


Inorganic Carbon Assimilation and Electrosynthesis of Platform Chemicals in Bioelectrochemical Systems (BESs) Inoculated with Clostridium saccharoperbutylacetonicum N1-H4

 and
and 
Abstract
:1. Introduction
CO2 Capture by Microorganisms
2. Materials and Methods
2.1. Microorganisms and Culture Media
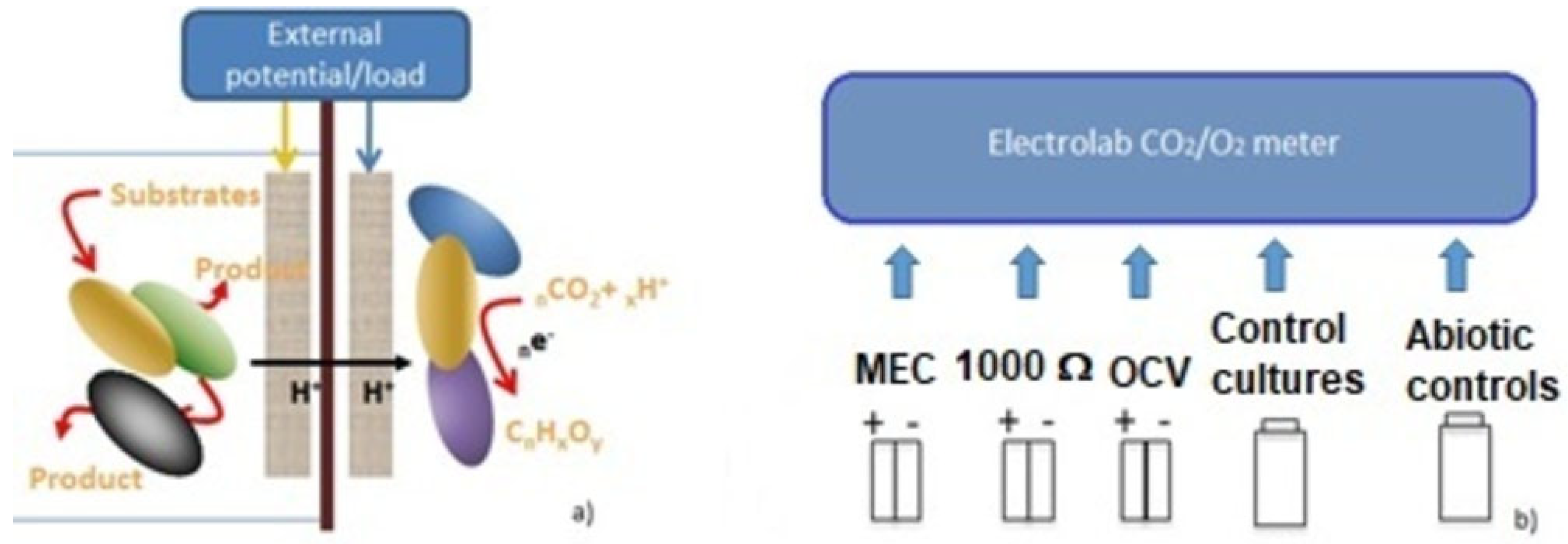
2.2. Bioanode and Biocathode Protocol
2.3. Set-Up of Two Chamber-MFCs for CO2 Capture
2.4. Metabolic Shift in C. saccharoperbutylacetonicum NT-1
2.5. CO2 Assimilation Efficiency
2.6. Energy Balance and CO2 Assimilation
2.7. Electrochemical Analyses
2.8. Screening of Electrosynthesis Products
2.9. Statistical Analysis
3. Results and Discussion
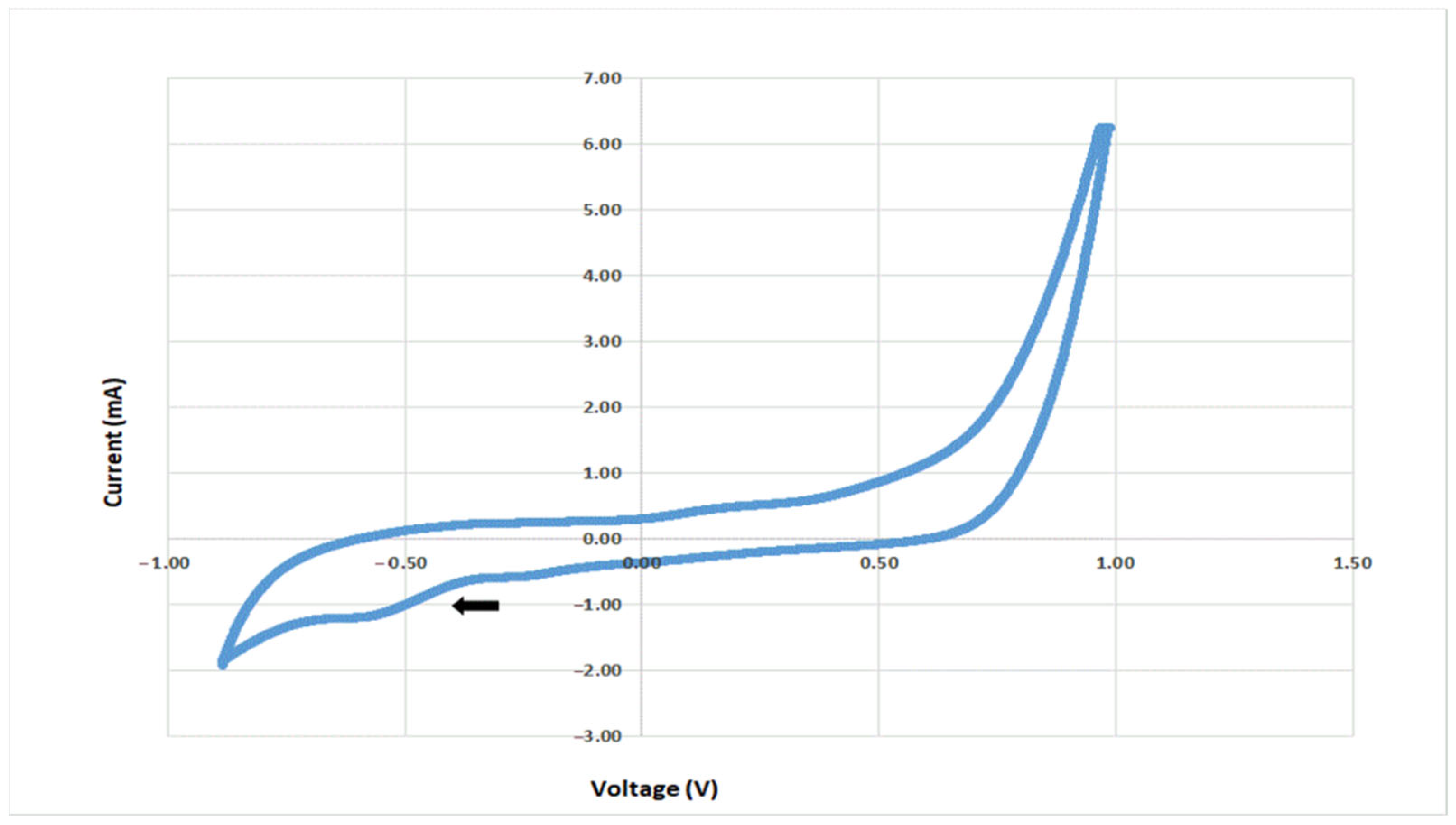
3.1. Electrochemical Potential along with Biofilm Preparation
3.2. Electrochemical Behavior
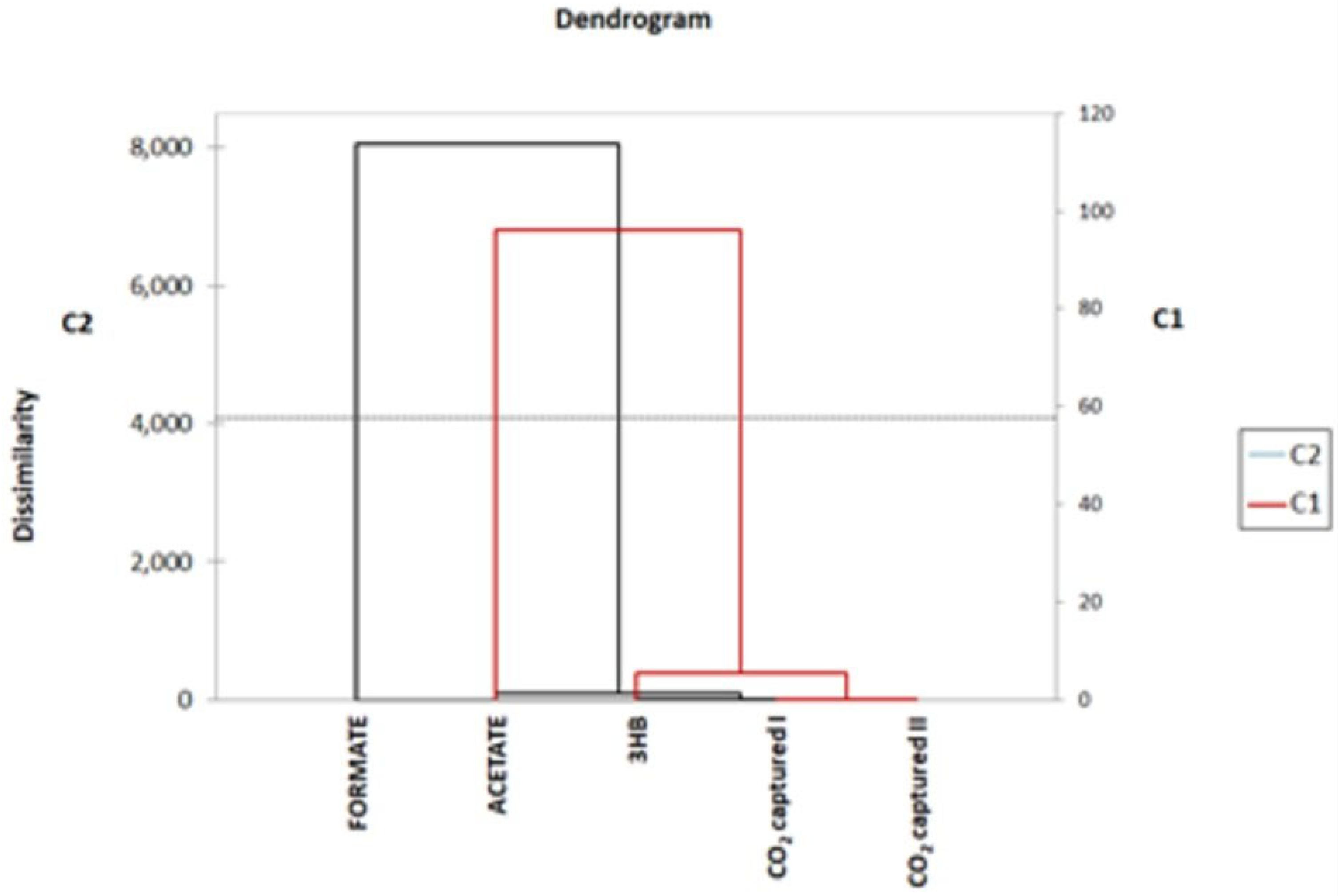
3.3. Inorganic Carbon Assimilation
3.4. Metabolic by-Products

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formate (mg/L*d) | CRform (%) | Acetate (mg/L*d) | CRac (%) | 3-D-Hydroxybutyrrate (mg/L*d) | CRbut (%) | |
|---|---|---|---|---|---|---|
| MFC_OCV | 87.1 ± 4.4 | 248.99 | 0 | 0 | 1.7 ± 0.3 | 0.081 |
| MFC_1000 Ω | 31.1 ± 2.8 | 52.8 | 0 | 0 | 2.3 ± 0.4 | 0.066 |
| MEC | 13.9 ± 0.8 | 29.1 | 11.4 ± 1.1 | 18.2 | 0 | 0 |
| Control culture | 39.1 ± 1.7 | 193.15 | 0 | 0 | 0.9 ± 0.5 | 0.07 |
3.5. Energy Balance
3.6. Metabolic Analysis Outcomes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Krebs, H.A. Carbon Dioxide Assimilation in Heterotrophic Organisms. Nature 1941, 147, 560–563. [Google Scholar] [CrossRef]
- Jones, S. Five ways to cycle carbon. Nat. Rev. Genet. 2008, 6, 95. [Google Scholar] [CrossRef]
