
The Conjusome—A Transient Organelle Linking Genome Rearrangements in the Parental and Developing Macronuclei

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture Conditions
2.2. Cytogamy and Environmental Shock
2.3. Drugs
2.4. Microcompression and Light Microscopy
2.5. Immunofluorescence
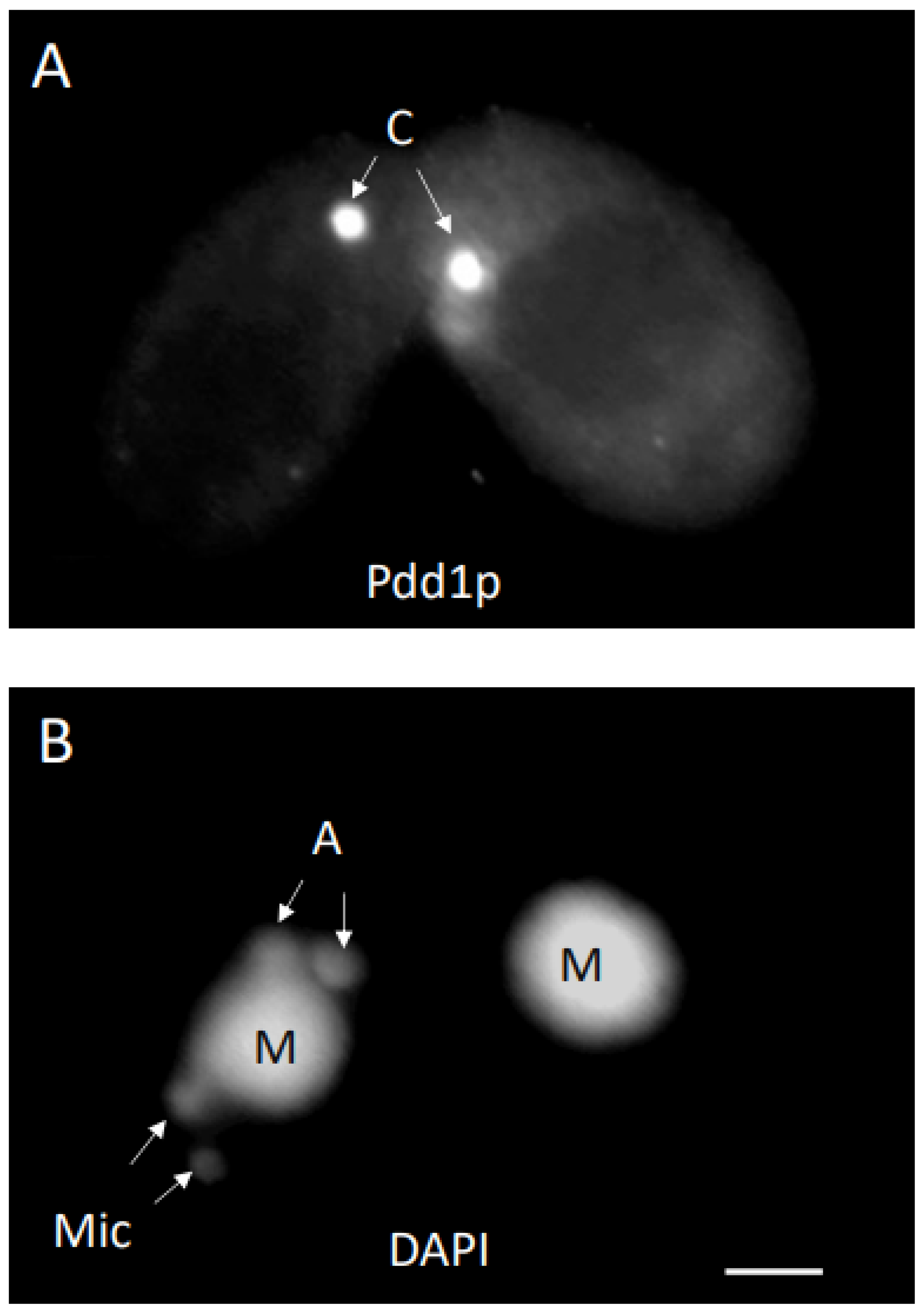
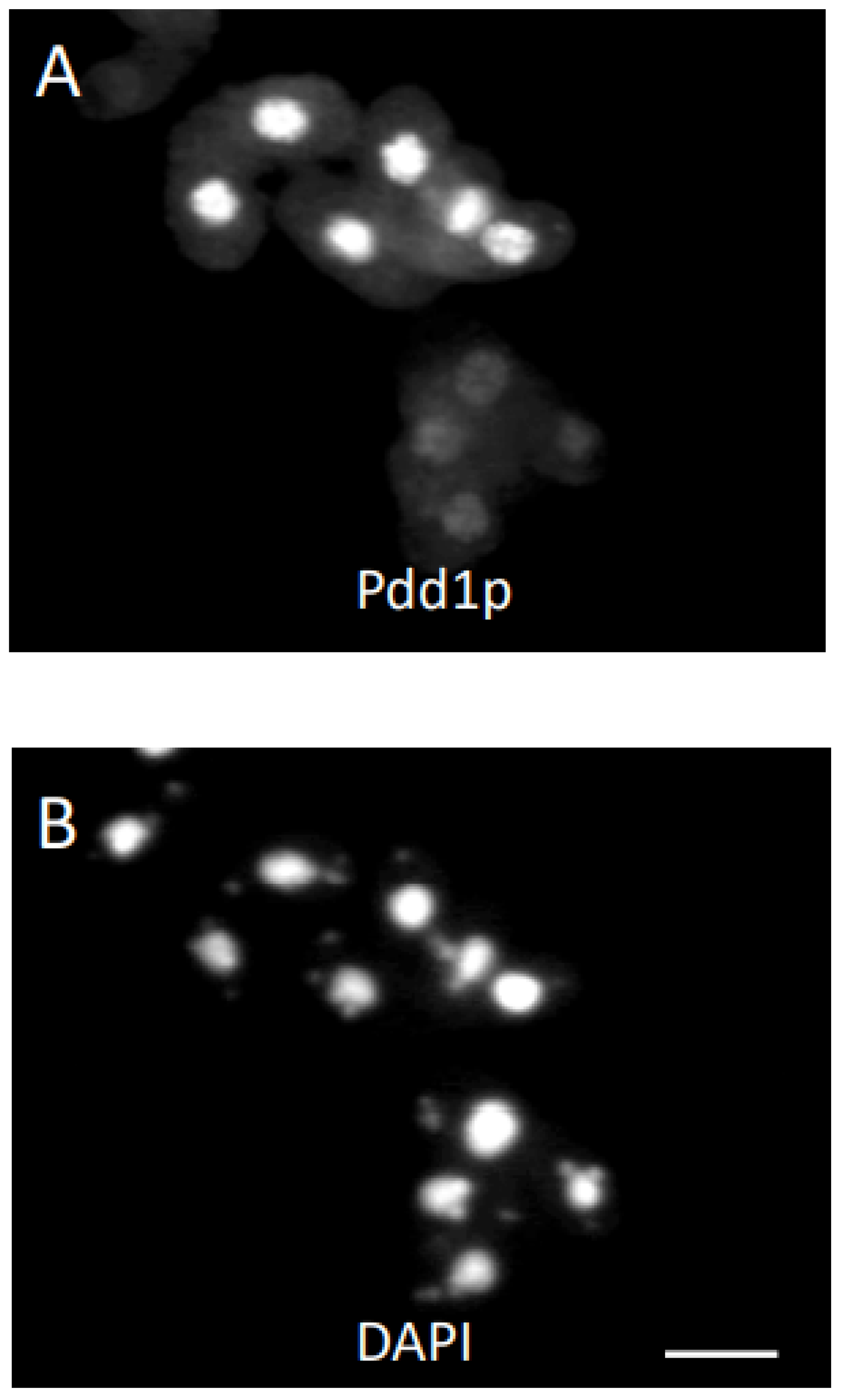
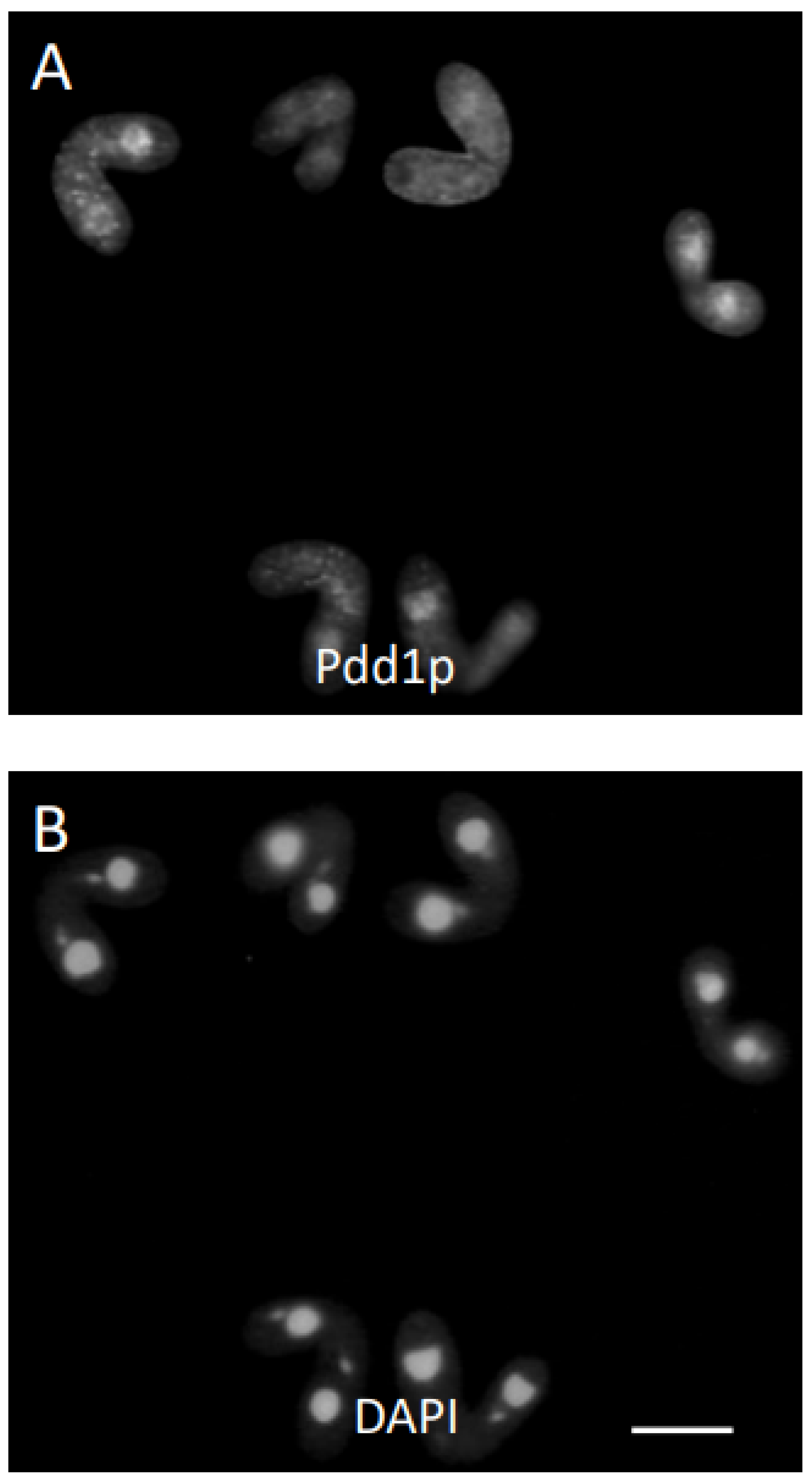
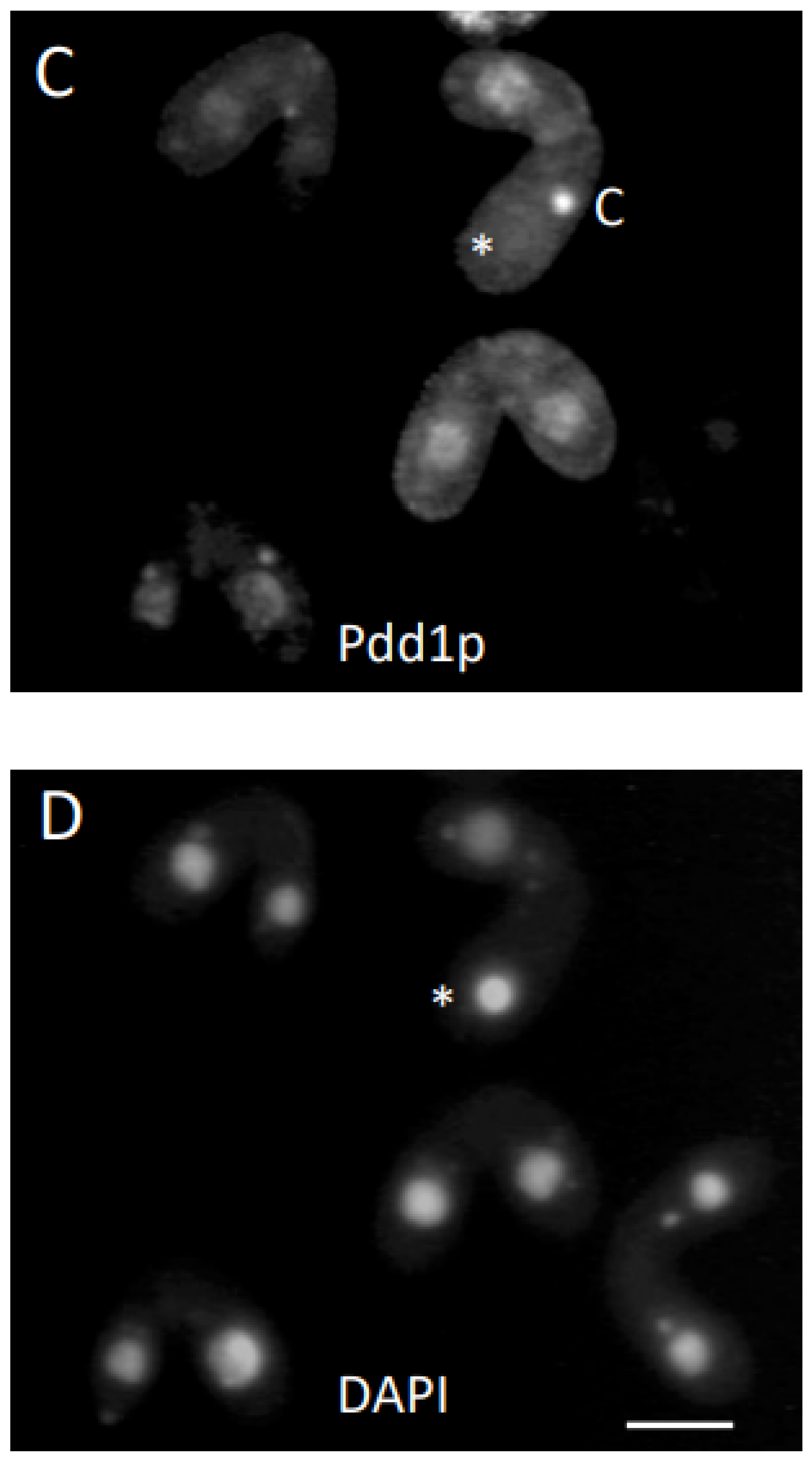
3. Results
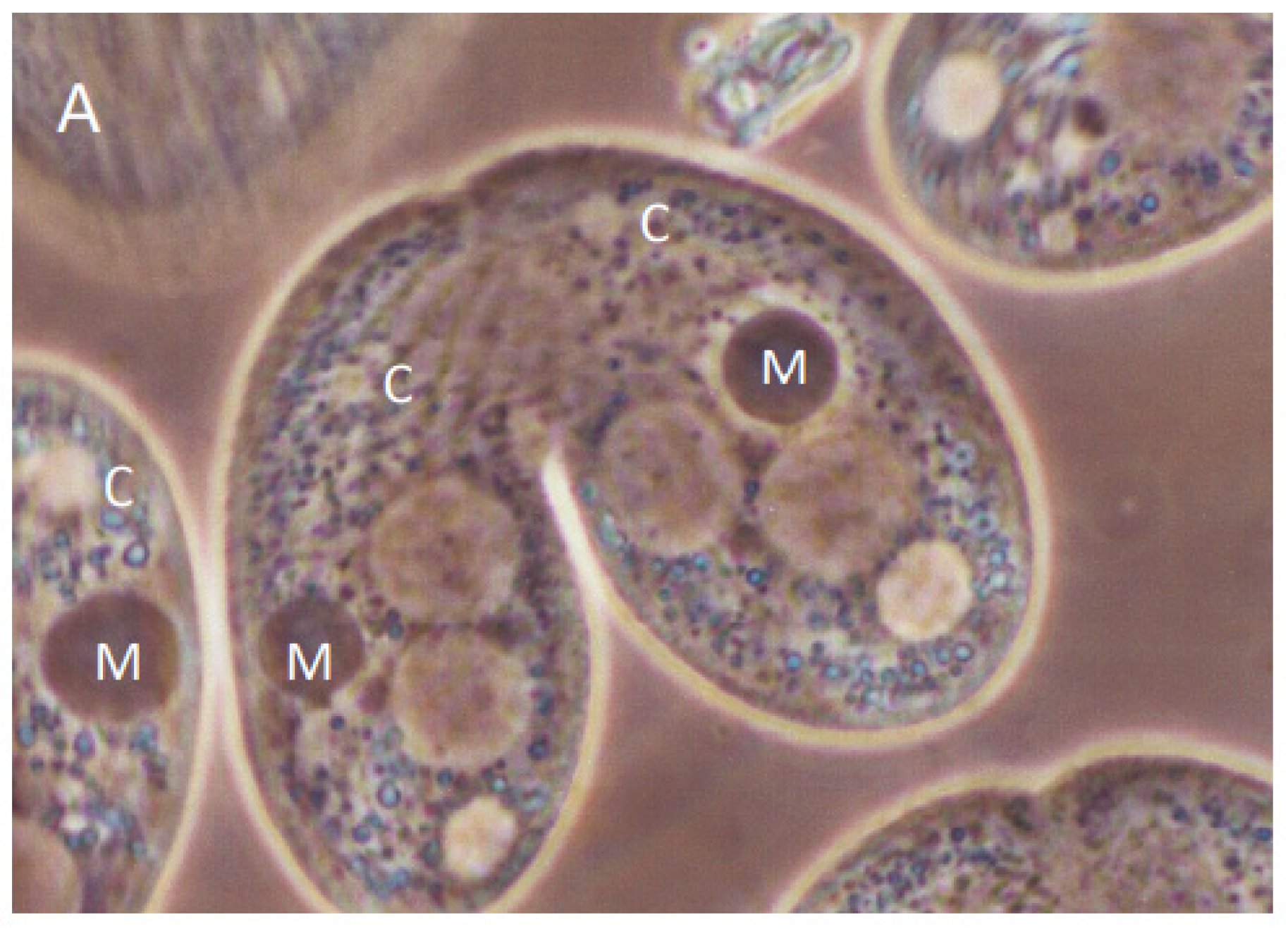
3.1. Analysis of Progeny Derived from Uniparental Cytogamy (UPC)
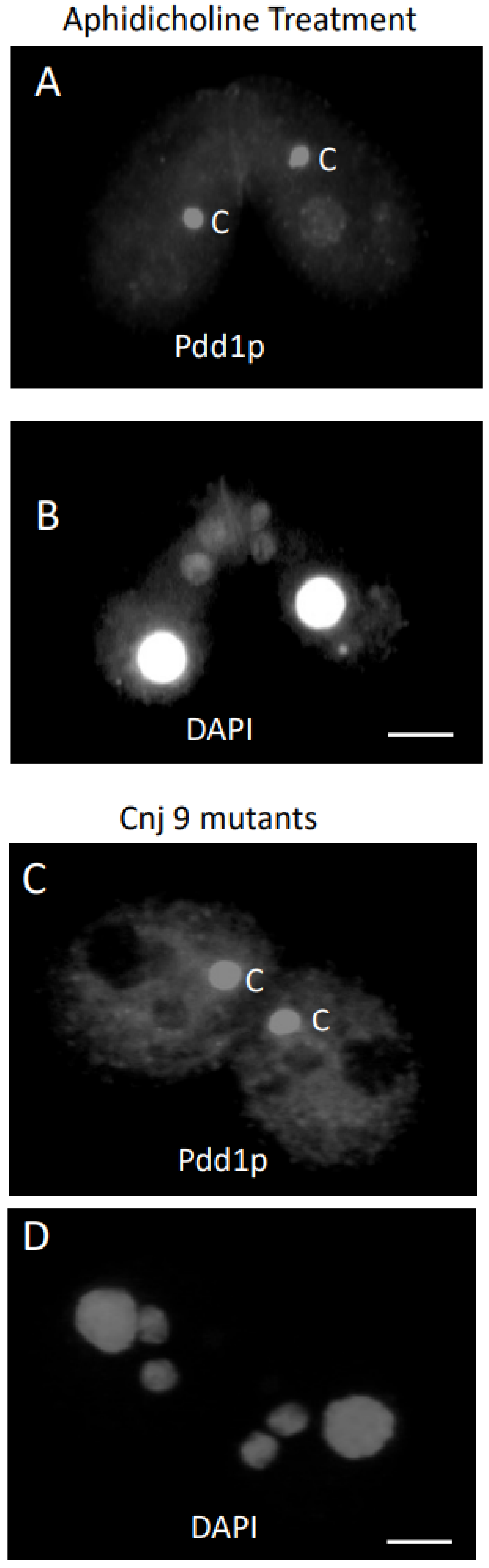
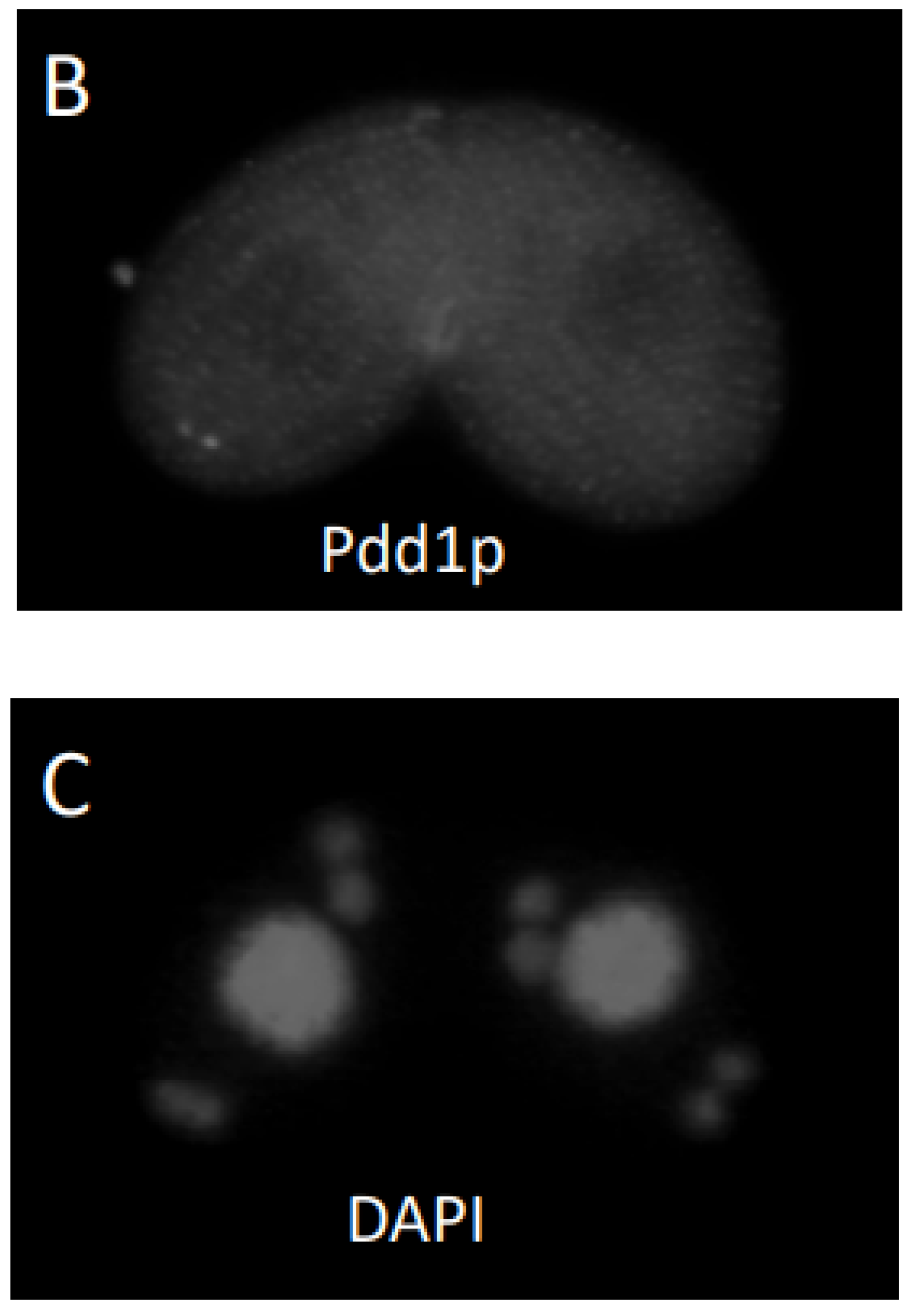
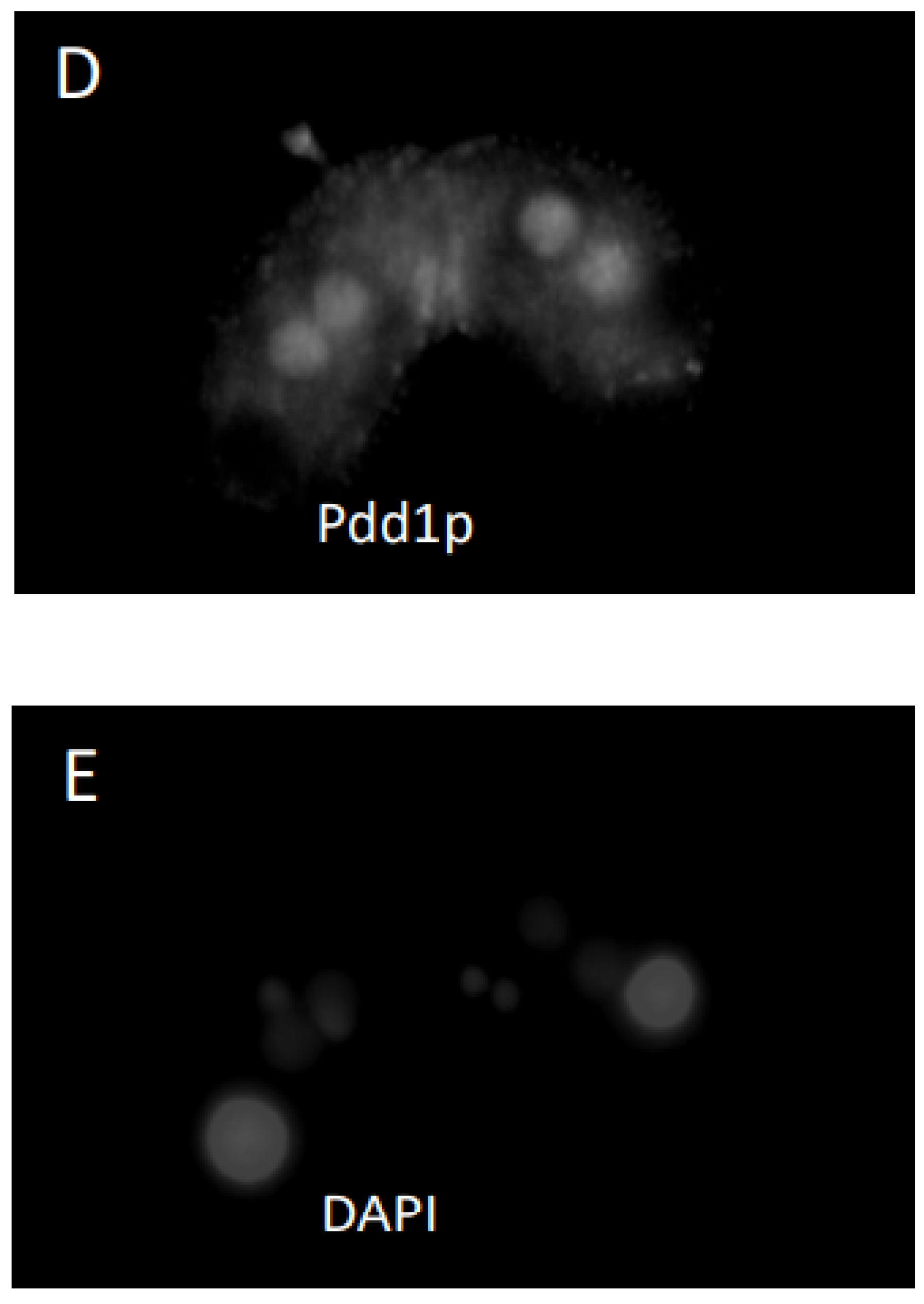
3.2. Response of Cells to Cycloheximide or Aphidicholine Treatment
3.3. Knockouts Have Conjusomes
3.4. Conjusomes Are found in Other Ciliates
4. Discussion
4.1. Conjusomes Appear to Be Regulated by an Independent Pathway
4.2. Pdd1p Is Trafficking between Nuclei
4.3. Conjusomes Might Be the Vehicles of Epigenetic Information
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ng, S.F. The somatic function of the micronucleus of ciliated protozoa. Prog. Protistol. 1986, 1, 215–286. [Google Scholar]
- Prescott, D.M. The DNA of ciliated protozoa. Microbiol. Rev. 1994, 58, 233–267. [Google Scholar] [CrossRef] [PubMed]
- Gorovsky, M.A. Genome Organization and Reorganization in Tetrahymena. Annu. Rev. Genet. 1980, 14, 203–239. [Google Scholar] [CrossRef] [PubMed]
