

Comparative Analysis between Salmonella enterica Isolated from Imported and Chinese Native Chicken Breeds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Strategy and Isolation and Identification of Salmonella
2.2. Serological Identification and Biochemical Identification of Salmonella Isolates
2.3. Antimicrobial Susceptibility Testing
2.4. ESBL Screening
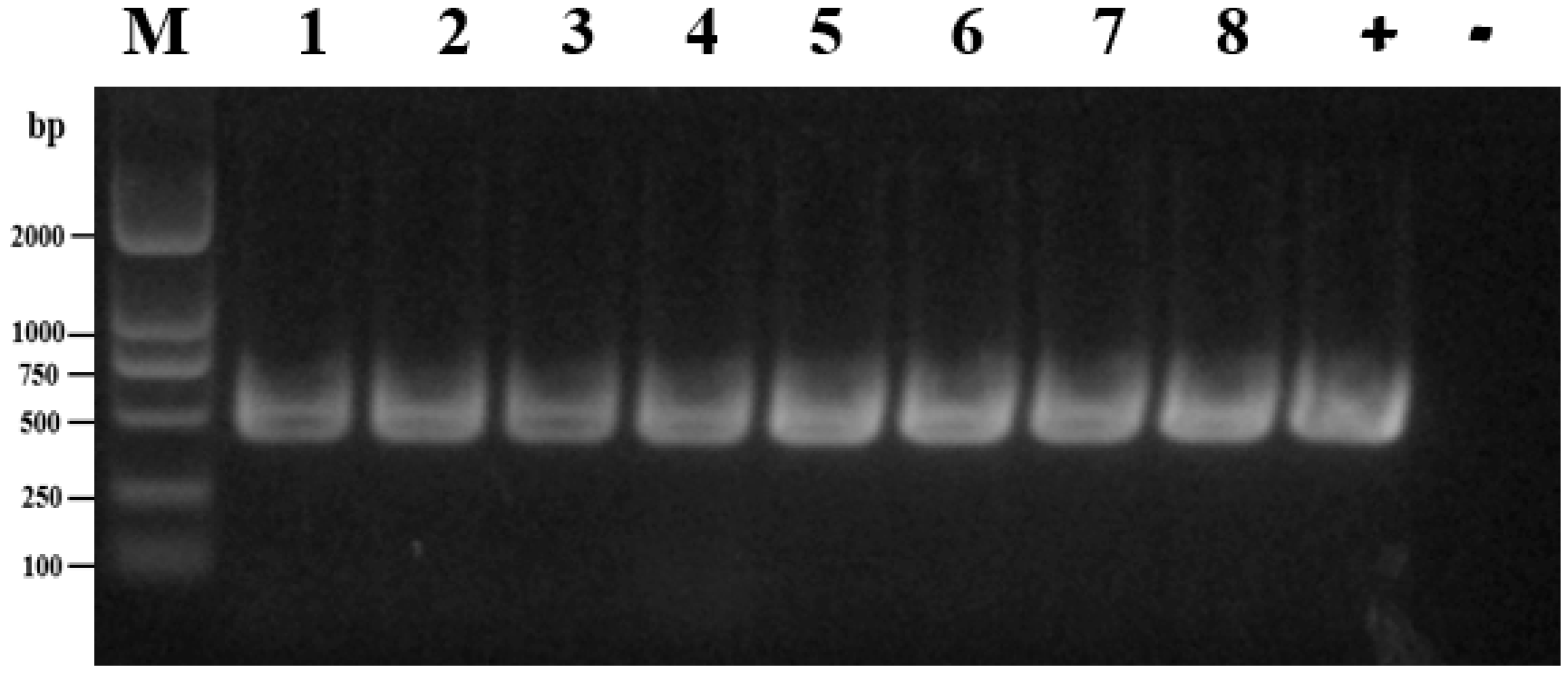
2.5. Molecular Detection of Antimicrobial Resistance-Associated Genes
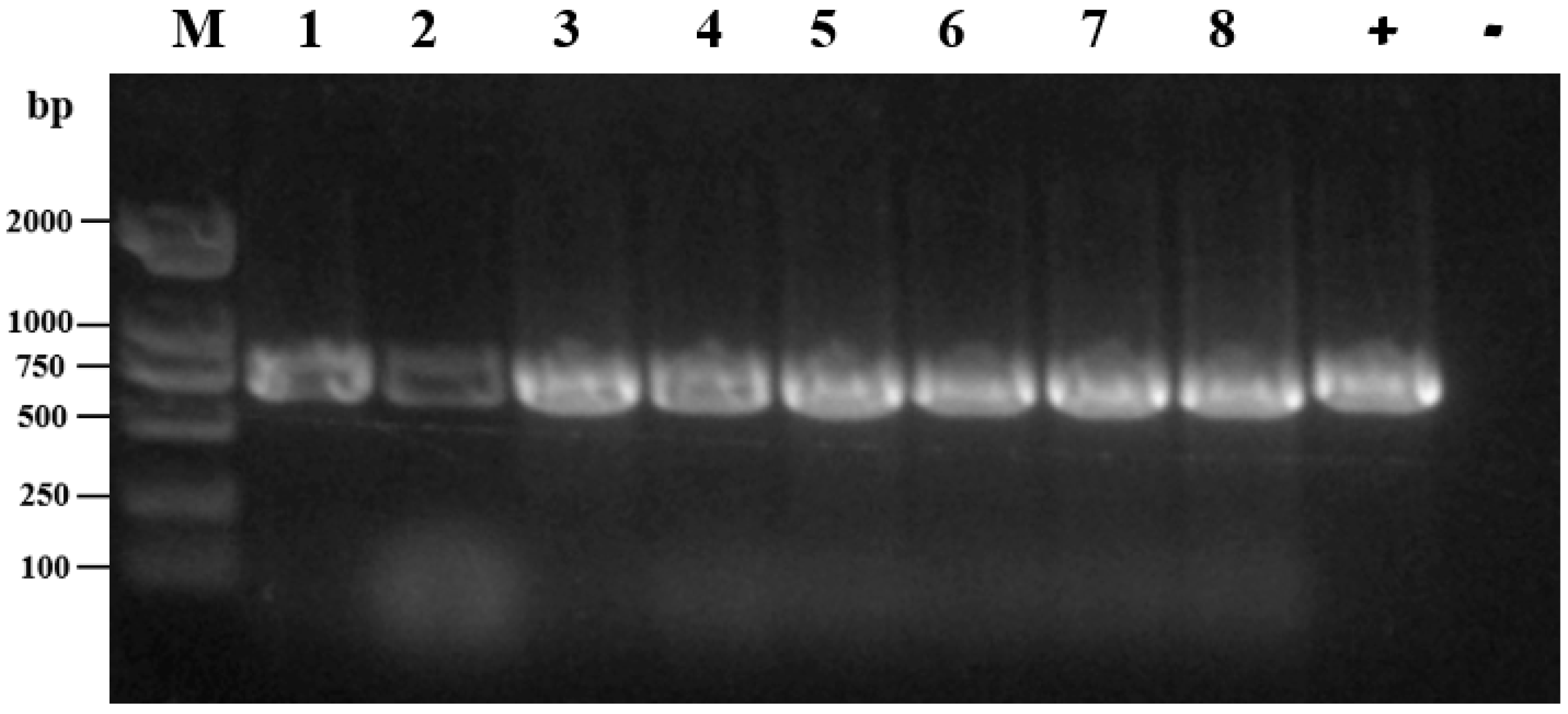
2.6. ERIC-PCR
2.7. Statistical Analysis
3. Results
3.1. Prevalence of Salmonella Isolates
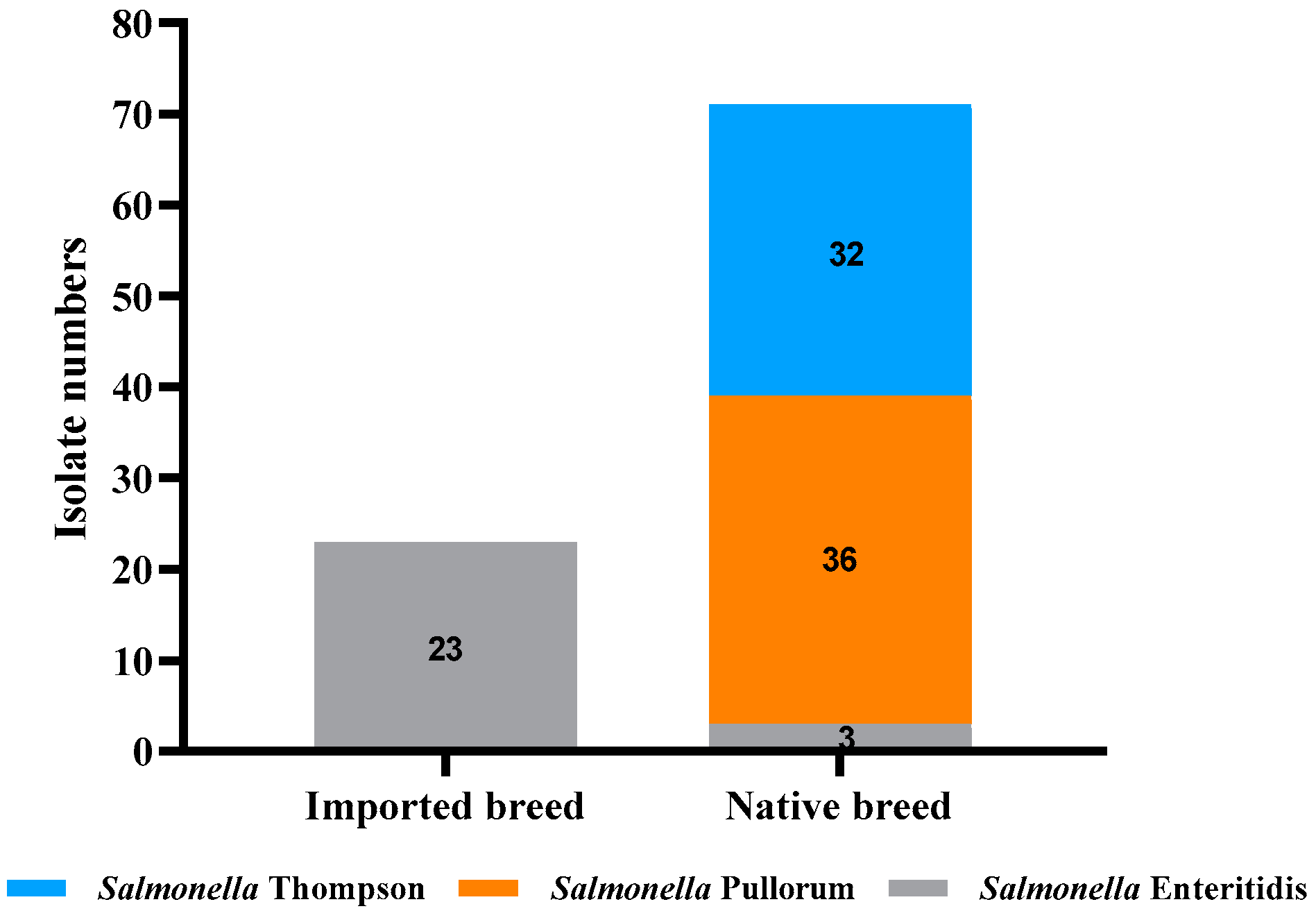
3.2. Distribution of Salmonella Serotypes
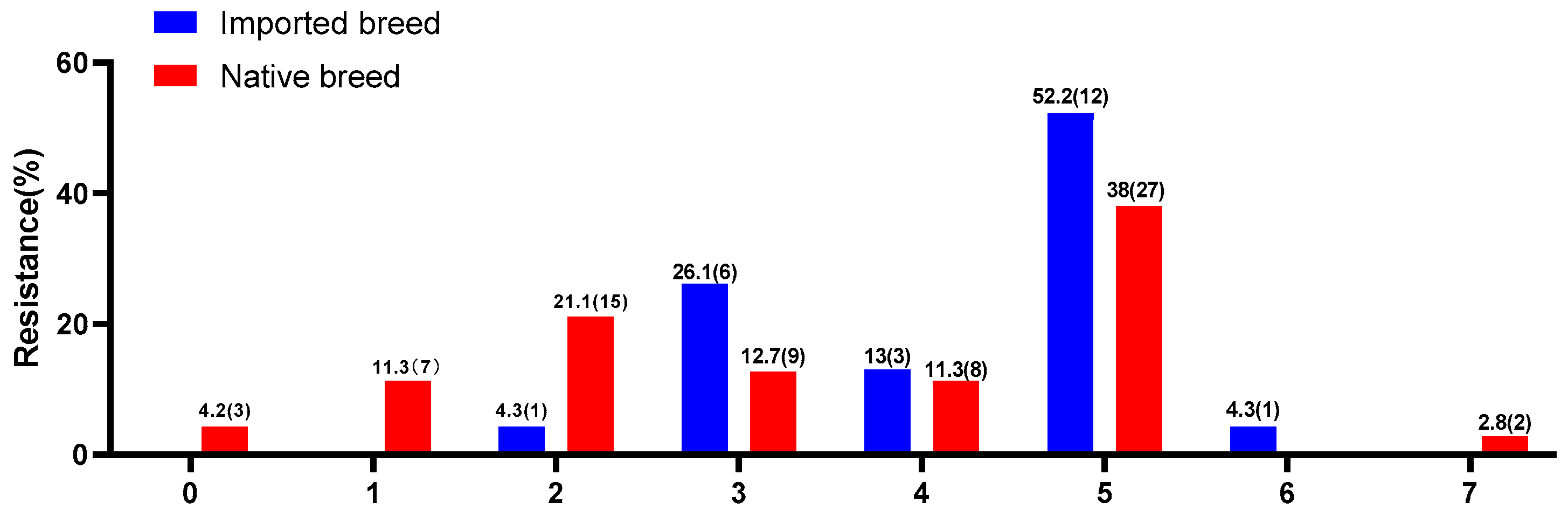
3.3. Antimicrobial Susceptibility and ESBL Production
3.4. Characterization of Antimicrobial Resistance Genes
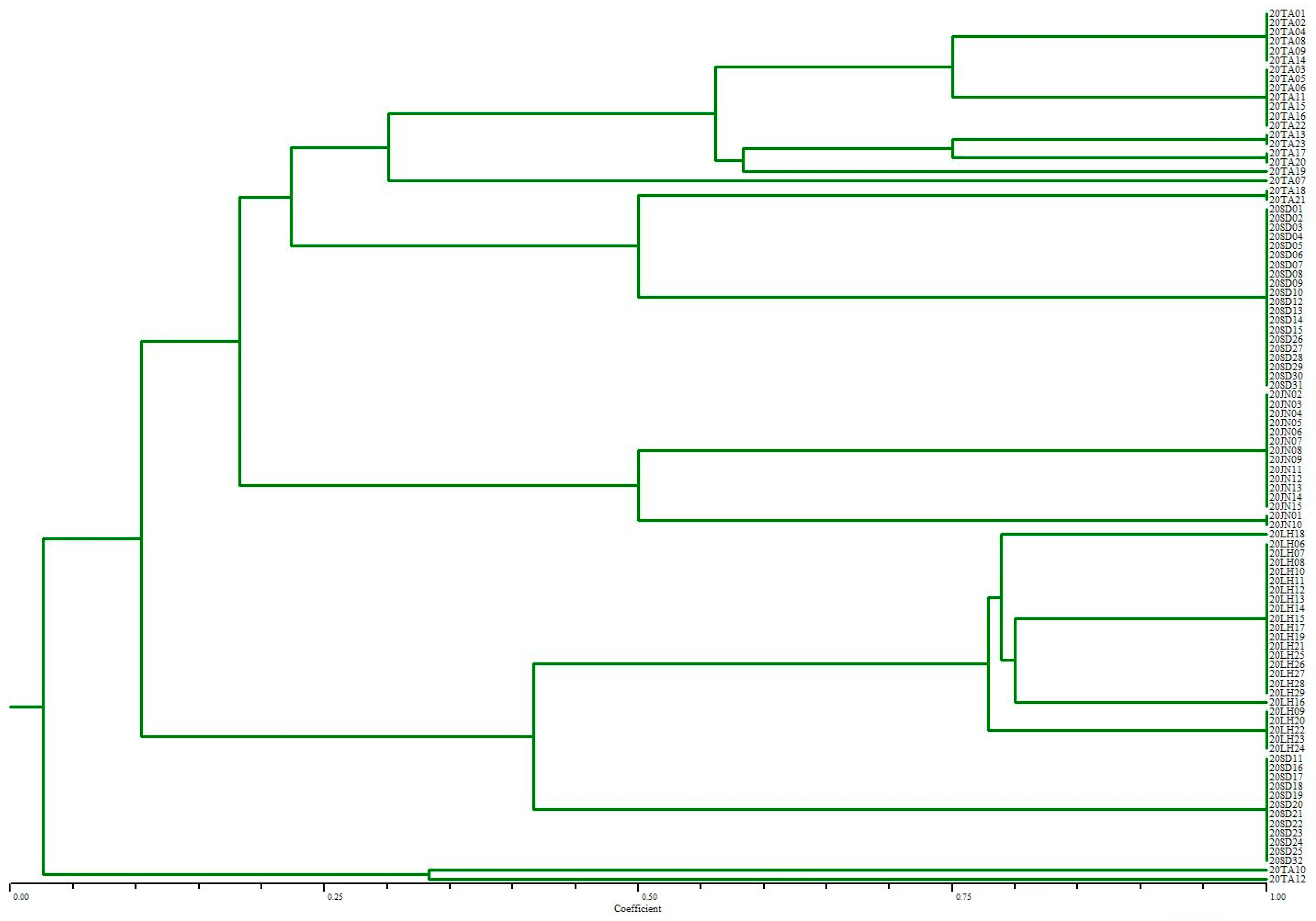
