
Prioritization of Microorganisms Isolated from the Indian Ocean Sponge Scopalina hapalia Based on Metabolomic Diversity and Biological Activity for the Discovery of Natural Products

 , ,
, ,  , ,
, ,  , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.1.1. Sponge
2.1.2. Microbial Strains
2.2. Strains Culture and Extracts Preparation
2.3. Chemical Analysis
2.3.1. HPLC-DAD-CAD Analysis
2.3.2. UHPLC-QTOF-MS/MS Analysis
2.4. Raw Data Processing
2.5. Ion Identity Molecular Network
2.6. Feature Annotations
2.7. Biological Activity Tests
2.7.1. Cytotoxic Activity
2.7.2. Antiplasmodial Activity
3. Results
3.1. Metabolomic Study
3.1.1. HPLC-DAD-CAD Analysis
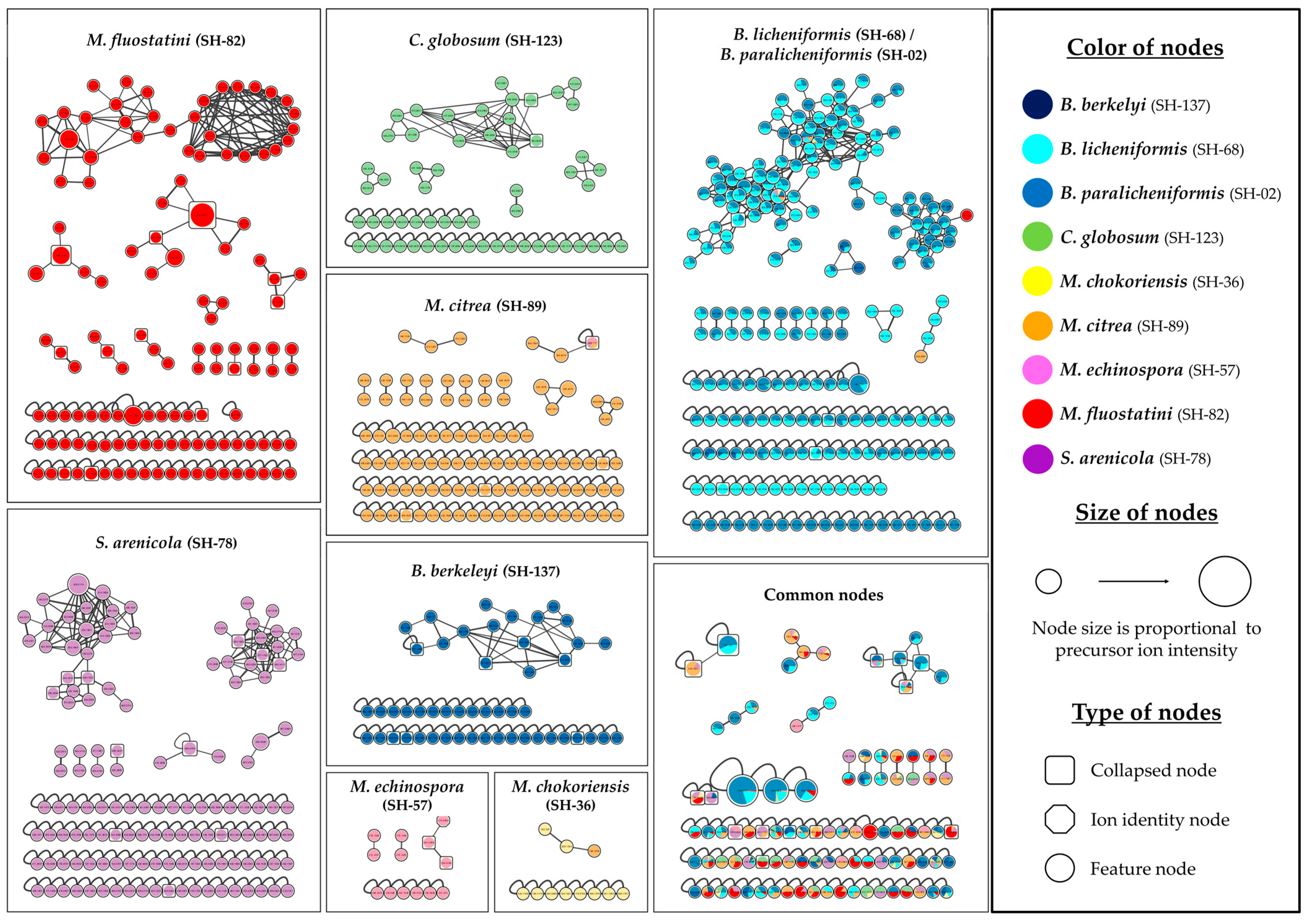
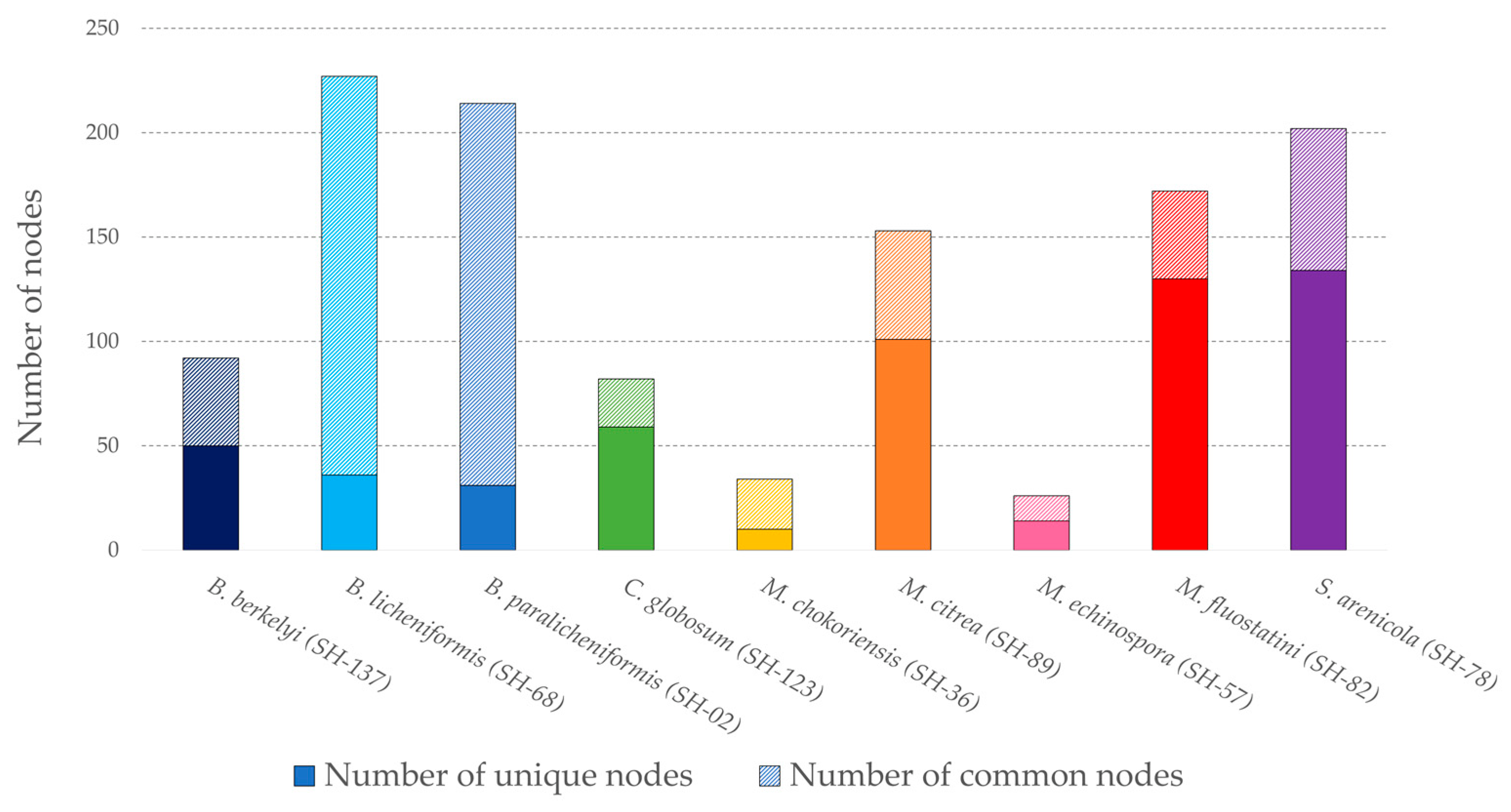
3.1.2. Ion Identity Molecular Network
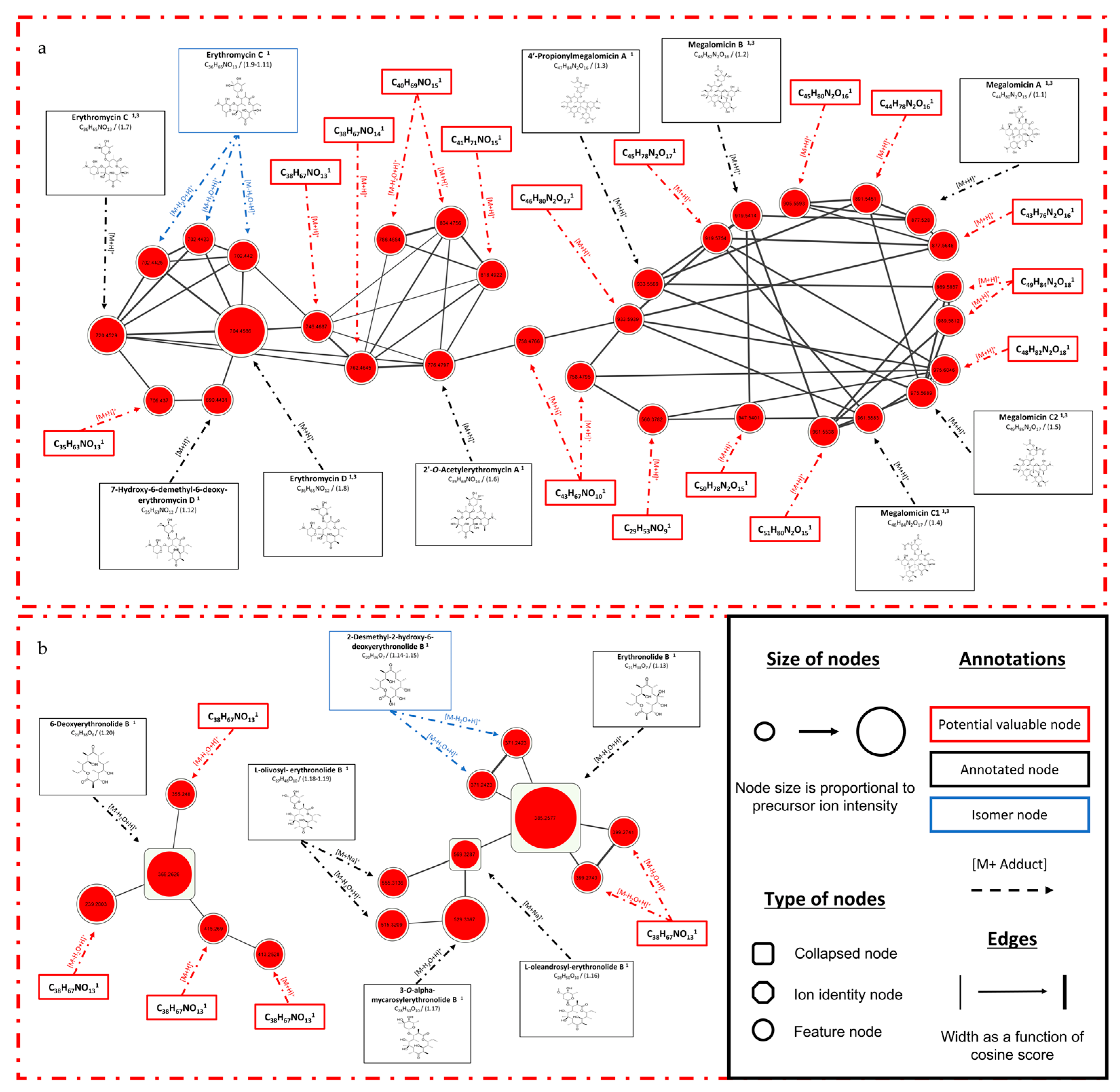
3.1.3. Focus on Micromonospora fluostatini SH-82
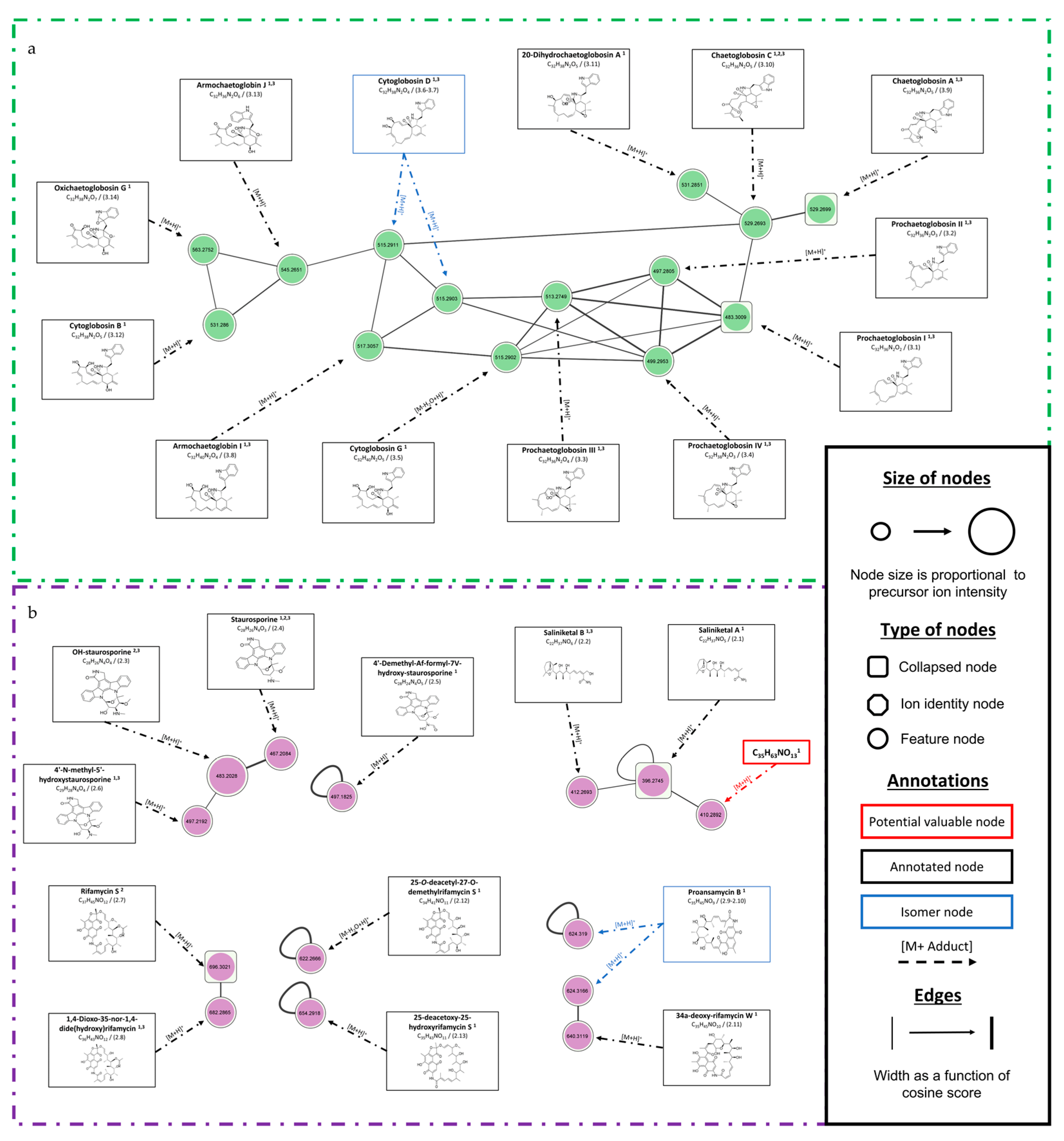
3.1.4. Focus on Chaetomium globosum SH-123 and Salinispora arenicola SH-78
3.2. Biological Activity
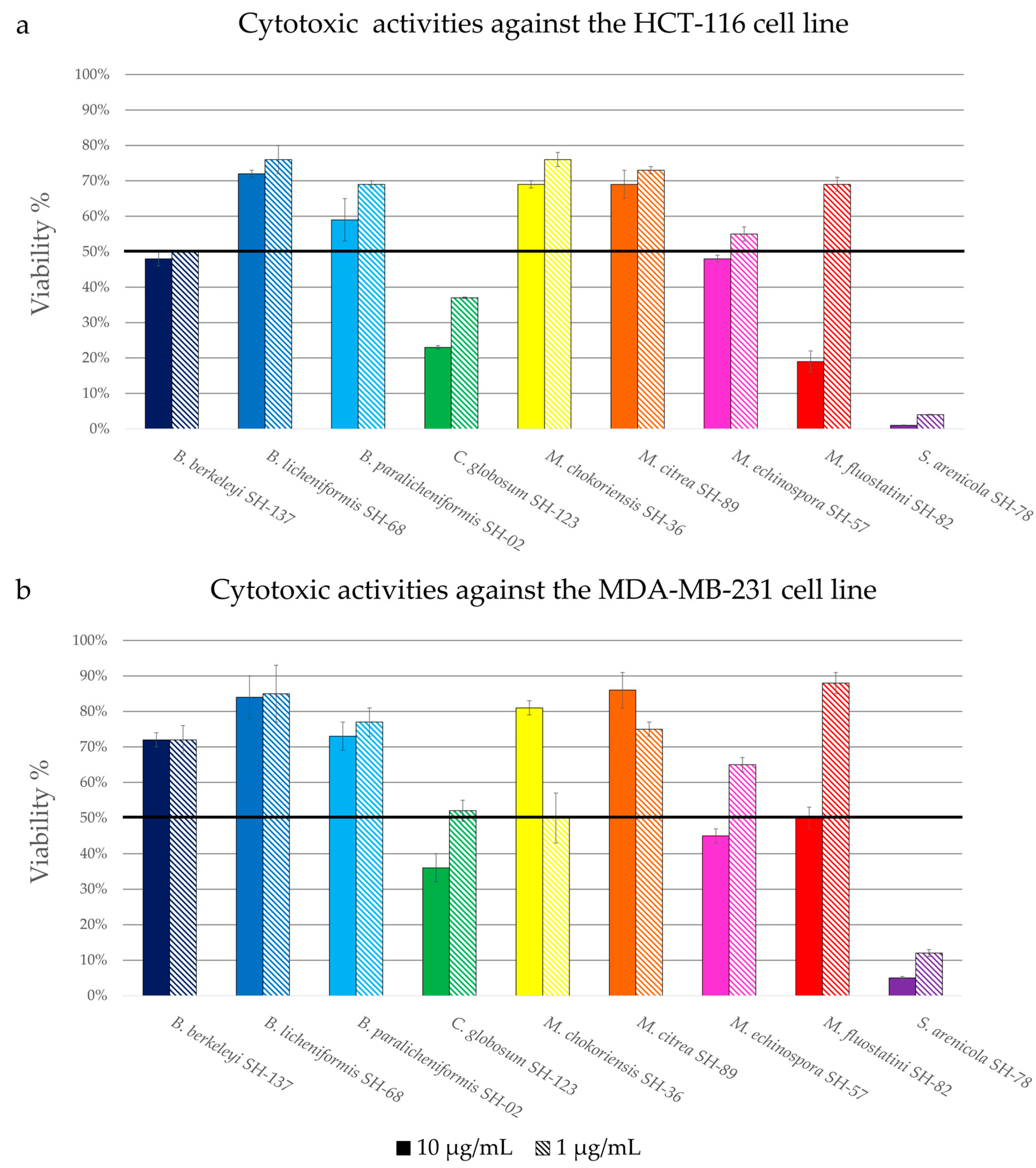
3.2.1. Cytotoxic Activity
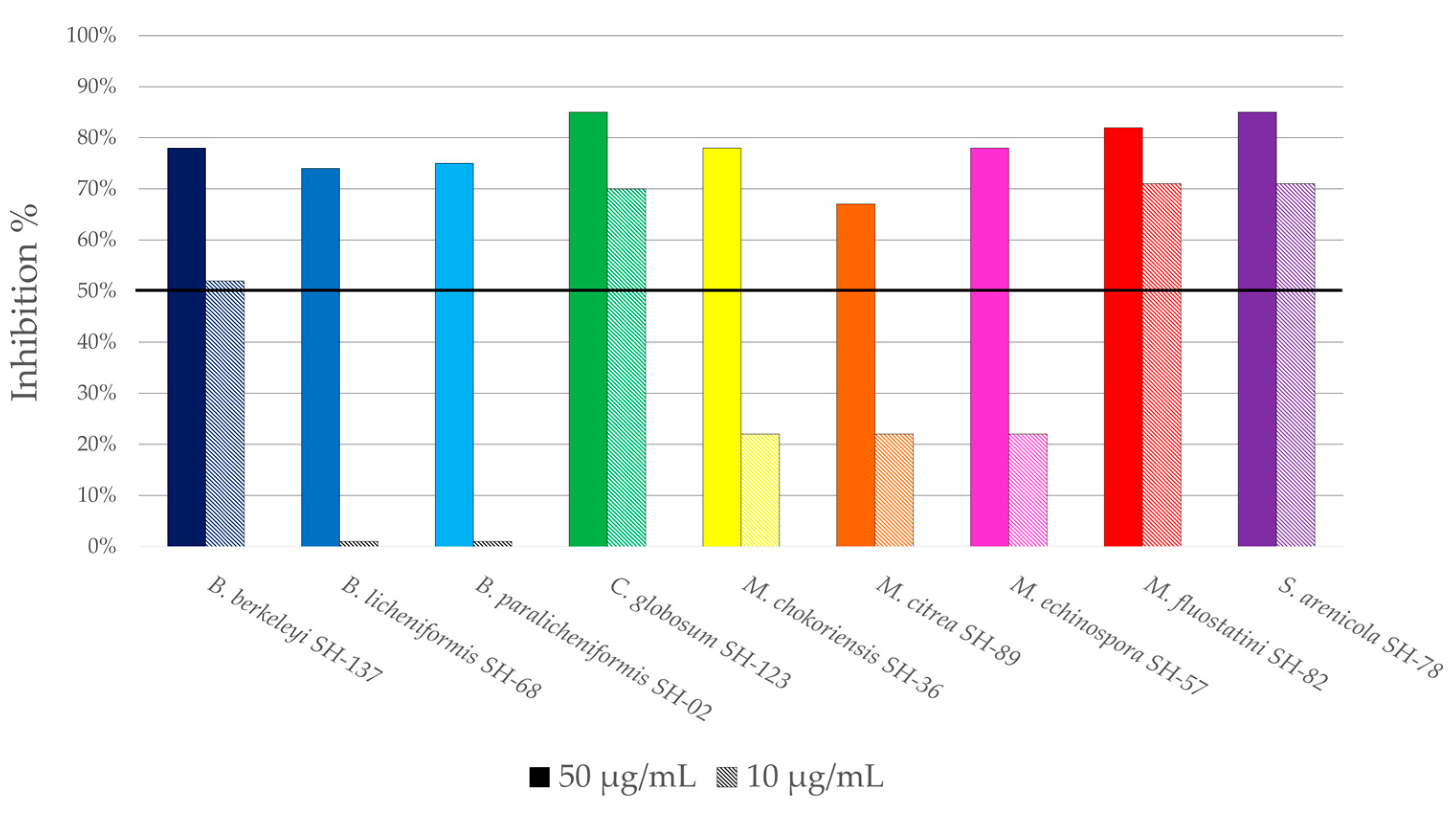
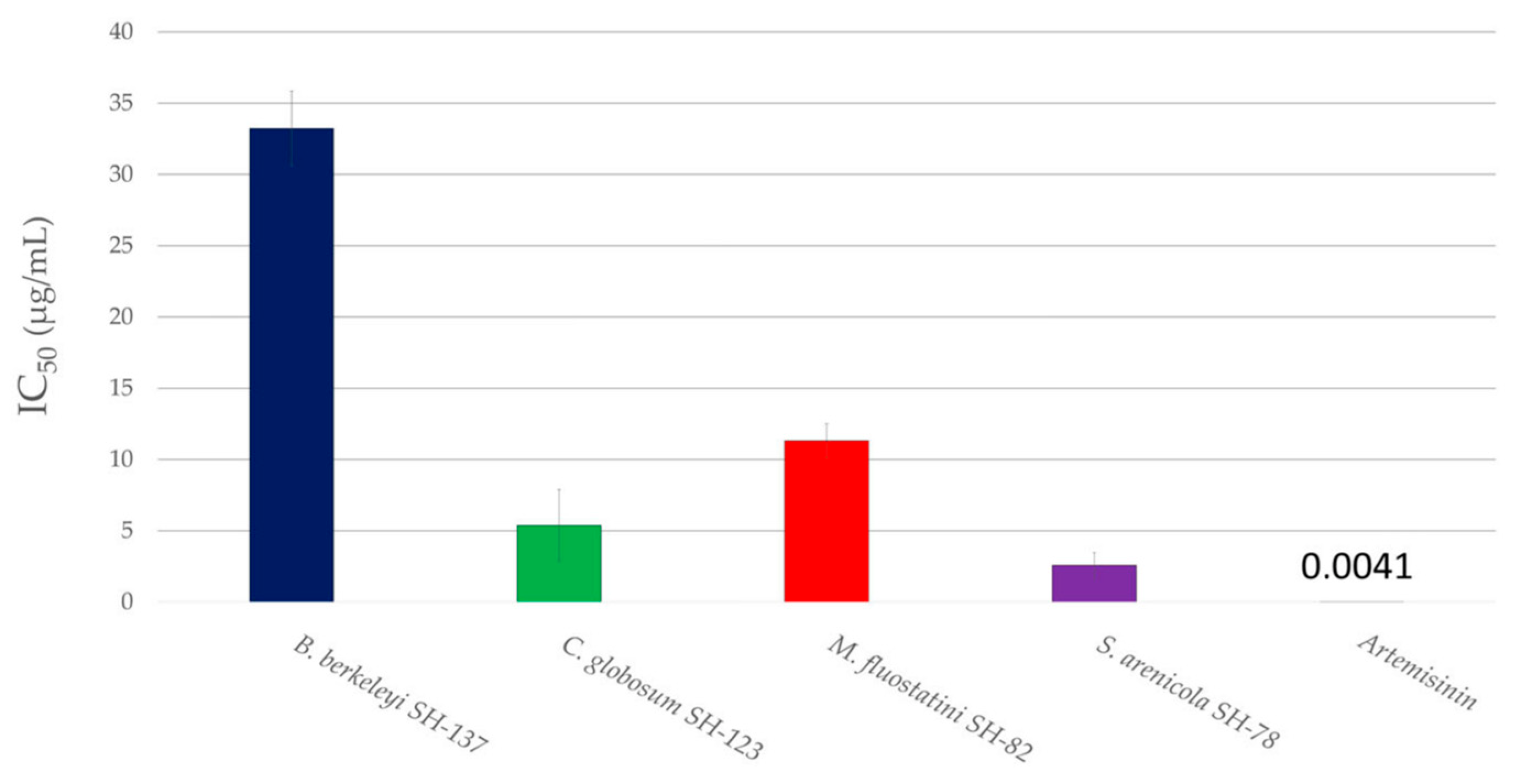
3.2.2. Antiplasmodial Activity
3.3. Strain Selection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization Website. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 16 November 2022).
- World Health Organization Website. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 16 November 2022).
- World Health Organization Website. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 16 November 2022).
- IARC. IARC Biennial Report 2020-2021; IARC Scientific Publications: Lyon, France, 1976; ISBN 978-92-832-1106-8. [Google Scholar]
- World Health Organization World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021; ISBN 978-92-4-004049-6.
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef]
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic Resistance and Persistence-Implications for Human Health and Treatment Perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef] [PubMed]
- Global Cancer Observatory, WHO Website. Available online: https://gco.iarc.fr/tomorrow/en (accessed on 16 November 2022).
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R.H. Natural Product Discovery: Past, Present, and Future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.-Y.; Li, H.-J.; Li, Q.-Y.; Wu, Y.-C. Application of Marine Natural Products in Drug Research. Bioorganic Med. Chem. 2021, 35, 116058. [Google Scholar] [CrossRef] [PubMed]
- Jaspars, M.; Pascale, D.D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The marine biodiscovery pipeline and ocean medicines of tomorrow. J. Mar. Biol. Assoc. U. K. 2016, 96, 151–158. [Google Scholar] [CrossRef]
