


A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
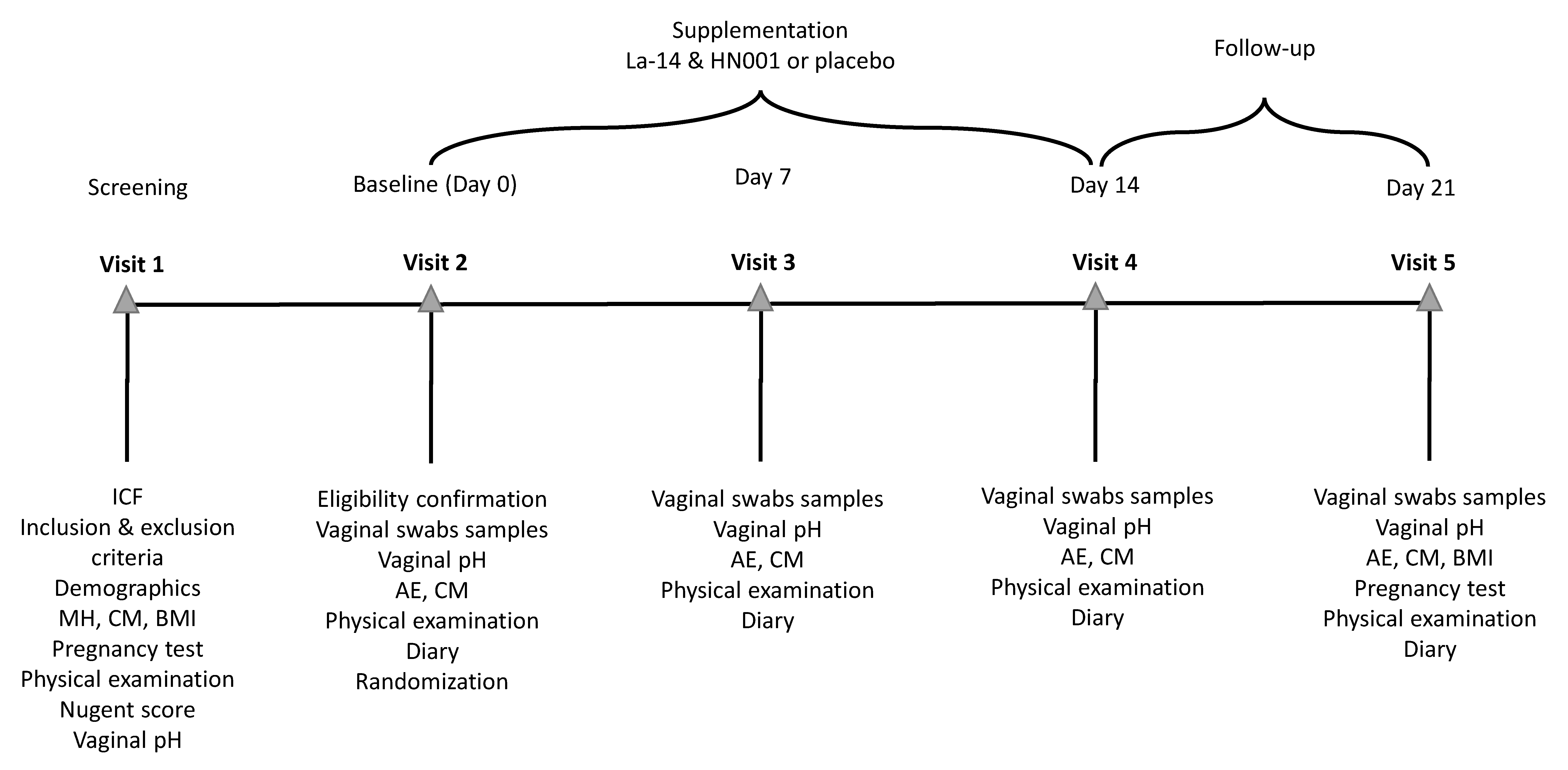
2.1. Study Design
2.2. Participants
2.3. Intervention
2.4. Outcomes
2.5. Sample Collection and Analysis
2.5.1. DNA Extraction
2.5.2. Detection of Colonisation with Quantitative PCR
2.5.3. Vaginal Microbiota Sequencing and Data Analysis
2.5.4. Vaginal Immune Marker Analysis
2.6. Determination of Sample Size
2.7. Randomisation and Blinding
2.8. Statistical Analysis
2.8.1. Primary and Secondary Outcomes
2.8.2. Exploratory Outcomes
3. Results
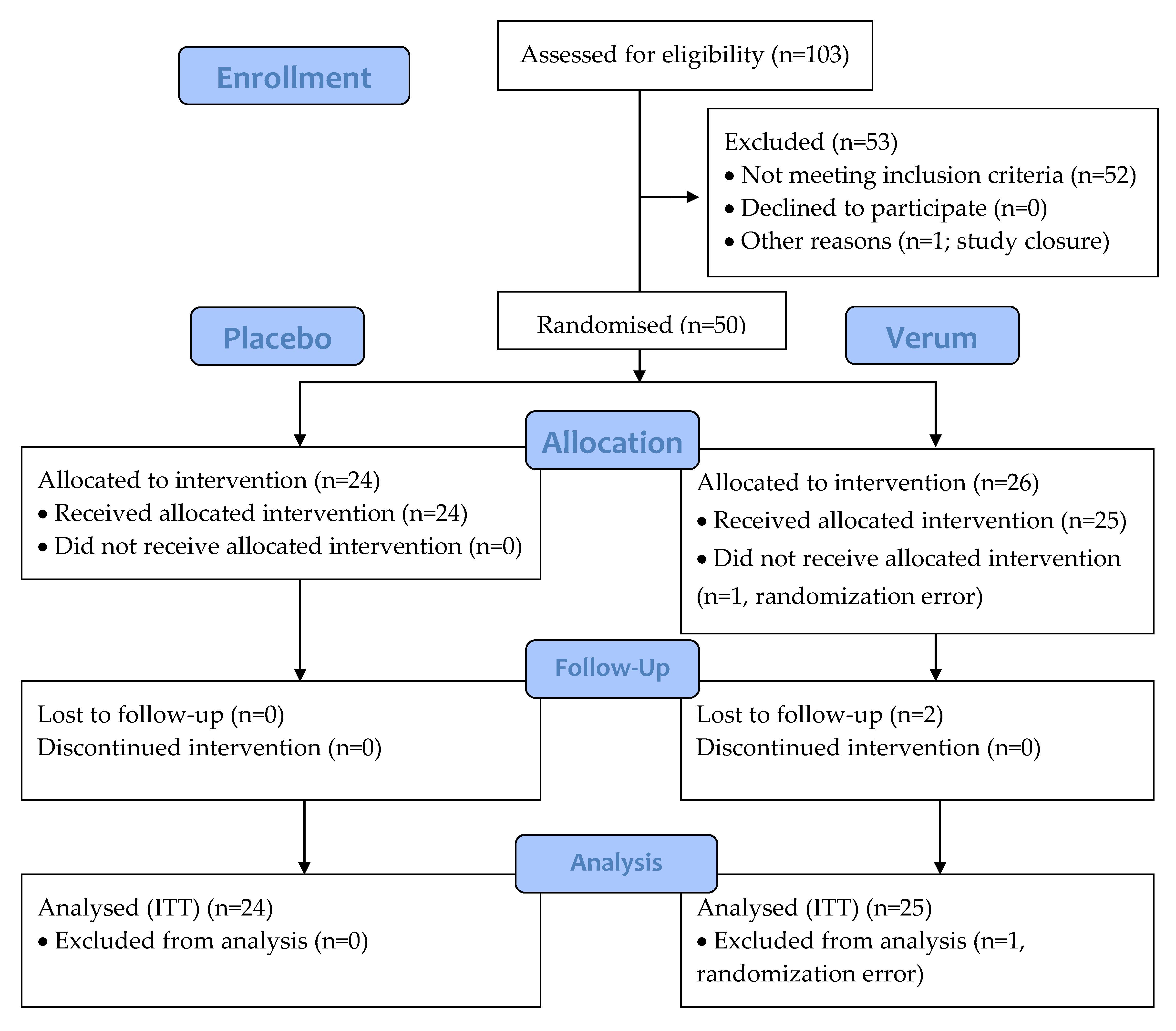
3.1. Participants
3.2. Vaginal Colonisation and pH
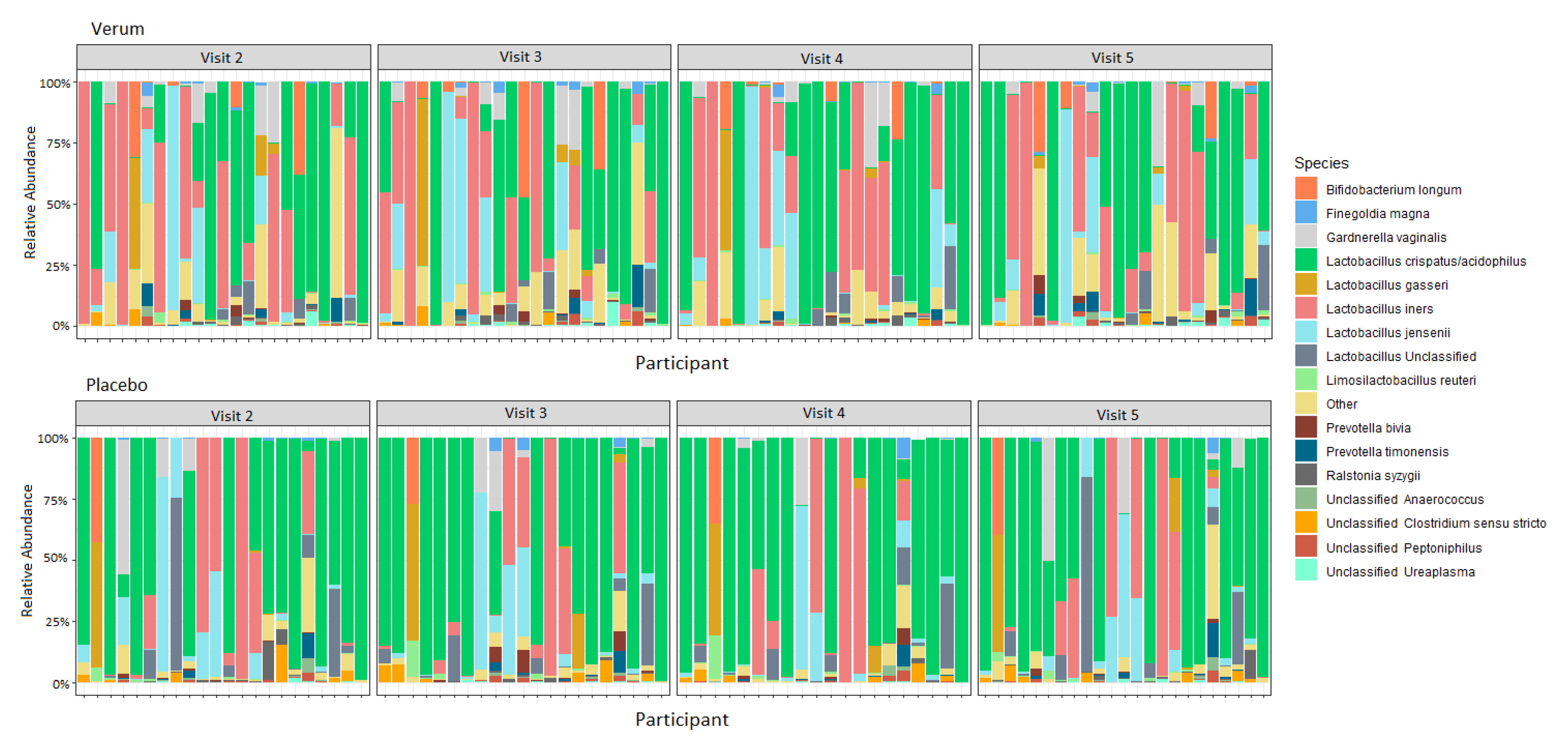
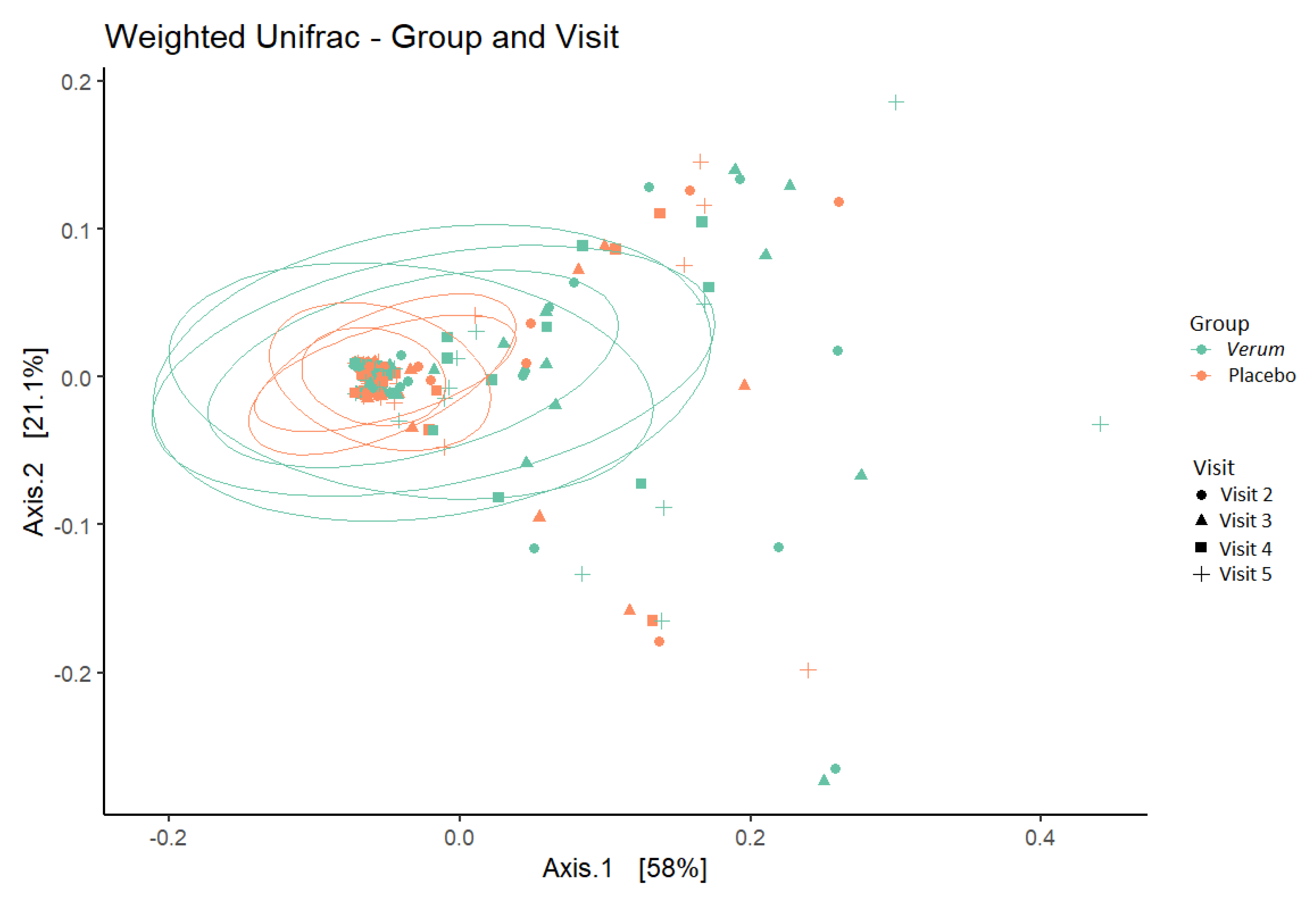
3.3. Vaginal Microbiota
3.4. Vaginal Immune Markers
3.5. Safety
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, X.; Brown, C.J.; Abdo, Z.; Davis, C.C.; Hansmann, M.A.; Joyce, P.; Foster, J.A.; Forney, L.J. Differences in the composition of vaginal microbial communities found in healthy Caucasian and black women. ISME J. 2007, 1, 121–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.B.; Ravel, J. The vaginal microbiota, host defence and reproductive physiology. J. Physiol. 2017, 595, 451–463. [Google Scholar] [CrossRef] [Green Version]
- France, M.T.; Ma, B.; Gajer, P.; Brown, S.; Humphrys, M.S.; Holm, J.B.; Waetjen, L.E.; Brotman, R.M.; Ravel, J. VALENCIA: A nearest centroid classification method for vaginal microbial communities based on composition. Microbiome 2020, 8, 166. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Bhattacharjee, M.J.; Mukherjee, A.K.; Khan, M.R. Recent advances in understanding of multifaceted changes in the vaginal microenvironment: Implications in vaginal health and therapeutics. Crit. Rev. Microbiol. 2022, 1–27, online ahead of print. [Google Scholar] [CrossRef]
- Lehtoranta, L.; Ala-Jaakkola, R.; Laitila, A.; Maukonen, J. Healthy Vaginal Microbiota and Influence of Probiotics across the Female Life Span. Front. Microbiol. 2022, 13, 819958. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtoranta, L.; Hibberd, A.A.; Reimari, J.; Junnila, J.; Yeung, N.; Maukonen, J.; Crawford, G.; Ouwehand, A.C. Recovery of Vaginal Microbiota After Standard Treatment for Bacterial Vaginosis Infection: An Observational Study. Microorganisms 2020, 8, 875. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Guo, R.; Wang, J.; Zhou, W.; Ling, Z. Contribution of Lactobacillus iners to Vaginal Health and Diseases: A Systematic Review. Front. Cell Infect. Microbiol. 2021, 11, 792787. [Google Scholar] [CrossRef]
