
Spray Drying Is a Viable Technology for the Preservation of Recombinant Proteins in Microalgae

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strains and Lab-Scale Cultivation
2.2. Design and Construction of the SAV Vaccine Transformation Plasmid
2.3. Transformation of the C. Reinhardtii Chloroplast
2.4. Cultivation in Single-Use ‘Hanging-Bag’ Photobioreactors
2.5. Harvesting by Centrifugation
2.6. Spray Drying
2.7. Freeze Drying
2.8. SDS-PAGE and Immunoblotting
2.8.1. Protein Ladder, Controls and Sample Preparation
2.8.2. Electrophoresis, Electroblotting and Antibody incubations
2.8.3. Visualisation and Densitometry Analysis
2.9. Statistical Analysis
2.10. Model Construction and Techno-Economic Analysis
2.10.1. Background and Definition of the Case Study
2.10.2. Mass Balances, Bioprocess Design and Operating Strategies
- i.
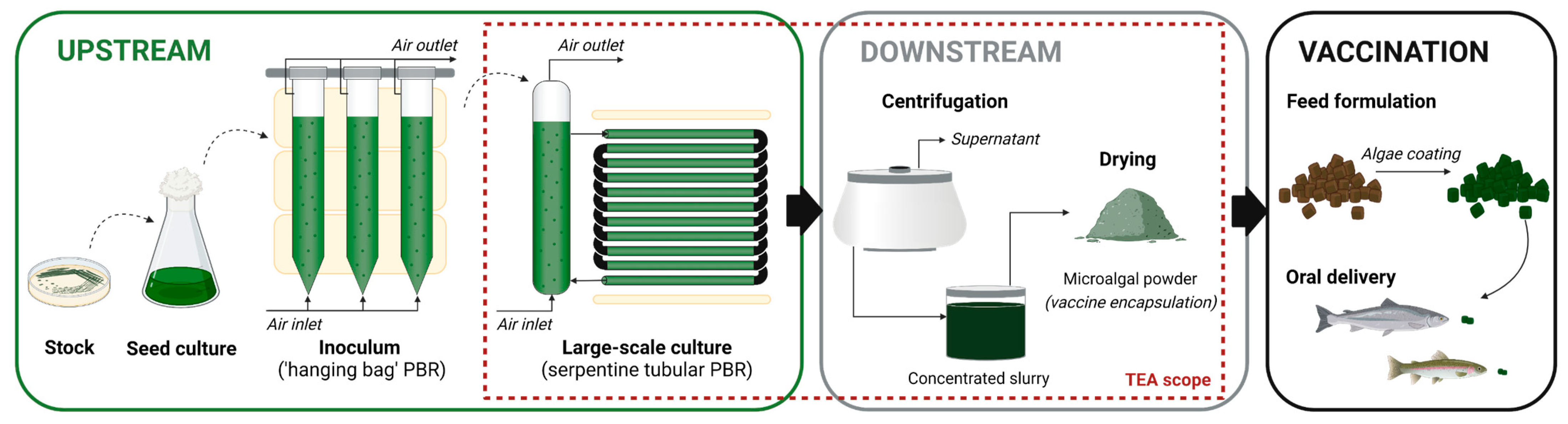
- Upstream: large-scale cultivation in a serpentine tubular photobioreactor
- ii.
- Centrifugation step
- iii.
- Freeze-drying strategy
- iv.
- Spray-drying strategy
2.10.3. Economic Model and Key Assumptions
- i.
- Capital expenditures (CAPEX)
- ii.
- Operating expenditures (OPEX)
- iii.
