
Validation of Multiplex PCR and Serology Detecting Helicobacter Species in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin, Housing and Sampling of Mice
2.2. Helicobacter Infection Experiment, Housing and Handling of Mice
2.3. DNA Preparation
2.4. Conventional PCR
2.5. Helicobacter Multiplex DNA Finder
2.6. Helicobacter Multiplex DNA Finder Controls
2.7. Cut-Off Definition of Helicobacter Multiplex DNA Finder
2.8. Helicobacter Multiplex Serology
2.9. Statistical Analysis
3. Results
3.1. Analytical Sensitivity of Helicobacter Multiplex DNA Finder
3.2. Specificity of Helicobacter Multiplex DNA Finder
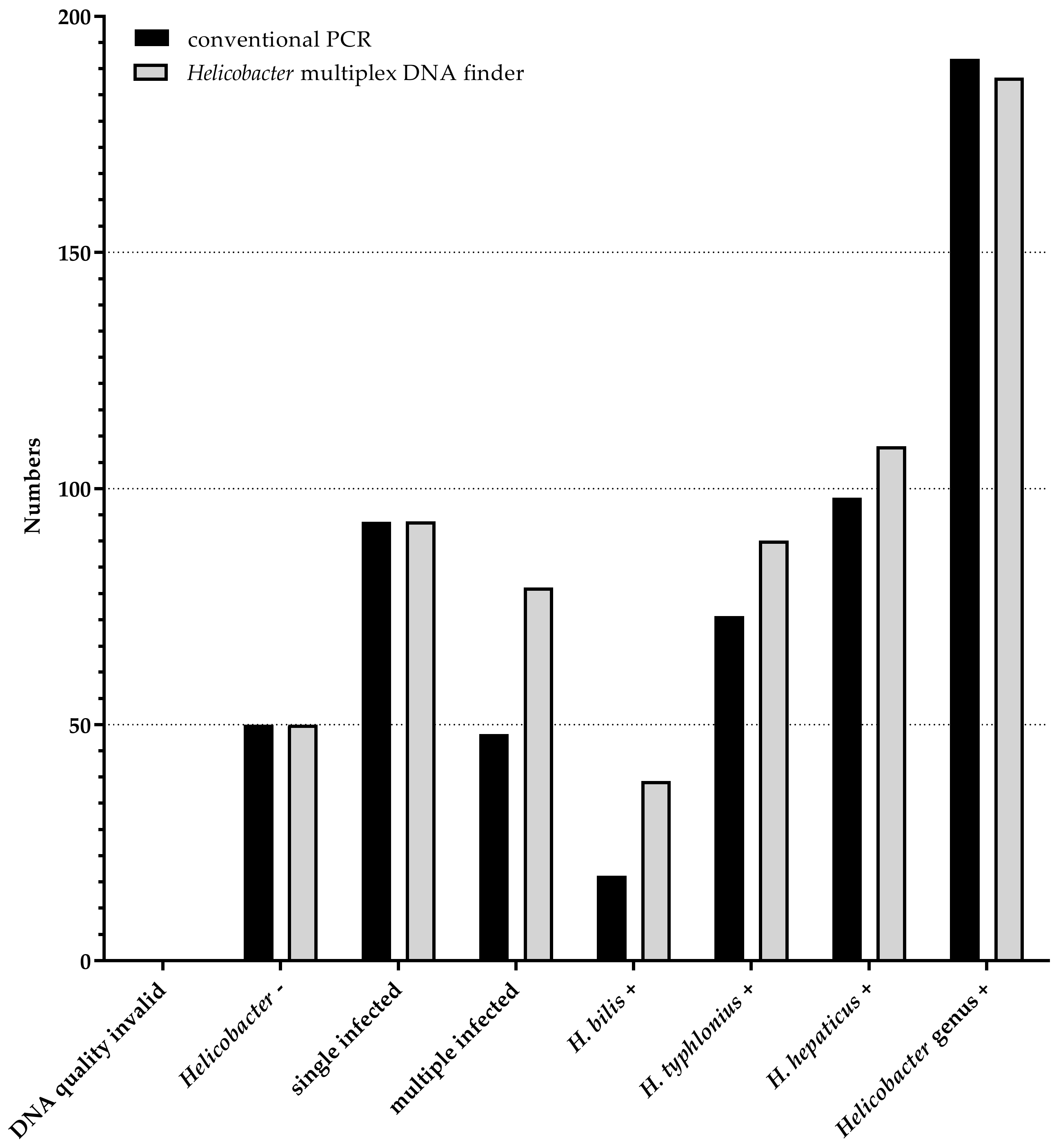
3.3. Comparison of Helicobacter Multiplex DNA Finder to Conventional PCR
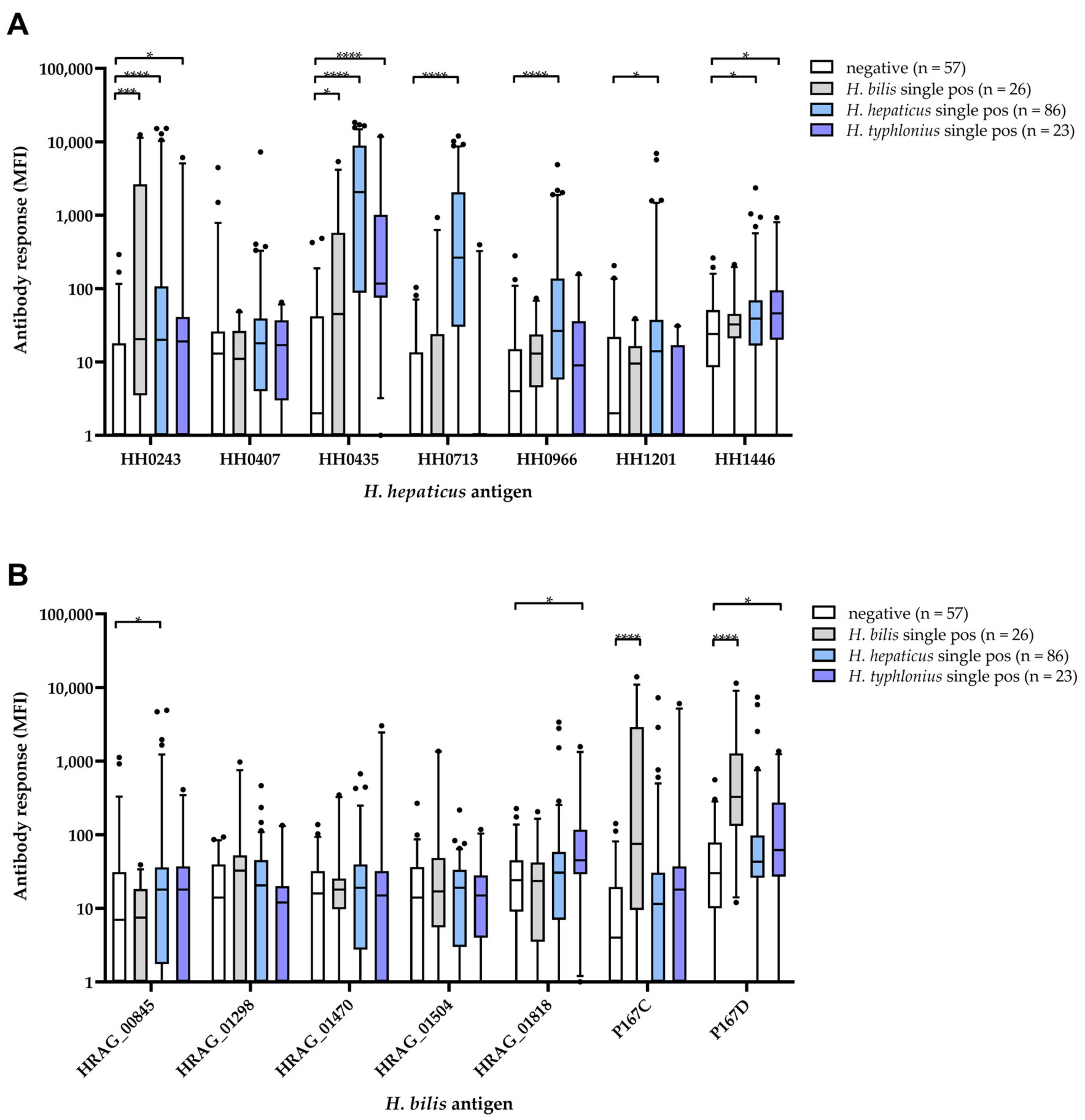
3.4. Comparison of H. hepaticus and H. bilis Multiplex Serology to Helicobacter Multiplex DNA Finder Results
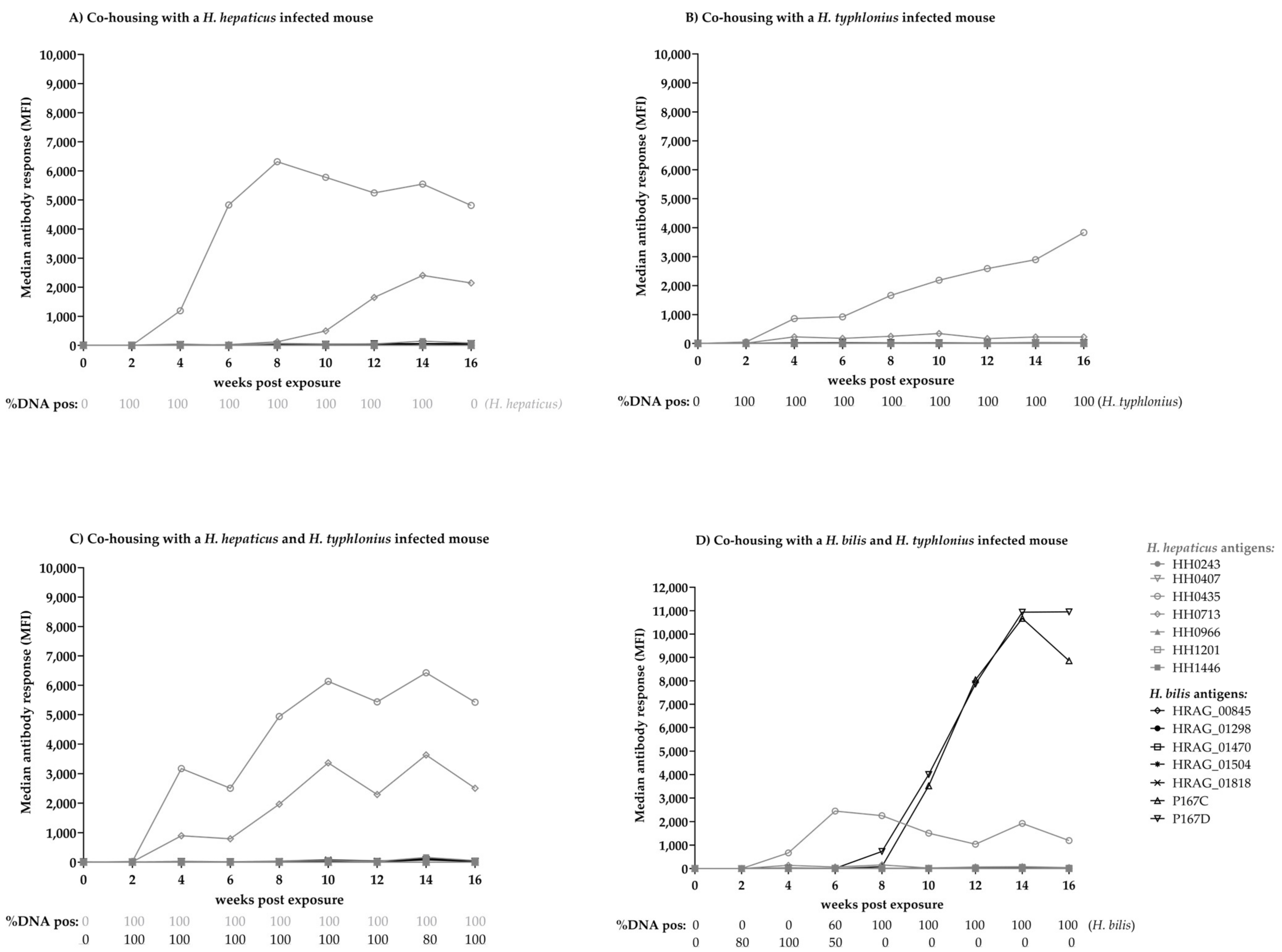
3.5. Experimental Infection of Mice with Helicobacter spp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mannion, A.; Shen, Z.; Fox, J.G. Comparative genomics analysis to differentiate metabolic and virulence gene potential in gastric versus enterohepatic Helicobacter species. BMC Genom. 2018, 19, 830. [Google Scholar] [CrossRef] [PubMed]
- Shames, B.; Fox, J.G.; Dewhirst, F.; Yan, L.; Shen, Z.; Taylor, N.S. Identification of widespread Helicobacter hepaticus infection in feces in commercial mouse colonies by culture and PCR assay. J. Clin. Microbiol. 1995, 33, 2968–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, J.G.; Dewhirst, F.E.; Tully, J.G.; Paster, B.J.; Yan, L.; Taylor, N.S.; Collins, M.J.; Gorelick, P.L.; Ward, J.M. Helicobacter hepaticus Sp-Nov, a Microaerophilic Bacterium Isolated from Livers and Intestinal Mucosal Scrapings from Mice. J. Clin. Microbiol. 1994, 32, 1238–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ramer-Tait, A.E.; Henderson, A.L.; Demirkale, C.Y.; Nettleton, D.; Wang, C.; Hostetter, J.M.; Jergens, A.E.; Wannemuehler, M.J. Helicobacter bilis colonization enhances susceptibility to Typhlocolitis following an inflammatory trigger. Dig. Dis. Sci. 2011, 56, 2838–2848. [Google Scholar] [CrossRef]
- Ge, Z.; Sterzenbach, T.; Whary, M.T.; Rickman, B.H.; Rogers, A.B.; Shen, Z.; Taylor, N.S.; Schauer, D.B.; Josenhans, C.; Suerbaum, S.; et al. Helicobacter hepaticus HHGI1 is a pathogenicity island associated with typhlocolitis in B6.129-IL10 tm1Cgn mice. Microbes Infect. Inst. Pasteur 2008, 10, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Lee, A.; Whary, M.T.; Rogers, A.B.; Maurer, K.J.; Taylor, N.S.; Schauer, D.B.; Fox, J.G. Helicobacter hepaticus urease is not required for intestinal colonization but promotes hepatic inflammation in male A/JCr mice. Microb. Pathog. 2008, 45, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Whary, M.T.; Fox, J.G. Natural and experimental Helicobacter infections. Comp. Med. 2004, 54, 128–158. [Google Scholar]