- Branduardi, P.; Sauer, M.A. Microbial carbon dioxide fixation: New tricks for an old game. FEMS Microbiol. Lett. 2017, 35, fnx269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, A.B. Autotrophic CO2 assimilation and the evolution of ribulose diphosphate carboxylase. Bacteriol. Rev. 1973, 37, 289–319. [Google Scholar] [CrossRef] [PubMed]
- Sumi, T.; Harada, K. Kinetics of the ancestral carbon metabolism pathways in deep-branching bacteria and archaea. Commun. Chem. 2021, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, R.; Dempo, Y.; Nakayama, Y.; Nakamura, S.; Bamba, T.; Fukusaki, E.; Fukui, T. New Insight into the Role of the Calvin Cycle: Reutilization of CO2 Emitted through Sugar Degradation. Sci. Rep. 2015, 5, 11617. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Espinola, W.J.; Chinn, M.S.; Thon, M.R.; Bruno-Bárcena, J.M. Evidence of mixotrophic carbon-capture by n-butanol-producer Clostridium beijerinckii. Sci. Rep. 2017, 7, 12759. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Chauhan, P.; Gupta, S.; Raghuvanshi, S.; Singh, R.P.; Jha, P.N. CO2 sequestration potential of halo-tolerant bacterium Pseudomonas aeruginosa SSL-4 and its application for recovery of fatty alcohols. Process. Saf. Environ. Prot. 2017, 111, 582–591. [Google Scholar] [CrossRef]
- Xiong, W.; Linb, P.P.; Magnusson, L.; Warner, L.; Liao, J.C.; Maness, P.C.; Chou, K.J. CO2-fixing one-carbon metabolism in a cellulose degrading bacterium Clostridium thermocellum. Proc. Natl. Acad. Sci. USA 2016, 113, 13180–13185. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Al Makishah, N.H.; Li, Q.; Li, Y.; Liu, W.; Sun, X.; Wen, Z.; Yang, S. Developing Clostridia as Cell Factories for Short- and Medium-Chain Ester Production. Front. Bioeng. Biotechnol. 2021, 9, 661694. [Google Scholar] [CrossRef]