3.5. ERIC-PCR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abd El-Ghany, W.A. Salmonellosis: A Food Borne Zoonotic and Public Health Disease in Egypt. J. Infect. Dev. Ctries 2020, 14, 674–678. [Google Scholar] [CrossRef]
- Ha, J.S.; Seo, K.W.; Kim, Y.B.; Kang, M.S.; Song, C.S.; Lee, Y.J. Prevalence and Characterization of Salmonella in Two Integrated Broiler Operations in Korea. Ir. Vet. J. 2018, 71, 3. [Google Scholar] [CrossRef]
- Ammar, A.M.; Mohamed, A.A.; Abd El-Hamid, M.I.; El-Azzouny, M.M. Virulence Genotypes of Clinical SalmonellaSerovars from Broilers in Egypt. J. Infect. Dev. Ctries 2016, 10, 337–346. [Google Scholar] [CrossRef]
- El-Sharkawy, H.; Tahoun, A.; El-Gohary, A.E.A.; El-Abasy, M.; El-Khayat, F.; Gillespie, T.; Kitade, Y.; Hafez, H.M.; Neubauer, H.; El-Adawy, H. Epidemiological, Molecular Mharacterization and Antibiotic Resistance of Salmonella enterica Serovars Isolated from Chicken Farms in Egypt. Gut. Pathog. 2017, 9, 8. [Google Scholar] [CrossRef]
