
Transgenic Microalgae Expressing Double-Stranded RNA as Potential Feed Supplements for Controlling White Spot Syndrome in Shrimp Aquaculture

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strain and Maintenance
2.2. Plasmid Design and Construction
2.3. Confirmation of dsRNA Expression in E. coli
2.4. Chloroplast Transformation and Transformant Selection
2.5. Double-Stranded RNA Detection and Quantification
2.6. Hanging-Bag Culture and Biomass Preparation
2.7. Preparation of Animals and Viral Inoculum
2.8. Assay for Viral Protection in Shrimp
2.9. Statement of Informed Animal Rights
3. Results
3.1. A New Vector for dsRNA Production in the C. reinhardtii Chloroplast through Convergent Transcription
3.2. Golden Gate Assembly Allows Quick and Efficient Construction of Chloroplast Transformation Plasmids
3.3. The Dual-rrnS Promoter System Is Capable of dsRNA Production in E. coli
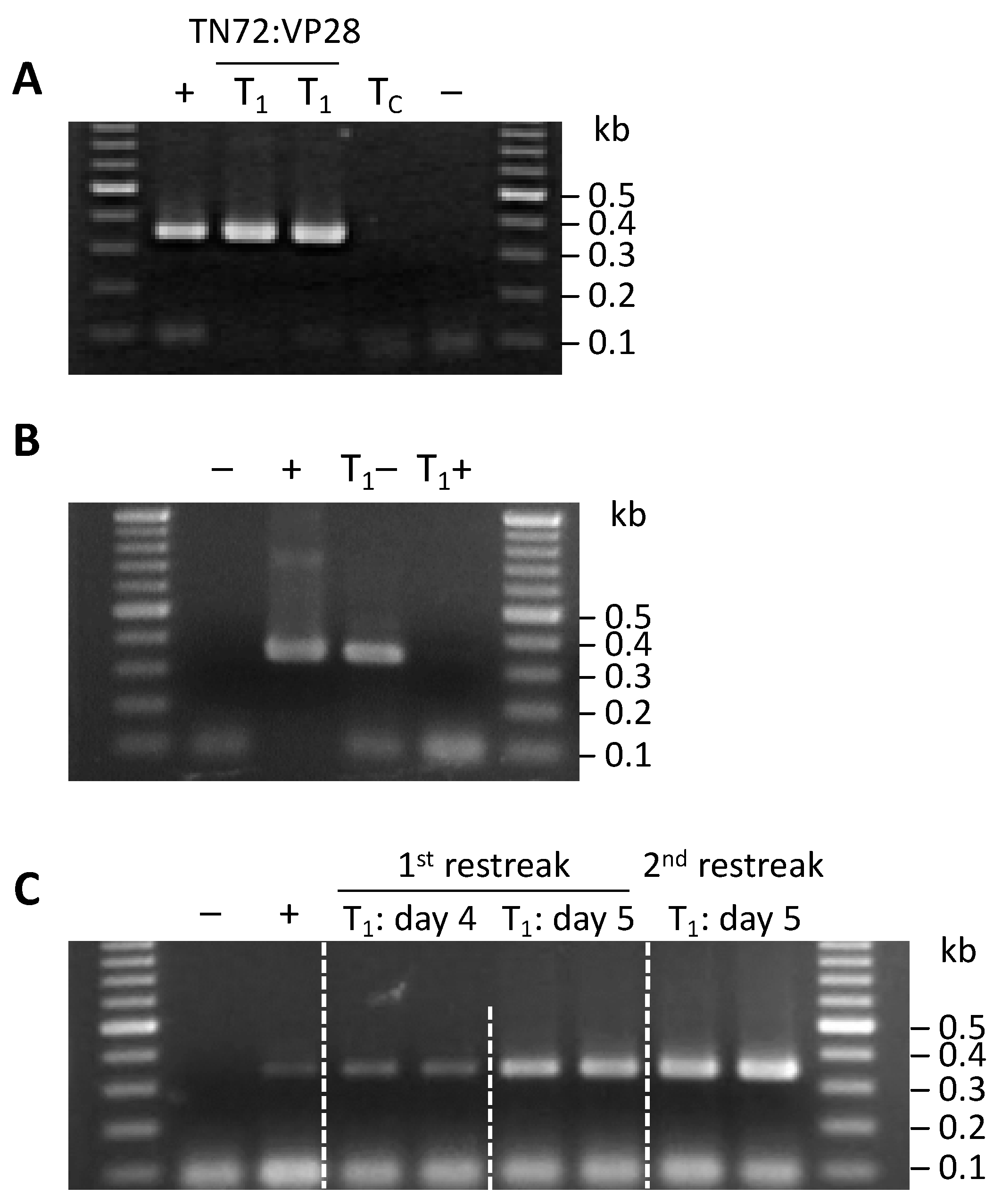
3.4. Generation of C. reinhardtii Transplastomic Lines
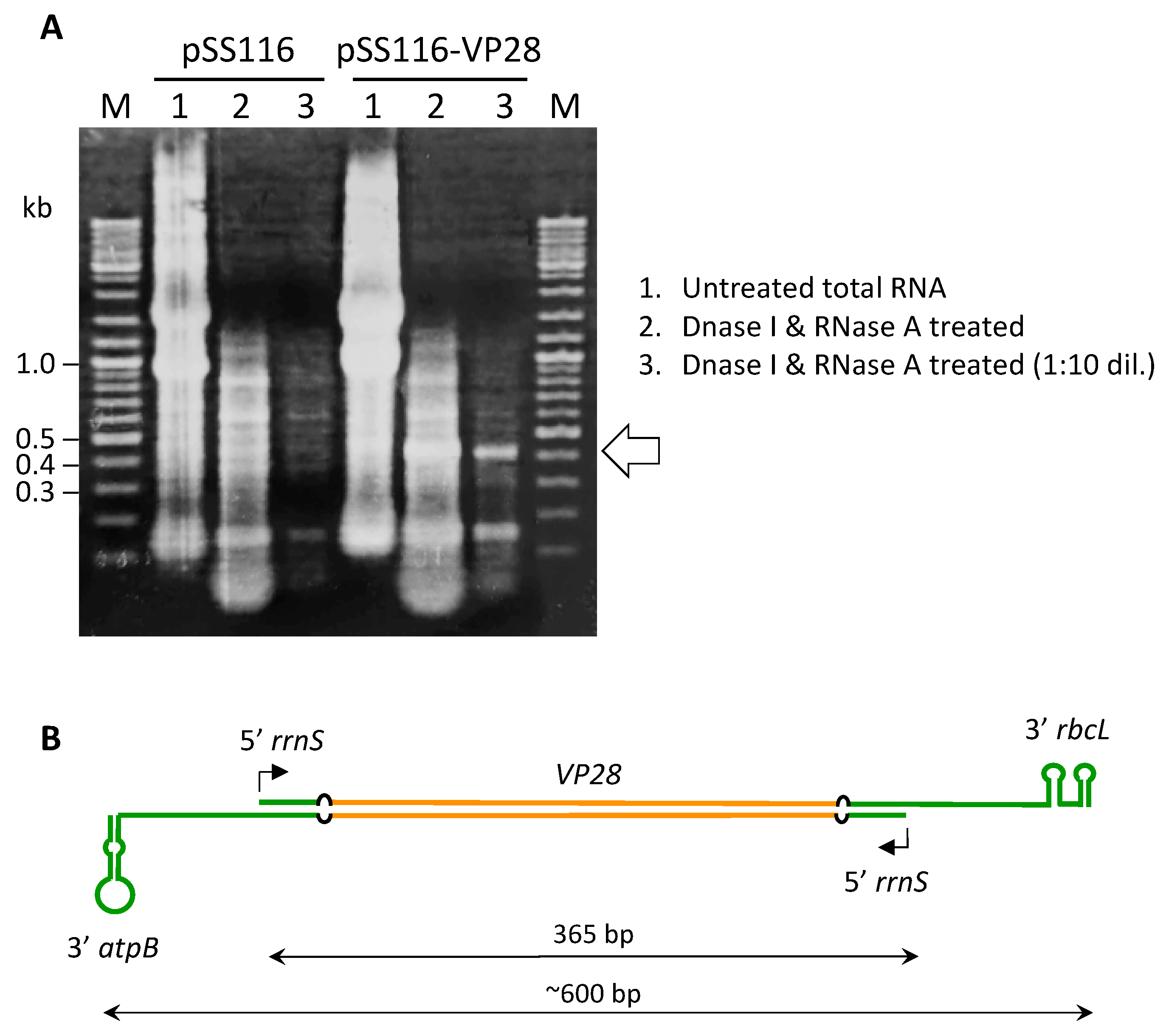
3.5. Convergent rrnS Promoters Enhance Production of dsRNA Relative to the Previous Study
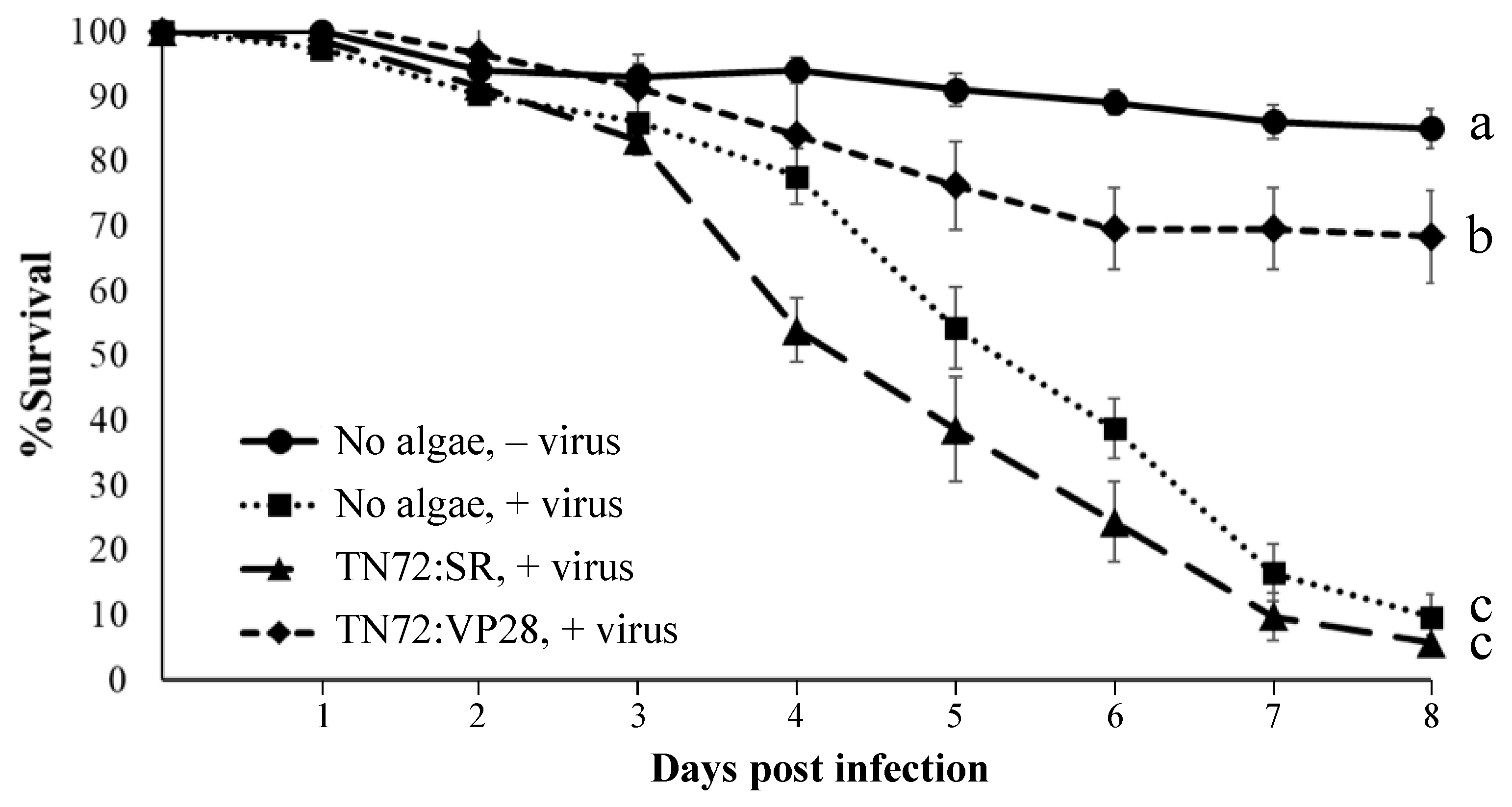
3.6. Shrimp Fed with the TN72-dsVP28 Line Show ~69% Survival against WSSV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oakey, J.; Smith, C.; Underwood, D.; Afsharnasab, M.; Alday-Sanz, V.; Dhar, A.; Sivakumar, S.; Sahul Hameed, A.S.; Beattie, K.; Crook, A. Global distribution of white spot syndrome virus genotypes determined using a novel genotyping assay. Arch. Virol. 2019, 164, 2061–2082. [Google Scholar] [CrossRef]
- Flegel, T.W.; Lightner, D.V.; Lo, C.F.; Owens, L.; Bondad-Reantaso, M.G.; Mohan, C.V.; Crumlish, M.; Subasinghe, R.P. Diseases in Asian Aquaculture VI; Fish Health Section, Asian Fisheries Society: Manila, Philippines, 2008; pp. 355–378. [Google Scholar]
