

The 4-α-Glucanotransferase AcbQ Is Involved in Acarbose Modification in Actinoplanes sp. SE50/110


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Construction of the Expression Systems
2.3. Preparation of Recombinant Proteins
2.4. Preparation of Substrates and Standards
2.5. In Vitro Reaction of AcbQ and Analysis
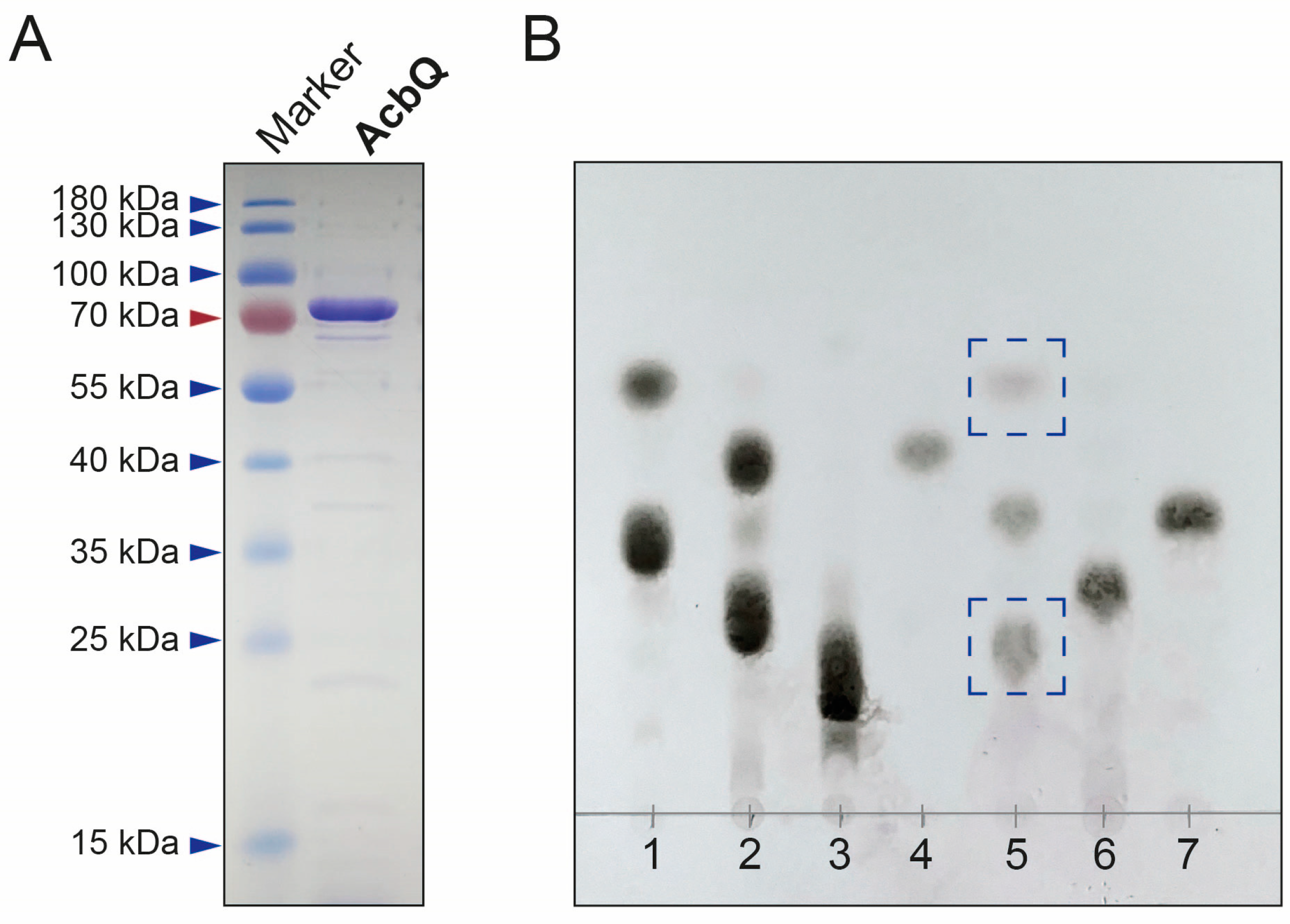
2.6. Thin-Layer Chromatography Analysis
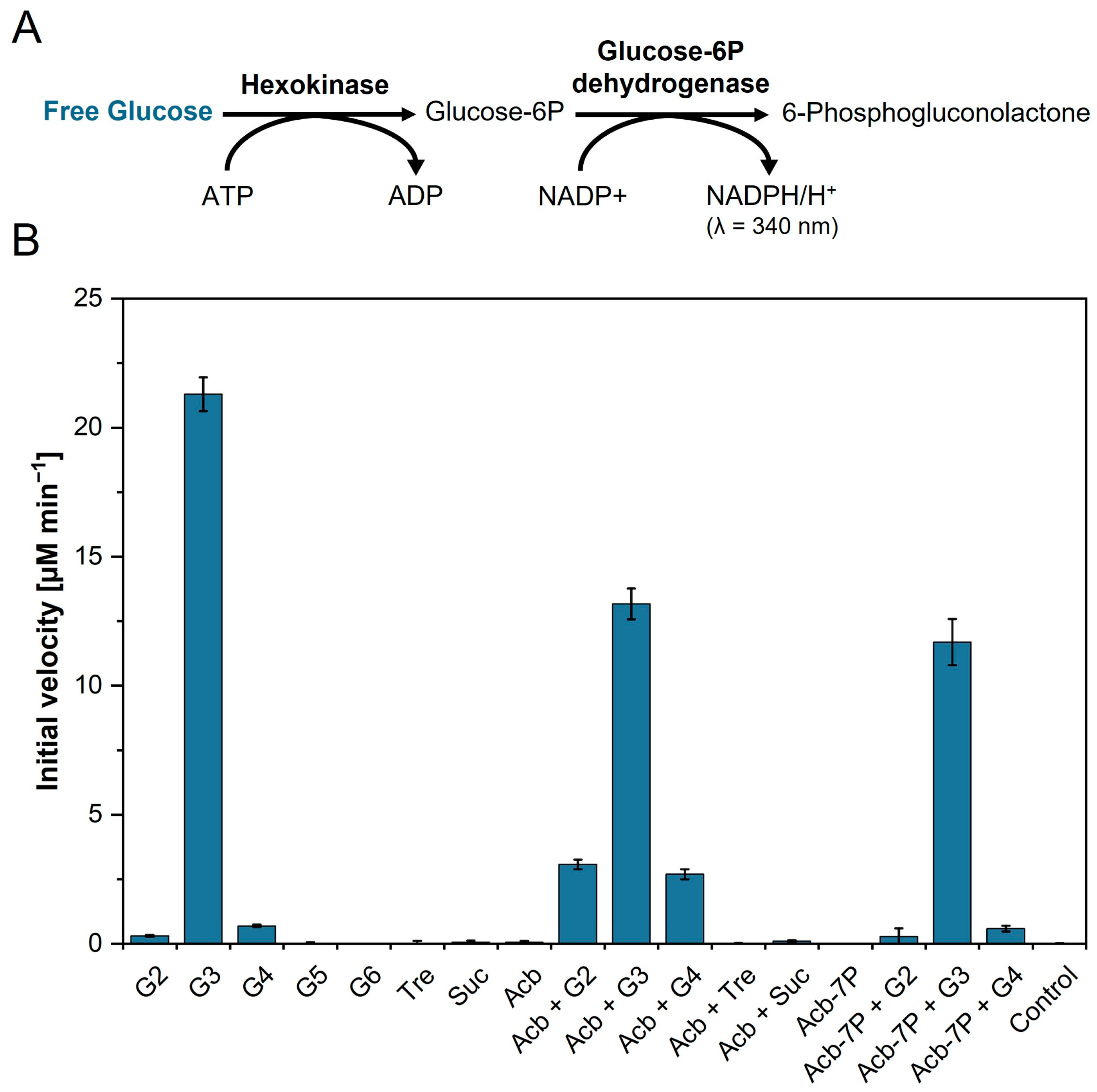
2.7. Initial Velocity of AcbQ with Different Substrates
2.8. HPAEC-PAD Analysis
2.9. LC-ESI-MS Analysis
3. Results
3.1. Simultaneous Assembly and Disassembly of Linear α-1,4-Glucans
3.2. Substrate Preference of the 4-α-Glucanotransferase AcbQ
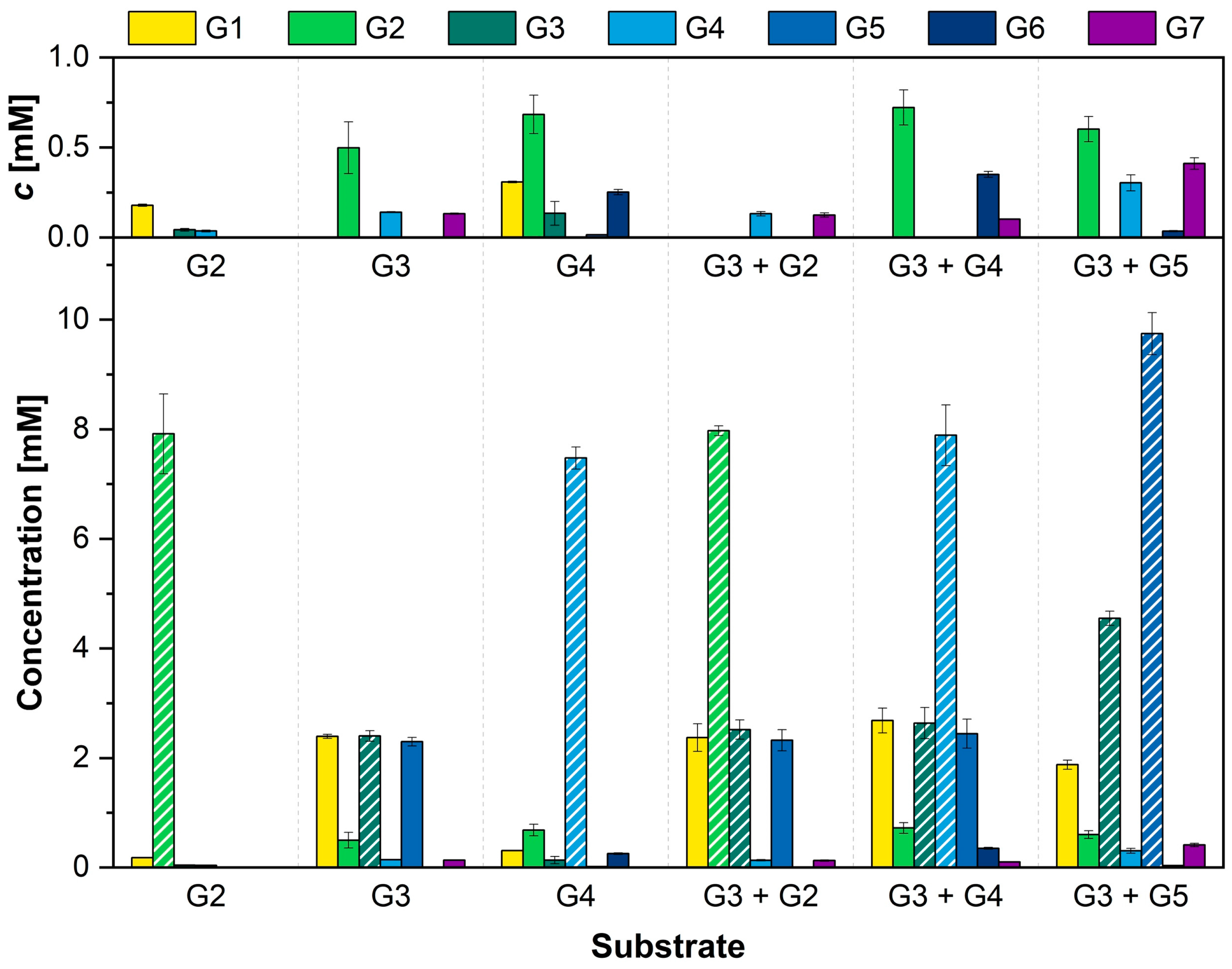
3.3. Combination of α-1,4-Glucans Results in Different Elongated Glucan Chains
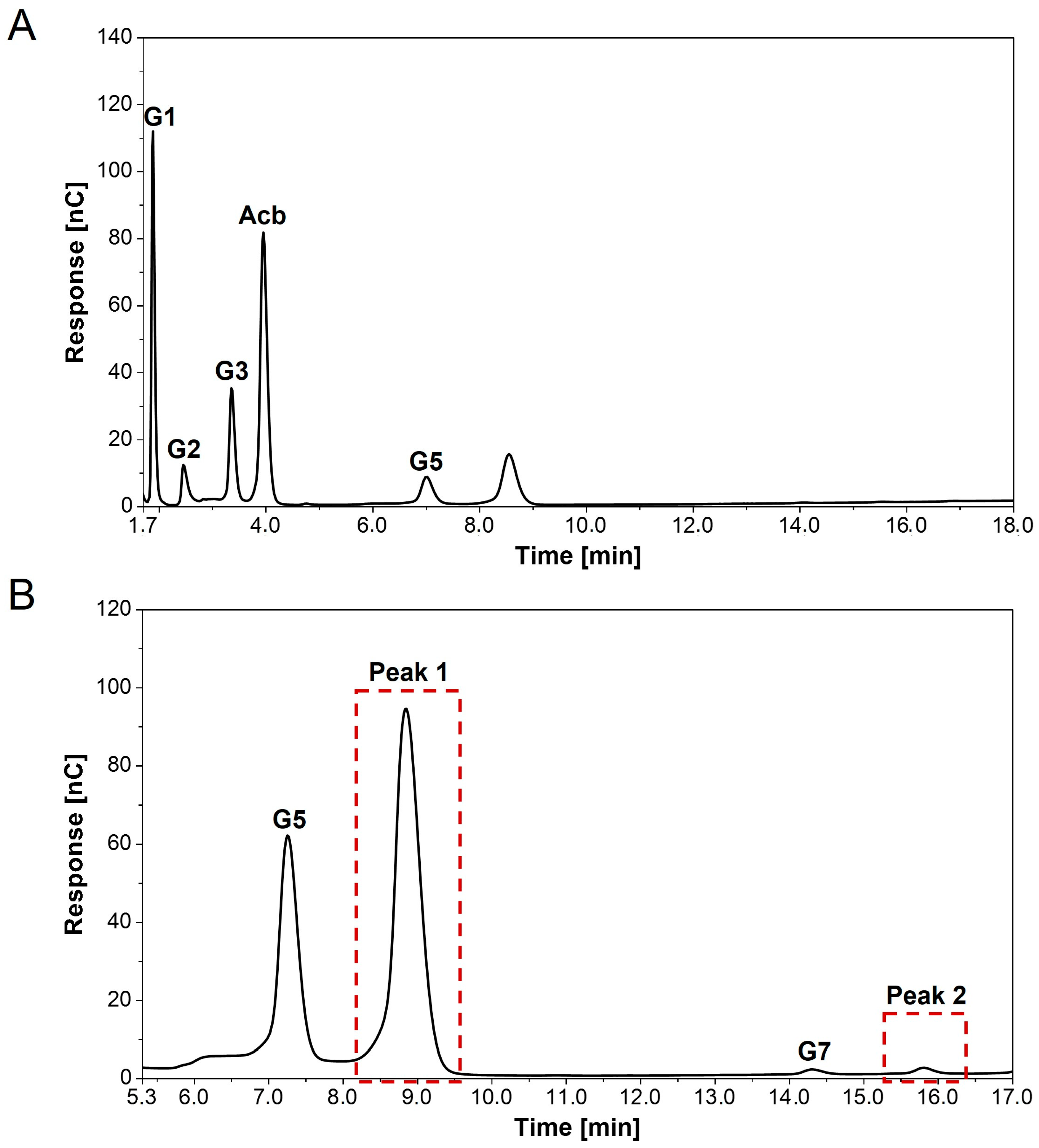
3.4. Detection of New Peaks in Assays with Acarbose and α-1,4-Glucans
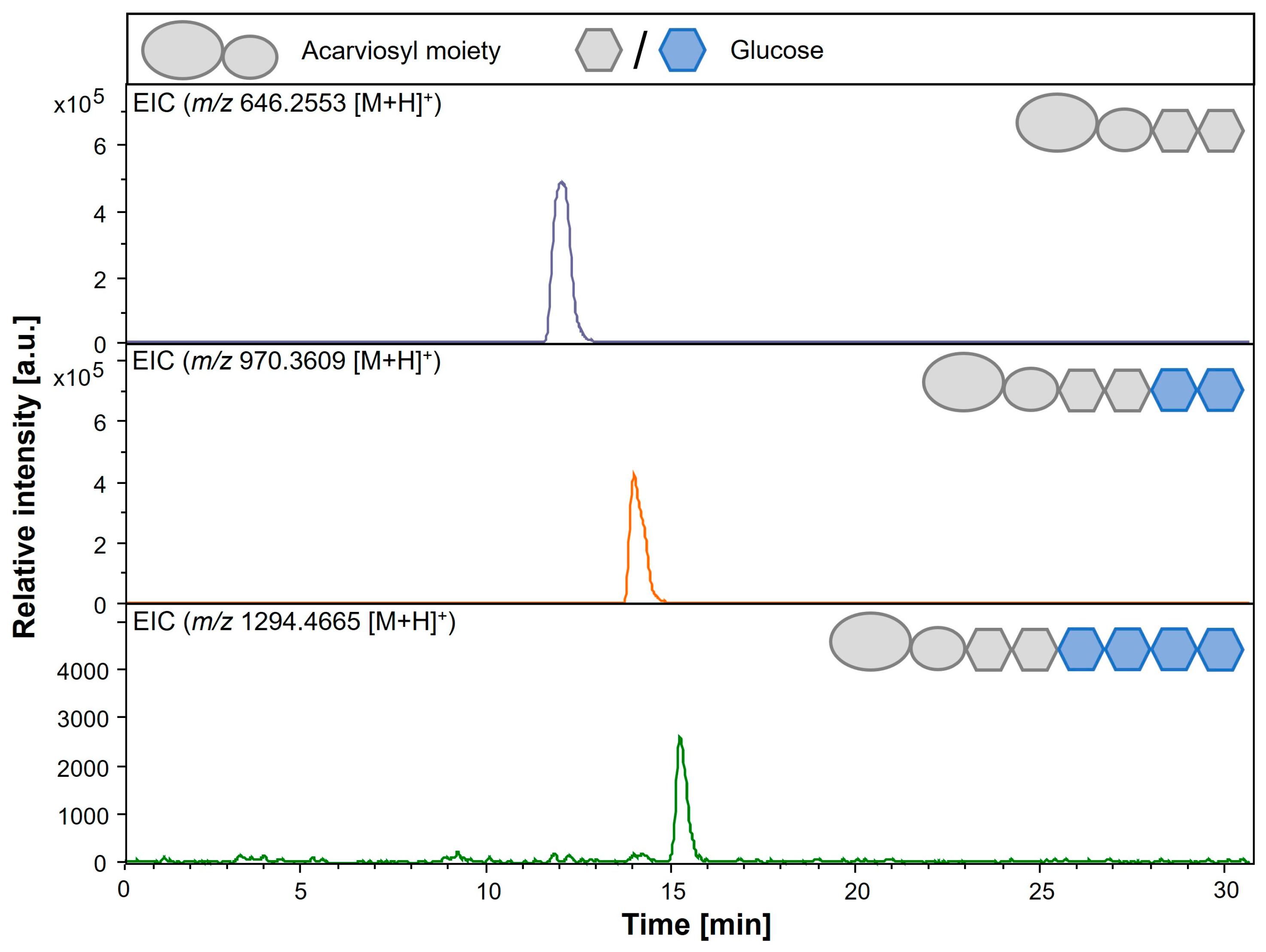
3.5. Detection of Elongated Acarviosyl Metabolites by LC-ESI-MS
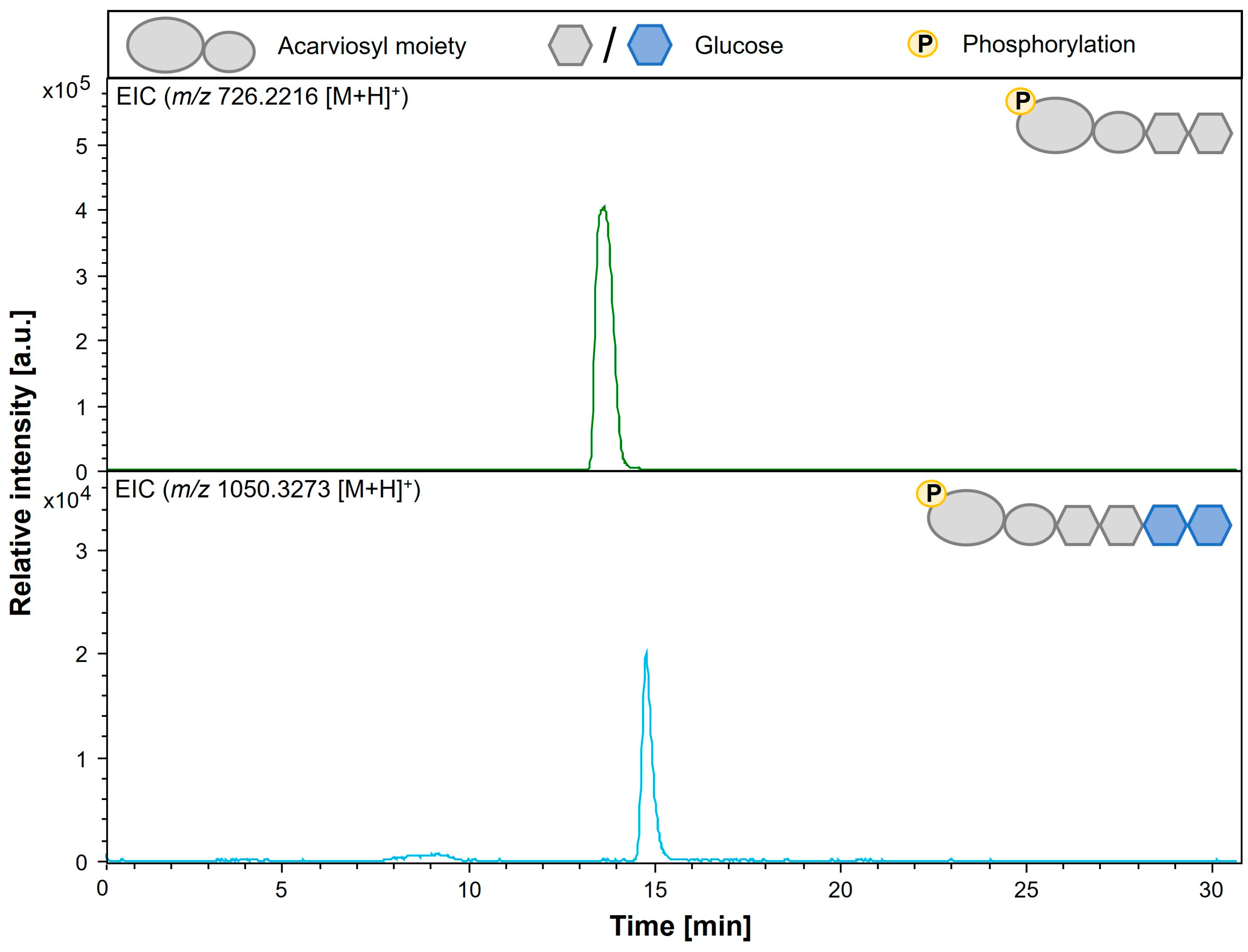
3.6. Analysis of Modification of Phosphorylated Acarbose by AcbQ
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas, 10th Ed. Available online: https://www.diabetesatlas.org/en/ (accessed on 9 January 2023).
- Wehmeier, U.F. Acarbose, ein therapeutisch eingesetzter Wirkstoff: Biosynthese und Funktion. BioSpektrum 2004, 10, 34–36. [Google Scholar]
- Laube, H. Acarbose: An update of its therapeutic use in diabetes treatment. Clin. Drug Investig. 2002, 22, 141–156. [Google Scholar] [CrossRef]