- Merwad, A.M.A.; Abdel-Haliem, M.E.F. Isolation and Initial Characterization of a Myoviridae Phage for Controlling Zoonotic Salmonella Typhimurium and Salmonella Enteritidis from Broilers in Egypt. Vet. Res. 2018, 55, 5–15. [Google Scholar]
- Paudyal, N.; Yue, M. Antimicrobial Resistance in the “Dark Matter”. Clin. Infect. Dis. 2019, 69, 379–380. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Food Safety. Available online: https://www.who.int/en/news-room/fact-sheets/detail/food-safety (accessed on 24 May 2021).
- Pan, H.; Zhou, X.; Chai, W.; Paudyal, N.; Li, S.; Zhou, X.; Zhou, K.; Wu, Q.; Wu, B.; Li, G.; et al. Diversified Sources for Human Infections by Salmonella enterica Serovar Newport. Transbound. Emerg. Dis. 2019, 66, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kang, X.; Ed-Dra, A.; Zhou, X.; Jia, C.; Müller, A.; Liu, Y.; Kehrenberg, C.; Yue, M. Genome-Based Assessment of Antimicrobial Resistance and Virulence Potential of Isolates of Non-Pullorum/Gallinarum Salmonella Serovars Recovered from Dead Poultry in China. Microbiol. Spectr. 2022, 10, e0096522. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, F.; Liu, Y.; Song, Y.; Zhang, L.; Zhang, F.; Gu, X.; Sun, S. Occurrence and Characterization of Salmonella Isolated from Chicken Breeder Flocks in Nine Chinese Provinces. Front. Vet. Sci. 2020, 7, 479. [Google Scholar] [CrossRef]
- Berchieri, A., Jr.; Murphy, C.K.; Marston, K.; Barrow, P.A. Observations on the Persistence and Vertical Transmission of Salmonella Enterica Serovars Pullorum and Gallinarum in Chickens: Effect of Bacterial and Host Genetic Background. Avian. Pathol. 2001, 30, 221–231. [Google Scholar] [CrossRef]
- OIE. Pullorum Disease Timelines (1996–2020). World Organisation for Animal Health (OIE), Paris, France 2020. Available online: https://www.oie.int/wahis_2/public/wahid.php/Diseaseinformation/Diseasetimelines (accessed on 14 December 2022).