- Chalker, D.L.; Yao, M.C. Non-Mendelian, heritable blocks to DNA rearrangement are induced by loading the somatic nucleus of Tetrahymena thermophila with germ line-limited DNA. Mol. Cell. Biol. 1996, 16, 3658–3667. [Google Scholar] [CrossRef]
- Duharcourt, S.; Keller, A.-M.; Meyer, E. Homology-Dependent Maternal Inhibition of Developmental Excision of Internal Eliminated Sequences in Paramecium tetraurelia. Mol. Cell. Biol. 1998, 18, 7075–7085. [Google Scholar] [CrossRef]
- Hamilton, E.P.; Kapusta, A.; Huvos, P.E.; Bidwell, S.L.; Zafar, N.; Tang, H.; Hadjithomas, M.; Krishnakumar, V.; Badger, J.H.; Caler, E.V.; et al. Structure of the germline genome of Tetrahymena thermophila and relationship to the massively rearranged somatic genome. eLife 2016, 5, e19090. [Google Scholar] [CrossRef]
- Yao, M.-C.; Choi, J.; Yokoyama, S.; Austerberry, C.F.; Yao, C.-H. DNA elimination in Tetrahymena: A developmental process involving extensive breakage and rejoining of DNA at defined sites. Cell 1984, 36, 433–440. [Google Scholar] [CrossRef]
- Yao, M.-C.; Gorovsky, M. Comparison of the sequences of macro- and micronuclear DNA of Tetrahymena pyriformis. Chromosoma 1974, 48, 1–18. [Google Scholar] [CrossRef]
- Chalker, D.L.; Meyer, E.; Mochizuki, K. Epigenetics of Ciliates. Cold Spring Harb. Perspect. Biol. 2013, 5, a017764. [Google Scholar] [CrossRef]