- Mahmud, T.; Lee, S.; Floss, H.G. The biosynthesis of acarbose and validamycin. Chem. Rec. 2001, 1, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, A.; Mahmud, T.; Lee, S.; Distler, J.; Floss, H.G.; Piepersberg, W. The AcbC protein from Actinoplanes species is a C7-cyclitol synthase related to 3-dehydroquinate synthases and is involved in the biosynthesis of the α-glucosidase inhibitor acarbose. J. Biol. Chem. 1999, 274, 10889–10896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.-S.; Podeschwa, M.; Altenbach, H.-J.; Piepersberg, W.; Wehmeier, U.F. The acarbose-biosynthetic enzyme AcbO from Actinoplanes sp. SE 50/110 is a 2-epi-5-epi-valiolone-7-phosphate 2-epimerase. FEBS Lett. 2003, 540, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-S.; Podeschwa, M.; Block, O.; Altenbach, H.-J.; Piepersberg, W.; Wehmeier, U.F. Identification of a 1-epi-valienol 7-kinase activity in the producer of acarbose, Actinoplanes sp. SE50/110. FEBS Lett. 2003, 540, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Luo, Y.; Zhang, X.; Kang, Q.; Zhang, D.; Zhang, L.; Bai, L.; Deng, Z. A severe leakage of intermediates to shunt products in acarbose biosynthesis. Nat. Commun. 2020, 11, 1468. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, T.; Samadi, A.; Burade, S.; Mahmud, T. Complete biosynthetic pathway to the antidiabetic drug acarbose. Nat. Commun. 2022, 13, 3455. [Google Scholar] [CrossRef]
- Zhang, C.-S.; Stratmann, A.; Block, O.; Brückner, R.; Podeschwa, M.; Altenbach, H.-J.; Wehmeier, U.F.; Piepersberg, W. Biosynthesis of the C7-cyclitol Moiety of Acarbose in Actinoplanes Species SE50/110. J. Biol. Chem. 2002, 277, 22853–22862. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Zhao, Q.; Jiang, M.; Kang, Q.; Bai, L. Biosynthetic pathway of deoxyaminosugar moiety in acarbose from Actinoplanes sp. SE50/110. Acta Microbiol. Sin. 2019, 60, 118–134. [Google Scholar] [CrossRef]