- Du, Y.; Zou, W.; Zhang, K.; Ye, G.; Yang, J. Advances and Applications of Clostridium Co-culture Systems in Biotechnology. Front. Microbiol. 2020, 11, 2842. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, C.M.; Minton, N.P. Advances in metabolic engineering in the microbial production of fuels and chemicals from C1 gas. Curr. Opin. Biotechnol. 2018, 50, 174–181. [Google Scholar] [CrossRef]
- Schievano, A.; Sciarria, T.P.; Vanbroekhoven, K.; De Wever, H.; Puig, S.; Andersen, S.J.; Rabaey, K.; Pant, D. Electro-Fermentation—Merging Electrochemistry with Fermentation in Industrial Applications. Trends Biotechnol. 2016, 34, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Maesen, M.; Dominguez-Benetton, X.; Vanbroekhoven, K.; Pant, D. Enzymatic electrosynthesis of formate through CO2 sequestration/reduction in a bioelectrochemical system (BES). Bioresour. Technol. 2014, 165, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; Srikanth, S.; Mohanakrishna, G.; Zacharia, R.; Strik, D.P.; Pant, D. Biotransformation of carbon dioxide in bioelectrochemical systems: State of the art and future prospects. J. Power Sources 2017, 356, 256–273. [Google Scholar] [CrossRef]
- Nastro, R.A.; Avignone-Rossa, C. Shewanella oneidensis MR1 and Pseudomonas aeruginosa PA1430/CO1 anodic consortium to sustain CO2 capture in Bioelectrochemical Systems (BESs). In Proceedings of the EU-ISMET—International Society for Microbial Electrochemistry and Technology, 4th European Meeting, Newcastle, UK, 12–14 September 2018. [Google Scholar]