- Nowacki, M.; Landweber, L.F. Epigenetic inheritance in ciliates. Curr. Opin. Microbiol. 2009, 12, 638–643. [Google Scholar] [CrossRef]
- Arnaiz, O.; Mathy, N.; Baudry, C.; Malinsky, S.; Aury, J.-M.; Wilkes, C.D.; Garnier, O.; Labadie, K.; Lauderdale, B.E.; Le Mouël, A.; et al. The Paramecium Germline Genome Provides a Niche for Intragenic Parasitic DNA: Evolutionary Dynamics of Internal Eliminated Sequences. PLoS Genet. 2012, 8, e1002984. [Google Scholar] [CrossRef] [PubMed]
- Noto, T.; Mochizuki, K. Small RNA-Mediated trans-Nuclear and trans-Element Communications in Tetrahymena DNA Elimination. Curr. Biol. 2018, 28, 1938–1949.e5. [Google Scholar] [CrossRef] [PubMed]
- Chalker, D.L. Transgenerational Inheritance: Parental Guidance Suggested. Curr. Biol. 2018, 28, R702–R704. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, K.; Fine, N.A.; Fujisawa, T.; Gorovsky, M.A. Analysis of a piwi-related gene implicates small RNAs in genome rearrangement in Tetrahymena. Cell 2002, 110, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Bouhouche, K.; Gout, J.-F.; Kapusta, A.; Bétermier, M.; Meyer, E. Functional specialization of Piwi proteins in Paramecium tetraurelia from post-transcriptional gene silencing to genome remodelling. Nucleic Acids Res. 2011, 39, 4249–4264. [Google Scholar] [CrossRef]
- Allen, S.E.; Nowacki, M. Roles of Noncoding RNAs in Ciliate Genome Architecture. J. Mol. Biol. 2020, 432, 4186–4198. [Google Scholar] [CrossRef]