- Drepper, A.; Pape, H. Acarbose 7-phosphotransferase from Actinoplanes sp.: Purification, properties, and possible physiological function. J. Antibiot. 1996, 49, 664–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehmeier, U.F.; Piepersberg, W. Biotechnology and molecular biology of the α-glucosidase inhibitor acarbose. Appl. Microbiol. Biotechnol. 2004, 63, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, T. The C 7 N aminocyclitol family of natural products. Nat. Prod. Rep. 2003, 20, 137–166. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.T.; Shin, C.S. Reduced Formation of Byproduct Component C in Acarbose Fermentation by Actinoplanes sp. CKD485-16. Biotechnol. Prog. 2003, 19, 1677–1682. [Google Scholar] [CrossRef]
- Zhao, Q.; Xie, H.; Peng, Y.; Wang, X.; Bai, L. Improving acarbose production and eliminating the by-product component C with an efficient genetic manipulation system of Actinoplanes sp. SE50/110. Synth. Syst. Biotechnol. 2017, 2, 302–309. [Google Scholar] [CrossRef]
- Wendler, S.; Ortseifen, V.; Persicke, M.; Klein, A.; Neshat, A.; Niehaus, K.; Schneiker-Bekel, S.; Walter, F.; Wehmeier, U.F.; Kalinowski, J. Carbon source dependent biosynthesis of acarviose metabolites in Actinoplanes sp. SE50/110. J. Biotechnol. 2014, 191, 113–120. [Google Scholar] [CrossRef]
- Ortseifen, V.; Kalinowski, J.; Pühler, A.; Rückert, C. The complete genome sequence of the actinobacterium Streptomyces glaucescens GLA.O (DSM 40922) carrying gene clusters for the biosynthesis of tetracenomycin C, 5-hydroxy streptomycin, and acarbose. J. Biotechnol. 2017, 262, 84–88. [Google Scholar] [CrossRef]