- Vitali, B.; Cruciani, F.; Picone, G.; Parolin, C.; Donders, G.; Laghi, L. Vaginal microbiome and metabolome highlight specific signatures of bacterial vaginosis. Eur. J. Clin.. Microbiol. Infect. Dis. 2015, 34, 2367–2376. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, S.; Kwon, M.Y.; Kim, M. Clinical Significance of Composition and Functional Diversity of the Vaginal Microbiome in Recurrent Vaginitis. Front. Microbiol. 2022, 13, 851670. [Google Scholar] [CrossRef]
- Nguyen, P.V.; Kafka, J.K.; Ferreira, V.H.; Roth, K.; Kaushic, C. Innate and adaptive immune responses in male and female reproductive tracts in homeostasis and following HIV infection. Cell Mol. Immunol. 2014, 11, 410–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anahtar, M.N.; Byrne, E.H.; Doherty, K.E.; Bowman, B.A.; Yamamoto, H.S.; Soumillon, M.; Padavattan, N.; Ismail, N.; Moodley, A.; Sabatini, M.E.; et al. Cervicovaginal bacteria are a major modulator of host inflammatory responses in the female genital tract. Immunity 2015, 42, 965–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wira, C.R.; Rodriguez-Garcia, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Quayle, A.J. The innate and early immune response to pathogen challenge in the female genital tract and the pivotal role of epithelial cells. J. Reprod. Immunol. 2002, 57, 61–79. [Google Scholar] [CrossRef]
- Klotman, M.E.; Chang, T.L. Defensins in innate antiviral immunity. Nat. Rev. Immunol. 2006, 6, 447–456. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; Kelly, R.W.; Sallenave, J.M.; Bocking, A.D.; Challis, J.R. Innate immune defences in the human uterus during pregnancy. Placenta 2007, 28, 1099–1106. [Google Scholar] [CrossRef]
- Stock, S.J.; Duthie, L.; Tremaine, T.; Calder, A.A.; Kelly, R.W.; Riley, S.C. Elafin (SKALP/Trappin-2/proteinase inhibitor-3) is produced by the cervix in pregnancy and cervicovaginal levels are diminished in bacterial vaginosis. Reprod. Sci. 2009, 16, 1125–1134. [Google Scholar] [CrossRef]
- Tramont, E.C. Inhibition of adherence of Neisseria gonorrhoeae by human genital secretions. J. Clin. Invest. 1977, 59, 117–124. [Google Scholar] [CrossRef]
- Breedveld, A.C.; Schuster, H.J.; van Houdt, R.; Painter, R.C.; Mebius, R.E.; van der Veer, C.; Bruisten, S.M.; Savelkoul, P.H.M.; van Egmond, M. Enhanced IgA coating of bacteria in women with Lactobacillus crispatus-dominated vaginal microbiota. Microbiome 2022, 10, 15. [Google Scholar] [CrossRef]
- Chen, R.; Li, R.; Qing, W.; Zhang, Y.; Zhou, Z.; Hou, Y.; Shi, Y.; Zhou, H.; Chen, M. Probiotics are a good choice for the treatment of bacterial vaginosis: A meta-analysis of randomized controlled trial. Reprod. Health 2022, 19, 137. [Google Scholar] [CrossRef]