- Krishnan, P.; Babu, P.G.; Rajendran, K.; Chaudhari, A. RNA interference-based therapeutics for shrimp viral diseases. Dis. Aquat. Org. 2009, 86, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Wang, Z.; Qi, Y.; Yao, L.; Qian, J.; Hu, L. Vp28 of Shrimp White Spot Syndrome Virus Is Involved in the Attachment and Penetration into Shrimp Cells. BMB Rep. 2004, 37, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Han, F.; Zhang, X. Silencing shrimp white spot syndrome virus (WSSV) genes by siRNA. Antivir. Res. 2007, 73, 126–131. [Google Scholar] [CrossRef]
- Thammasorn, T.; Sangsuriya, P.; Meemetta, W.; Senapin, S.; Jitrakorn, S.; Rattanarojpong, T.; Saksmerprome, V. Large-scale production and antiviral efficacy of multi-target double-stranded RNA for the prevention of white spot syndrome virus (WSSV) in shrimp. BMC Biotechnol. 2015, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Ward, V. Production of recombinant and therapeutic proteins in microalgae. Curr. Opin. Biotechnol. 2022, 78, 102784. [Google Scholar] [CrossRef] [PubMed]
- Fabris, M.; Abbriano, R.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiumparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef]
- Muller-Feuga, A. The role of microalgae in aquaculture: Situation and trends. J. Appl. Phycol. 2000, 12, 527–534. [Google Scholar] [CrossRef]
- Kumar, G.; Shekh, A.; Jakhu, S.; Sharma, Y.; Kapoor, R.; Sharma, T.R. Bioengineering of Microalgae: Recent Advances, Perspectives, and Regulatory Challenges for Industrial Application. Front. Bioeng. Biotechnol. 2020, 8, 914. [Google Scholar] [CrossRef]
- Cutolo, E.A.; Mandalà, G.; Dall’osto, L.; Bassi, R. Harnessing the Algal Chloroplast for Heterologous Protein Production. Microorganisms 2022, 10, 743. [Google Scholar] [CrossRef]
- Fajardo, C.; De Donato, M.; Carrasco, R.; Martínez-Rodríguez, G.; Mancera, J.M.; Fernández-Acero, F.J. Advances and challenges in genetic engineering of microalgae. Rev. Aquac. 2019, 12, 365–381. [Google Scholar] [CrossRef]