- Hemker, M.; Stratmann, A.; Goeke, K.; Schröder, W.; Lenz, J.; Piepersberg, W.; Pape, H. Identification, cloning, expression, and characterization of the extracellular acarbose-modifying glycosyltransferase, AcbD, from Actinoplanes sp. strain SE50. J. Bacteriol. 2001, 183, 4484–4492. [Google Scholar] [CrossRef] [Green Version]
- Wendler, S.; Otto, A.; Ortseifen, V.; Bonn, F.; Neshat, A.; Schneiker-Bekel, S.; Walter, F.; Wolf, T.; Zemke, T.; Wehmeier, U.F. Comprehensive proteome analysis of Actinoplanes sp. SE50/110 highlighting the location of proteins encoded by the acarbose and the pyochelin biosynthesis gene cluster. J. Proteom. 2015, 125, 1–16. [Google Scholar] [CrossRef]
- Droste, J.; Ortseifen, V.; Schaffert, L.; Persicke, M.; Schneiker-Bekel, S.; Pühler, A.; Kalinowski, J. The expression of the acarbose biosynthesis gene cluster in Actinoplanes sp. SE50/110 is dependent on the growth phase. BMC Genom. 2020, 21, 818. [Google Scholar] [CrossRef]
- Boos, W.; Shuman, H. Maltose/maltodextrin system of Escherichia coli: Transport, metabolism, and regulation. Microbiol. Mol. Biol. Rev. 1998, 62, 204–229. [Google Scholar] [CrossRef] [Green Version]
- Lan, T.P.; Nguyen, T.M. Action Modes of MalQ isolated from Escherichia coli K12. J. Vietnam Agric. Sci. Technol. 2018, 3, 82–87. [Google Scholar]
- Seibold, G.M.; Wurst, M.; Eikmanns, B.J. Roles of maltodextrin and glycogen phosphorylases in maltose utilization and glycogen metabolism in Corynebacterium glutamicum. Microbiology 2009, 155, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, T.N.; Ryman, B.E.; Whelan, W.J. The action pattern of amylomaltase from Escherichia coli. Eur. J. Biochem. 1976, 69, 105–115. [Google Scholar] [CrossRef]
