

Conversion of Mixed Waste Food Substrates by Carotenogenic Yeasts of Rhodotorula sp. Genus

 , , , ,
, , , ,
Abstract
:1. Introduction
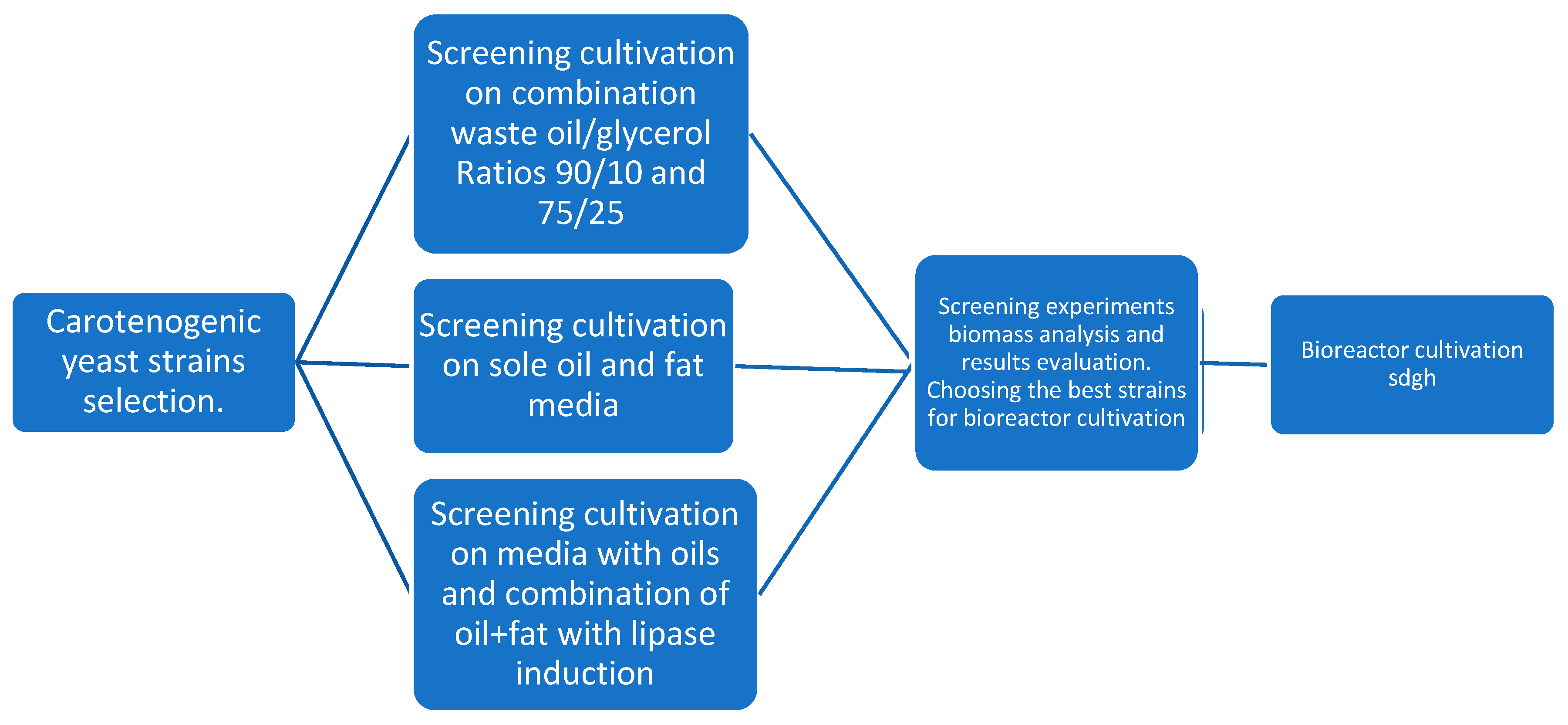
2. Materials and Methods
2.1. Yeast Strains
2.2. Microorganisms Cultivation, Hydrolysate, and Media Preparation
2.2.1. Yeast Inoculation