- Haefner, B. Drugs from the Deep: Marine Natural Products as Drug Candidates. Drug Discov. Today 2003, 8, 536–544. [Google Scholar] [CrossRef]
- Taufa, T.; Subramani, R.; Northcote, P.T.; Keyzers, R.A. Natural Products from Tongan Marine Organisms. Molecules 2021, 26, 4534. [Google Scholar] [CrossRef]
- Bai, X.; Liu, Y.; Wang, H.; Zhang, H. Natural Products from the Marine Sponge Subgenus Reniera. Molecules 2021, 26, 1097. [Google Scholar] [CrossRef]
- Morrow, C.; Cárdenas, P. Proposal for a Revised Classification of the Demospongiae (Porifera). Front. Zool. 2015, 12, 7. [Google Scholar] [CrossRef]
- Helber, S.; de Voogd, N.; Muhando, C.; Rohde, S.; Schupp, P. Anti-Predatory Effects of Organic Extracts of 10 Common Reef Sponges from Zanzibar. Hydrobiologia 2017, 790. [Google Scholar] [CrossRef]
- Saïd Hassane, C.; Herbette, G.; Garayev, E.; Mabrouki, F.; Clerc, P.; de Voogd, N.J.; Greff, S.; Trougakos, I.P.; Ouazzani, J.; Fouillaud, M.; et al. New Metabolites from the Marine Sponge Scopalina hapalia Collected in Mayotte Lagoon. Mar. Drugs 2022, 20, 186. [Google Scholar] [CrossRef]
- Helber, S.B.; Steinert, G.; Wu, Y.-C.; Rohde, S.; Hentschel, U.; Muhando, C.A.; Schupp, P.J. Sponges from Zanzibar Host Diverse Prokaryotic Communities with Potential for Natural Product Synthesis. FEMS Microbiol. Ecol. 2019, 95, fiz026. [Google Scholar] [CrossRef]
- Helber, S.B.; Hoeijmakers, D.J.J.; Muhando, C.A.; Rohde, S.; Schupp, P.J. Sponge Chemical Defenses Are a Possible Mechanism for Increasing Sponge Abundance on Reefs in Zanzibar. PLoS ONE 2018, 13, e0197617. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, C.; Tulasi, B.R.; Raju, M.; Thakur, N.; Dufossé, L. Marine Natural Products from Tunicates and Their Associated Microbes. Mar. Drugs 2021, 19, 308. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, X.-N.; Bi, H.-Y.; Wang, G.-Y. Antimicrobial Biosynthetic Potential and Phylogenetic Analysis of Culturable Bacteria Associated with the Sponge Ophlitaspongia sp. from the Yellow Sea, China. Mar. Drugs 2022, 20, 588. [Google Scholar] [CrossRef] [PubMed]
- Said Hassane, C.; Fouillaud, M.; Le Goff, G.; Sklirou, A.D.; Boyer, J.B.; Trougakos, I.P.; Jerabek, M.; Bignon, J.; de Voogd, N.J.; Ouazzani, J.; et al. Microorganisms Associated with the Marine Sponge Scopalina hapalia: A Reservoir of Bioactive Molecules to Slow Down the Aging Process. Microorganisms 2020, 8, 1262. [Google Scholar] [CrossRef]
- Cheng, C.; MacIntyre, L.; Abdelmohsen, U.R.; Horn, H.; Polymenakou, P.N.; Edrada-Ebel, R.; Hentschel, U. Biodiversity, Anti-Trypanosomal Activity Screening, and Metabolomic Profiling of Actinomycetes Isolated from Mediterranean Sponges. PLoS ONE 2015, 10, e0138528. [Google Scholar] [CrossRef]