- Wendler, S.; Hürtgen, D.; Kalinowski, J.; Klein, A.; Niehaus, K.; Schulte, F.; Schwientek, P.; Wehlmann, H.; Wehmeier, U.F.; Pühler, A. The cytosolic and extracellular proteomes of Actinoplanes sp. SE50/110 led to the identification of gene products involved in acarbose metabolism. J. Biotechnol. 2013, 167, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Seibold, G.; Auchter, M.; Berens, S.; Kalinowski, J.; Eikmanns, B.J. Utilization of soluble starch by a recombinant Corynebacterium glutamicum strain: Growth and lysine production. J. Biotechnol. 2006, 124, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Xavier, K.B.; Peist, R.; Kossmann, M.; Boos, W.; Santos, H. Maltose metabolism in the hyperthermophilic archaeon Thermococcus litoralis: Purification and characterization of key enzymes. J. Bacteriol. 1999, 181, 3358–3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwientek, P.; Szczepanowski, R.; Rückert, C.; Kalinowski, J.; Klein, A.; Selber, K.; Wehmeier, U.F.; Stoye, J.; Pühler, A. The complete genome sequence of the acarbose producer Actinoplanes sp. SE50/110. BMC Genom. 2012, 13, 112. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Hung, Y.-J.; Qamruddin, K.; Aziz, M.F.A.; Stein, H.; Schmidt, B. International noninterventional study of acarbose treatment in patients with type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2011, 92, 57–64. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Pauli, G.F.; Chen, S.-N.; Friesen, J.B.; McAlpine, J.B.; Jaki, B.U. Analysis and purification of bioactive natural products: The AnaPurNa study. J. Nat. Prod. 2012, 75, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.C.; Skerra, A.; Schiefner, A. Structural basis for the interconversion of maltodextrins by MalQ, the amylomaltase of Escherichia coli. J. Biol. Chem. 2015, 290, 21352–21364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.H.D.; Park, S.