- Noori, T.; Mohan, S.V.; Min, B. Microbial electrosynthesis of multi-carbon volatile fatty acids under the influence of different imposed potentials. Sustain. Energy Technol. Assess. 2021, 45, 101118. [Google Scholar] [CrossRef]
- Chandrasekhar, K.; Kondaveeti, S.; Nastro, R.A.; Naradasu, D.; Singh, M.; Mudumbai, H.; Enamala, M.K.; Chavali, M.; Kumar, P.S.M. Strategies in the direction of scaling-up aspects of microbial electrolysis cells. In Advances in Green and Sustainable Chemistry, Scaling Up of Microbial Electrochemical Systems; Jadhav, D.A., Pandit, S., Gajalakshmi, S., Shah, M.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 359–375. ISBN 9780323907651. [Google Scholar] [CrossRef]
- Saratale, G.D.; Banu, J.R.; Nastro, R.A.; Kadier, A.; Ashokkumar, V.; Lay, C.-H.; Jung, J.-H.; Shin, H.S.; Saratale, R.G.; Chandrasekhar, K. Bioelectrochemical systems in aid of sustainable biorefineries for the production of value-added products and resource recovery from wastewater: A critical review and future perspectives. Bioresour. Technol. 2022, 359, 127435. [Google Scholar] [CrossRef]
- Nelabhotla, A.B.T.; Dinamarca, C. Bioelectrochemical CO2 Reduction to Methane: MES Integration in Biogas Production Processes. Appl. Sci. 2019, 9, 1056. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Noori, M.T.; Núñez, A.E.; Verma, N. An insight into the bioelectrochemical photoreduction of CO2 to value-added chemicals. Iscience 2021, 24, 102294. [Google Scholar] [CrossRef]
- Im, C.; Valgepea, K.; Modin, O.; Nygård, Y. Clostridium ljungdahlii as a biocatalyst in microbial electrosynthesis—Effect of culture conditions on product formation. Bioresour. Technol. Rep. 2022, 19, 101156. [Google Scholar] [CrossRef]