- Santos, J.D.; Vitorino, I.; De la Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Pérez-Moreno, G.; Gonzalez Martinez, I.; Tormo, J.R.; Martín, J.M.; et al. Bioactivities and Extract Dereplication of Actinomycetales Isolated from Marine Sponges. Front. Microbiol. 2019, 10, 727. [Google Scholar] [CrossRef] [PubMed]
- Siro, G.; Pipite, A.; Christi, K.; Srinivasan, S.; Subramani, R. Marine Actinomycetes Associated with Stony Corals: A Potential Hotspot for Specialized Metabolites. Microorganisms 2022, 10, 1349. [Google Scholar] [CrossRef]
- Valliappan, K.; Sun, W.; Li, Z. Marine Actinobacteria Associated with Marine Organisms and Their Potentials in Producing Pharmaceutical Natural Products. Appl. Microbiol. Biotechnol. 2014, 98, 7365–7377. [Google Scholar] [CrossRef]
- Jose, P.A.; Maharshi, A.; Jha, B. Actinobacteria in Natural Products Research: Progress and Prospects. Microbiol. Res. 2021, 246, 126708. [Google Scholar] [CrossRef]
- Al-shaibani, M.M.; Radin Mohamed, R.M.S.; Sidik, N.M.; Enshasy, H.A.E.; Al-Gheethi, A.; Noman, E.; Al-Mekhlafi, N.A.; Zin, N.M. Biodiversity of Secondary Metabolites Compounds Isolated from Phylum Actinobacteria and Its Therapeutic Applications. Molecules 2021, 26, 4504. [Google Scholar] [CrossRef]
- Leal, M.C.; Sheridan, C.; Osinga, R.; Dionísio, G.; Rocha, R.J.M.; Silva, B.; Rosa, R.; Calado, R. Marine Microorganism-Invertebrate Assemblages: Perspectives to Solve the “Supply Problem” in the Initial Steps of Drug Discovery. Mar. Drugs 2014, 12, 3929–3952. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T.; International Natural Product Sciences Taskforce. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Kalkreuter, E.; Pan, G.; Cepeda, A.J.; Shen, B. Targeting Bacterial Genomes for Natural Product Discovery. Trends Pharmacol. Sci. 2020, 41, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Steele, A.D.; Teijaro, C.N.; Yang, D.; Shen, B. Leveraging a Large Microbial Strain Collection for Natural Product Discovery. J. Biol. Chem. 2019, 294, 16567–16576. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.C.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A Computational Framework to Explore Large-Scale Biosynthetic Diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]
- Yan, X.; Ge, H.; Huang, T.; Hindra; Yang, D.; Teng, Q.; Crnovčić, I.; Li, X.; Rudolf, J.D.; Lohman, J.R.; et al. Strain Prioritization and Genome Mining for Enediyne Natural Products. mBio 2016, 7, e02104-16. [Google Scholar] [CrossRef] [PubMed]
- Hindra; Huang, T.; Yang, D.; Rudolf, J.D.; Xie, P.; Xie, G.; Teng, Q.; Lohman, J.R.; Zhu, X.; Huang, Y.; et al. Strain Prioritization for Natural Product Discovery by a High-Throughput Real-Time PCR Method. J. Nat. Prod. 2014, 77, 2296–2303. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kirchmair, J. Cheminformatics in Natural Product-Based Drug Discovery. Mol. Inform. 2020, 39, e2000171. [Google Scholar] [CrossRef]