- Capital charges (A) and total cost of production (TCOP)
3. Results and Discussion
3.1. Investigating the Feasibility of Spray Drying for Downstream Processing
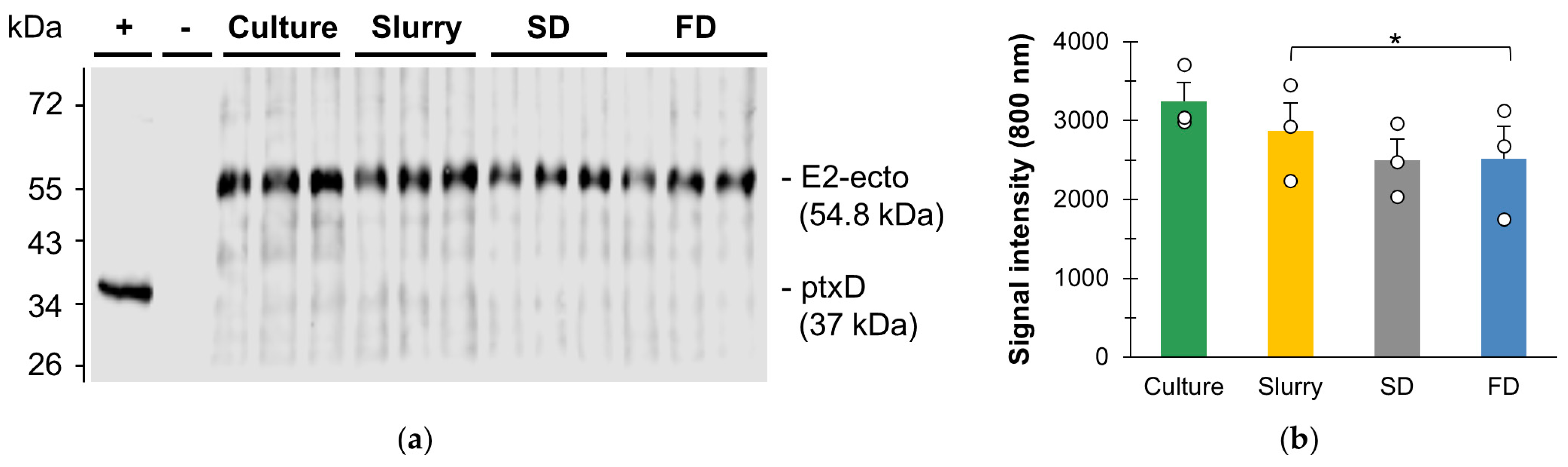
3.1.1. Influence of Spray Drying on Recombinant Protein Integrity
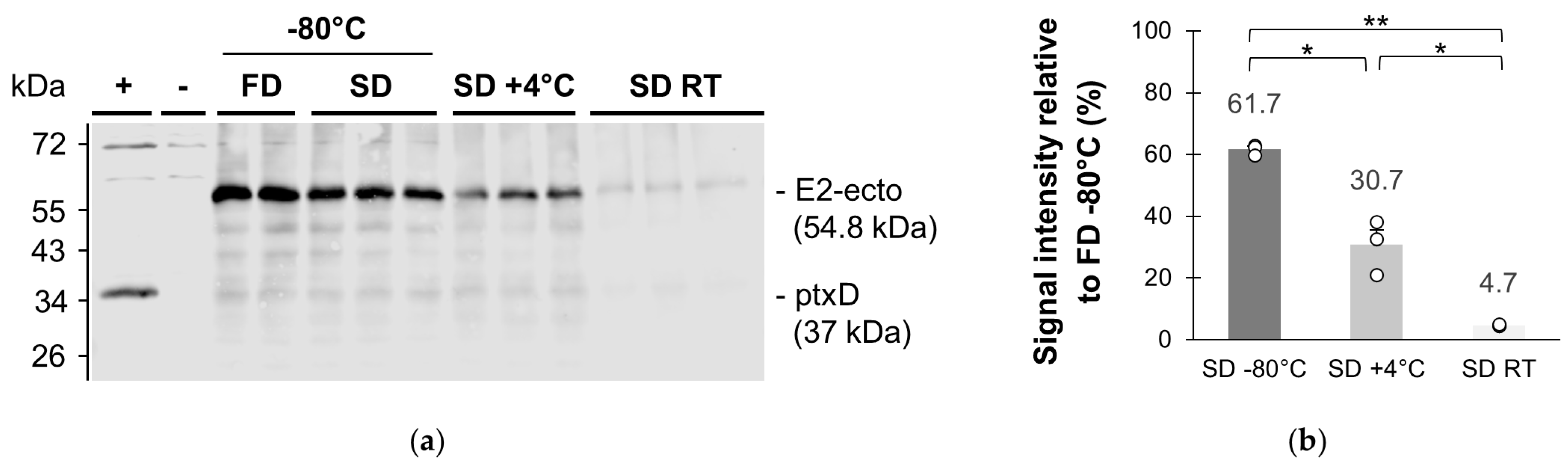
3.1.2. Stability of SAV Vaccine in Spray-Dried Powder over Time
3.2. Evaluation of Spray-drying Potential at Industrial Scale
3.2.1. Base Case Scenario: Economics and Scale of Production
3.2.2. Influence of the Drying Stage
3.2.3. Influence of the Vaccine Yield
3.2.4. Additional Considerations on the Selection of the Drying Technology
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Process-Flow Diagrams (PFD)


Appendix B. Design Details, Capital Investment and Operating Costs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PFD Code | Process Step | Equipment | Capacity | Nb Units | Lifespan (Years) | Total Installed Cost 1 |
|---|---|---|---|---|---|---|
| B1 | Inoculation | Plastic reservoir tanks | 5.3 a–5.9 b m3 | 1 | 10 | £21,700 a–£24,100 b |
| B2 | 0.08 a–0.09 b m3 | 1 | 10 | £300 a–£400 b | ||
| P1 | Pumps | 5 m3/h | 1 | 20 | £20,600 | |
| P2 | 0.1 m3/h | 1 | 20 | £16,200 | ||
| C1, F1, K1, F2, P3, K2 | Cell culture | Serpentine tubular PBR with LED array | 6.01 m3 | 1 | 10 | £165,400 |
| Upstream sub-total | £224,200 a–£226,700 b | |||||
| P4, S1 | Harvesting | Disc-stack centrifuge | 5 m3/h | 1 | 50 | £260,700 |
| B3 | Mixing tank b | 0.05 m3 | 1 | 20 | £17,700 b | |
| Harvesting sub-total | £260,700 a–£278,400 b | |||||
| H1, D1, C2, F3, V1 | Drying | Freeze dryer a | 50 kgice | 2 | 20 | £1,711,900 a |
| C2, F3, H1, P5, K3, S2, C3, F4 | Spray dryer b | 15 kgwater/h | 1 | 20 | £1,701,800 b | |
| Drying sub-total | £1,711,900a–£1,701,800 b | |||||
| PFD Code | Process Step/Details | Equipment | Energy | Annual Consumption | Annual Costs |
|---|---|---|---|---|---|
| P3 | Culture recirculation | PBR 2 | 200 W/m3culture | 5844 a–6493 b kWh/year | £871 a–£967 b |
| K2 | Culture illumination | 4 W/LPBR | 72,146 kWh/year | £10,750 | |
| - | Chemicals (TAP medium) 1 | - | - | £34,954 a–£38,838 b | |
| - | Water (TAP medium) | - | 478 a–531 b m3/year | £717 a–£797 b | |
| P4, S1 | Centrifugation | Disc-stack centrifuge 3 | 1.5 kWh/m 3,4 | 730 a–812 b kWh/year | £109 a–£121 b |
| H1, D1 | Drying | Freeze dryer a | 2 kWh/h | 24,000 kWh/year | £3576 a |
| H1, K3 | Spray dryer b | 50 kWh/h | 9969 kWh/year | £1485 b | |
| Total variable production costs (utilities + consumables) | £50,976 a–£52,958 b | ||||