- Liu, H.F.; Yi, N. A systematic review and meta-analysis on the efficacy of probiotics for bacterial vaginosis. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, R.; Edu, A.; De Seta, F. Study on the effects of an oral lactobacilli and lactoferrin complex in women with intermediate vaginal microbiota. Arch. Gynecol. Obstet. 2018, 298, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Karadja, E.; De Seta, F. Evidence-based mixture containing Lactobacillus strains and lactoferrin to prevent recurrent bacterial vaginosis: A double blind, placebo controlled, randomised clinical trial. Benef. Microbes 2019, 10, 19–26. [Google Scholar] [CrossRef]
- Anukam, K.C.; Osazuwa, E.; Osemene, G.I.; Ehigiagbe, F.; Bruce, A.W.; Reid, G. Clinical study comparing probiotic Lactobacillus GR-1 and RC-14 with metronidazole vaginal gel to treat symptomatic bacterial vaginosis. Microbes Infect. 2006, 8, 2772–2776. [Google Scholar] [CrossRef]
- Heczko, P.B.; Tomusiak, A.; Adamski, P.; Jakimiuk, A.J.; Stefanski, G.; Mikolajczyk-Cichonska, A.; Suda-Szczurek, M.; Strus, M. Supplementation of standard antibiotic therapy with oral probiotics for bacterial vaginosis and aerobic vaginitis: A randomised, double-blind, placebo-controlled trial. BMC Womens Health 2015, 15, 115. [Google Scholar] [CrossRef] [Green Version]
- Koirala, R.; Gargari, G.; Arioli, S.; Taverniti, V.; Fiore, W.; Grossi, E.; Anelli, G.M.; Cetin, I.; Guglielmetti, S. Effect of oral consumption of capsules containing Lactobacillus paracasei LPC-S01 on the vaginal microbiota of healthy adult women: A randomized, placebo-controlled, double-blind crossover study. FEMS Microbiol. Ecol. 2020, 96, fiaa084. [Google Scholar] [CrossRef]
- De Leo, V.; Lazzeri, E.; Governini, L.; Cuppone, A.M.; Colombini, L.; Teodori, L.; Ciprandi, G.; Iannelli, F.; Pozzi, G. Vaginal colonization of women after oral administration of Lactobacillus crispatus strain NTCVAG04 from the human microbiota. Minerva Obstet. Gynecol. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Bohbot, J.M.; Cardot, J.M. Vaginal impact of the oral administration of total freeze-dried culture of LCR 35 in healthy women. Infect. Dis. Obstet. Gynecol. 2012, 2012, 503648. [Google Scholar] [CrossRef] [Green Version]
- De Alberti, D.; Russo, R.; Terruzzi, F.; Nobile, V.; Ouwehand, A.C. Lactobacilli vaginal colonisation after oral consumption of Respecta((R)) complex: A randomised controlled pilot study. Arch. Gynecol. Obstet. 2015, 292, 861–867. [Google Scholar] [CrossRef]