-H.; Tran, P.L.; Kim, J.-W.; Le, Q.T.; Boos, W.; Park, J.-T. Characterization of the transglycosylation reaction of 4-α-glucanotransferase (MalQ) and its role in glycogen breakdown in Escherichia coli. J. Microbiol. Biotechnol. 2019, 29, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Wendler, S.; Otto, A.; Ortseifen, V.; Bonn, F.; Neshat, A.; Schneiker-Bekel, S.; Wolf, T.; Zemke, T.; Wehmeier, U.F.; Hecker, M. Comparative proteome analysis of Actinoplanes sp. SE50/110 grown with maltose or glucose shows minor differences for acarbose biosynthesis proteins but major differences for saccharide transporters. J. Proteom. 2016, 131, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Ball, J.; Reis, R.A.G.; Agniswamy, J.; Weber, I.T.; Gadda, G. Steric hindrance controls pyridine nucleotide specificity of a flavin-dependent NADH: Quinone oxidoreductase. Protein Sci. 2019, 28, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Feeney, R.; Clarke, A.R.; Holbrook, J.J. A single amino acid substitution in lactate dehydrogenase improves the catalytic efficiency with an alternative coenzyme. Biochem. Biophys. Res. Commun. 1990, 166, 667–672. [Google Scholar] [CrossRef]
- Wehmeier, U.F.; Piepersberg, W. Enzymology of aminoglycoside biosynthesis—Deduction from gene clusters. Methods Enzymol. 2009, 459, 459–491. [Google Scholar]
- Rockser, Y.; Wehmeier, U.F. The gac-gene cluster for the production of acarbose from Streptomyces glaucescens GLA.O—Identification, isolation and characterization. J. Biotechnol. 2009, 140, 114–123. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Yanisch-Perron, C.; Vieira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: Nucleotide sequences of the M13mpl8 and pUC19 vectors. Gene 1985, 33, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.; Bóna-Lovász, J.; Beuttler, H.; Altenbuchner, J. In vivo and in vitro studies on the carotenoid cleavage oxygenases from Sphingopyxis alaskensis RB2256 and Plesiocystis pacifica SIR-1 revealed their substrate specificities and non-retinal-forming cleavage activities. FEBS J. 2012, 279, 3911–3924. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
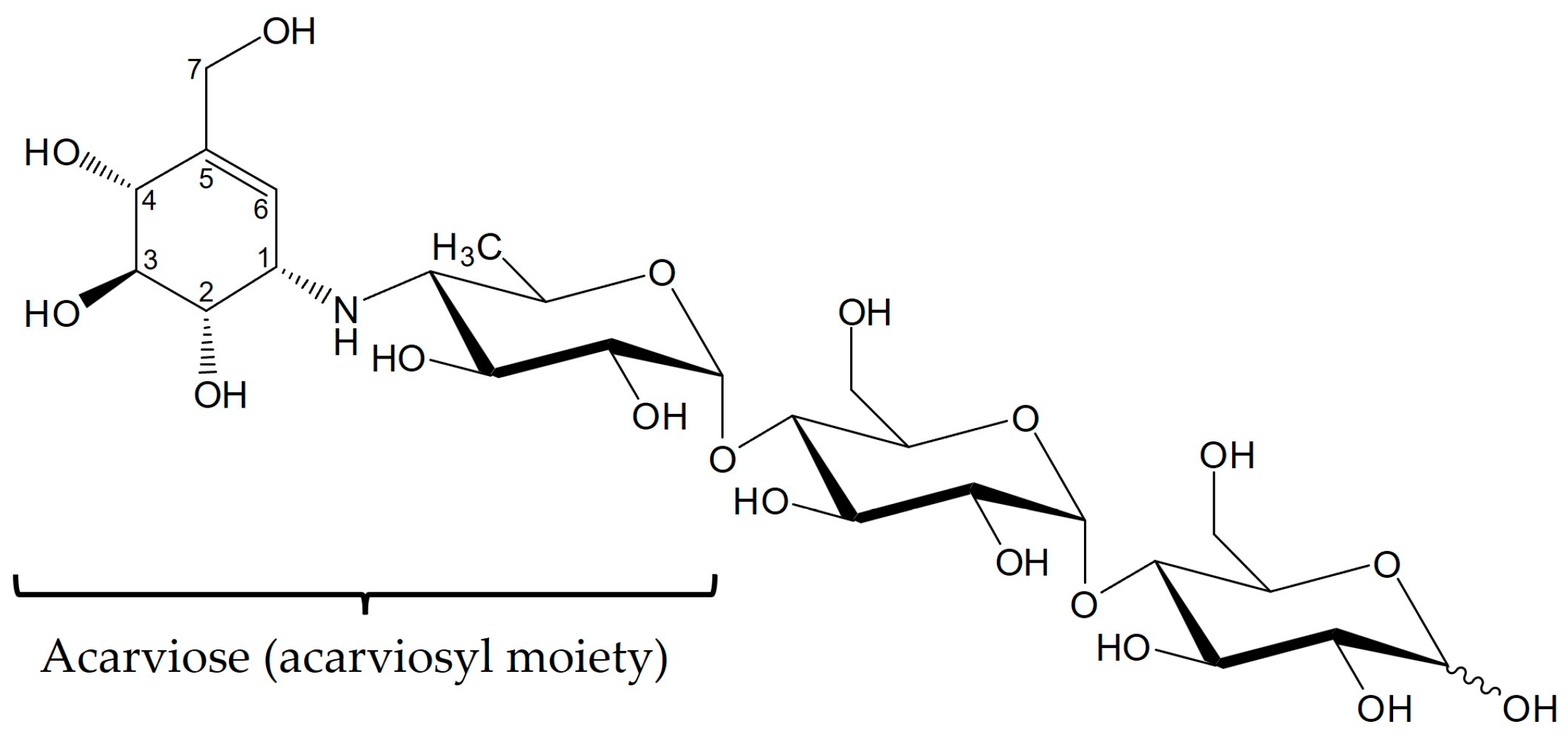
| Name | Specific Name (Used in This Study) | Structure |
|---|---|---|
| Component 1 | Acarviose | Ac |
| Component 2 | Acarviosyl-glucose | Ac-1,4-Glc |
| Component 3 | Acarviosyl-maltose (Acarbose) | Ac-1,4-Glc-1,4-Glc |
| Component 3a | Ac-1,4-Glc-1,4-Fru | |
| Component 3b | Ac-1,4-Glc-1,4-(1-epi-valienol) | |
| Component 3c | Ac-1,4-Glc-1,1-Glc | |
| Component 3d | Ac-1,4-Glc-1,4-Man | |
| Component 4a | Ac-1,4-Glc-1,4-Glc-1,4-Fru | |
| Component 4b | Acarviosyl-maltotriose | Ac-1,4-Glc-1,4-Glc-1,4-Glu |
| Component 4c | Ac-1,4-Glc-1,4-Glc-1,1-Glu | |
| Component 5–7 | Acarviosyl-maltotetraose/ -pentaose/-hexaose | Ac-1,4-Glc-1,4-Glc-1,4-Glu-1,4-Glu/ -1,4-Glu/-1,4-Glu |
| Acarviosyl Metabolites | Abbreviation | Sum Formula | [M + H]+ m/z |
|---|---|---|---|
| Acarviosyl-maltose (Acarbose) | Acb | C25H43NO18 | 646.2553 |
| Acarviosyl-maltotriose | Ac-G3 | C31H53NO23 | 808.3081 |
| Acarviosyl-maltotetraose | Ac-G4 | C37H63NO28 | 970.3609 |
| Acarviosyl-maltopentaose | Ac-G5 | C43H73NO33 | 1132.4138 |
| Acarviosyl-maltohexaose | Ac-G6 | C49H83NO38 | 1294.4665 |
| Acarviosyl-maltose 7-phosphate (Acarbose 7-phosphate) | Acb-7P | C25H44NO21P | 726.2216 |