| Value | Details | Annual Costs |
|---|---|---|
| Labour | 1× full-time biochemical engineer | £40,000 |
| 2× part-time (2/5) production operators | £20,000 | |
| Supervision, management | 25% operating labour [41] | £15,000 |
| Direct salary overhead | 50% operating labour + supervision [41] | £37,500 |
| Maintenance | 5% ISBL [41] | £109,838 a–£110,346 b |
| Insurance | 1% ISBL [41] | £21,968 a–£22,069 b |
| Total fixed production costs (labour + maintenance + insurance) | £244,306 a–£244,915 b | |
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Deperasińska, I.; Schulz, P.; Siwicki, A.K. Salmonid Alphavirus (SAV). J. Vet. Res. 2018, 62, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, M.D.; Bang Jensen, B.; McLoughlin, M.F.; Rodger, H.D.; Taksdal, T.; Sindre, H.; Graham, D.A.; Lillehaug, A. The Epidemiology of Pancreas Disease in Salmonid Aquaculture: A Summary of the Current State of Knowledge. J. Fish Dis. 2017, 40, 141–155. [Google Scholar] [CrossRef]
- Bedekar, M.K.; Kole, S. Fundamentals of Fish Vaccination. In Vaccine Design. Methods in Molecular Biology, 2nd ed.; Thomas, S., Ed.; Humana Press Inc.: New York, NY, USA, 2022; Volume 2411, pp. 147–173. [Google Scholar] [CrossRef]
- Wesolowska, A.; Ljunggren, M.K.; Jedlina, L.; Basalaj, K.; Legocki, A.; Wedrychowicz, H.; Kesik-Brodacka, M. A Preliminary Study of a Lettuce-Based Edible Vaccine Expressing the Cysteine Proteinase of Fasciola Hepatica for Fasciolosis Control in Livestock. Front. Immunol. 2018, 9, 2592. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Kim, H.J.; Lan, N.T.; Han, H.J.; Lee, D.-C.; Hwang, J.Y.; Kwon, M.-G.; Kang, B.K.; Han, S.Y.; Moon, H.; et al. Oral Vaccination through Voluntary Consumption of the Convict Grouper Epinephelus Septemfasciatus with Yeast Producing the Capsid Protein of Red-Spotted Grouper Nervous Necrosis Virus. Vet. Microbiol. 2017, 204, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Yang, R. Recent Progress in Bacillus Subtilis Spore-Surface Display: Concept, Progress, and Future. Appl. Microbiol. Biotechnol. 2017, 101, 933–949. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Bao, Q.; Wu, Y.; Chen, S.; Zhao, S.; Wu, H.; Fan, J. Evaluation of Microalgae as Immunostimulants and Recombinant Vaccines for Diseases Prevention and Control in Aquaculture. Front. Bioeng. Biotechnol. 2020, 8, 590431. [Google Scholar] [CrossRef]
- Dehghani, J.; Adibkia, K.; Movafeghi, A.; Maleki-Kakelar, H.; Saeedi, N.; Omidi, Y. Towards a New Avenue for Producing Therapeutic Proteins: Microalgae as a Tempting Green Biofactory. Biotechnol. Adv. 2020, 40, 107499. [Google Scholar] [CrossRef]