- Shalev, E.; Battino, S.; Weiner, E.; Colodner, R.; Keness, Y. Ingestion of yogurt containing Lactobacillus acidophilus compared with pasteurized yogurt as prophylaxis for recurrent candidal vaginitis and bacterial vaginosis. Arch. Fam. Med. 1996, 5, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Strus, M.; Chmielarczyk, A.; Kochan, P.; Adamski, P.; Chelmicki, Z.; Chelmicki, A.; Palucha, A.; Heczko, P.B. Studies on the effects of probiotic Lactobacillus mixture given orally on vaginal and rectal colonization and on parameters of vaginal health in women with intermediate vaginal flora. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 163, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Hao, L.; Zhang, Z.; Tian, L.; Zhang, X.; Zhu, J.; Jie, Z.; Tong, X.; Xiao, L.; Zhang, T.; et al. Cervicovaginal microbiome dynamics after taking oral probiotics. J. Genet. Genom. 2021, 48, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Superti, F.; Karadja, E.; De Seta, F. Randomised clinical trial in women with Recurrent Vulvovaginal Candidiasis: Efficacy of probiotics and lactoferrin as maintenance treatment. Mycoses 2019, 62, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Ku, Y.W.; Chu, F.Y. Influence of bovine lactoferrin on the growth of selected probiotic bacteria under aerobic conditions. Biometals 2014, 27, 905–914. [Google Scholar] [CrossRef]
- Pino, A.; Giunta, G.; Randazzo, C.L.; Caruso, S.; Caggia, C.; Cianci, A. Bacterial biota of women with bacterial vaginosis treated with lactoferrin: An open prospective randomized trial. Microb. Ecol. Health Dis. 2017, 28, 1357417. [Google Scholar] [CrossRef] [Green Version]
- Giunta, G.; Giuffrida, L.; Mangano, K.; Fagone, P.; Cianci, A. Influence of lactoferrin in preventing preterm delivery: A pilot study. Mol. Med. Rep. 2012, 5, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.E.; Jeong, J.J.; Choi, S.Y.; Kim, H.; Han, M.J.; Kim, D.H. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 Attenuate Gardnerella vaginalis-Infected Bacterial Vaginosis in Mice. Nutrients 2017, 9, 531. [Google Scholar] [CrossRef] [Green Version]
- Bertuccini, L.; Russo, R.; Iosi, F.; Superti, F. Effects of Lactobacillus rhamnosus and Lactobacillus acidophilus on bacterial vaginal pathogens. Int. J. Immunopathol. Pharmacol. 2017, 30, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.; Morgan, X.C.; Wang, X.Y.; Wickens, K.; Purdie, G.; Fitzharris, P.; Otal, A.; Lawley, B.; Stanley, T.; Barthow, C.; et al. Eczema-protective probiotic alters infant gut microbiome functional capacity but not composition: Sub-sample analysis from a RCT. Benef. Microbes 2019, 10, 5–17. [Google Scholar] [CrossRef]