- Giddings, C.G.S.; Nevin, K.P.; Ewoodward, T. Simplifying microbial electrosynthesis reactor design. Front. Microbiol. 2015, 6, 468. [Google Scholar] [CrossRef] [Green Version]
- Mohanakrishna, G.; Vanbroekhoven, K.; Pant, D. Imperative role of applied potential and inorganic carbon source on acetate production through microbial electrosynthesis. J. CO2 Util. 2016, 15, 57–64. [Google Scholar] [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. USA 2012, 109, 10042–10046. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Feng, J.; Guo, L.; Fasina, O.; Wang, Y. Engineering Clostridium saccharoperbutylacetonicum for High Level Isopropanol-Butanol-Ethanol (IBE) Production from Acetic Acid Pretreated Switchgrass Using the CRISPR-Cas9 System. ACS Sustain. Chem. Eng. 2019, 7, 18153–18164. [Google Scholar] [CrossRef]
- Das, S.; Das, S.; Das, I.; Ghangrekar, M. Application of bioelectrochemical systems for carbon dioxide sequestration and concomitant valuable recovery: A review. Mater. Sci. Energy Technol. 2019, 2, 687–696. [Google Scholar] [CrossRef]
- Bajracharya, S.; Heijne, A.; Benetton, X.D.; Vanbroekhoven, K.; Buisman, C.J.N.; Strik, D.P.B.T.B.; Pant, D. Carbon dioxide reduction by mixed and pure cultures in microbial electrosynthesis using an assembly of graphite felt and stainless steel as a cathode. Bioresour. Technol. 2015, 195, 14–24. [Google Scholar] [CrossRef]
- Geerlof, A. Helmholtz Center Munich. 2010. Available online: https://static.igem.org/mediawiki/2019/2/20/T--Tuebingen--M9_recipe.pdf (accessed on 1 October 2022).