- Zhang-Barber, L.; Turner, A.K.; Barrow, P.A. Vaccination for Control of Salmonella in Poultry. Vaccine 1999, 17, 2538–2545. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, J.; Zhang, Y.; Liu, S.; Chen, L.; Xiao, C.; Zeng, H.; Wei, X.; Gu, Q.; Li, Y.; et al. Prevalence, Abundance, Serovars and Antimicrobial Resistance of Salmonella Isolated from Retail Raw Poultry Meat in China. Sci. Total Environ. 2020, 713, 136385. [Google Scholar] [CrossRef]
- Fall-Niang, N.K.; Sambe-Ba, B.; Seck, A.; Deme, S.N.; Wane, A.A.; Bercion, R.; Alambedji-Bada, R.; Gassama-Sow, A. Antimicrobial Resistance Profile of Salmonella Isolates in Chicken Carcasses in Dakar, Senegal. Foodborne Pathog. Dis. 2019, 16, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Stephen, J.; Lekshmi, M.; Ojha, M.; Wenzel, N.; Sanford, L.; Hernandez, A.; Parvathi, A.; Kumar, S.H. Bacterial Resistance to Antimicrobial Agents. Antibiotics 2021, 10, 593. [Google Scholar] [CrossRef]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A Review of Antibiotic Use in Food Animals: Perspective, Policy, and Potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef]
- Alekshun, M.N.; Levy, S.B. Molecular Mechanisms of Antibacterial Multidrug Resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef]
- McDermott, P.F.; Zhao, S.; Tate, H. Antimicrobial Resistance in Nontyphoidal Salmonella. Microbiol. Spectr. 2018, 6, 261–287. [Google Scholar] [CrossRef]
- Jajere, S.M. A Review of Salmonella Enterica with Particular Focus on the Pathogenicity and Virulence Factors, Host Specificity and Antimicrobial Resistance Including Multidrug Resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef]
- Rincón-Gamboa, S.M.; Poutou-Piñales, R.A.; Carrascal-Camacho, A.K. Antimicrobial Resistance of Non-Typhoid Salmonella in Meat and Meat Products. Foods 2021, 10, 1731. [Google Scholar] [CrossRef]
- Sheykhsaran, E.; Baghi, H.B.; Soroush, M.H.; Ghotaslou, R. An Overview of Tetracyclines and Related Resistance Mechanisms. Rev. Res. Med. Microbiol. 2019, 30, 69–75. [Google Scholar] [CrossRef]
- Lees, P.; Pelligand, L.; Giraud, E.; Toutain, P.L. A History of Antimicrobial Drugs in Animals: Evolution and Revolution. J. Vet. Pharmacol. Ther. 2021, 44, 137–171. [Google Scholar] [CrossRef]
- Xiang, R.; Ye, X.; Tuo, H.; Zhang, X.; Zhang, A.; Lei, C.; Yang, Y.; Wang, H. Co-Occurrence of Mcr-3 and Bla(Ndm-5) Genes in Multidrug-Resistant Klebsiella Pneumoniae St709 from a Commercial Chicken Farm in China. Int. J. Antimicrob. Agents 2018, 52, 519–520. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Feng, C.; Wang, P.; Yu, L.; Liu, D.; Sun, S.; Wang, F. Distribution Characteristics and Potential Risks of Heavy Metals and Antimicrobial Resistant Escherichia coli in Dairy Farm Wastewater in Tai’an, China. Chemosphere 2021, 262, 127768. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yu, L.; Zhang, Y.; Dai, Y.; Wang, P.; Feng, C.; Liu, M.; Sun, S.; Xie, Z.; Wang, F. Prevalence and Characteristics of Multidrug-Resistant Mcr-1-Positive Escherichia coli Isolates from Broiler Chickens in Tai’an, China. Poult. Sci. 2020, 99, 1117–1123. [Google Scholar] [CrossRef]
- Archambault, M.; Petrov, P.; Hendriksen, R.S.; Asseva, G.; Bangtrakulnonth, A.; Hasman, H.; Aarestrup, F.M. Molecular Characterization and Occurrence of Extended-Spectrum Beta-Lactamase Resistance Genes among Salmonella enterica Serovar Corvallis from Thailand, Bulgaria, and Denmark. Microb. Drug. Resist. 2006, 12, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Dinh, A.; Duran, C.; Singh, S.; Tesmoingt, C.; Bouabdallah, L.; Hamon, A.; Antignac, M.; Ourghanlian, C.; Loustalot, M.C.; Pain, J.B.; et al. Real-life Temocillin Use in Greater Paris Area, Effectiveness and Risk Factors for Failure in Infections Caused by ESBL-Producing Enterobacterales: A Multicentre Retrospective Study. JAC-Antimicrob. Resist. 2022, 5, dlac132. [Google Scholar] [CrossRef]
- Orabi, A.; Armanious, W.; Radwan, I.A.; Girh, Z.M.S.A.; Hammad, E.; Diab, M.S.; Elbestawy, A.R. Genetic Correlation of Virulent Salmonella Serovars (Extended Spectrum β-Lactamases) Isolated from Broiler Chickens and Human: A Public Health Concern. Pathogens 2022, 11, 1196. [Google Scholar] [CrossRef]
- The Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; CDC: Atlanta, GA, USA, 2013.
- WHO. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis. 2017. Available online: www.who.int/medicines/areas/rational_use/PPLreport_2017_09_19.pdf?ua=1 (accessed on 14 December 2022).
- Yang, J.; Gao, S.; Chang, Y.; Su, M.; Xie, Y.; Sun, S. Occurrence and Characterization of Salmonella Isolated from Large-Scale Breeder Farms in Shandong Province, China. BioMed Res. Int. 2019, 8159567. [Google Scholar] [CrossRef] [PubMed]
- ISO 6579:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella. International Organization for Standardization: Geneva, Switzerland, 2017.
- GB 4789.4-2016; National Food Safety Standard Food Microbiological Examination: Salmonella. National Health and Family Planning Commission of the People’s Republic of China and China Food and Drug Administration: Beijing, China, 2016.