- Mussgnug, J.H. Genetic tools and techniques for Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2015, 99, 5407–5418. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.O.; Taunt, H.N.; Mordaka, P.M.; Smith, A.G.; Purton, S. The Algal Chloroplast as a Testbed for Synthetic Biology Designs Aimed at Radically Rewiring Plant Metabolism. Front. Plant Sci. 2021, 12, 708370. [Google Scholar] [CrossRef] [PubMed]
- De Vitry, C.; Vallon, O. Mutants of Chlamydomonas: Tools to study thylakoid membrane structure, function and biogenesis. Biochimie 1999, 81, 631–643. [Google Scholar] [CrossRef]
- Esland, L.; Larrea-Alvarez, M.; Purton, S. Selectable Markers and Reporter Genes for Engineering the Chloroplast of Chlamydomonas reinhardtii. Biology 2018, 7, 46. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; Solís-Andrade, K.I.; Márquez-Escobar, V.A.; González-Ortega, O.; Bañuelos-Hernandez, B. Current advances in the algae-made biopharmaceuticals field. Expert Opin. Biol. Ther. 2020, 20, 751–766. [Google Scholar] [CrossRef]
- Charoonnart, P.; Worakajit, N.; Zedler, J.A.Z.; Meetam, M.; Robinson, C.; Saksmerprome, V. Generation of microalga Chlamydomonas reinhardtii expressing shrimp antiviral dsRNA without supplementation of antibiotics. Sci. Rep. 2019, 9, 3164. [Google Scholar] [CrossRef]
- Wannathong, T.; Waterhouse, J.C.; Young, R.E.B.; Economou, C.K.; Purton, S. New tools for chloroplast genetic engineering allow the synthesis of human growth hormone in the green alga Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2016, 100, 5467–5477. [Google Scholar] [CrossRef]
- Kropat, J.; Hong-Hermesdorf, A.; Casero, D.; Ent, P.; Castruita, M.; Pellegrini, M.; Merchant, S.S.; Malasarn, D. A revised mineral nutrient supplement increases biomass and growth rate in Chlamydomonas reinhardtii. Plant J. 2011, 66, 770–780. [Google Scholar] [CrossRef]
- Taylor, G.M.; Mordaka, P.M.; Heap, J.T. Start-Stop Assembly: A functionally scarless DNA assembly system optimized for metabolic engineering. Nucleic Acids Res. 2018, 47, e17. [Google Scholar] [CrossRef]