- Kim, C.; Song, Y.E.; Lee, C.R.; Jeon, B.-H.; Kim, J.R. Glycerol-fed microbial fuel cell with a co-culture of Shewanella oneidensis MR-1 and Klebsiella pneumonae J2B. J. Ind. Microbiol. Biotechnol. 2016, 43, 1397–1403. [Google Scholar] [CrossRef]
- Scoffield, J.; Silo-Suh, L. Glycerol metabolism promotes biofilm formation by Pseudomonas aeruginosa. Can. J. Microbiol. 2016, 62, 704–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoareau, M.; Erable, B.; Bergel, A. Microbial electrochemical snorkels (MESs): A budding technology for multiple applications. A mini review. Electrochem. Commun. 2019, 104, 106473. [Google Scholar] [CrossRef]
- Gambino, E.; Toscanesi, M.; Del Prete, F.; Flagiello, F.; Falcucci, G.; Minutillo, M.; Trifuoggi, M.; Guida, M.; Nastro, R.A.; Jannelli, E. Polycyclic Aromatic Hydrocarbons (PAHs) Degradation and Detoxification of Water Environment in Single-chamber Air-cathode Microbial Fuel Cells (MFCs). Fuel Cells 2017, 17, 618–626. [Google Scholar] [CrossRef]
- Liu, C.-G.; Qin, J.-C.; Lin, Y.-H. Fermentation and redox potential. In Fermentation Processes; Jozala, A.F., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Minutillo, M.; Nastro, R.A.; Di Micco, S.; Jannelli, E.; Cioffi, R.; Di Giuseppe, M. Performance Assessment of Multi-Electrodes Reactors for Scaling-up Microbial Fuel Cells. E3S Web Conf. 2020, 197, 08020. [Google Scholar] [CrossRef]
- Kracke, F.; Virdis, B.; Bernhardt, P.V.; Rabaey, K.; Krömer, J.O. Redox dependent metabolic shift in Clostridium autoethanogenum by extracellular electron supply. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumontet, S.; Cavoski, I.; Ricciuti, P.; Mondelli, D.; Jarrar, M.; Pasquale, V.; Crecchio, C. Metabolic and genetic patterns of soil microbial communities in response to different amendments under organic farming system. Geoderma 2017, 296, 79–85. [Google Scholar] [CrossRef]
- Nastro, R.; Leccisi, E.; Toscanesi, M.; Liu, G.; Trifuoggi, M.; Ulgiati, S. Exploring Avoided Environmental Impacts as Well as Energy and Resource Recovery from Microbial Desalination Cell Treatment of Brine. Energies 2021, 14, 4453. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal component analysis: A review and recent developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef] [Green Version]
- Funari, R.; Shen, A.Q. Detection and Characterization of Bacterial Biofilms and Biofilm-Based Sensors. ACS Sens. 2022, 7, 347–357. [Google Scholar] [CrossRef]
- Corrigan, P.S.; Tirsch, J.L.; Silakov, A. Investigation of the Unusual Ability of the [FeFe] Hydrogenase from Clostridium beijerinckii to Access an O2-Protected State. J. Am. Chem. Soc. 2020, 142, 12409–12419. [Google Scholar] [CrossRef]
- Westphal, L.; Wiechmann, A.; Baker, J.; Minton, N.P.; Müller, V. The Rnf Complex Is an Energy-Coupled Transhydrogenase Essential To Reversibly Link Cellular NADH and Ferredoxin Pools in the Acetogen Acetobacterium woodii. J. Bacteriol. 2018, 200, e00357. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.; Ching, K.; Kumar, P.V.; Zhao, C.; Kumar, N.; Chen, X.J.; Das, B. Vitamin B12 on Graphene for Highly Efficient CO2 Electroreduction. ACS Appl. Mater. Interfaces 2020, 2020 12, 41288–41293. [Google Scholar] [CrossRef]
- Wan, N.; Wang, H.; Ng, C.K.; Mukherjee, M.; Ren, D.; Cao, B.; Tang, Y.J. Bacterial Metabolism During Biofilm Growth Investigated by 13C Tracing. Front. Microbiol. 2018, 9, 2657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, J.; Humphreys, J.R.; Magnusson, L.; Wachter, B.; Urban, C.; Hebdon, S.D.; Xiong, W.; Chou, K.J.; Ching, M.P. Acetogenic production of 3-Hydroxybutyrate using a native 3-Hydroxybutyryl-CoA Dehydrogenase. Front. Microbiol. 2022, 13, 948369. [Google Scholar] [CrossRef] [PubMed]