- Zhang, F.; Song, Y.; Xu, Y.; Ju, Z.; Cui, Z.; Sun, S. Establishment of Identification Medium and Pcr Assay for Salmonella from Poultry. Shangdong J. Anim. Sci. Vet. Med. 2019, 10. [Google Scholar]
- Yang, B.; Niu, Q.; Yang, Y.; Dai, P.; Yuan, T.; Xu, S.; Pan, X.; Yang, Y.; Zhu, G. Self-made Salmonella Pullorum Agglutination Antigen Development and Its Potential Practical Application. Poult. Sci. 2019, 98, 6326–6332. [Google Scholar] [CrossRef]
- Grimont, P.A.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars. WHO Collab. Cent. Ref. Res. Salmonella 2007, 9, 1–166. [Google Scholar]
- Xiong, D.; Song, L.; Pan, Z.; Jiao, X. Identification and Discrimination of Salmonella Enterica Serovar Gallinarum Biovars Pullorum and Gallinarum Based on a One-Step Multiplex PCR Assay. Front. Microbiol. 2018, 9, 1718. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. M100, 32nd Ed: Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Motoi, Y.; Sato, M.; Maruyama, A.; Watanabe, H.; Fukumoto, Y.; Shimamoto, T. Zoo Animals as Reservoirs of Gram-Negative Bacteria Harboring Integrons and Antimicrobial Resistance Genes. Appl. Environ. Microbiol. 2007, 73, 6686–6690. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex Pcr for Detection of Acquired Carbapenemase Genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Pérez-Pérez, F.J.; Hanson, N.D. Detection of Plasmid-Mediated Ampc Beta-Lactamase Genes in Clinical Isolates by Using Multiplex Pcr. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Guo, Q.; Xu, X.; Wang, X.; Ye, X.; Wu, S.; Hooper, D.C.; Wang, M. New Plasmid-mediated Quinolone Resistance Gene, QnrC, Found in a Clinical Isolate of Proteus Mirabilis. Antimicrob. Agents Chemother. 2009, 53, 1892–1897. [Google Scholar] [CrossRef]
- Hansen, L.H.; Sørensen, S.J.; Jørgensen, H.S.; Jensen, L.B. The prevalence of the OqxAB Multidrug Efflux Pump Amongst Olaquindox-Resistant Escherichia Coli in Pigs. Microb. Drug Resist. 2005, 11, 378–382. [Google Scholar] [CrossRef]
- Sáenz, Y.; Briñas, L.; Domínguez, E.; Ruiz, J.; Zarazaga, M.; Vila, J.; Torres, C. Mechanisms of Resistance in Multiple-Antibiotic-Resistant Escherichia Coli Strains of Human, Animal, and Food Origins. Antimicrob. Agents Chemother. 2004, 48, 3996–4001. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.Y.; Wang, H.N.; Tian, G.B.; Zhang, Y.; Yang, X.; Xia, Q.Q.; Tang, J.N.; Zou, L.K. Phenotypic and Genotypic Characterisation of Antimicrobial Resistance in Faecal Bacteria from 30 Giant Pandas. Int. J. Antimicrob. Agents 2009, 33, 456–460. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel Plasmid-Mediated Colistin Resistance Mcr-4 Gene in Salmonella and Escherichia Coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Shu, R.; Zhao, Y.; Zhang, Q.; Xu, X.; Zhou, G. Analysis of ERIC-PCR Genomic Polymorphism of Salmonella Isolates from Chicken Slaughter Line. Eur. Food Res. Technol. 2014, 239, 543–548. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, X.; Jiang, Z.; Qi, Y.; Ed-Dra, A.; Yue, M. Epidemiological Investigation and Antimicrobial Resistance Profiles of Salmonella Isolated from Breeder Chicken Hatcheries in Henan, China. Front. Cell Infect Microbiol. 2020, 10, 497. [Google Scholar] [CrossRef]
- Zhao, X.; Ju, Z.; Wang, G.; Yang, J.; Wang, F.; Tang, H.; Zhao, X.; Sun, S. Prevalence and Antimicrobial Resistance of Salmonella Isolated from Dead-in-Shell Chicken Embryos in Shandong, China. Front. Vet. Sci. 2021, 8, 581946. [Google Scholar] [CrossRef]
- Abdeen, E.; Elmonir, W.; Suelam, I.I.A.; Mousa, W.S. Antibiogram and Genetic Diversity of Salmonella enterica with Zoonotic Potential Isolated from Morbid Native Chickens and Pigeons in Egypt. J. Appl. Microbiol. 2018, 124, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Liu, Q.; Jiang, Z.; Song, Y.; Yi, S.; Qiu, J.; Hao, G.; Sun, S. Characteristics of Salmonella from Chinese Native Chicken Breeds Fed on Conventional or Antibiotic-Free Diets. Front. Vet. Sci. 2021, 8, 607491. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, J.; Xu, M.; Zhu, C.; Yu, Y.; Liu, X.; Kelly, P.; Xu, B.; Wang, C. Prevalence and Fimbrial Genotype Distribution of Poultry Salmonella Isolates in China (2006 to 2012). Appl. Environ. Microbiol. 2014, 80, 687–693. [Google Scholar] [CrossRef]