- Papić, L.; Rivas, J.; Toledo, S.; Romero, J. Double-stranded RNA production and the kinetics of recombinant Escherichia coli HT115 in fed-batch culture. Biotechnol. Rep. 2018, 20, e00292. [Google Scholar] [CrossRef] [PubMed]
- Economou, C.; Wannathong, T.; Szaub, J.; Purton, S. A Simple, Low-Cost Method for Chloroplast Transformation of the Green Alga Chlamydomonas reinhardtii. Methods Mol. Biol. 2014, 1132, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Juntadech, T.; Yokthongwattana, K.; Tangphatsornruang, S.; Yap, Y.-K.; Katzenmeier, G.; Angsuthanasombat, C. Efficient transcription of the larvicidal cry4Ba gene from Bacillus thuringiensis in transgenic chloroplasts of the green algal Chlamydomonas reinhardtii. Adv. Biosci. Biotechnol. 2012, 3, 362–369. [Google Scholar] [CrossRef]
- Cui, J.; Purton, S.; Baganz, F. Characterisation of a simple ‘hanging bag’ photobioreactor for low-cost cultivation of microalgae. J. Chem. Technol. Biotechnol. 2021, 97, 608–619. [Google Scholar] [CrossRef]
- Srivihok, C.; Prachumwat, A.; Taengchaiyaphum, S.; Sritunyalucksana, K.; Thitamadee, S. Characterization of survivability-associated transcript of Penaeus monodon upon WSSV infection. In The 11th National Science Research Conference; Srinakharinwirot University: Bangkok, Thailand, 2019; pp. 152–159. [Google Scholar]
- Blowers, A.D.; Ellmore, G.S.; Klein, U.; Bogorad, L. Transcriptional analysis of endogenous and foreign genes in chloroplast transformants of Chlamydomonas. Plant Cell 1990, 2, 1059–1070. [Google Scholar] [CrossRef]
- Rott, R.; Liveanu, V.; Drager, R.G.; Stern, D.B.; Schuster, G. The sequence and structure of the 3′-untranslated regions of chloroplast transcripts are important determinants of mRNA accumulation and stability. Plant Mol. Biol. 1998, 36, 307–314. [Google Scholar] [CrossRef]
- Bird, J.E.; Marles-Wright, J.; Giachino, A. A User’s Guide to Golden Gate Cloning Methods and Standards. ACS Synth. Biol. 2022, 11, 3551–3563. [Google Scholar] [CrossRef] [PubMed]