| Acarviosyl-maltotriose 7-phosphate | Ac-G3-7P | C31H54NO26P | 888.2744 |
| Acarviosyl-maltotetraose 7-phosphate | Ac-G4-7P | C37H64NO31P | 1050.3273 |
| Acarviosyl-maltopentaose 7-phosphate | Ac-G5-7P | C43H74NO36P | 1212.3801 |
| Acarviosyl-maltohexaose 7-phosphate | Ac-G6-7P | C49H84NO41P | 1374.4329 |
| Ac-G3 | Ac-G4 | Ac-G5 | Ac-G6 | |
| Acb + G2 | + | + | ||
| Acb + G3 | (+) 1 | + | + | |
| Acb + G4 | (+) 1 | + | + | |
| Ac-G3-7P | Ac-G4-7P | Ac-G5-7P | Ac-G6-7P | |
| Acb-7P + G2 | (+) 1 | + | ||
| Acb-7P + G3 | + | |||
| Acb-7P + G4 | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nölting, S.; März, C.; Jacob, L.; Persicke, M.; Schneiker-Bekel, S.; Kalinowski, J. The 4-α-Glucanotransferase AcbQ Is Involved in Acarbose Modification in Actinoplanes sp. SE50/110. Microorganisms 2023, 11, 848. https://doi.org/10.3390/microorganisms11040848
Nölting S, März C, Jacob L, Persicke M, Schneiker-Bekel S, Kalinowski J. The 4-α-Glucanotransferase AcbQ Is Involved in Acarbose Modification in Actinoplanes sp. SE50/110. Microorganisms. 2023; 11(4):848. https://doi.org/10.3390/microorganisms11040848
Chicago/Turabian StyleNölting, Sophia, Camilla März, Lucas Jacob, Marcus Persicke, Susanne Schneiker-Bekel, and Jörn Kalinowski. 2023. "The 4-α-Glucanotransferase AcbQ Is Involved in Acarbose Modification in Actinoplanes sp. SE50/110" Microorganisms 11, no. 4: 848. https://doi.org/10.3390/microorganisms11040848
APA StyleNölting, S., März, C., Jacob, L., Persicke, M., Schneiker-Bekel, S., & Kalinowski, J. (2023). The 4-α-Glucanotransferase AcbQ Is Involved in Acarbose Modification in Actinoplanes sp. SE50/110. Microorganisms, 11(4), 848. https://doi.org/10.3390/microorganisms11040848