- Mayfield, S.P.; Franklin, S.E.; Lerner, R.A. Expression and Assembly of a Fully Active Antibody in Algae. Proc. Natl. Acad. Sci. USA 2003, 100, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Scaife, M.A.; Nguyen, G.T.D.T.; Rico, J.; Lambert, D.; Helliwell, K.E.; Smith, A.G. Establishing Chlamydomonas Reinhardtii as an Industrial Biotechnology Host. Plant J. 2015, 82, 532–546. [Google Scholar] [CrossRef]
- Dyo, Y.M.; Purton, S. The Algal Chloroplast as a Synthetic Biology Platform for Production of Therapeutic Proteins. Microbiology 2018, 164, 113–121. [Google Scholar] [CrossRef]
- Charoonnart, P.; Purton, S.; Saksmerprome, V. Applications of Microalgal Biotechnology for Disease Control in Aquaculture. Biology 2018, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Kiataramgul, A.; Maneenin, S.; Purton, S.; Areechon, N.; Hirono, I.; Brocklehurst, T.W.; Unajak, S. An Oral Delivery System for Controlling White Spot Syndrome Virus Infection in Shrimp Using Transgenic Microalgae. Aquaculture 2020, 521, 735022. [Google Scholar] [CrossRef]
- Taunt, H.N.; Stoffels, L.; Purton, S. Green Biologics: The Algal Chloroplast as a Platform for Making Biopharmaceuticals. Bioengineered 2018, 9, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Molina Grima, E.; Belarbi, E.-H.; Acién Fernández, F.G.; Robles Medina, A.; Chisti, Y. Recovery of Microalgal Biomass and Metabolites: Process Options and Economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- Milledge, J.J.; Heaven, S. A Review of the Harvesting of Micro-Algae for Biofuel Production. Rev. Environ. Sci. Biotechnol. 2013, 12, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Leach, G.; Oliveira, G.; Morais, R. Spray-Drying of Dunaliella Salina to Produce a β-Carotene Rich Powder. J. Ind. Microbiol. Biotechnol. 1998, 20, 82–85. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, T.; Li, J.; Pan, B.; Hu, Q.; Duan, M.; Zhang, X. Study on the Effect of Spray Drying Process on the Quality of Microalgal Biomass: A Comprehensive Biocomposition Analysis of Spray-Dried S. Acuminatus Biomass. Bioenergy Res. 2022, 15, 320–333. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Muylaert, K.; Eeckhout, M.; Ruyssen, T.; Foubert, I. Influence of Drying and Storage on Lipid and Carotenoid Stability of the Microalga Phaeodactylum Tricornutum. J. Agric. Food Chem. 2011, 59, 11063–11069. [Google Scholar] [CrossRef]
- Lin, L.P. Microstructure of Spray-Dried and Freeze-Dried Microalgal Powders. Food Microstruct. 1985, 4, 341–348. [Google Scholar]