- Macintyre, L.; Zhang, T.; Viegelmann, C.; Martinez, I.J.; Cheng, C.; Dowdells, C.; Abdelmohsen, U.R.; Gernert, C.; Hentschel, U.; Edrada-Ebel, R. Metabolomic Tools for Secondary Metabolite Discovery from Marine Microbial Symbionts. Mar. Drugs 2014, 12, 3416–3448. [Google Scholar] [CrossRef]
- Clarke, C.J.; Haselden, J.N. Metabolic Profiling as a Tool for Understanding Mechanisms of Toxicity. Toxicol. Pathol. 2008, 36, 140–147. [Google Scholar] [CrossRef]
- Hou, Y.; Braun, D.R.; Michel, C.R.; Klassen, J.L.; Adnani, N.; Wyche, T.P.; Bugni, T.S. Microbial Strain Prioritization Using Metabolomics Tools for the Discovery of Natural Products. Anal. Chem. 2012, 84, 4277–4283. [Google Scholar] [CrossRef]
- Soldatou, S.; Eldjárn, G.H.; Ramsay, A.; van der Hooft, J.J.J.; Hughes, A.H.; Rogers, S.; Duncan, K.R. Comparative Metabologenomics Analysis of Polar Actinomycetes. Mar. Drugs 2021, 19, 103. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.R.; Mouillon, J.-M.; Pohl, S.; Arnau, J. Secondary Metabolite Production and the Safety of Industrially Important Members of the Bacillus subtilis Group. FEMS Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.C.; Wilke, D.V.; Ferreira, E.G.; Takeara, R.; de Moraes, M.O.; da Silveira, E.R.; da Cruz Lotufo, T.M.; Lopes, N.P.; Costa-Lotufo, L.V. Structure Elucidation and Anticancer Activity of 7-Oxostaurosporine Derivatives from the Brazilian Endemic Tunicate Eudistoma vannamei. Mar. Drugs 2012, 10, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Diao, A. Marizomib, A Potent Second Generation Proteasome Inhibitor from Natural Origin. ACAMC 2015, 15, 298–306. [Google Scholar] [CrossRef]
- Goodman, C.; Useglio, M.; Peirú, S.; Labadie, G.; Mcfadden, G.; Rodriguez, E.; Gramajo, H. Chemobiosynthesis of New Antimalarial Macrolides. Antimicrob. Agents Chemother. 2012, 57, 907–913. [Google Scholar] [CrossRef]
- Schmid, R.; Petras, D.; Nothias, L.-F.; Wang, M.; Aron, A.T.; Jagels, A.; Tsugawa, H.; Rainer, J.; Garcia-Aloy, M.; Dührkop, K.; et al. Ion Identity Molecular Networking for Mass Spectrometry-Based Metabolomics in the GNPS Environment. Nat. Commun. 2021, 12, 3832. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A Rapid Tool for Turning Tandem Mass Spectra into Metabolite Structure Information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching Molecular Structure Databases with Tandem Mass Spectra Using CSI: FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef] [PubMed]