- Ouyang, J.P.T.; Folkmann, A.; Bernard, L.; Lee, C.-Y.; Seroussi, U.; Charlesworth, A.G.; Claycomb, J.M.; Seydoux, G. P Granules Protect RNA Interference Genes from Silencing by piRNAs. Dev. Cell 2019, 50, 716–728.e6. [Google Scholar] [CrossRef]
- Janetopoulos, C.; Cole, E.; Smothers, J.; Allis, C.; Aufderheide, K. The conjusome: A novel structure in Tetrahymena found only during sexual reorganization. J. Cell Sci. 1999, 112, 1003–1011. [Google Scholar] [CrossRef]
- Yao, M.-C.; Yao, C.-H.; Halasz, L.M.; Fuller, P.; Rexer, C.H.; Wang, S.H.; Jain, R.; Coyne, R.S.; Chalker, D.L. Identification of novel chromatin-associated proteins involved in programmed genome rearrangements in Tetrahymena. J. Cell Sci. 2007, 120, 1978–1989. [Google Scholar] [CrossRef]
- Rexer, C.H.; Chalker, D.L. Lia1p, a Novel Protein Required during Nuclear Differentiation for Genome-Wide DNA Rearrangements in Tetrahymena thermophila. Eukaryot. Cell 2007, 6, 1320–1329. [Google Scholar] [CrossRef]
- Xu, J.; Yuan, Y.; Liang, A.; Wang, W. Chromodomain protein Tcd1 is required for macronuclear genome rearrangement and repair in Tetrahymena. Sci. Rep. 2015, 5, 10243. [Google Scholar] [CrossRef] [Green Version]
- Chalker, D.L.; Yao, M.-C. Nongenic, bidirectional transcription precedes and may promote developmental DNA deletion in Tetrahymena thermophila. Genes Dev. 2001, 15, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, K.; Gorovsky, M.A. RNA Polymerase II