2.2.2. Yeast Production Media
2.2.3. Large-Scale Bioreactor Co-Cultivation
2.3. Waste Materials
2.3.1. Waste Glycerol
2.3.2. Waste Oils and Fats
2.4. Analytical Methods
2.4.1. Waste Glycerol Analysis
2.4.2. Phenolics in Coffee Oil
2.4.3. Cell Dry Weight
2.4.4. Lipid Metabolite Analysis
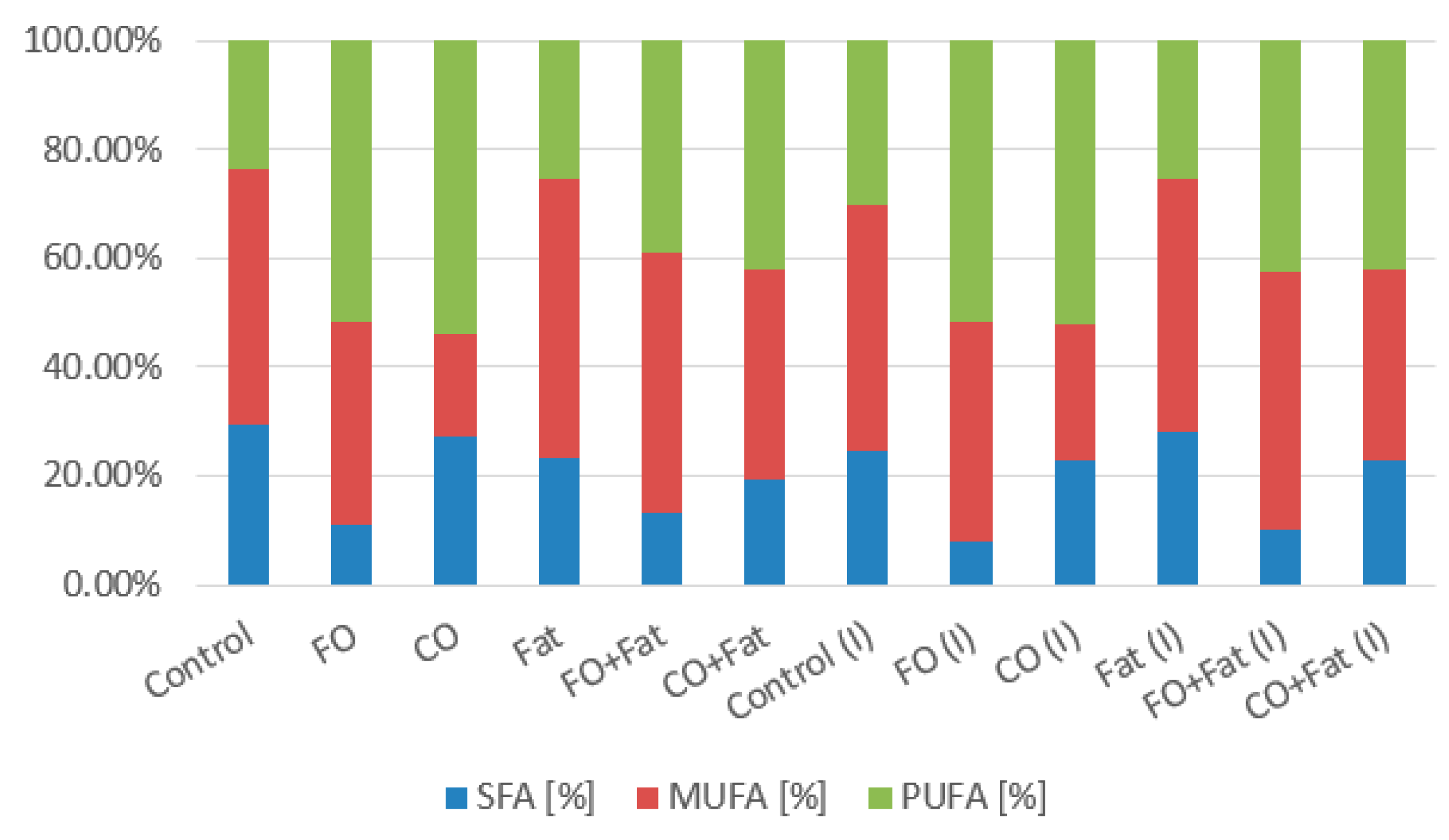