- Rutz, A.; Dounoue-Kubo, M.; Ollivier, S.; Bisson, J.; Bagheri, M.; Saesong, T.; Ebrahimi, S.N.; Ingkaninan, K.; Wolfender, J.-L.; Allard, P.-M. Taxonomically Informed Scoring Enhances Confidence in Natural Products Annotation. Front. Plant Sci. 2019, 10, 1329. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. mZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based Molecular Networking in the GNPS Analysis Environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Dührkop, K.; Nothias, L.-F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C.; et al. Systematic Classification of Unknown Metabolites Using High-Resolution Fragmentation Mass Spectra. Nat. Biotechnol. 2021, 39, 462–471. [Google Scholar] [CrossRef]
- Rutz, A.; Sorokina, M.; Galgonek, J.; Mietchen, D.; Willighagen, E.; Gaudry, A.; Graham, J.G.; Stephan, R.; Page, R.; Vondrášek, J.; et al. The LOTUS Initiative for Open Knowledge Management in Natural Products Research. eLife 2022, 11, e70780. [Google Scholar] [CrossRef]
- da Silva, R.R.; Wang, M.; Nothias, L.-F.; van der Hooft, J.J.J.; Caraballo-Rodríguez, A.M.; Fox, E.; Balunas, M.J.; Klassen, J.L.; Lopes, N.P.; Dorrestein, P.C. Propagating Annotations of Molecular Networks Using in Silico Fragmentation. PLoS Comput. Biol. 2018, 14, e1006089. [Google Scholar] [CrossRef]
- Ernst, M.; Kang, K.B.; Caraballo-Rodríguez, A.M.; Nothias, L.-F.; Wandy, J.; Chen, C.; Wang, M.; Rogers, S.; Medema, M.H.; Dorrestein, P.C.; et al. MolNetEnhancer: Enhanced Molecular Networks by Integrating Metabolome Mining and Annotation Tools. Metabolites 2019, 9, 144. [Google Scholar] [CrossRef] [PubMed]
- Djoumbou Feunang, Y.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated Chemical Classification with a Comprehensive, Computable Taxonomy. J. Cheminformatics 2016, 8, 61. [Google Scholar] [CrossRef]
- Trager, W.; Jensen, J.B. Human Malaria Parasites in Continuous Culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Ledoux, A.; Mamede, L.; Palazzo, C.; Furst, T.; Jansen, O.; De Tullio, P.; Kagisha, V.; Pendeville, H.; Fillet, M.; Piel, G.; et al. Heparin-Coated Liposomes Improve Antiplasmodial Activity and Reduce the Toxicity of Poupartone B. Planta Med. Int. Open 2020, 07, e73–e80. [Google Scholar] [CrossRef]
- Rajput, A.; Dominguez San Martin, I.; Rose, R.; Beko, A.; LeVea, C.; Sharratt, E.; Mazurchuk, R.; Hoffman, R.M.; Brattain, M.G.; Wang, J. Characterization of HCT116 Human Colon Cancer Cells in an Orthotopic Model. J. Surg. Res. 2008, 147, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J. Chapter 40—Animal Models for Studying Prevention and Treatment of Breast Cancer. In Animal Models for the Study of Human Disease; Conn, P.M., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 997–1018. ISBN 978-0-12-415894-8. [Google Scholar]
- Dictonary of Natural Products Website. Available online: https://dnp.chemnetbase.com/ (accessed on 11 November 2022).