Localizes in Tetrahymena thermophila Meiotic Micronuclei When Micronuclear Transcription Associated with Genome Rearrangement Occurs. Eukaryot. Cell 2004, 3, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Noto, T.; Kataoka, K.; Suhren, J.H.; Hayashi, A.; Woolcock, K.J.; Gorovsky, M.A.; Mochizuki, K. Small-RNA-Mediated Genome-wide trans-Recognition Network in Tetrahymena DNA Elimination. Mol. Cell 2015, 59, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Taverna, S.D.; Coyne, R.S.; Allis, C. Methylation of Histone H3 at Lysine 9 Targets Programmed DNA Elimination in Tetrahymena. Cell 2002, 110, 701–711. [Google Scholar] [CrossRef]
- Liu, Y.; Taverna, S.D.; Muratore, T.L.; Shabanowitz, J.; Hunt, D.F.; Allis, C.D. RNAi-dependent H3K27 methylation is required for heterochromatin formation and DNA elimination in Tetrahymena. Genes Dev. 2007, 21, 1530–1545. [Google Scholar] [CrossRef]
- Suhren, J.H.; Noto, T.; Kataoka, K.; Gao, S.; Liu, Y.; Mochizuki, K. Negative Regulators of an RNAi-Heterochromatin Positive Feedback Loop Safeguard Somatic Genome Integrity in Tetrahymena. Cell Rep. 2017, 18, 2494–2507. [Google Scholar] [CrossRef]
- Kataoka, K.; Mochizuki, K. Phosphorylation of an HP1-like Protein Regulates Heterochromatin Body Assembly for DNA Elimination. Dev. Cell 2015, 35, 775–788. [Google Scholar] [CrossRef]
