|---|---|---|
| Monkeypox virus | Milder rash and lower mortality | RT-PCR, NGS, electron microscope, Immunological | No effective measures | No effective curative treatment | [9,10] |
| Monkey B virus | Oral herpes-like lesions, such as oral ulcers, conjunctivitis | RT-PCR, NGS, Immunological, Histopathological | No effective measures | Effective suppressive treatment but not curative | [11] |
| Ebola virus; Marburg virus | Severe hemorrhagic fever fatal cases | Epidemiological data, clinical manifestations, positive laboratory tests for viral antigen and positive serum | No effective measures | mAbs therapy, Inmazeb, Ebanga | [12] |
| Simian cytomegalovirus | Basically recessive and does not show any clinical symptoms and signs | virus isolation and culture, serum antibody testing, antigen testing, and viral nucleic acid molecular biology testing | No effective measures | No effective curative treatment | [13] |
| Simian immunodeficiency virus | Monkey-acquired immunodeficiency (SAIDS) | Laboratory virus isolation, serological tests, histopathological diagnosis, and molecular biological to diagnosis | No effective measures | No effective curative treatment | [14] |
| Simian type D retrovirus | SAIDS | Serology and viral molecular biology | Inactivated SRV-Ⅰ, outer membrane glycoprotein vaccines expressing simian retrovirus | AZT and TDF, potently inhibited SRV-4 infection | [15,16,17] |
| Simian hemorrhagic fever virus | Hemorrhage and fever | Clinical symptoms, pathological, epidemic characteristics, serological tests, molecular biological diagnosis | No effective measures | No effective curative treatment | [18] |
| Simian adenovirus | Pneumonia, pharyngitis, gastroenteritis, conjunctivitis | Pathogen isolation, PCR, transmission electron microscopy, immunofluorescence detection, nucleic acid hybridization | No effective measures | No effective treatment | [19] |
| Simian parvovirus | Not apparent or causes only mild febrile illness | Clinical symptoms, epidemic characteristics, serological tests, hemagglutination and hemagglutination inhibition, histopathological and molecular biological | No effective measures | No effective curative treatment | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Fan, Z.; Li, S.; Yin, H. A Review on Zoonotic Pathogens Associated with Non-Human Primates: Understanding the Potential Threats to Humans. Microorganisms 2023, 11, 246. https://doi.org/10.3390/microorganisms11020246
Jiang X, Fan Z, Li S, Yin H. A Review on Zoonotic Pathogens Associated with Non-Human Primates: Understanding the Potential Threats to Humans. Microorganisms. 2023; 11(2):246. https://doi.org/10.3390/microorganisms11020246
Chicago/Turabian StyleJiang, Xinjie, Zhenyu Fan, Shijia Li, and Haichang Yin. 2023. "A Review on Zoonotic Pathogens Associated with Non-Human Primates: Understanding the Potential Threats to Humans" Microorganisms 11, no. 2: 246. https://doi.org/10.3390/microorganisms11020246
APA StyleJiang, X., Fan, Z., Li, S., & Yin, H. (2023). A Review on Zoonotic Pathogens Associated with Non-Human Primates: Understanding the Potential Threats to Humans. Microorganisms, 11(2), 246. https://doi.org/10.3390/microorganisms11020246