- Purton, S. Tools and Techniques for Chloroplast Transformation of Chlamydomonas. Adv. Exp. Med. Biol. 2007, 616, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.A.; Muto, M.; Sullivan, J.; Mayfield, S.P. Improved heterologous protein expression in the chloroplast of Chlamydomonas reinhardtii through promoter and 5′ untranslated region optimization. Plant Biotechnol. J. 2011, 9, 674–683. [Google Scholar] [CrossRef]
- García, K.; Ramírez-Araya, S.; Díaz, Á.; Reyes-Cerpa, S.; Espejo, R.T.; Higuera, G.; Romero, J. Inactivated E. coli transformed with plasmids that produce dsRNA against infectious salmon anemia virus hemagglutinin show antiviral activity when added to infected ASK cells. Front. Microbiol. 2015, 6, 300. [Google Scholar] [CrossRef]
- Kim, E.; Park, Y.; Kim, Y. A Transformed Bacterium Expressing Double-Stranded RNA Specific to Integrin β1 Enhances Bt Toxin Efficacy against a Polyphagous Insect Pest, Spodoptera exigua. PLoS ONE 2015, 10, e0132631. [Google Scholar] [CrossRef]
- Chen, Z.; He, J.; Luo, P.; Li, X.; Gao, Y. Production of functional double-stranded RNA using a prokaryotic expression system in Escherichia coli. MicrobiologyOpen 2018, 8, e787. [Google Scholar] [CrossRef]
- Kumar, A.; Wang, S.; Ou, R.; Samrakandi, M.; Beerntsen, B.; Sayre, R. Development of an RNAi based microalgal larvicide to control mosquitoes. Malaria World J. 2013, 4, 6. [Google Scholar]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Murbach, T.S.; Glávits, R.; Endres, J.R.; Hirka, G.; Vértesi, A.; Béres, E.; Szakonyiné, I.P. A Toxicological Evaluation of Chlamydomonas reinhardtii, a Green Algae. Int. J. Toxicol. 2018, 37, 53–62. [Google Scholar] [CrossRef]
- Lee, W.; Matthews, A.; Moore, D. Safety Evaluation of a Novel Algal Feed Additive for Poultry Production. Avian Dis. 2022, 66, 326–336. [Google Scholar] [CrossRef]
- He, W.; Xu, W.; Xu, L.; Fu, K.; Guo, W.; Bock, R.; Zhang, J. Length-dependent accumulation of double-stranded RNAs in plastids affects RNA interference efficiency in the Colorado potato beetle. J. Exp. Bot. 2020, 71, 2670–2677. [Google Scholar] [CrossRef]