- NPatlas Website. Available online: https://www.npatlas.org/ (accessed on 11 November 2022).
- Le Goff, G.; Lopes, P.; Arcile, G.; Vlachou, P.; Van Elslande, E.; Retailleau, P.; Gallard, J.-F.; Weis, M.; Benayahu, Y.; Fokialakis, N.; et al. Impact of the Cultivation Technique on the Production of Secondary Metabolites by Chrysosporium lobatum TM-237-S5, Isolated from the Sponge Acanthella Cavernosa. Mar. Drugs 2019, 17, 678. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Guo, Q.; Yu, W.; Zhang, Y.; He, B. Bioactivities and Future Perspectives of Chaetoglobosins. Evid. Based Complement Altern. Med. 2020, 2020, 8574084. [Google Scholar] [CrossRef]
- Li, H.; Xiao, J.; Gao, Y.-Q.; Tang, J.; Zhang, A.-L.; Gao, J.-M. Chaetoglobosins from Chaetomium globosum, an Endophytic Fungus in Ginkgo biloba, and Their Phytotoxic and Cytotoxic Activities. J. Agric. Food Chem. 2014, 62, 3734–3741. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Moore, B.S.; Fenical, W. The Marine Actinomycete Genus Salinispora: A Model Organism for Secondary Metabolite Discovery. Nat. Prod. Rep. 2015, 32, 738–751. [Google Scholar] [CrossRef]
- METACYC Website. Available online: https://metacyc.org/ (accessed on 15 January 2023).
- Stratmann, A.; Schupp, T.; Toupet, C.; Schilling, W.; Oberer, L.; Traber, R. New Insights into Rifamycin B Biosynthesis: Isolation of Proansamycin B and 34a-Deoxy-Rifamycin W as Early Macrocyclic Intermediates Indicating Two Separated Biosynthetic Pathways. J. Antibiot. 2002, 55, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.J.; Wagman, G.H.; Marquez, J.A.; Testa, R.T.; Oden, E.; Waitz, J.A. Megalomicin, a New Macrolide Antibiotic Complex Produced by Micromonospora. J. Antibiot. 1969, 22, 253–258. [Google Scholar] [CrossRef]
- Useglio, M.; Peirú, S.; Rodríguez, E.; Labadie, G.R.; Carney, J.R.; Gramajo, H. TDP-l-Megosamine Biosynthesis Pathway Elucidation and Megalomicin A Production in Escherichia coli. Appl. Environ. Microbiol. 2010, 76, 3869. [Google Scholar] [CrossRef]
- Volchegursky, Y.; Hu, Z.; Katz, L.; McDaniel, R. Biosynthesis of the Anti-Parasitic Agent Megalomicin: Transformation of Erythromycin to Megalomicin in Saccharopolyspora erythraea. Mol. Microbiol. 2000, 37, 752–762. [Google Scholar] [CrossRef]
- Perley, J.E.; Stowe, B.B. The Production of Tryptamine from Tryptophan by Bacillus cereus (KVT). Biochem. J. 1966, 100, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, H.; Li, W.; Zhu, X.; Ding, W.; Li, C. Bioactive Chaetoglobosins from the Mangrove Endophytic Fungus Penicillium chrysogenum. Mar. Drugs 2016, 14, 172. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Li, H.; Xu, M.; Li, T.; Wang, J.; Sun, C.; Hong, K.; Li, W. Staurosporine Derivatives Generated by Pathway Engineering in a Heterologous Host and Their Cytotoxic Selectivity. J. Nat. Prod. 2018, 81, 1745–1751. [Google Scholar] [CrossRef]
- Renner, M.K.; Shen, Y.-C.; Cheng, X.-C.; Jensen, P.R.; Frankmoelle, W.; Kauffman, C.A.; Fenical, W.; Lobkovsky, E.; Clardy, J. Cyclomarins A−C, New Antiinflammatory Cyclic Peptides Produced by a Marine Bacterium ( Streptomyces sp.). J. Am. Chem. Soc. 1999, 121, 11273–11276. [Google Scholar] [CrossRef]
- Schulze, C.J.; Navarro, G.; Ebert, D.; DeRisi, J.; Linington, R.G. Salinipostins A–K, Long-Chain Bicyclic Phosphotriesters as a Potent and Selective Antimalarial Chemotype. J. Org. Chem. 2015, 80, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Quiros-Guerrero, L.-M.; Nothias, L.-F.; Gaudry, A.; Marcourt, L.; Allard, P.-M.; Rutz, A.; David, B.; Queiroz, E.F.; Wolfender, J.-L. Inventa: A Computational Tool to Discover Structural Novelty in Natural Extracts Libraries. Front. Mol. Biosci. 2022, 9, 1028334. [Google Scholar] [CrossRef]
- Crüsemann, M.; O’Neill, E.C.; Larson, C.B.; Melnik, A.V.; Floros, D.J.; da Silva, R.R.; Jensen, P.R.; Dorrestein, P.C.; Moore, B.S. Prioritizing Natural Product Diversity in a Collection of 146 Bacterial Strains Based on Growth and Extraction Protocols. J. Nat. Prod. 2017, 80, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Huang, C.; Luo, Y. Recent Advances in Silent Gene Cluster Activation in Streptomyces. Front. Bioeng. Biotechnol. 2021, 9, 632230. [Google Scholar] [CrossRef]
- Liu, S.-W.; Jadambaa, N.; Nikandrova, A.A.; Osterman, I.A.; Sun, C.-H. Exploring the Diversity and Antibacterial Potentiality of Cultivable Actinobacteria from the Soil of the Saxaul Forest in Southern Gobi Desert in Mongolia. Microorganisms 2022, 10, 989. [Google Scholar] [CrossRef] [PubMed]
- Ortlieb, N.; Klenk, E.; Kulik, A.; Niedermeyer, T.H.J. Development of an Agar-Plug Cultivation System for Bioactivity Assays of Actinomycete Strain Collections. PLoS ONE 2021, 16, e0258934. [Google Scholar] [CrossRef] [PubMed]
- Crater, J.S.; Lievense, J.C. Scale-up of Industrial Microbial Processes. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Species | Code | Selected Regions for Genetic Characterization |
|---|---|---|---|
| Bacilli | Bacillus berkeleyi | SH-137 | |
| Bacillus paralicheniformis | SH-02a | ADNr 16s (from V1 to V5) | |
| Bacillus licheniformis | SH-68 | ||
| Sordariomycete | Chaetomium globosum | SH-123 | ITS/BenA |
| Micromonospora chokoriensis | SH-36 | ||
| Micromonospora citrea | SH-89 | ||
| Actinobacteria | Micromonospora echinospora | SH-57 | ADNr 16s (from V1 to V5) |