- Fox, J.G.; Rogers, A.B.; Whary, M.T.; Taylor, N.S.; Xu, S.; Feng, Y.; Keys, S. Helicobacter bilis-associated hepatitis in outbred mice. Comp. Med. 2004, 54, 571–577. [Google Scholar]
- Li, X.; Fox, J.G.; Whary, M.T.; Yan, L.; Shames, B.; Zhao, Z. SCID/NCr mice naturally infected with Helicobacter hepaticus develop progressive hepatitis, proliferative typhlitis, and colitis. Infect. Immun. 1998, 66, 5477–5484. [Google Scholar] [CrossRef] [Green Version]
- Franklin, C.L.; Riley, L.K.; Livingston, R.S.; Beckwith, C.S.; Besch-Williford, C.L.; Hook, R.R., Jr. Enterohepatic lesions in SCID mice infected with Helicobacter bilis. Lab. Anim. Sci. 1998, 48, 334–339. [Google Scholar]
- Ward, J.M.; Anver, M.R.; Haines, D.C.; Melhorn, J.M.; Gorelick, P.; Yan, L.; Fox, J.G. Inflammatory large bowel disease in immunodeficient mice naturally infected with Helicobacter hepaticus. Lab. Anim. Sci. 1996, 46, 15–20. [Google Scholar]
- Fox, J.G.; Yan, L.; Shames, B.; Campbell, J.; Murphy, J.C.; Li, X. Persistent hepatitis and enterocolitis in germfree mice infected with Helicobacter hepaticus. Infect. Immun. 1996, 64, 3673–3681. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.G.; Li, X.; Yan, L.; Cahill, R.J.; Hurley, R.; Lewis, R.; Murphy, J.C. Chronic proliferative hepatitis in A/JCr mice associated with persistent Helicobacter hepaticus infection: A model of helicobacter-induced carcinogenesis. Infect. Immun. 1996, 64, 1548–1558. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.M.; Fox, J.G.; Anver, M.R.; Haines, D.C.; George, C.V.; Collins, M.J., Jr.; Gorelick, P.L.; Nagashima, K.; Gonda, M.A.; Gilden, R.V.; et al. Chronic active hepatitis and associated liver tumors in mice caused by a persistent bacterial infection with a novel Helicobacter species. J. Natl. Cancer Inst. 1994, 86, 1222–1227. [Google Scholar] [CrossRef]
- FELASAWorking Group on Revision of Guidelines for Health Monitoring of Rodents Rabbits; Mähler, M.; Berard, M.; Feinstein, R.; Gallagher, A.; Illgen-Wilcke, B.; Pritchett-Corning, K.; Raspa, M. FELASA recommendations for the health monitoring of mouse, rat, hamster, guinea pig and rabbit colonies in breeding and experimental units. Lab. Anim. 2014, 48, 178–192. [Google Scholar]
- Poynter, S.; Phipps, J.D.; Naranjo-Pino, A.; Sanchez-Morgado, J.M. Difficulties in the molecular diagnosis of Helicobacter rodent infections. Vet. Microbiol. 2009, 134, 272–278. [Google Scholar] [CrossRef] [Green Version]