- Taverniti, V.; Koirala, R.; Dalla Via, A.; Gargari, G.; Leonardis, E.; Arioli, S.; Guglielmetti, S. Effect of Cell Concentration on the Persistence in the Human Intestine of Four Probiotic Strains Administered through a Multispecies Formulation. Nutrients 2019, 11, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogensen, G.S.; O’Brien, J.; Ouwenhand, A.; Holzapfel, W.; Shortt, C.; Fonden, R.; Miller, G.D.; Donohue, D.; Playne, M.J.; Crittenden, R.G.; et al. Inventory of Microorganisms with a documented history of use in food. Bull. Int. Dairy Fed. 2002, 377, 10–19. [Google Scholar]
- Bourdichon, F.; Casaregola, S.; Farrokh, C.; Frisvad, J.C.; Gerds, M.L.; Hammes, W.P.; Harnett, J.; Huys, G.; Laulund, S.; Ouwehand, A.; et al. Food fermentations: Microorganisms with technological beneficial use. Int. J. Food Microbiol. 2012, 154, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yang, S.M.; Lim, B.; Park, S.H.; Rackerby, B.; Kim, H.Y. Design of PCR assays to specifically detect and identify 37 Lactobacillus species in a single 96 well plate. BMC Microbiol. 2020, 20, 96. [Google Scholar] [CrossRef] [Green Version]
- Byun, R.; Nadkarni, M.A.; Chhour, K.L.; Martin, F.E.; Jacques, N.A.; Hunter, N. Quantitative analysis of diverse Lactobacillus species present in advanced dental caries. J. Clin. Microbiol. 2004, 42, 3128–3136. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.J.Z.; Tang, P.; Kiefer, A.; Galles, K.; Wong, C.; Morovic, W. Droplet Digital PCR Is an Improved Alternative Method for High-Quality Enumeration of Viable Probiotic Strains. Front. Microbiol. 2019, 10, 3025. [Google Scholar] [CrossRef]
- Song, Y.; Kato, N.; Liu, C.; Matsumiya, Y.; Kato, H.; Watanabe, K. Rapid identification of 11 human intestinal Lactobacillus species by multiplex PCR assays using group- and species-specific primers derived from the 16S-23S rRNA intergenic spacer region and its flanking 23S rRNA. FEMS Microbiol. Lett. 2000, 187, 167–173. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Author Correction: Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 1091. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Faith, D.P. Systematics and Conservation: On Predicting the Feature Diversity of Subsets of Taxa. Cladistics 1992, 8, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The R Project for Statistical Computing. Available online: http://www.R-project.org (accessed on 20 April 2018).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis. Available online: http://CRAN.R-project.org/package=ggplot2 (accessed on 20 April 2018).