- Coyne, R.S.; Nikiforov, M.A.; Smothers, J.F.; Allis, C.; Yao, M.-C. Parental Expression of the Chromodomain Protein Pdd1p Is Required for Completion of Programmed DNA Elimination and Nuclear Differentiation. Mol. Cell 1999, 4, 865–872. [Google Scholar] [CrossRef]
- Cole, E.; Bruns, P.J. Uniparental cytogamy: A novel, efficient method for bringing mutations of Tetrahymena into homozygous expression with precocious sexual maturity. Genetics 1992, 132, 1017–1031. [Google Scholar] [CrossRef]
- Dryl, S. Antigenic transformation in Paramecium aurelia after homologous antiserum treatment during conjugation or autogamy. J. Protozool. 1959, 6, 25. [Google Scholar]
- Martindale, D.; Allis, C.; Bruns, P. Conjugation in Tetrahymena thermophila: A temporal analysis of cytological stages. Exp. Cell Res. 1982, 140, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Kaczanowski, A.; Kaczanowska, J. Induction of Blocks in Nuclear Divisions and Overcondensation of Meiotic Chromosomes with Cycloheximide During Conjugation of Tetrahymena thermophila. J. Eukaryot. Microbiol. 1996, 43, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Aufderheide, K.J. Identification of the basal bodies and kinetodesmal fibers in living cells of Paramecium tetraurelia Sonneborn, 1975 and Paramecium sonneborni Aufderheide, Daggett & Nerad, 1983. J. Protozool. 1986, 33, 77–80. [Google Scholar] [PubMed]