- Riley, L.K.; Franklin, C.L.; Hook, R.R., Jr.; Besch-Williford, C. Identification of murine helicobacters by PCR and restriction enzyme analyses. J. Clin. Microbiol. 1996, 34, 942–946. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.G.; Yan, L.L.; Dewhirst, F.E.; Paster, B.J.; Shames, B.; Murphy, J.C.; Hayward, A.; Belcher, J.C.; Mendes, E.N. Helicobacter bilis sp. nov., a novel Helicobacter species isolated from bile, livers, and intestines of aged, inbred mice. J. Clin. Microbiol. 1995, 33, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Franklin, C.L.; Riley, L.K.; Livingston, R.S.; Beckwith, C.S.; Hook, R.R., Jr.; Besch-Williford, C.L.; Hunziker, R.; Gorelick, P.L. Enteric lesions in SCID mice infected with “Helicobacter typhlonicus”, a novel urease-negative Helicobacter species. Lab. Anim. Sci. 1999, 49, 496–505. [Google Scholar]
- Boutin, S.R.; Shen, Z.; Roesch, P.L.; Stiefel, S.M.; Sanderson, A.E.; Multari, H.M.; Pridhoko, E.A.; Smith, J.C.; Taylor, N.S.; Lohmiller, J.J.; et al. Helicobacter pullorum outbreak in C57BL/6NTac and C3H/HeNTac barrier-maintained mice. J. Clin. Microbiol. 2010, 48, 1908–1910. [Google Scholar] [CrossRef] [Green Version]
- Pisani, P.; Whary, M.T.; Nilsson, I.; Sriamporn, S.; Wadstrom, T.; Fox, J.G.; Ljungh, A.; Forman, D. Cross-reactivity between immune responses to Helicobacter bilis and Helicobacter pylori in a population in Thailand at high risk of developing cholangiocarcinoma. Clin. Vaccine Immunol. 2008, 15, 1363–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananieva, O.; Nilsson, I.; Vorobjova, T.; Uibo, R.; Wadstrom, T. Immune Responses to Bile-Tolerant Helicobacter Species in Patients with Chronic Liver Diseases, a Randomized Population Group, and Healthy Blood Donors. Clin. Vaccine Immunol. 2002, 9, 1160–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, I.; Lindgren, S.; Eriksson, S.; Wadstrom, T. Serum antibodies to Helicobacter hepaticus and Helicobacter pylori in patients with chronic liver disease. Gut 2000, 46, 410–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Kendall, L.V.; Hodzic, E.; Wong, S.; Lorenzana, E.; Freet, K.; Ku, K.S.; Luciw, P.A.; Barthold, S.W.; Khan, I.H. Recombinant Helicobacter bilis protein P167 for mouse serodiagnosis in a multiplex microbead assay. Clin. Diagn. Lab. Immunol. 2004, 11, 1094–1099. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Hodzic, E.; Kendall, L.V.; Smith, A.; Freet, K.; Barthold, S.W. Cloning and expression of a Helicobacter bilis immunoreactive protein. Clin. Diagn. Lab. Immunol. 2002, 9, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Livingston, R.S.; Riley, L.K.; Hook, R.R., Jr.; Besch-Williford, C.L.; Franklin, C.L. Cloning and expression of an immunogenic membrane-associated protein of Helicobacter hepaticus for use in an enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol. 1999, 6, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Michel, A.; Waterboer, T.; Kist, M.; Pawlita, M. Helicobacter pylori multiplex serology. Helicobacter 2009, 14, 525–535. [Google Scholar] [CrossRef]
- Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:de:PDF (accessed on 1 December 2022).
- Hofler, D.; Nicklas, W.; Mauter, P.; Pawlita, M.; Schmitt, M. A bead-based multiplex assay for the detection of DNA viruses infecting laboratory rodents. PLoS ONE 2014, 9, e97525. [Google Scholar] [CrossRef]
- Schmitt, M.; Bravo, I.G.; Snijders, P.J.; Gissmann, L.; Pawlita, M.; Waterboer, T. Bead-based multiplex genotyping of human papillomaviruses. J. Clin. Microbiol. 2006, 44, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Chien, C.C.; Taylor, N.S.; Ge, Z.; Schauer, D.B.; Young, V.B.; Fox, J.G. Identification of cdtB homologues and cytolethal distending toxin activity in enterohepatic Helicobacter spp. J. Med. Microbiol. 2000, 49, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Suerbaum, S.; Josenhans, C.; Sterzenbach, T.; Drescher, B.; Brandt, P.; Bell, M.; Droge, M.; Fartmann, B.; Fischer, H.P.; Ge, Z.; et al. The complete genome sequence of the carcinogenic bacterium Helicobacter hepaticus. Proc. Natl. Acad. Sci. USA 2003, 100, 7901–7906. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 3 December 2020).
- Waterboer, T.; Sehr, P.; Michael, K.M.; Franceschi, S.; Nieland, J.D.; Joos, T.O.; Templin, M.F.; Pawlita, M. Multiplex human papillomavirus serology based on in situ-purified glutathione s-transferase fusion proteins. Clin. Chem. 2005, 51, 1845–1853. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.G.; Gorelick, P.L.; Kullberg, M.C.; Ge, Z.; Dewhirst, F.E.; Ward, J.M. A novel urease-negative Helicobacter species associated with colitis and typhlitis in IL-10-deficient mice. Infect. Immun. 1999, 67, 1757–1762. [Google Scholar] [CrossRef]
- Donovan, J.C.; Mayo, J.G.; Rice, J.M.; Ward, J.M.; Fox, J.G. Helicobacter-associated hepatitis of mice. Lab. Anim. Sci. 1993, 43, 403. [Google Scholar]
- Whary, M.T.; Fox, J.G. Detection, eradication, and research implications of Helicobacter infections in laboratory rodents. Lab. Anim. 2006, 35, 25–36. [Google Scholar] [CrossRef]
- Neubert, V.; Sadek, A.; Burell, T.; Ralser, A.; Erhard, M.; Gerhard, M.; Seidel, K.; Kalali, B. Validation and improvement of a multiplex PCR method to detect murine Helicobacter species in feces samples of mice. Helicobacter 2022, 27, e12888. [Google Scholar] [CrossRef]
- Ge, Z.; White, D.A.; Whary, M.T.; Fox, J.G. Fluorogenic PCR-based quantitative detection of a murine pathogen, Helicobacter hepaticus. J. Clin. Microbiol. 2001, 39, 2598–2602. [Google Scholar] [CrossRef] [Green Version]
- Drazenovich, N.L.; Franklin, C.L.; Livingston, R.S.; Besselsen, D.G. Detection of rodent Helicobacter spp. by use of fluorogenic nuclease polymerase chain reaction assays. Comp. Med. 2002, 52, 347–353. [Google Scholar]