- Prévoteau, A.; Carvajal-Arroyo, J.M.; Ganigué, R.; Rabaey, K. Microbial electrosynthesis from CO2: Forever a promise? Curr. Opin. Biotechnol. 2020, 62, 48–57. [Google Scholar] [CrossRef]
- Roy, M.; Aryal, N.; Zhang, Y.; Patil, S.A.; Pant, D. Technological progress and readiness level of microbial electrosynthesis and electrofermentation for carbon dioxide and organic wastes valorization. Curr. Opin. Green Sustain. Chem. 2022, 35, 100605. [Google Scholar] [CrossRef]
- Omidi, M.; Mashkour, M.; Biswas, J.K.; Garlapati, V.K.; Singh, L.; Rahimnejad, M.; Pant, D. From Electricity to Products: Recent Updates on Microbial Electrosynthesis (MES). Top. Catal. 2021. [Google Scholar] [CrossRef]
- Junkins, E.N.; McWhirter, J.B.; McCall, L.I.; Stevenson, B.S. Environmental structure impacts microbial composition and secondary metabolism. ISME Commun. 2022, 2, 15. [Google Scholar] [CrossRef]





| Sample | Residual CO2 in the Headspace (%) | Residual CO2 in the Headspace (%) | Residual Concentration in the Medium (M) | Residual Concentration in the Medium (M) | nCO2 Captured (mol) | nCO2 Captured (mol) | % CO2 Captured (mol) | % CO2 Captured (mol) |
|---|---|---|---|---|---|---|---|---|
| 1st Measurement * | 2nd Measurement ’ | 1st Measurement * | 2nd Measurement ’ | 1st Measurement * | 2nd Measurement ’ | 1st Measurement * | 2nd Measurement ’ | |
| MEC | 0.46 | 1.34 | 0.078 | 0.227 | 0.261 | 0.112 | 76.99 | 32.98 |
| MFC_OCV | 0.595 | 1.38 | 0.101 | 0.234 | 0.238 | 0.105 | 70.24 | 30.98 |
| MFC_1000 Ω) | 0.09 | 1.43 | 0.015 | 0.242 | 0.324 | 0.097 | 95.50 | 28.47 |
| MEC (blank) | 0.89 | 1.45 | 0.151 | 0.246 | 0.188 | 0.093 | 55.48 | 27.47 |
| MFC(blank)_1000 Ω | 1.7 | 1.58 | 0.288 | 0.268 | 0.051 | 0.071 | 14.97 | 20.97 |
| MFC(blank)_OCV | 1.54 | 1.57 | 0.261 | 0.266 | 0.078 | 0.073 | 22.97 | 21.47 |
| Control culture | 1.55 | 1.65 | 0.263 | 0.280 | 0.076 | 0.059 | 22.47 | 17.47 |
| Abiotic control | 1.6 | 1.7 | 0.271 | 0.288 | 0.068 | 0.051 | 19.97 | 14.97 |
| AwCD_BES_24 h | AwCD_Coll_24 h | AwCD_BES_144 h | AwCD_Coll_144 h | |
|---|---|---|---|---|
| Carbohydrates | 0.009 | 0.027 | 0.0794 | 0.637 |
| Polymers | 0.019 | 0 | 0 | 0 |
| Ammines | 0 | 0 | 0 | 0 |
| Amminoacids | 0 | 0 | 0.085 | 0 |
| Phenolic Compounds | 0 | 0 | 0 | 0 |
| Average | 0.005 | 0.009 | 0.101 | 0.219 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nastro, R.A.; Salvian, A.; Kuppam, C.; Pasquale, V.; Pietrelli, A.; Rossa, C.A. Inorganic Carbon Assimilation and Electrosynthesis of Platform Chemicals in Bioelectrochemical Systems (BESs) Inoculated with Clostridium saccharoperbutylacetonicum N1-H4. Microorganisms 2023, 11, 735. https://doi.org/10.3390/microorganisms11030735
Nastro RA, Salvian A, Kuppam C, Pasquale V, Pietrelli A, Rossa CA. Inorganic Carbon Assimilation and Electrosynthesis of Platform Chemicals in Bioelectrochemical Systems (BESs) Inoculated with Clostridium saccharoperbutylacetonicum N1-H4. Microorganisms. 2023; 11(3):735. https://doi.org/10.3390/microorganisms11030735
Chicago/Turabian StyleNastro, Rosa Anna, Anna Salvian, Chandrasekhar Kuppam, Vincenzo Pasquale, Andrea Pietrelli, and Claudio Avignone Rossa. 2023. "Inorganic Carbon Assimilation and Electrosynthesis of Platform Chemicals in Bioelectrochemical Systems (BESs) Inoculated with Clostridium saccharoperbutylacetonicum N1-H4" Microorganisms 11, no. 3: 735. https://doi.org/10.3390/microorganisms11030735
APA StyleNastro, R. A., Salvian, A., Kuppam, C., Pasquale, V., Pietrelli, A., & Rossa, C. A. (2023). Inorganic Carbon Assimilation and Electrosynthesis of Platform Chemicals in Bioelectrochemical Systems (BESs) Inoculated with Clostridium saccharoperbutylacetonicum N1-H4. Microorganisms, 11(3), 735. https://doi.org/10.3390/microorganisms11030735