- Virtanen, S.; Kalliala, I.; Nieminen, P.; Salonen, A. Comparative analysis of vaginal microbiota sampling using 16S rRNA gene analysis. PLoS ONE 2017, 12, e0181477. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbo, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [Green Version]
- Onderdonk, A.B.; Delaney, M.L.; Fichorova, R.N. The Human Microbiome during Bacterial Vaginosis. Clin. Microbiol. Rev. 2016, 29, 223–238. [Google Scholar] [CrossRef] [Green Version]
- van de Wijgert, J.; Jespers, V. The global health impact of vaginal dysbiosis. Res. Microbiol. 2017, 168, 859–864. [Google Scholar] [CrossRef]
- Falconi-McCahill, A. Bacterial Vaginosis: A Clinical Update with a Focus on Complementary and Alternative Therapies. J. Midwifery Womens Health 2019, 64, 578–591. [Google Scholar] [CrossRef]
- Boetius Hertz, F.; Holm, J.B.; Palleja, A.; Bjornsdottir, M.K.; Mikkelsen, L.S.; Brandsborg, E.; Frimodt-Moller, N. The vaginal microbiome following orally administered probiotic. APMIS 2022, 130, 605–611. [Google Scholar] [CrossRef]
- Allen, N.G.; Edupuganti, L.; Edwards, D.J.; Jimenez, N.R.; Buck, G.A.; Jefferson, K.K.; Strauss, J.F., 3rd; Vaginal Microbiome, C.; Wickham, E.P., 3rd; Fettweis, J.M. The vaginal microbiome in women of reproductive age with healthy weight versus overweight/obesity. Obesity 2022, 30, 142–152. [Google Scholar] [CrossRef]
- Ahannach, S.; Delanghe, L.; Spacova, I.; Wittouck, S.; Van Beeck, W.; De Boeck, I.; Lebeer, S. Microbial enrichment and storage for metagenomics of vaginal, skin, and saliva samples. iScience 2021, 24, 103306. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Marques, J.; Hout, A.; Ferreira, R.M.; Weber, M.; Pinto-Ribeiro, I.; van Doorn, L.J.; Knetsch, C.W.; Figueiredo, C. Impact of Host DNA and Sequencing Depth on the Taxonomic Resolution of Whole Metagenome Sequencing for Microbiome Analysis. Front. Microbiol. 2019, 10, 1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattei, V.; Murugesan, S.; Al Hashmi, M.; Mathew, R.; James, N.; Singh, P.; Kumar, M.; Lakshmanan, A.P.; Terranegra, A.; Al Khodor, S.; et al. Evaluation of Methods for the Extraction of Microbial DNA From Vaginal Swabs Used for Microbiome Studies. Front. Cell. Infect. Microbiol. 2019, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Antonio, M.A.; Meyn, L.A.; Murray, P.J.; Busse, B.; Hillier, S.L. Vaginal colonization by probiotic Lactobacillus crispatus CTV-05 is decreased by sexual activity and endogenous Lactobacilli. J. Infect. Dis. 2009, 199, 1506–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcotte, H.; Larsson, P.G.; Andersen, K.K.; Zuo, F.; Mikkelsen, L.S.; Brandsborg, E.; Gray, G.; Laher, F.; Otwombe, K. An exploratory pilot study evaluating the supplementation of standard antibiotic therapy with probiotic lactobacilli in south African women with bacterial vaginosis. BMC Infect. Dis. 2019, 19, 824. [Google Scholar] [CrossRef]
- Brandt, K.; Barrangou, R. Adaptive response to iterative passages of five Lactobacillus species in simulated vaginal fluid. BMC Microbiol. 2020, 20, 339. [Google Scholar] [CrossRef]
- Finlay, B.B.; Amato, K.R.; Azad, M.; Blaser, M.J.; Bosch, T.C.G.; Chu, H.; Dominguez-Bello, M.G.; Ehrlich, S.D.; Elinav, E.; Geva-Zatorsky, N.; et al. The hygiene hypothesis, the COVID pandemic, and consequences for the human microbiome. Proc. Natl. Acad. Sci. USA 2021, 118, e2010217118. [Google Scholar] [CrossRef]
- Miquel, S.; Verlaguet, J.; Garcin, S.; Bertran, T.; Evrard, B.; Forestier, C.; Vareille-Delarbre, M. Lacticaseibacillus rhamnosus Lcr35 Stimulates Epithelial Vaginal Defenses upon Gardnerella vaginalis Infection. Infect. Immun. 2022, 90, e0030922. [Google Scholar] [CrossRef]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Ghosh, S.K.; Flyckt, R.; Kalinowska, M.; Starks, D.; Jurevic, R.; Weinberg, A.; Lederman, M.M.; Rodriguez, B. Bacterial colonization and beta defensins in the female genital tract in HIV infection. Curr. HIV Res. 2012, 10, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Orfanelli, T.; Jayaram, A.; Doulaveris, G.; Forney, L.J.; Ledger, W.J.; Witkin, S.S. Human epididymis protein 4 and secretory leukocyte protease inhibitor in vaginal fluid: Relation to vaginal components and bacterial composition. Reprod. Sci. 2014, 21, 538–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
|
|
| Primer/Probe (5′–3′) | Master Mix | Annealing Temperature |
|---|---|---|
| Primary outcome assays | ||
| L. acidophilus (species level) [44] | 1 × SYBR FAST | 60 °C |
| F: CCTTTCTAAGGAAGCGAAGGAT (400 nM) | ||
| R: ACGCTTGGTATTCCAAATCG (400 nM) | ||