- Dewhirst, F.E.; Shen, Z.; Scimeca, M.S.; Stokes, L.N.; Boumenna, T.; Chen, T.; Paster, B.J.; Fox, J.G. Discordant 16S and 23S rRNA gene phylogenies for the genus Helicobacter: Implications for phylogenetic inference and systematics. J. Bacteriol. 2005, 187, 6106–6118. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, M.; Pawlita, M. High-throughput detection and multiplex identification of cell contaminations. Nucleic Acids Res. 2009, 37, e119. [Google Scholar] [CrossRef]
- Kendall, L.V.; Feng, S.; Hodzic, E.; Freet, K.; Barthold, S.W. Use of the P167 recombinant antigen for serodiagnosis of Helicobacter bilis. Comp. Med. 2004, 54, 44–48. [Google Scholar] [PubMed]
- Schmidt, K.; Butt, J.; Mauter, P.; Vogel, K.; Erles-Kemna, A.; Pawlita, M.; Nicklas, W. Development of a multiplex serological assay reveals a worldwide distribution of murine astrovirus infections in laboratory mice. PLoS ONE 2017, 12, e0187174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whary, M.T.; Cline, J.H.; King, A.E.; Hewes, K.M.; Chojnacky, D.; Salvarrey, A.; Fox, J.G. Monitoring sentinel mice for Helicobacter hepaticus, H. rodentium, and H. bilis infection by use of polymerase chain reaction analysis and serologic testing. Comp. Med. 2000, 50, 436–443. [Google Scholar] [PubMed]
- IARC; World Health Organization. Schistosomes, Liver Flukes and Helicobacter pylori. In Monographs on the Evaluation of Carcinogenic Risks to Humans; Iarc Working Group on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1994; Volume 61, pp. 1–241. [Google Scholar]
- Kosaka, T.; Tajima, Y.; Kuroki, T.; Mishima, T.; Adachi, T.; Tsuneoka, N.; Fukuda, K.; Kanematsu, T. Helicobacter bilis colonization of the biliary system in patients with pancreaticobiliary maljunction. Br. J. Surg. 2010, 97, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Yokota, K.; Ayada, K.; Hirai, K.; Kamada, T.; Haruma, K.; Chayama, K.; Oguma, K. Detection of Helicobacter hepaticus in human bile samples of patients with biliary disease. Helicobacter 2009, 14, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Rocha, M.; Avenaud, P.; Menard, A.; Le Bail, B.; Balabaud, C.; Bioulac-Sage, P.; de Magalhaes Queiroz, D.M.; Megraud, F. Association of Helicobacter species with hepatitis C cirrhosis with or without hepatocellular carcinoma. Gut 2005, 54, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Pellicano, R.; Mazzaferro, V.; Grigioni, W.F.; Cutufia, M.A.; Fagoonee, S.; Silengo, L.; Rizzetto, M.; Ponzetto, A. Helicobacter species sequences in liver samples from patients with and without hepatocellular carcinoma. World J. Gastroenterol. 2004, 10, 598–601. [Google Scholar] [CrossRef]
- Fox, J.G.; Dewhirst, F.E.; Shen, Z.; Feng, Y.; Taylor, N.S.; Paster, B.J.; Ericson, R.L.; Lau, C.N.; Correa, P.; Araya, J.C.; et al. Hepatic Helicobacter species identified in bile and gallbladder tissue from Chileans with chronic cholecystitis. Gastroenterology 1998, 114, 755–763. [Google Scholar] [CrossRef]