- Li, H.F.; Hu, Y.; Hu, H.; Song, C.; Shu, J.T.; Zhu, C.H.; Zhang, S.J.; Fan, J.H.; Chen, W.W. Genetic Differ in Tlr4 Gene Polymorphisms and Expression Involved in Salmonella Natural and Artificial Infection Respectively in Chinese Native Chicken Breeds. Mol. Biol. Rep. 2013, 40, 5005–5012. [Google Scholar] [CrossRef]
- Lei, C.W.; Zhang, Y.; Kang, Z.Z.; Kong, L.H.; Tang, Y.Z.; Zhang, A.Y.; Yang, X.; Wang, H.N. Vertical Transmission of Salmonella Enteritidis with Heterogeneous Antimicrobial Resistance from Breeding Chickens to Commercial Chickens in China. Vet. Microbiol. 2020, 240, 108538. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, J.; Zhang, B.; Sun, S.; Chang, W. Characterization of Integrons and Resistance Genes in Salmonella Isolates from Farm Animals in Shandong Province, China. Front. Microbiol. 2017, 8, 1300. [Google Scholar] [CrossRef] [PubMed]
- Elbediwi, M.; Tang, Y.; Shi, D.; Ramadan, H.; Xu, Y.; Xu, S.; Li, Y.; Yue, M. Genomic Investigation of Antimicrobial-Resistant Salmonella enterica Isolates from Dead Chick Embryos in China. Front. Microbiol. 2021, 12, 684400. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The Antibiotic Resistome: The Nexus of Chemical and Genetic Diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.M.; Attia, A.M.; Abd El-Aziz, N.K.; Abd El Hamid, M.I.; El-Demerdash, A.S. Class 1 Integron and Associated Gene Cassettes Mediating Multiple-Drug Resistance in Some Food Borne Pathogens. Int. Food Res. J. 2016, 23, 332. [Google Scholar]
- Castro-Vargas, R.E.; Herrera-Sanchez, M.P.; Rodriguez-Hernandez, R.; Rondon-Barragan, I.S. Antibiotic Resistance in Salmonella Spp. Isolated from Poultry: A Global Overview. Vet. World 2020, 13, 2070–2084. [Google Scholar] [CrossRef] [PubMed]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The Ecology of Extended-Spectrum Β-Lactamases (Esbls) in the Developed World. J. Travel. Med. 2017, 24, S44–S51. [Google Scholar] [CrossRef]
- Kuang, D.; Zhang, J.; Xu, X.; Shi, W.; Yang, X.; Su, X.; Shi, X.; Meng, J. Increase in Ceftriaxone Resistance and Widespread Extended-Spectrum Β-Lactamases Genes among Salmonella enterica from Human and Nonhuman Sources. Foodborne Pathog. Dis. 2018, 15, 770–775. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Deekshit, V.K.; Kumar, B.K.; Rai, P.; Srikumar, S.; Karunasagar, I.; Karunasagar, I. Detection of class 1 integrons in Salmonella Weltevreden and silent antibiotic resistance genes in some seafood-associated nontyphoidal isolates of Salmonella in south-west coast of India. J. Appl. Microbiol. 2012, 112, 1113–1122. [Google Scholar] [CrossRef]
- Neuert, S.; Nair, S.; Day, M.R.; Doumith, M.; Ashton, P.M.; Mellor, K.C.; Jenkins, C.; Hopkins, K.L.; Woodford, N.; de Pinna, E.; et al. Prediction of Phenotypic Antimicrobial Resistance Profiles from Whole Genome Sequences of Non-typhoidal Salmonella enterica. Front. Microbiol. 2018, 9, 592. [Google Scholar] [CrossRef]
- Galán-Relaño, Á.; Sánchez-Carvajal, J.M.; Gómez-Gascón, L.; Vera, E.; Huerta, B.; Cardoso-Toset, F.; Gómez-Laguna, J.; Astorga, R.J. Phenotypic and genotypic antibiotic resistance patterns in Salmonella Typhimurium and its monophasic variant from pigs in southern Spain. Res. Vet. Sci. 2022, 152, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Schwan, C.L.; Lomonaco, S.; Bastos, L.M.; Cook, P.W.; Maher, J.; Trinetta, V.; Bhullar, M.; Phebus, R.K.; Gragg, S.; Kastner, J.; et al. Genotypic and Phenotypic Characterization of Antimicrobial Resistance Profiles in Non-typhoidal Salmonella enterica Strains Isolated from Cambodian Informal Markets. Front. Microbiol. 2021, 12, 711472. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, W.; Elbediwi, M.; Pan, H.; Wang, L.; Zhou, C.; Zhao, B.; Xu, X.; Li, D.; Yan, X.; et al. Characterization of Salmonella Resistome and Plasmidome in Pork Production System in Jiangsu, China. Front. Vet. Sci. 2020, 7, 617. [Google Scholar] [CrossRef]
- Kaye, K.S.; Pogue, J.M.; Tran, T.B.; Nation, R.L.; Li, J. Agents of Last Resort: Polymyxin Resistance. Infect. Dis. Clin. N. Am. 2016, 30, 391–414. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Farm | Breed | Type of Sample | No. of Sample | No. of Positive Sample (%) |
|---|---|---|---|---|---|
| Tai’an | Farm A | White feather broiler | Dead embryos | 60 | 23 (38.3) |