| Micromonospora fluostatini | SH-82 | ||
| Salinispora arenicola | SH-78 |
| Microbial Strain | Visible Peaks (Height > 10 pA) | Major Peaks (Height > 40 pA) |
|---|---|---|
| Bacillus berkeleyi SH-137 | 6 | 2 |
| Bacillus paralicheniformis SH-02a | 6 | 3 |
| Bacillus licheniformis SH-68 | 4 | 2 |
| Chaetomium globosum SH-123 | 10 | 3 |
| Micromonospora chokoriensis SH-36 | 0 | 0 |
| Micromonospora citrea SH-89 | 1 | 0 |
| Micromonospora echinospora SH-57 | 1 | 0 |
| Micromonospora fluostatini SH-82 | 17 | 8 |
| Salinispora arenicola SH-78 | 7 | 0 |
| Strain | Compound ID | Retention Time (min) | m/z [Adduct] | Molecular Formula (1) | Compound Name (1,2,3) | Superclass (a)/Natural Product Class (b) | Similarity Score (1,2,3) |
|---|---|---|---|---|---|---|---|
| Micromonospora fluostatini SH-82 | 1.1 | 6.14 | 877.5648 [M+H]+ | C44H80N2O15 | Megalomicin A (1,3) | Organic oxygen compounds(a)/ Erythromycins(b) | 92.2% (1)/0.22 (3) |
| 1.2 | 6.34 | 919.5754 [M+H]+ | C45H78N2O17 | Megalomicin B (1,3) | 88.61% (1)/0.47 (3) | ||
| 1.3 | 6.52 | 933.5939 [M+H]+ | C47H84N2O16 | 4′-Propionylmegalomicin A (1) | 89.01% (1) | ||
| 1.4 | 6.59 | 961.5883 [M+H]+ | C48H84N2O17 | Megalomicin C1 (1,3) | 87.19% (1)/0.47 (3) | ||
| 1.5 | 6.77 | 975.6046 [M+H]+ | C48H86N2O17 | Megalomicin C2 (1,3) | 83.55% (1)/0.40 (3) | ||
| 1.6 | 7.26 | 776.4797 [M+H]+ | C39H69NO14 | 2′-O-Acetylerythromycin A (1) | 83.58% (1) | ||
| 1.7 | 6.75 | 720.4529 [M+H]+ | C36H65NO13 | Erythromycin C (1,3) | 95.82% (1)/0.22 (3) | ||
| 1.8 | 7.08 | 704.4586 [M+H]+ | C36H65NO12 | Erythromycin D (1,3) | 93.53% (1)/0.22 (3) | ||
| 1.9 | 7.61 | 702.4425 [M-H2O+H]+ | C36H65NO13 | Erythromycin C (1) | 95.55% (1) | ||
| 1.10 | 7.36 | 702.4423 [M-H2O+H]+ | C36H65NO13 | 93.29% (1) | |||
| 1.11 | 7.13 | 702.442 [M-H2O+H]+ | C36H65NO13 | 94.40% (1) | |||
| 1.12 | 6.98 | 690.4431 [M+H]+ | C36H65NO12 | 7-Hydroxy-6-demethyl-6-deoxy-erythromycin D (1) | 83.46% (1) | ||
| 1.13 | 7.54 | 385.2577 [M-H2O+H]+ | C21H38O7 | Erythronolide B (1) | Phenylpropanoids and polyketides (a) | 61.07% (1) | |
| 1.14 | 7.30 | 371.2423 [M-H2O+H]+ | C20H36O7 | 2-Desmethyl-2-hydroxy-6-deoxyerythronolide B (1) | 68.90% (1) | ||
| 1.15 | 7.23 | 371.2423 [M-H2O+H]+ | 68.90% (1) | ||||
| 1.16 | 7.97 | 569.3287 [M+Na]+ | C28H50O10 | L-oleandrosyl-erythronolide B (1) | 75.17% (1) | ||
| 1.17 | 7.99 | 529.3367 [M-H2O+H]+ | 3-O-alpha-mycarosylerythronolide B (1) | 68.21% (1) | |||
| 1.18 | 7.71 | 555.3136 [M+Na]+ | C27H48O10 | L-olivosyl- erythronolide B (1) | 75.93% (1) | ||
| 1.19 | 515.3209 [M-H2O+H]+ | 75.87% (1) | |||||
| 1.20 | 8.25 | 369.2626 [M-H2O+H]+ | C21H38O6 | 6-Deoxyerythronolide B (1) | 63.47% (1) | ||
| 1.21 | 2.06 | 136.0618 [M+H]+ | C5H5N5 | Adenine (1,3) | Organoheterocyclic compounds (a)/Purine alkaloids (b) | 100% (1)/0.63 (3) | |
| Salinispora arenicola SH-78 | 2.1 | 7.84 | 396.2745 [M+H]+ | C22H37NO5 | Saliniketal A (1) | Lipids and lipid-like molecules (a)/Open-chain polyketides (b) | 49.50% (1) |
| 2.2 | 6.90 | 412.2693 [M+H]+ | C22H37NO6 | Saliniketal B (1,3) | 55.44% (1)/0.45 (3) | ||
| 2.3 | 7.20 | 483.2028 [M+H]+ | C28H26N4O4 | OH-staurosporine (2,3) | Organoheterocyclic compounds (a)/Carbazole alkaloids (b) | 0.79 (2)/0.30 (3) | |
| 2.4 | 7.33 | 467.2085 [M+H]+ | C28H26N4O3 | Staurosporine (1,2,3) | 98.11% (1)/0.96 (2)/0.30 (3) | ||
| 2.5 | 7.53 | 497.1825 [M+H]+ | C28H24N4O5 | 4′-Demethyl-Af-formyl-7V-hydroxy-staurosporine (1) | 81.75% (1) | ||
| 2.6 | 7.30 | 497.2192 [M+H]+ | C29H28N4O4 | 4′-N-methyl-5′-hydroxystaurosporine (1,3) | 62.56% (1)/0.28 (3) | ||
| 2.7 | 9.70 | 696.3022 [M+H]+ | C37H45NO12 | Rifamycin S (2,3) | Phenylpropanoids and polyketides (a) / Ansa macrolides (b) | 0.71 (2)/0.35 (3) | |
| 2.8 | 9.24 | 682.2866 [M+H]+ | C36H43NO12 | 1,4-Dioxo-35-nor-1,4-dide(hydroxy)rifamycin (1,3) | 74.69% (1)/0.39 (3) | ||
| 2.9 | 9.39 | 624.3166 [M+H]+ | C34H41NO10 | Proansamycin B (1) | 43.29% (1) | ||
| 2.10 | 8.30 | 624.3190 [M+H]+ | 64.34% (1) | ||||
| 2.11 | 9.75 | 640.3119 [M+H]+ | C35H45NO10 | 34a-deoxy-rifamycin W (1) | 54.25% (1) | ||
| 2.12 | 9.01 | 622.2666 [M-H2O+H]+ | C34H41NO11 | 25-O-deacetyl-27-O-demethylrifamycin S (1) | 52.72% (1) | ||
| 2.13 | 8.68 | 654.2918 [M+H]+ | C35H43NO11 | 25-deacetoxy-25-hydroxyrifamycin S (1) | 71.40% (1) | ||
| 2.14 | 6.84 | 176.0707 [M+H]+ | C10H9NO2 | 4-Hydroxy-1-methyl-2-quinolone (1,3) | Organoheterocyclic compounds (a) / Quinoline alkaloids (b) | 75.64% (1)/0.54 (3) | |
| Chaetomium globosum SH-123 | 3.1 | 10.4 | 483.3009 [M+H]+ | C32H38N2O2 | Prochaetoglobosin I (1,3) | Organoheterocyclic compounds (a)/Cytochalasan alkaloids (b) | 64.04% (1)/0.45 (3) |
| 3.2 | 10.27 | 497.2805 [M+H]+ | C32H36N2O3 | Prochaetoglobosin II (1,3) | 69.16% (1)/0.61 (3) | ||
| 3.3 | 9.94 | 513.2749 [M+H]+ | C32H36N2O4 | Prochaetoglobosin III (1,3) | 81.75% (1)/0.40 (3) | ||
| 3.4 | 9.74 | 499.2953 [M+H]+ | C32H38N2O3 | Prochaetoglobosin IV (1,3) | 80.73% (1)/0.56 (3) | ||
| 3.5 | 9.49 | 515.2902 [M-H2O+H]+ | C32H40N2O5 | Cytoglobosin G (1) | 86.58% (1) | ||
| 3.6 | 9.02 | 515.2903 [M+H]+ | C32H38N2O4 | Cytoglobosin D (1,3) | 77.72% (1)/0.61 (3) | ||
| 3.7 | 8.24 | 515.2911 [M+H]+ | C28H24N4O5 | 4′-demethyl-Af-formyl-7V-hydroxy-staurosporine (1) | 84.31% (1)/0.53 (3) | ||
| 3.8 | 8.97 | 517.3057 [M+H]+ | C32H40N2O4 | Armochaetoglobin I (1,3) | 79.76% (1)/0.61 (3) | ||