- Segura-Lopez, F.K.; Aviles-Jimenez, F.; Guitron-Cantu, A.; Valdez-Salazar, H.A.; Leon-Carballo, S.; Guerrero-Perez, L.; Fox, J.G.; Torres, J. Infection with Helicobacter bilis but not Helicobacter hepaticus was Associated with Extrahepatic Cholangiocarcinoma. Helicobacter 2015, 20, 223–230. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, J.D.; Weng, M.Z.; Zhang, Y.; Wang, X.F.; Gong, W.; Quan, Z.W. Infections of Helicobacter spp. in the biliary system are associated with biliary tract cancer: A meta-analysis. Eur. J. Gastroenterol. Hepatol. 2013, 25, 447–454. [Google Scholar] [CrossRef]
- Alvarez, C.S.; Florio, A.A.; Butt, J.; Rivera-Andrade, A.; Kroker-Lobos, M.F.; Waterboer, T.; Camargo, M.C.; Freedman, N.D.; Graubard, B.I.; Lazo, M.; et al. Associations between Helicobacter pylori with nonalcoholic fatty liver disease and other metabolic conditions in Guatemala. Helicobacter 2020, 25, e12756. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PCR Target | Gene Target | Primer/Probe Name | Sequence (5′-3′) | Fragment Size | Reference |
|---|---|---|---|---|---|
| Helicobacter genus | 16S rRNA | h276 forward | CTATGACGGGTATCCGGC | 400 bp | Riley et al. [17] |
| h676 reverse | ATTCCACCTACCTCTCCCA | ||||
| H. hepaticus | 16S rRNA | B38 forward | GCATTTGAAACTGTTACTCTG | 417 bp | Shames et al. [2] |
| B39 reverse | CTGTTTTCAAGCTCCCC | ||||
| H. typhlonius | 16S rRNA | Ht 184 forward | TTAAAGATATTCTAGGGGTATAT | 474 bp | Franklin et al. [19] |
| Ht 640 reverse | TCTCCCATACTCTAGAGTGA | ||||
| H. bilis | 16S rRNA | C62 forward | AGAACTGCATTTGAAACTACTTT | 638 bp | Fox et al. [18] |
| C12 reverse | GGTATTGCATCTCTTTGTATGT |
| PCR Target | Gene Target | Primer/Probe Name | Sequence (5′-3′) | Fragment Size |
|---|---|---|---|---|
| H. muridarum | gyAa | fw2_murid | CCAAATGCCAGAGATGGAT | 122 bp |
| bw1_murid | CCGATTACATCACCAACTAT | |||
| p_murid | TATGAATGAGCTAAACCTTACACA | |||
| ggtb | fw2_mur | GCCACTAGAGATATGTATCTTG | 112 bp | |
| bw2_mur | GCATAGCACTCATTCCTT | |||
| p1_mur | AGATGTCCAATAATAGAA | |||
| H. pylori | gyA | fw1_pylori | GATCGCYGTRGGGATGGC | 144 bp |
| bw1_pylori | AGTGGGAAARTCAGGCCCTT | |||
| p_pylori | CTTTAGYGCATGTCTTAGAA | |||
| ggt | fw1_pylori | TTAGACAAGCAAGGCAATGT | 94 bp | |
| bw2_pylori | ACATCGCTTCCATGCCCGC | |||
| p1_pylori | AAATAGCCATCTTCRCTG | |||
| H. bilis | gyA | fw1_bilis | TGCCTAATGCAAGAGATGG | 126 bp |
| bw1_bilis | CCATTACTTCCCCCACAAT | |||
| p_bilis | CGATGAAGCATAATCTAGGG | |||
| ggt | fw2_bilis | GGATAATAAAGGTAATGTGATTCC | 138 bp | |
| bw2_bilis | GCAGGTTGCATGAGTTC | |||
| p3_bilis | GCTAAATATCCAAGTGTTGAAGCA | |||
| H. heilmannii | gyA | Fw2_Hheilm | CTTGCAAATAGGCGATCT | 129 bp |
| bw1_Hheilm | CGCATGATCTAAGTGAAG | |||
| p2_Hheilm | TTCTCCTGCTCTAGCCC | |||