- Zinskie, J.A.; Shribak, M.; Bruist, M.F.; Aufderheide, K.J.; Janetopoulos, C. A mechanical microcompressor for high resolution imaging of motile specimens. Exp. Cell Res. 2015, 337, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Wenkert, D.; Allis, C.D. Timing of the appearance of macronuclear-specific histone variant hv1 and gene expression in developing new macronuclei of Tetrahymena thermophila. J. Cell Biol. 1984, 98, 2107–2117. [Google Scholar] [CrossRef]
- Madireddi, M.T.; Davis, M.C.; Allis, C. Identification of a Novel Polypeptide Involved in the Formation of DNA-Containing Vesicles during Macronuclear Development in Tetrahymena. Dev. Biol. 1994, 165, 418–431. [Google Scholar] [CrossRef]
- Cole, E.S.; Soelter, T.A. A mutational analysis of conjugation in Tetrahymena thermophila. 2. Phenotypes affecting middle and late development: Third prezygotic nuclear division, pronuclear exchange, pronuclear fusion and postzygotic development. Dev. Biol. 1997, 189, 233–245. [Google Scholar] [CrossRef]
- Madireddi, M.T.; Coyne, R.S.; Smothers, J.F.; Mickey, K.M.; Yao, M.-C.; Allis, C. Pdd1p, A Novel Chromodomain-Containing Protein, Links Heterochromatin Assembly and DNA Elimination in Tetrahymena. Cell 1996, 87, 75–84. [Google Scholar] [CrossRef]
- Smothers, J.F.; Madireddi, M.T.; Warner, F.D.; Allis, C.D. Programmed DNA Degradation and Nucleolar Biogenesis Occur in Distinct Organelles During Macronuclear Development in Tetrahymena. J. Eukaryot. Microbiol. 1997, 44, 79–88. [Google Scholar] [CrossRef]