| 3.9 | 9.31 | 529.2693 [M+H]+ | C32H36N2O5 | Chaetoglobosin A (1,3) | 97.67% (1)/0.59 (3) | ||
| 3.10 | 8.62 | 529.2699 [M+H]+ | C32H36N2O5 | Chaetoglobosin C (1,2,3) | 91.01% (1)/0.80 (2)/0.55 (3) | ||
| 3.11 | 8.74 | 531.2851 [M+H]+ | C32H38N2O5 | 20-Dihydrochaetoglobosin A (1) | 94.02% (1) | ||
| 3.12 | 8.26 | 531.286 [M+H]+ | C32H38N2O5 | Cytoglobosin B (1) | 88.00% (1) | ||
| 3.13 | 8.67 | 545.2651 [M+H]+ | C32H36N2O6 | Armochaetoglobin J (1,3) | 84.42% (1)/0.59 (3) | ||
| 3.14 | 8.39 | 563.2752 [M+H]+ | C32H38N2O7 | Oxichaetoglobosin G (1) | 69.84% (1) | ||
| Bacillus licheniformis SH-68 / Bacillus paralicheniformis SH-02 | 4.1 | 10.89 | 385.3206 [M+H]+ | C25H40N2O | N-[2-(1H-indol-3-yl)ethyl]pentadecanamide (1,2) | Lipids and lipid-like molecules (a)/N-acyl amines (b) | 97.53% (1)/0.93 (2) |
| 4.2 | 11.27 | 413.3514 [M+H]+ | C27H44N2O | Heptadecanoic acid tryptamide (1,2) | 87.54% (1)/0.93 (2) | ||
| 4.3 | 10.74 | 371.3046 [M+H]+ | C24H38N2O | Myristoyl tryptamine (1,2) | Organoheterocyclic compounds (a)/Cytochalasan alkaloids (b) | 92.12% (1)/0.93 (2) | |
| 4.4 | 11.13 | 399.3365 [M+H]+ | C26H42N2O | N-palmitoyltryptamine (1,2) | 100% (1)/0.92 (2) | ||
| 4.5 | 2.07 | 161.1070 [M+H]+ | C10H12N2 | Tryptamine (1,2,3) | Organoheterocyclic compounds (a)/Simple indole alkaloids (b) | 100% (1)/0.77 (2)/0.38 (3) | |
| 4.6 | 8.02 | 245.1646 [M+H]+ | C15H20N2O | N-[2-(1H-indol-3-yl)ethyl]-2-methylbutanamide (2,3) | 87.03% (1)/0.15 (3) | ||
| 4.7 | 4.23 | 173.1072 [M+H]+ | C11H12N2 | Triptoline (1,2,3) | Organoheterocyclic compounds (a)/Carboline alkaloids (b) | 98.95% (1)/0.77 (2)/0.12 (3) | |
| Micromonospora chokoriensis SH-36 | 5.1 | 6.52 | 176.0703 [M+H]+ | C10H9NO2 | Indole-3-acetic acid (1,2) | Organoheterocyclic compounds (a)/Simple indole alkaloids (b | 100% (1)/0.94 (2) |
| Chemical score (CS) (1) | |||||||
|---|---|---|---|---|---|---|---|
| HPLC-DAD-CAD Analysis | Ion Identity Molecular Network (IIMN) Analysis | ||||||
| Number of Visible Peaks (VP) | Score VP | Number of Major Peaks (MP) | Score MP | Number of Total | Score TN | Number of Unique Nodes (UN) | Score UN |
| (CSVP) | (CSMP) | Nodes (TN) | (CSTN) | (CSUN) | |||
| 0–2 | 1 | 0–1 | 1 | 0–40 | 1 | 0–30 | 1 |
| 3–5 | 2 | 2–3 | 2 | 40–80 | 2 | 30–60 | 2 |
| 6–8 | 3 | 4–5 | 3 | 80–120 | 3 | 60–90 | 3 |
| 9–11 | 4 | 6–7 | 4 | 120–160 | 4 | 90–120 | 4 |
| >11 | 5 | >7 | 5 | >160 | 5 | >120 | 5 |
| Biological Score (BS) (2) | Novelty Score (NS) | ||||||
| Cytotoxic Activity | CA Score | Antiplasmodial Activity (AA) | AA Score | Average of Bibliographic Reference | Score (NS) | ||
| (CA) | (BSCA) | (BSAA) | |||||
| >80% | 5 | >70% | 5 | >50 | 0 | ||
| 80–50% | 4 | 70–50% | 4 | 30–50 | 1 | ||
| 50–40% | 3 | 50–40% | 3 | 10–30 | 2 | ||
| 40–30% | 2 | 40–30% | 2 | <10 | 3 | ||
| <30% | 1 | <30% | 1 | ||||
| >80% | 5 | >70% | 5 | ||||
| 80–50% | 4 | 70–50% | 4 | ||||
| 50–40% | 3 | 50–40% | 3 | ||||
| Strain | Biological Score | Chemical Score | Novelty Score | Selection Score | Ranking |
|---|---|---|---|---|---|
| Bacillus berkeleyi SH-137 | 6.5 | 5 | 3 | 6.3 | 4 |
| Bacillus paralicheniformis SH-02 | 2.5 | 6 | 3 | 5.0 | 5 |
| Bacillus licheniformis SH-68 | 1.5 | 5 | 1 | 3.3 | 8 |
| Chaetomium globosum SH-123 | 9 | 6 | 0 | 6.5 | 3 |
| Micromonospora chokoriensis SH-36 | 3.5 | 2 | 3 | 3.7 | 7 |
| Micromonospora citrea SH-89 | 1.5 | 5 | 3 | 4.6 | 6 |
| Micromonospora echinospora SH-57 | 3.5 | 2 | 1 | 3.3 | 8 |
| Micromonospora fluostatini SH-82 | 6.5 | 10 | 3 | 8.5 | 1 |
| Salinispora arenicola SH-78 | 10 | 7 | 1 | 7.8 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Loarer, A.; Marcellin-Gros, R.; Dufossé, L.; Bignon, J.; Frédérich, M.; Ledoux, A.; Queiroz, E.F.; Wolfender, J.-L.; Gauvin-Bialecki, A.; Fouillaud, M. Prioritization of Microorganisms Isolated from the Indian Ocean Sponge Scopalina hapalia Based on Metabolomic Diversity and Biological Activity for the Discovery of Natural Products. Microorganisms 2023, 11, 697. https://doi.org/10.3390/microorganisms11030697
Le Loarer A, Marcellin-Gros R, Dufossé L, Bignon J, Frédérich M, Ledoux A, Queiroz EF, Wolfender J-L, Gauvin-Bialecki A, Fouillaud M. Prioritization of Microorganisms Isolated from the Indian Ocean Sponge Scopalina hapalia Based on Metabolomic Diversity and Biological Activity for the Discovery of Natural Products. Microorganisms. 2023; 11(3):697. https://doi.org/10.3390/microorganisms11030697
Chicago/Turabian StyleLe Loarer, Alexandre, Rémy Marcellin-Gros, Laurent Dufossé, Jérôme Bignon, Michel Frédérich, Allison Ledoux, Emerson Ferreira Queiroz, Jean-Luc Wolfender, Anne Gauvin-Bialecki, and Mireille Fouillaud. 2023. "Prioritization of Microorganisms Isolated from the Indian Ocean Sponge Scopalina hapalia Based on Metabolomic Diversity and Biological Activity for the Discovery of Natural Products" Microorganisms 11, no. 3: 697. https://doi.org/10.3390/microorganisms11030697
APA StyleLe Loarer, A., Marcellin-Gros, R., Dufossé, L., Bignon, J., Frédérich, M., Ledoux, A., Queiroz, E. F., Wolfender, J. -L., Gauvin-Bialecki, A., & Fouillaud, M. (2023). Prioritization of Microorganisms Isolated from the Indian Ocean Sponge Scopalina hapalia Based on Metabolomic Diversity and Biological Activity for the Discovery of Natural Products. Microorganisms, 11(3), 697. https://doi.org/10.3390/microorganisms11030697