| H. typhlonius | gyA | fw2_typhlonius | ATTGTAGGTAGGGCGTTA | 149 bp |
| bw2_typhlonius | TGGTATTTACCAATCACATC | |||
| p_typhlonius | GATGAACGAGCTAAGCCTTTCACC | |||
| H. cinaedi | gyA | fw1_cinaedi | TACCAGATGCTAAAGATGG | 122 bp |
| bw1_cinaedi | AATCACATCGCCAACAAT | |||
| p_cinaedi | AATGAATGAGCTAAGCCTCTCT | |||
| H. hepaticus | gyA | fw1_hepaticus | CCTGACGCAAAAGATGG | 122 bp |
| bw1_hepaticus | ATTTACCAATTACATCGCCTAC | |||
| p_hepaticus | AATGAATGAGCTTAATCTCTCACC | |||
| H. pullorum | gyA | fw1_Hpull | AATGGAATAAGAGAGGCTTA | 128 bp |
| bw_Hpull | GCTTTATTGACCTGATAGGGA | |||
| p1_Hpull | TTAATGCGCCCTCTCCCTG | |||
| C. jejuni | gyA | fw2_Cjejuni | ATGAAACTTGGTCGTTTAACA | 178 bp |
| bw2_Cjejuni | GAGTAATACGTGGCACA | |||
| p_Cjejuni | CTTGCTTGAAAATTTAATTCG | |||
| ggt | fw2_Cjejuni | TGTATCTTGATAGCAAAGGAGAA | 105 bp | |
| bw1_ Cjejuni | GATCAAGCATAGCACTCATACC | |||
| p1_Cjeju | CAGCTAGATAACCTATAGT | |||
| Helicobacter genus | 16S rRNA | fw2.3_heli_uni | GAGTATGGGAGAGGTAGGTGGAATTC | 110 bp |
| bw2.2_heli_uni | TAATCCTGTTTGCTCCCCACGC | |||
| p2.2_heli_uni | CAATGAGTATTCCTCTTGA |
| Species | Gene | Analytical Sensitivity [# of Copies/PCR] a |
|---|---|---|
| H. muridarum | gyA | 10 |
| ggt | 100 | |
| H. pylori | gA | 10 |
| ggt | 10 | |
| H. bilis | gyA | 100 |
| ggt | 10 | |
| H. heilmannii | gyA | 1000 |
| H. typhlonius | gyA | 1000 |
| H. cinaedi | gyA | 10 |
| H. hepaticus | gyA | 10 |
| H. pullorum | gyA | 10 |
| C. jejuni | gyA | 10,000 |
| ggt | 10 | |
| Helicobacter genus | 16SrRNA | 10 |
| Mus musculus | polA | 10 |
| Homo sapiens | polA | 10 |
| Bacteria-Specific Probe | QC Probe | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H. muridarum | H. muridarum | H. pylori | H. pylori | H. bilis | H. bilis | H. heilmannii | H. typhlonius | H. cinaedi | H. hepaticus | H. pullorum | C. jejuni | C. jejuni | Heli | |||
| PCR-Template | Gene | gA | ggt | gyA | ggt | gA | ggt | gyA | gA | gyA | gyA | gyA | gyA | ggt | 16SrRNA | polA |
| H. muridarum | gyA | 340a | 1 b | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ggt | 1 | 124 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| H. pylori | gyA | 1 | 1 | 30 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ggt | 1 | 1 | 1 | 136 | 1 | 1 | 1 | 1 | 4 c | 1 | 1 | 1 | 1 | 1 | 1 | |
| H. bilis | gA | 1 | 1 | 1 | 1 | 259 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ggt | 1 | 1 | 1 | 1 | 1 | 644 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| H. heilmannii | gyA | 1 | 1 | 1 | 1 | 1 | 1 | 87 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| H. typhlonius | gA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 123 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| H. cinaedi | gyA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 5 | 226 | 1 | 1 | 1 | 1 | 1 | 1 |
| H. hepaticus | gyA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 639 | 1 | 1 | 1 | 1 | 1 |