- Wannathong, T.; Waterhouse, J.C.; Young, R.E.B.; Economou, C.K.; Purton, S. New Tools for Chloroplast Genetic Engineering Allow the Synthesis of Human Growth Hormone in the Green Alga Chlamydomonas Reinhardtii. Appl. Microbiol. Biotechnol. 2016, 100, 5467–5477. [Google Scholar] [CrossRef] [Green Version]
- Gorman, D.S.; Levine, R.P. Cytochrome f and Plastocyanin: Their Sequence in the Photosynthetic Electron Transport Chain of Chlamydomonas Reinhardi. Proc. Natl. Acad. Sci. USA 1965, 54, 1665–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropat, J.; Hong-Hermesdorf, A.; Casero, D.; Ent, P.; Castruita, M.; Pellegrini, M.; Merchant, S.S.; Malasarn, D. A Revised Mineral Nutrient Supplement Increases Biomass and Growth Rate in Chlamydomonas Reinhardtii. Plant J. 2011, 66, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Petterson, E.; Stormoen, M.; Evensen, Ø.; Mikalsen, A.B.; Haugland, Ø. Natural Infection of Atlantic Salmon (Salmo salar L.) with Salmonid Alphavirus 3 Generates Numerous Viral Deletion Mutants. J. Gen. Virol. 2013, 94, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.A.; Topol, A.B.; Doerner, D.Z.; Mayfield, S. Alga-Produced Cholera Toxin-Pfs25 Fusion Proteins as Oral Vaccines. Appl. Environ. Microbiol. 2013, 79, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- Taylor, G.M.; Mordaka, P.M.; Heap, J.T. Start-Stop Assembly: A Functionally Scarless DNA Assembly System Optimized for Metabolic Engineering. Nucleic Acids Res. 2019, 47, e17. [Google Scholar] [CrossRef] [Green Version]
- Weber, E.; Engler, C.; Gruetzner, R.; Werner, S.; Marillonnet, S. A Modular Cloning System for Standardized Assembly of Multigene Constructs. PLoS ONE 2011, 6, e16765. [Google Scholar] [CrossRef]
- Cui, J.; Purton, S.; Baganz, F. Characterisation of a Simple ‘Hanging Bag’ Photobioreactor for Low-Cost Cultivation of Microalgae. J. Chem. Technol. Biotechnol. 2022, 97, 608–619. [Google Scholar] [CrossRef]
- Changko, S.; Rajakumar, P.D.; Young, R.E.B.; Purton, S. The Phosphite Oxidoreductase Gene, PtxD as a Bio-Contained Chloroplast Marker and Crop-Protection Tool for Algal Biotechnology Using Chlamydomonas. Appl. Microbiol. Biotechnol. 2020, 104, 675–686. [Google Scholar] [CrossRef] [Green Version]
- FAO. Fishery and Aquaculture Statistics. Global Aquaculture Production 1950-2020 (FishStatJ). In FAO Fisheries and Aquaculture Division; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/fishery/en/topic/166235?lang=en (accessed on 7 December 2022).