- Smothers, J.F.; Mizzen, C.A.; Tubbert, M.M.; Cook, R.G.; Allis, C.D. Pdd1p associates with germline-restricted chromatin and a second novel anlagen-enriched protein in developmentally programmed DNA elimination structures. Development 1997, 124, 4537–4545. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-Y.S.; Putnam, A.; Lu, T.; He, S.; Ouyang, J.P.T.; Seydoux, G. Recruitment of mRNAs to P granules by condensation with intrinsically-disordered proteins. eLife 2020, 9, e52896. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janetopoulos, C.; Aufderheide, K.J. The Conjusome—A Transient Organelle Linking Genome Rearrangements in the Parental and Developing Macronuclei. Microorganisms 2023, 11, 418. https://doi.org/10.3390/microorganisms11020418
Janetopoulos C, Aufderheide KJ. The Conjusome—A Transient Organelle Linking Genome Rearrangements in the Parental and Developing Macronuclei. Microorganisms. 2023; 11(2):418. https://doi.org/10.3390/microorganisms11020418
Chicago/Turabian StyleJanetopoulos, Christopher, and Karl J. Aufderheide. 2023. "The Conjusome—A Transient Organelle Linking Genome Rearrangements in the Parental and Developing Macronuclei" Microorganisms 11, no. 2: 418. https://doi.org/10.3390/microorganisms11020418
APA StyleJanetopoulos, C., & Aufderheide, K. J. (2023). The Conjusome—A Transient Organelle Linking Genome Rearrangements in the Parental and Developing Macronuclei. Microorganisms, 11(2), 418. https://doi.org/10.3390/microorganisms11020418