| H. pullorum | gyA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 512 | 1 | 1 | 1 | 1 |
| C. jejuni | gyA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 7 | 1 | 1 | 1 |
| ggt | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 92 | 1 | 1 | |
| Helicobacter genus | 16SrRNA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 442 | 1 |
| M. musculus | polA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 12 | 1 | 1 | 62 | 1 | 1 | 1 | 1152 |
| H. canis | whole genome | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 610 | 1 |
| C. lari | whole genome | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 16 | 1 | 1 | 1 | 862 |
| C. coli | whole genome | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| murine faeces | whole genome | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 10 | 1 | 1 | 50 | 1 | 1 | 1 | 1095 |
| N (%) by DNA Result | |||||
|---|---|---|---|---|---|
| Antigen | Cut-Off [MFI] a | Negative (n = 57) | H. bilis (n = 26) | H. hepaticus (n = 86) | H. typhlonius (n = 23) |
| H. hepaticus | |||||
| HH0243 | 128 | 2 (4) | 8 (31) | 20 (23) | 3 (13) |
| HH0407 | 956 | 2 (4) | 0 (0) | 1 (1) | 0 (0) |
| HH0435 | 247 | 2 (4) | 9 (35) | 61 (71) | 10 (44) |
| HH0713 | 73 | 1 (2) | 1 (4) | 55 (64) | 1 (4) |
| HH0966 | 115 | 2 (4) | 0 (0) | 25 (29) | 2 (9) |
| HH1201 | 136 | 2 (4) | 0 (0) | 11 (13) | 0 (0) |
| HH1446 | 168 | 2 (4) | 1 (4) | 11 (13) | 4 (17) |
| H. bilis | |||||
| HRAG_00845 | 475 | 2 (4) | 0 (0) | 4 (5) | 0 (0) |
| HRAG_01298 | 84 | 2 (4) | 4 (15) | 6 (7) | 2 (9) |
| HRAG_01470 | 95 | 2 (4) | 2 (8) | 8 (9) | 2 (9) |
| HRAG_01504 | 89 | 2 (4) | 4 (15) | 1 (1) | 1 (4) |
| HRAG_01818 | 146 | 2 (4) | 2 (8) | 6 (7) | 5 (22) |
| P167C | 89 | 2 (4) | 12 (46) | 11 (13) | 4 (17) |
| P167D | 288 | 2 (4) | 16 (62) | 9 (11) | 5 (22) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butt, J.; Schmitz, M.; Berkus, B.; Schmidt, K.; Höfler, D. Validation of Multiplex PCR and Serology Detecting Helicobacter Species in Mice. Microorganisms 2023, 11, 249. https://doi.org/10.3390/microorganisms11020249
Butt J, Schmitz M, Berkus B, Schmidt K, Höfler D. Validation of Multiplex PCR and Serology Detecting Helicobacter Species in Mice. Microorganisms. 2023; 11(2):249. https://doi.org/10.3390/microorganisms11020249
Chicago/Turabian StyleButt, Julia, Mareike Schmitz, Bernhard Berkus, Katja Schmidt, and Daniela Höfler. 2023. "Validation of Multiplex PCR and Serology Detecting Helicobacter Species in Mice" Microorganisms 11, no. 2: 249. https://doi.org/10.3390/microorganisms11020249
APA StyleButt, J., Schmitz, M., Berkus, B., Schmidt, K., & Höfler, D. (2023). Validation of Multiplex PCR and Serology Detecting Helicobacter Species in Mice. Microorganisms, 11(2), 249. https://doi.org/10.3390/microorganisms11020249