| L. rhamnosus (species level) [45] | 1 × SYBR FAST | 62 °C |
| F: TGCTTGCATCTTGATTTAATTTTG (400 nM) | ||
| R: GGTTCTTGGATYTATGCGGTATTAG (400 nM) | ||
| Exploratory outcome assays | ||
| L. acidophilus La-14 (strain level) [46] | 1 × Taq FAST Advanced | 60 °C |
| F: CCGGTTAATAAAATCTTTTCACCTTG (600 nM) | ||
| R: GCAGTTATTAATCGTGATTTGCATATAAATT (600 nM) | ||
| P: FAM-AGTTGATCAGTCAGCAAGTAGTGTTATGG-IowaBlack (300 nM) | ||
| L. rhamnosus HN001 (strain level) | 1 × Taq FAST Advanced | 60 °C |
| F: CTGGAGGAGATCACAACGACT (400 nM) | ||
| R: ATTGTCCCAACGCTGAATGC (400 nM) | ||
| P: FAM-TGAAGACAAGGTTGCGCCCTGTACACTGTTA-IowaBlack (200 nM) | ||
| Post hoc analysis | ||
| L. acidophilus (species level) [47] | 1 × SYBR FAST | 62 °C |
| F: TGCAAAGTGGTAGCGTAAGC (400 nM) | ||
| R: CCTTTCCCTCACGGTACTG (400 nM) |
| Item | Statistic | Placebo (n = 24) | Verum (n = 25) |
|---|---|---|---|
| Age (years) | Mean (SD) | 30.9 (6.89) | 32.4 (7.94) |
| Median | 30.0 | 33.0 | |
| Min, Max | 21, 41 | 20,47 | |
| BMI (kg/m2) | Mean (SD) | 26.9 (6.94) | 26.5 (6.41) |
| Median | 25.3 | 23.8 | |
| Min, Max | 16.8, 41.5 | 19.9, 43.6 | |
| Smoking status | |||
| Every day smoker | n (%) | 0 (0.0) | 2 (8.0) |
| Former smoker | n (%) | 7 (29.2) | 3 (12.0) |
| Never smoker | n (%) | 16 (66.7) | 16 (64.0) |
| Someday smoker | n (%) | 1 (4.2) | 4 (16.0) |
| Alcohol use | |||
| No | n (%) | 4 (16.7) | 4 (16.0) |
| Yes | n (%) | 20 (83.3) | 21 (84.0) |
| Contraceptive method | |||
| Hormonal | n (%) | 14 (58.3) | 9 (36.0) |
| Not hormonal | n (%) | 10 (41.7) | 16 (64.0) |
| Nugent score | |||
| 0 | n (%) | 19 (79.2) | 16 (64.0) |
| 1 | n (%) | 3 (12.5) | 3 (12.0) |
| 2 | n (%) | 2 (8.3) | 2 (8.0) |
| 3 | n (%) | 0 (0.0) | 3 (12.0) |
| Item | Statistic | Placebo (n = 24) | Verum (n = 25) |
|---|---|---|---|
| Duration of menstruation (days) | n | 18 | 19 |
| Mean (SD) | 4.83 (1.15) | 4.53 (1.07) | |
| Median | 5.00 | 5.00 | |
| Min, Max | 3.00, 7.00 | 2.00, 7.00 | |
| Length of menstrual cycle (days) | n | 18 | 19 |
| Mean (SD) | 27.61 | 28.47 | |
| Median | 28.00 | 28.00 | |
| Min, Max | 24.00, 29.00 | 25.00, 35.00 | |
| Menarche age (years) | n | 24 | 25 |
| Mean (SD) | 12.63 (1.53) | 13.04 (1.59) | |
| Median | 12.50 | 13.00 | |
| Min, Max | 10.00, 16.00 | 10.00, 16.00 | |
| n (%) | n (%) | ||
| Intermenstrual bleeding | N/A | 5 (20.8) | 5 (20.0) |
| No | 19 (79.2) | 20 (80.0) | |
| Menstrual flow | Mild | 6 (25.0) | 5 (20.0) |
| Moderate | 12 (50.0) | 14 (56.0) | |
| Post-coital bleeding | No | 24 (100.0) | 25 (100.0) |
| Leucorrhea | No | 23 (95.8) | 25 (100.0) |
| Yes | 1 (4.2) | 0 (0.0) | |
| Number of pregnancies | 0 | 14 (58.3) | 12 (48.0) |
| 1 | 4 (16.7) | 3 (12.0) | |
| 2 | 5 (20.8) | 7 (28.0) | |
| 3 | 1 (4.2) | 3 (12.0) | |
| Operations | No | 22 (91.7) | 23 (92.0) |
| Yes | 2 (8.3) | 2 (8.0) | |
| Pain in the lower abdomen | No | 24 (100.0) | 25 (100.0) |
| Pelvic or abdominal pelvic pain | No | 24 (100.0) | 24 (96.0) |
| Yes | 0 (0.0) | 1 (4.0) | |
| Presence of vaginal inflammations, dryness, itchiness, stinging | No | 24 (100.0) | 25 (100.0) |
| Sterility | No | 24 (100.0) | 25 (100.0) |
| Vulvovaginal infections | No | 24 (100.0) | 25 (100.0) |
| Assay | Visit | Placebo (n = 20–23) | Verum (n = 23–25) |
|---|---|---|---|
| Genomes detected | log 10 genomes/swab | log 10 genomes/swab | |
| L. acidophilus | V2 | <5.29 (0.00) | <5.29 (0.00) |
| V3 | <5.29 (0.00) | <5.29 (0.00) | |
| V4 | <5.29 (0.00) | <5.29 (0.00) | |
| V5 | <5.29 (0.00) | <5.29 (0.00) | |
| L. rhamnosus | V2 | 5.12 (0.056) | <5.11 (0.000) |
| V3 | <5.11 (0.017) | <5.11 (0.000) | |
| V4 | <5.11 (0.009) | <5.11 (0.000) | |
| V5 | 5.12 (0.048) | <5.11 (0.000) |
| Group | % (n/n) of Participants with > 2-Fold Increase from Baseline in Genome Copies/ng | % (n/n) of Participants with > 2-Fold Increase from Baseline in Genome Copies/swab | ||
|---|---|---|---|---|
| V3 | V4 | V3 | V4 | |
| Verum | 36 (9/25) | 40 (10/25) | 40 (10/25) | 36 (9/25) |
| Placebo | 30 (7/23) | 15 (3/20) | 26 (6/23) | 30 (6/20) |
| Visit | Categorised Change in Vaginal pH from Baseline | Placebo (n = 22–24) | Verum (n = 23–25) |
|---|---|---|---|
| n (%) | n (%) | ||
| V3 | Decrease | 5 (20.8) | 5 (20.0) |
| No change | 13 (54.2) | 16 (64.0) | |
| Increase | 6 (25.0) | 4 (16.0) | |
| V4 | Decrease | 4 (16.7) | 8 (32.0) |
| No change | 14 (58.3) | 14 (56.0) | |
| Increase | 4 (16.7) | 3 (12.0) | |
| V5 | Decrease | 2 (8.3) | 10 (40.0) |
| No change | 16 (66.7) | 8 (32.0) | |
| Increase | 5 (20.8) | 5 (20.0) |
| Group | Visit | Samples | CST I | CST II | CST III | CST IV-B | CST IV-A | CST V |
|---|---|---|---|---|---|---|---|---|