- Munro, L.A. Scottish Fish Farm Production Survey 2018. Available online: https://www.gov.scot/publications/scottish-fish-farm-production-survey-2018/ (accessed on 7 December 2022).
- Singh, G.; Patidar, S.K. Microalgae Harvesting Techniques: A Review. J. Environ. Manag. 2018, 217, 499–508. [Google Scholar] [CrossRef]
- Boychyn, M.; Yim, S.S.S.; Ayazi Shamlou, P.; Bulmer, M.; More, J.; Hoare, M. Characterization of Flow Intensity in Continuous Centrifuges for the Development of Laboratory Mimics. Chem. Eng. Sci. 2001, 56, 4759–4770. [Google Scholar] [CrossRef]
- Stoffels, L.; Finlan, A.; Mannall, G.; Purton, S.; Parker, B. Downstream Processing of Chlamydomonas Reinhardtii TN72 for Recombinant Protein Recovery. Front. Bioeng. Biotechnol. 2019, 7, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, F.; Li, Y.; Fanning, K.; Netzel, M.; Schenk, P.M. Effect of Drying, Storage Temperature and Air Exposure on Astaxanthin Stability from Haematococcus Pluvialis. Food Res. Int. 2015, 74, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopmann, I.K.; Möller, S.; Elle, C.; Hindersin, S.; Kramer, A.; Labes, A. Optimization of Astaxanthin Recovery in the Downstream Process of Haematococcus Pluvialis. Foods 2022, 11, 1352. [Google Scholar] [CrossRef] [PubMed]
- OECD. Monthly Monetary and Financial Statistics (MEI): Exchange Rates (USD Monthly Averages). Available online: https://stats.oecd.org/index.aspx?queryid=169 (accessed on 7 December 2022).
- Towler, G.; Sinnott, R. Chapter 7—Capital Cost Estimating. In Chemical Engineering Design; Towler, G., Sinnott, R., Eds.; Butterworth-Heinemann: Oxford, UK, 2013; pp. 307–354. [Google Scholar] [CrossRef]
- Department for Business, Energy & Industrial Strategy. Gas and Electricity Prices in the Non-Domestic Sector. 2022. Available online: https://www.gov.uk/government/statistical-data-sets/gas-and-electricity-prices-in-the-non-domestic-sector (accessed on 7 December 2022).
- Towler, G.; Sinnott, R. Chapter 8—Estimating Revenues and Production Costs. In Chemical Engineering Design; Towler, G., Sinnott, R., Eds.; Butterworth-Heinemann: Oxford, UK, 2013; pp. 355–387. [Google Scholar] [CrossRef]
- Towler, G.; Sinnott, R. Chapter 9—Economic Evaluation of Projects. In Chemical Engineering Design; Towler, G., Sinnott, R., Eds.; Butterworth-Heinemann: Oxford, UK, 2013; pp. 389–429. [Google Scholar] [CrossRef]
- Raposo, M.F.J.; Morais, A.M.M.B.; Morais, R.M.S.C. Effects of Spray-Drying and Storage on Astaxanthin Content of Haematococcus Pluvialis Biomass. World J. Microbiol. Biotechnol. 2012, 28, 1253–1257. [Google Scholar] [CrossRef] [Green Version]
- Norsker, N.H.; Barbosa, M.J.; Vermuë, M.H.; Wijffels, R.H. Microalgal Production—A Close Look at the Economics. Biotechnol. Adv. 2011, 29, 24–27. [Google Scholar] [CrossRef]
- Tredici, M.R.; Rodolfi, L.; Biondi, N.; Bassi, N.; Sampietro, G. Techno-Economic Analysis of Microalgal Biomass Production in a 1-Ha Green Wall Panel (GWP®) Plant. Algal Res. 2016, 19, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Romero, B.; Perales, J.A.; Pereira, H.; Barbosa, M.; Ruiz, J. Techno-Economic Assessment of Microalgae Production, Harvesting and Drying for Food, Feed, Cosmetics, and Agriculture. Sci. Total Environ. 2022, 837, 155742. [Google Scholar] [CrossRef]