| Jining | Farm B | Bairi | Dead embryos | 238 | 15 (6.3) |
| Rizhao | Farm C | Langya | Dead embryos | 100 | 29 (29) |
| Cloacal swabs | 100 | 1 (1) | |||
| Water | 15 | - | |||
| Feed | 15 | 2 (13.3) | |||
| Jining | Farm D | Luhua | Dead embryos | 60 | - |
| Meconium of newly hatched chicks | 70 | 24 (34.3) | |||
| Environmental swabs | 30 | - | |||
| Water | 5 | - | |||
| Grand total | 693 | 94 (13.6) |
| Antibiotic Classes | Antibiotic | Number (%) of Resistant Isolates (n = 94) | p Value | |
|---|---|---|---|---|
| Imported Breed | Native Breed | |||
| (n = 23) | (n = 71) | |||
| β-lactames | Ampicillin | 23 (100) | 48 (67.6) | 0.012 * |
| Amoxicillin | 20 (87) | 49 (69) | 0.150 | |
| Cefoxitin | 0 (4.3) | 3 (4.2) | 0.040 * | |
| Ceftazidime | 0 | 15 (21.1) | 0.000 ** | |
| Tetracyclines | Tetracycline | 19 (82.6) | 9 (12.7) | 0.000 ** |
| Doxycycline | 15 (65.2) | 6 (8.4) | 0.000 ** | |
| Quinolones | Ofloxacin | 0 | 0 | - |
| Aminoglycosides | Gentamicin | 1 (4.3) | 29 (40.8) | 0.000 ** |
| Streptomycin | 19 (82.6) | 59 (83.1) | 0.964 | |
| Sulphonamides | Sulfamethoxazole-trimethoprim | 0 | 29 (40.8) | 0.000 ** |
| Number | Source | Breed | Serotype | Resistant Phenotype | Resistant Genes |
|---|---|---|---|---|---|
| 1 | Meconium of newly hatched chicks | Luhua | S. pullorum | STR—CAZ | blaTEM—tetB |
| 2 | Meconium of newly hatched chicks | Luhua | S. pullorum | STR—CAZ | blaTEM |
| 3 | Meconium of newly hatched chicks | Luhua | S. pullorum | STR—CAZ | blaTEM—tetA—tetB |
| 4 | Meconium of newly hatched chicks | Luhua | S. pullorum | TET—STR—AMX—AMP—CAZ | blaTEM—tetB |
| 5 | Meconium of newly hatched chicks | Luhua | S. pullorum | STR—AMX—AMP—CAZ | blaTEM—tetB |
| 6 | Meconium of newly hatched chicks | Luhua | S. pullorum | STR—CAZ | blaTEM—tetA—tetB—sul1—sul2 |
| 7 | Meconium of newly hatched chicks | Luhua | S. pullorum | STR—CAZ | blaTEM—tetA—tetB |
| 8 | Meconium of newly hatched chicks | Luhua | S. pullorum | CAZ | blaTEM |
| Serotypes | Antibiotics | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SXT | GM | STR | OF | AMP | AMX | DOX | TET | CAZ | FOX | |
| Pullorum | 0 | 0 | 75% | 0 | 42% | 39% | 3% | 11% | 53% | 6% |
| Thompson | 91% | 91% | 91% | 0 | 94% | 100% | 6% | 6% | 0 | 0 |
| Enteritidis | 0 | 4% | 85% | 0 | 100% | 88% | 69% | 85% | 0% | 4% |
| Serotypes | Antibiotic Agents | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Pullorum | 100 | 100 | 94 | 40 | 27 | 5 | 0 | 0 |
| Thompson | 0 | 0 | 0 | 20 | 46 | 56 | 0 | 100 |
| Enteritidis | 0 | 0 | 6 | 40 | 27 | 39 | 100 | 0 |
| Antimicrobial Resistance Genes | Number (%) of Resistant Isolates (n = 94) | p Value | |
|---|---|---|---|
| Imported Breed | Native Breed | ||
| (n = 23) | (n = 71) | ||
| blaTEM | 23 (100) | 69 (97.2) | 0.841 |
| AAC2 | 1 (4.3) | 0 | 0.038 * |
| tetA | 20 (87) | 25 (35.2) | 0.000 ** |
| tetB | 16 (70) | 26 (36.6) | 0.001 ** |
| sul1 | 15 (65.2) | 10 (14.1) | 0.000 ** |
| sul2 | 0 | 2 (2.8) | 0.094 |
| Serotypes | Antimicrobial Resistance Genes | |||||
|---|---|---|---|---|---|---|
| blaTEM | AAC2 | tetA | tetB | sul1 | sul2 | |
| Pullorum | 100 | 0 | 61.1 | 61.1 | 22.2 | 2.8 |
| Thompson | 100 | 3.8 | 84.6 | 69.2 | 65.4 | 0 |
| Enteritidis | 93.8 | 0 | 12.5 | 6.2 | 0 | 3.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; Tang, W.; Li, Y.; Han, Y.; Cui, L.; Sun, S. Comparative Analysis between Salmonella enterica Isolated from Imported and Chinese Native Chicken Breeds. Microorganisms 2023, 11, 390. https://doi.org/10.3390/microorganisms11020390
Shi W, Tang W, Li Y, Han Y, Cui L, Sun S. Comparative Analysis between Salmonella enterica Isolated from Imported and Chinese Native Chicken Breeds. Microorganisms. 2023; 11(2):390. https://doi.org/10.3390/microorganisms11020390
Chicago/Turabian StyleShi, Wenjian, Wenli Tang, Yafei Li, Yu Han, Lulu Cui, and Shuhong Sun. 2023. "Comparative Analysis between Salmonella enterica Isolated from Imported and Chinese Native Chicken Breeds" Microorganisms 11, no. 2: 390. https://doi.org/10.3390/microorganisms11020390
APA StyleShi, W., Tang, W., Li, Y., Han, Y., Cui, L., & Sun, S. (2023). Comparative Analysis between Salmonella enterica Isolated from Imported and Chinese Native Chicken Breeds. Microorganisms, 11(2), 390. https://doi.org/10.3390/microorganisms11020390