| n | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | ||
| Verum | All visits | 90 | 38 (42.2) | 3 (3.3) | 26 (28.9) | 3 (3.3) | 5 (5.6) | 15 (16.7) |
| V2 | 23 | 9 (39.1) | 1 (4.3) | 8 (34.8) | 1 (4.3) | 1 (4.3) | 3 (13.0) | |
| V3 | 23 | 8 (34.8) | 1 (4.3) | 5 (21.7) | 1 (4.3) | 3 (13.0) | 5 (21.7) | |
| V4 | 22 | 10 (45.5) | 1 (4.5) | 7 (31.8) | 0 (0.0) | 0 (0.0) | 4 (18.2) | |
| V5 | 22 | 11 (50.0) | 0 (0.0) | 6 (27.3) | 1 (4.5) | 1 (4.5) | 3 (13.6) | |
| Placebo | All visits | 86 | 56 (65.1) | 4 (4.7) | 14 (16.3) | 1 (1.2) | 2 (2.3) | 9 (10.5) |
| V2 | 22 | 13 (59.1) | 1 (4.5) | 4 (18.2) | 1 (4.5) | 0 (0.0) | 3 (13.6) | |
| V3 | 21 | 14 (66.7) | 1 (4.8) | 3 (14.3) | 0 (0.0) | 0 (0.0) | 3 (14.3) | |
| V4 | 20 | 14 (70.0) | 1 (5.0) | 3 (15.0) | 0 (0.0) | 1 (5.0) | 1 (5.0) | |
| V5 | 23 | 15 (65.2) | 1 (4.3) | 4 (17.4) | 0 (0.0) | 1 (4.3) | 2 (8.7) |
| Group | Visit | N | Gardnerella vaginalis | Lactobacillus spp. | Lactobacillus crispatus | Lactobacillus jensenii | Lactobacillus gasseri | Lactobacillus iners |
|---|---|---|---|---|---|---|---|---|
| Percentage of participants with >1% increase in the relative abundance of an ASV. n (%) | ||||||||
| Verum | V3 | 23 | 4 (17) | 3 (13) | 5 (22) | 7 (30) | 3 (13) | 4 (17) |
| V4 | 22 | 3 (14) | 4 (18) | 7 (32) | 7 (32) | 2 (9) | 6 (27) | |
| V5 | 22 | 4 (18) | 2 (9) | 8 (36) | 5 (23) | 0 (0) | 7 (32) | |
| Placebo | V3 | 20 | 4 (20) | 3 (15) | 5 (25) | 4 (20) | 3 (15) | 3 (15) |
| V4 | 19 | 2 (11) | 4 (21) | 8 (42) | 2 (11) | 2 (11) | 4 (21) | |
| V5 | 22 | 4 (18) | 3 (14) | 7 (32) | 5 (23) | 2 (9) | 2 (9) | |
| Percentage of participants with >1% decrease in the relative abundance of an ASV. n (%) | ||||||||
| Verum | V3 | 23 | 4 (17) | 2 (9) | 6 (26) | 1 (4) | 1 (4) | 9 (39) |
| V4 | 22 | 5 (23) | 1 (5) | 5 (23) | 2 (9) | 0 (0) | 7 (32) | |
| V5 | 22 | 5 (23) | 1 (5) | 3 (14) | 4 (18) | 3 (14) | 7 (32) | |
| Placebo | V3 | 20 | 0 (0) | 2 (10) | 6 (30) | 4 (20) | 0 (0) | 4 (20) |
| V4 | 19 | 1 (5) | 1 (5) | 5 (26) | 5 (26) | 1 (5) | 3 (16) | |
| V5 | 22 | 2 (9) | 3 (14) | 7 (32) | 6 (27) | 1 (5) | 5 (23) | |
| Analysis Method | Parameter | Visit | Verum vs. Placebo | p |
|---|---|---|---|---|
| RM-ANCOVA (log-transformed) | SLPI (p = 0.15) | V3 | −0.194 (−0.793, 0.406) | 0.52 |
| V4 | −0.148 (−0.657, 0.362) | 0.56 | ||
| V5 | −0.488 (−0.988, 0.013) | 0.056 | ||
| Elafin (p = 0.022) | V3 | −0.310 (−0.648, 0.028) | 0.071 | |
| V4 | −0.305 (−0.626, 0.015) | 0.062 | ||
| V5 | −0.318 (−0.640, 0.004) | 0.051 | ||
| IgA (p = 0.82) | V3 | 0.038 (−0.401, 0.477) | 0.86 | |
| V4 | −0.079 (−0.577, 0.420) | 0.75 | ||
| V5 | −0.083 (−0.625, 0.460) | 0.76 | ||
| RM-ANCOVA (square-root-transformed) | HBD-1 (p = 0.97) | V3 | −1.48 (−11.78, 8.83) | 0.77 |
| V4 | 2.50 (−7.42, 12.43) | 0.61 | ||
| V5 | −1.51 (−9.40, 6.38) | 0.70 | ||
| HBD-3 (p = 0.028) | V3 | −6.34 (−24.72, 12.05) | 0.49 | |
| V4 | −13.86 (−31.69, 3.97) | 0.12 | ||
| V5 | −22.70 (−39.03, −6.37) | 0.008 |
| Finegoldia magna | Gardnerella vaginalis | Lactobacillus crispatus/acidophilus | Lactobacillus jensenii | Prevotella bivia | |
|---|---|---|---|---|---|
| Average R-Value (p) | |||||
| HBD-1 | −0.173 (0.05) | NS | NS | NS | −0.231 (0.01) |
| HBD-3 | −0.2 (0.02) | −0.197 (0.03) | NS | −0.193 (0.03) | NS |
| IgA | NS | NS | −0.258 (0.004) | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyra, A.; Ala-Jaakkola, R.; Yeung, N.; Datta, N.; Evans, K.; Hibberd, A.; Lehtinen, M.J.; Forssten, S.D.; Ibarra, A.; Pesonen, T.; et al. A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial. Microorganisms 2023, 11, 499. https://doi.org/10.3390/microorganisms11020499
Lyra A, Ala-Jaakkola R, Yeung N, Datta N, Evans K, Hibberd A, Lehtinen MJ, Forssten SD, Ibarra A, Pesonen T, et al. A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial. Microorganisms. 2023; 11(2):499. https://doi.org/10.3390/microorganisms11020499
Chicago/Turabian StyleLyra, Anna, Reeta Ala-Jaakkola, Nicolas Yeung, Neeta Datta, Kara Evans, Ashley Hibberd, Markus J. Lehtinen, Sofia D. Forssten, Alvin Ibarra, Tommi Pesonen, and et al. 2023. "A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial" Microorganisms 11, no. 2: 499. https://doi.org/10.3390/microorganisms11020499
APA StyleLyra, A., Ala-Jaakkola, R., Yeung, N., Datta, N., Evans, K., Hibberd, A., Lehtinen, M. J., Forssten, S. D., Ibarra, A., Pesonen, T., Junnila, J., Ouwehand, A. C., Baranowski, K., Maukonen, J., Crawford, G., & Lehtoranta, L. (2023). A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial. Microorganisms, 11(2), 499. https://doi.org/10.3390/microorganisms11020499