- Acién, F.G.; Fernández, J.M.; Magán, J.J.; Molina, E. Production Cost of a Real Microalgae Production Plant and Strategies to Reduce It. Biotechnol. Adv. 2012, 30, 1344–1353. [Google Scholar] [CrossRef]
- Tredici, M.; Rodolfi, L. Reactor for Industrial Culture of Photosynthetic Microorganisms. WO/2004/074423, 2 September 2004. [Google Scholar]
- Fernández, F.G.A.; Reis, A.; Wijffels, R.H.; Barbosa, M.; Verdelho, V.; Llamas, B. The Role of Microalgae in the Bioeconomy. New Biotechnol. 2021, 61, 99–107. [Google Scholar] [CrossRef]
- Coutteau, P.; Sorgeloos, P. The Use of Algal Substitutes and the Requirement for Live Algae in the Hatchery and Nursery Rearing of Bivalve Molluscs: An International Survey. J. Shellfish. Res. 1992, 11, 467–476. [Google Scholar]
- Oostlander, P.C.; van Houcke, J.; Wijffels, R.H.; Barbosa, M.J. Microalgae Production Cost in Aquaculture Hatcheries. Aquaculture 2020, 525, 735310. [Google Scholar] [CrossRef]
- Fasaei, F.; Bitter, J.H.; Slegers, P.M.; van Boxtel, A.J.B. Techno-Economic Evaluation of Microalgae Harvesting and Dewatering Systems. Algal Res. 2018, 31, 347–362. [Google Scholar] [CrossRef]
- Tredici, M.R.; Bassi, N.; Prussi, M.; Biondi, N.; Rodolfi, L.; Chini Zittelli, G.; Sampietro, G. Energy Balance of Algal Biomass Production in a 1-Ha “Green Wall Panel” Plant: How to Produce Algal Biomass in a Closed Reactor Achieving a High Net Energy Ratio. Appl. Energy 2015, 154, 1103–1111. [Google Scholar] [CrossRef] [Green Version]





| Parameters | Details | Reference/Notes |
|---|---|---|
| Location | United Kingdom (UK) | Market, process location |
| Year of study | 2019 | |
| Vaccination targets | Atlantic salmon and rainbow trout | 82,000,000 fish/year (UK) 1 |
| Vaccine dose | 10 µgvaccine/fish | Industry communication |
| Microalgal strain | TN72:E2-ecto | |
| Vaccine yield | 3.15 gvaccine/kgDCW | Experimental results (this study) |
| Annual production | 286 kgDCW/year | 10% safety margin |
| Operating time | 300 days/year |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilatte, A.; Spencer-Milnes, X.; Jackson, H.O.; Purton, S.; Parker, B. Spray Drying Is a Viable Technology for the Preservation of Recombinant Proteins in Microalgae. Microorganisms 2023, 11, 512. https://doi.org/10.3390/microorganisms11020512
Vilatte A, Spencer-Milnes X, Jackson HO, Purton S, Parker B. Spray Drying Is a Viable Technology for the Preservation of Recombinant Proteins in Microalgae. Microorganisms. 2023; 11(2):512. https://doi.org/10.3390/microorganisms11020512
Chicago/Turabian StyleVilatte, Anaëlle, Xenia Spencer-Milnes, Harry Oliver Jackson, Saul Purton, and Brenda Parker. 2023. "Spray Drying Is a Viable Technology for the Preservation of Recombinant Proteins in Microalgae" Microorganisms 11, no. 2: 512. https://doi.org/10.3390/microorganisms11020512
APA StyleVilatte, A., Spencer-Milnes, X., Jackson, H. O., Purton, S., & Parker, B. (2023). Spray Drying Is a Viable Technology for the Preservation of Recombinant Proteins in Microalgae. Microorganisms, 11(2), 512. https://doi.org/10.3390/microorganisms11020512