- Fasaei, F.; Bitter, J.; Slegers, P.; van Boxtel, A. Techno-economic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018, 31, 347–362. [Google Scholar] [CrossRef]
- Vilatte, A.; Spencer-Milnes, X.; Jackson, H.O.; Purton, S.; Parker, B. Spray Drying Is a Viable Technology for the Preservation of Recombinant Proteins in Microalgae. Microorganisms 2023, 11, 512. [Google Scholar] [CrossRef]
- Kongkumnerd, J.; Saleetid, N. Screening survey for diseases in marine shrimp hatchery and nursery. E-Thai Fish. Gaz. 2019, 2, 23–48. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charoonnart, P.; Taunt, H.N.; Yang, L.; Webb, C.; Robinson, C.; Saksmerprome, V.; Purton, S. Transgenic Microalgae Expressing Double-Stranded RNA as Potential Feed Supplements for Controlling White Spot Syndrome in Shrimp Aquaculture. Microorganisms 2023, 11, 1893. https://doi.org/10.3390/microorganisms11081893
Charoonnart P, Taunt HN, Yang L, Webb C, Robinson C, Saksmerprome V, Purton S. Transgenic Microalgae Expressing Double-Stranded RNA as Potential Feed Supplements for Controlling White Spot Syndrome in Shrimp Aquaculture. Microorganisms. 2023; 11(8):1893. https://doi.org/10.3390/microorganisms11081893
Chicago/Turabian StyleCharoonnart, Patai, Henry Nicholas Taunt, Luyao Yang, Conner Webb, Colin Robinson, Vanvimon Saksmerprome, and Saul Purton. 2023. "Transgenic Microalgae Expressing Double-Stranded RNA as Potential Feed Supplements for Controlling White Spot Syndrome in Shrimp Aquaculture" Microorganisms 11, no. 8: 1893. https://doi.org/10.3390/microorganisms11081893
APA StyleCharoonnart, P., Taunt, H. N., Yang, L., Webb, C., Robinson, C., Saksmerprome, V., & Purton, S. (2023). Transgenic Microalgae Expressing Double-Stranded RNA as Potential Feed Supplements for Controlling White Spot Syndrome in Shrimp Aquaculture. Microorganisms, 11(8), 1893. https://doi.org/10.3390/microorganisms11081893