2.4.5. Lipids and Fatty Acids
2.4.6. β-Glucan Determination
Total Glucan Assay
α-Glucan Assay
2.5. Statistical Analysis
3. Results
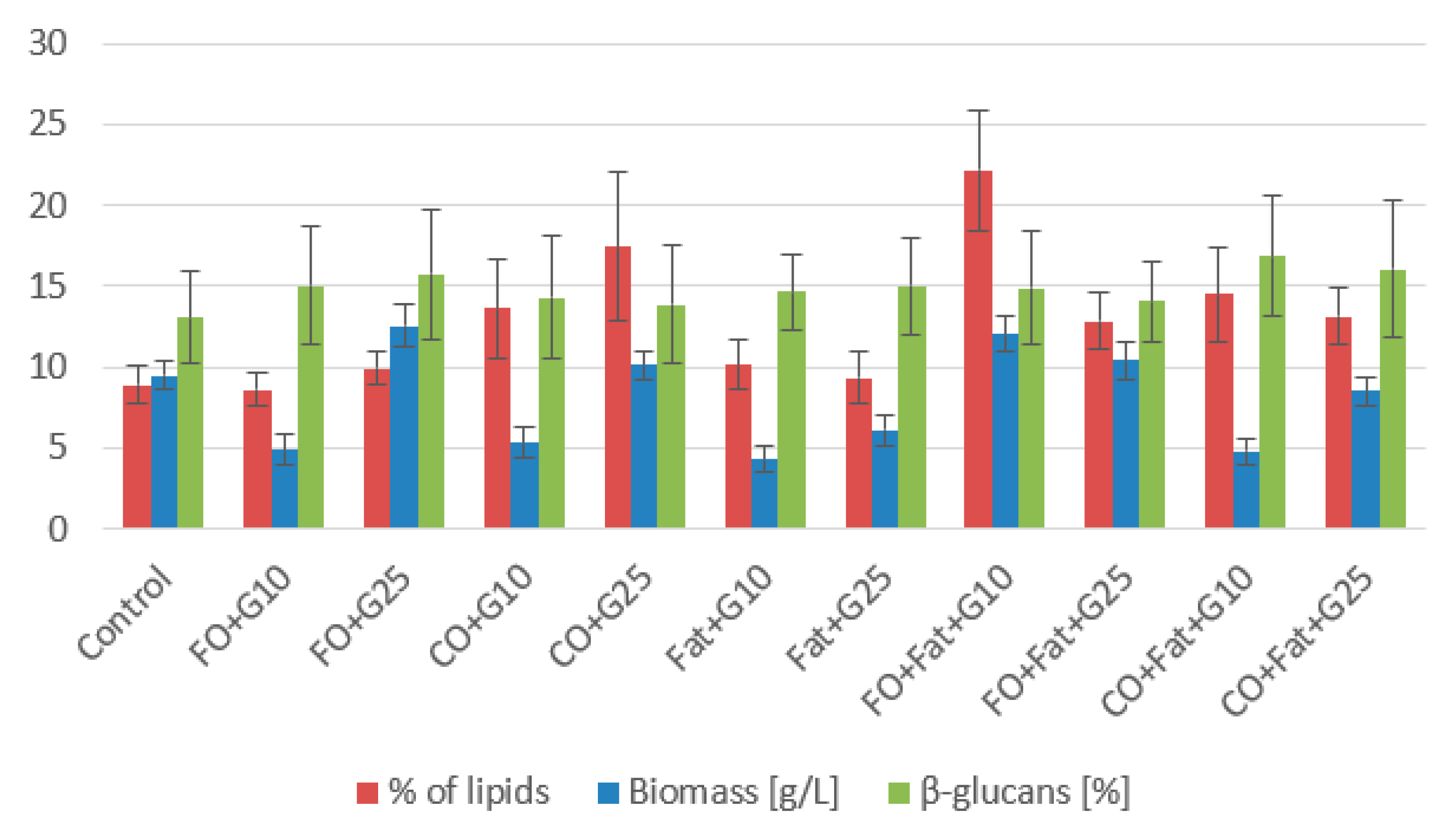
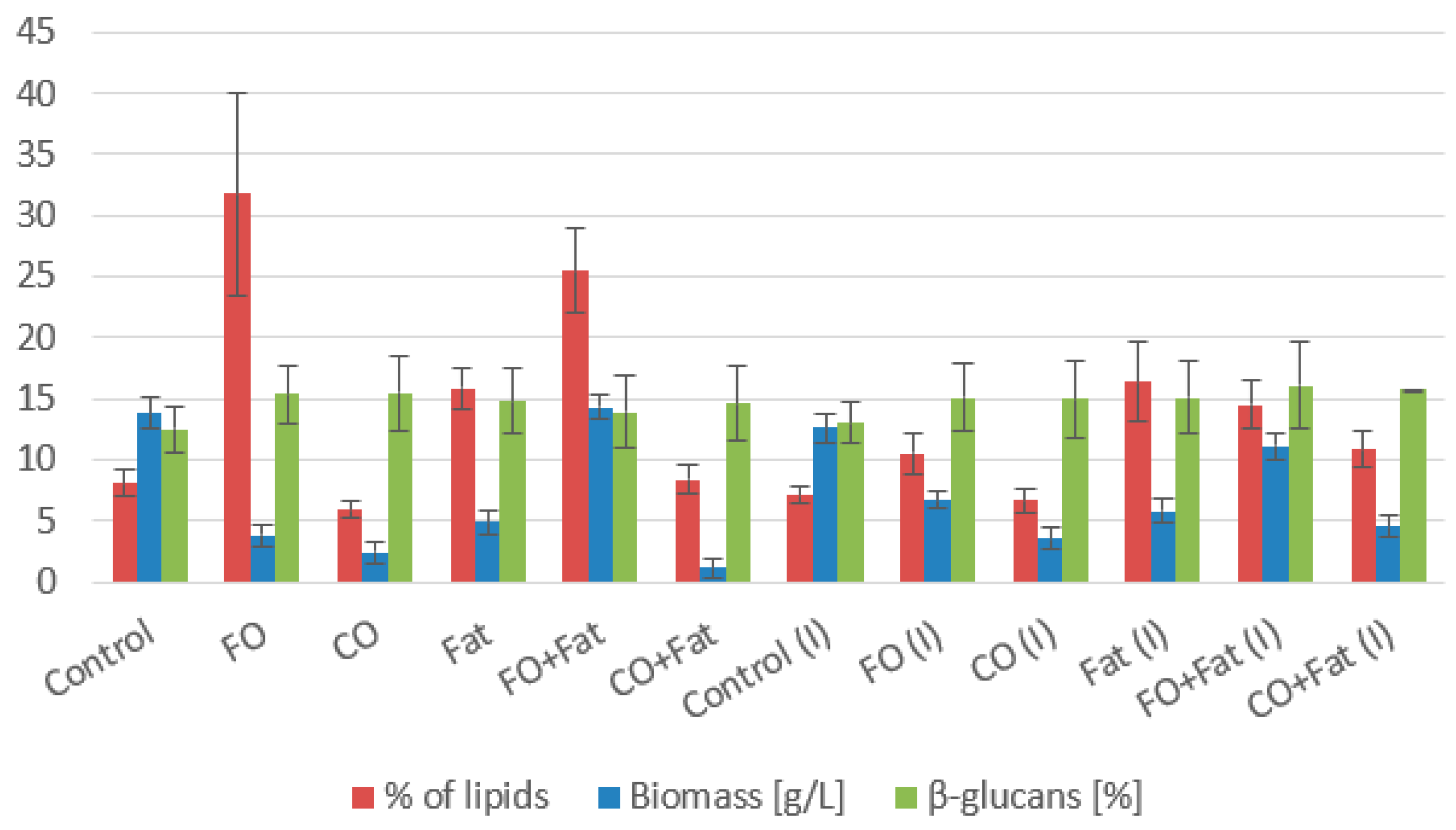
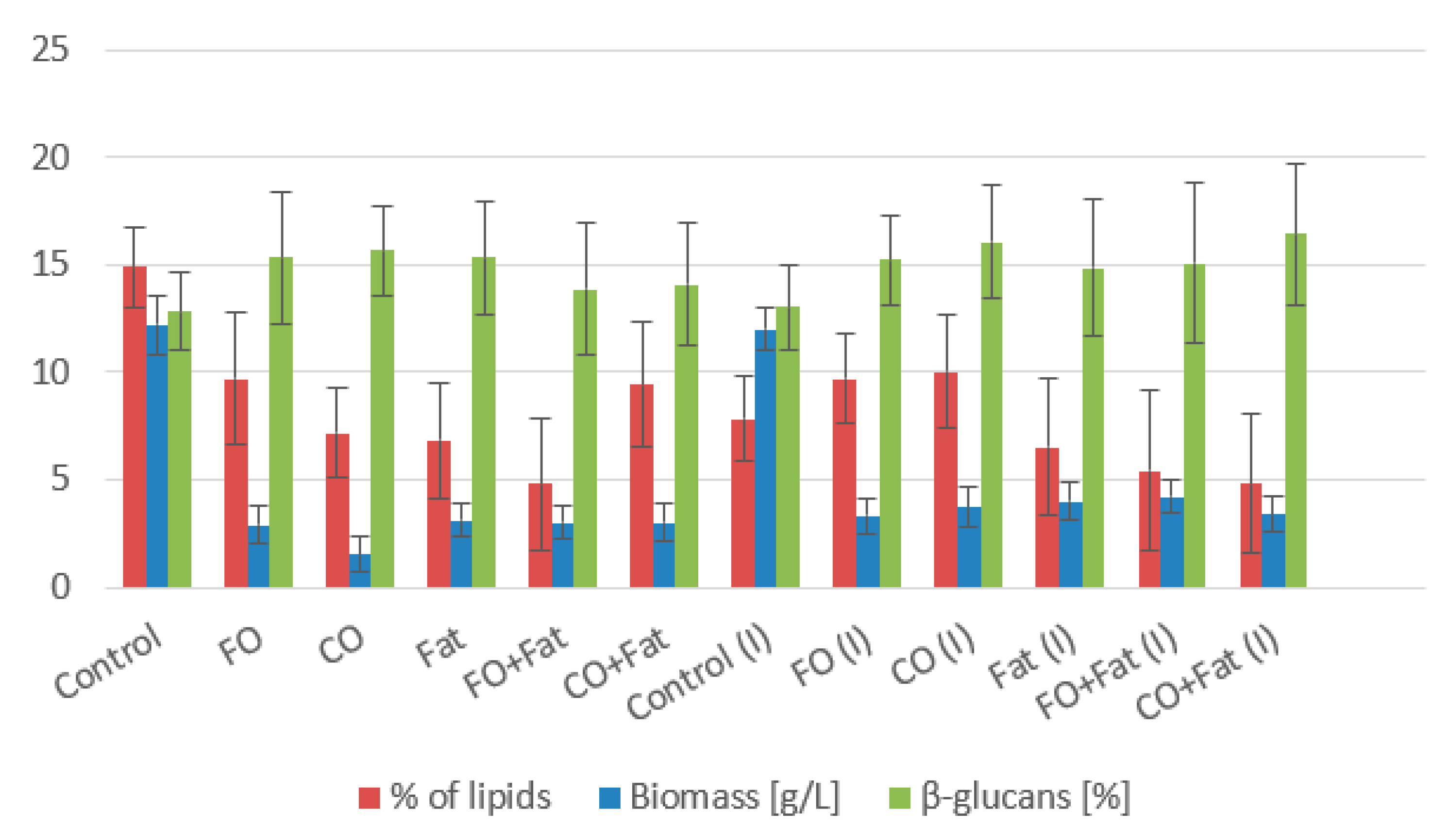
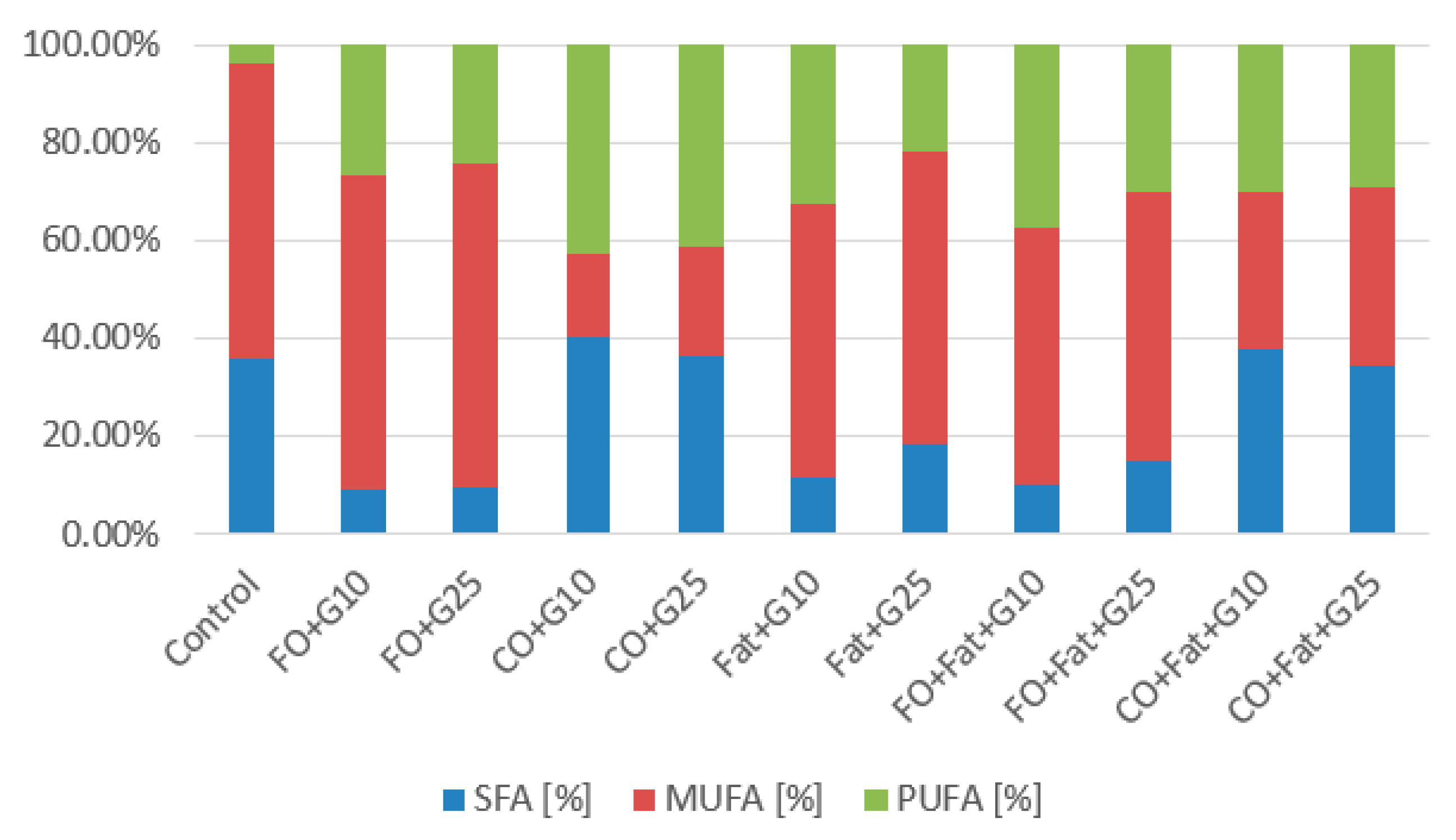
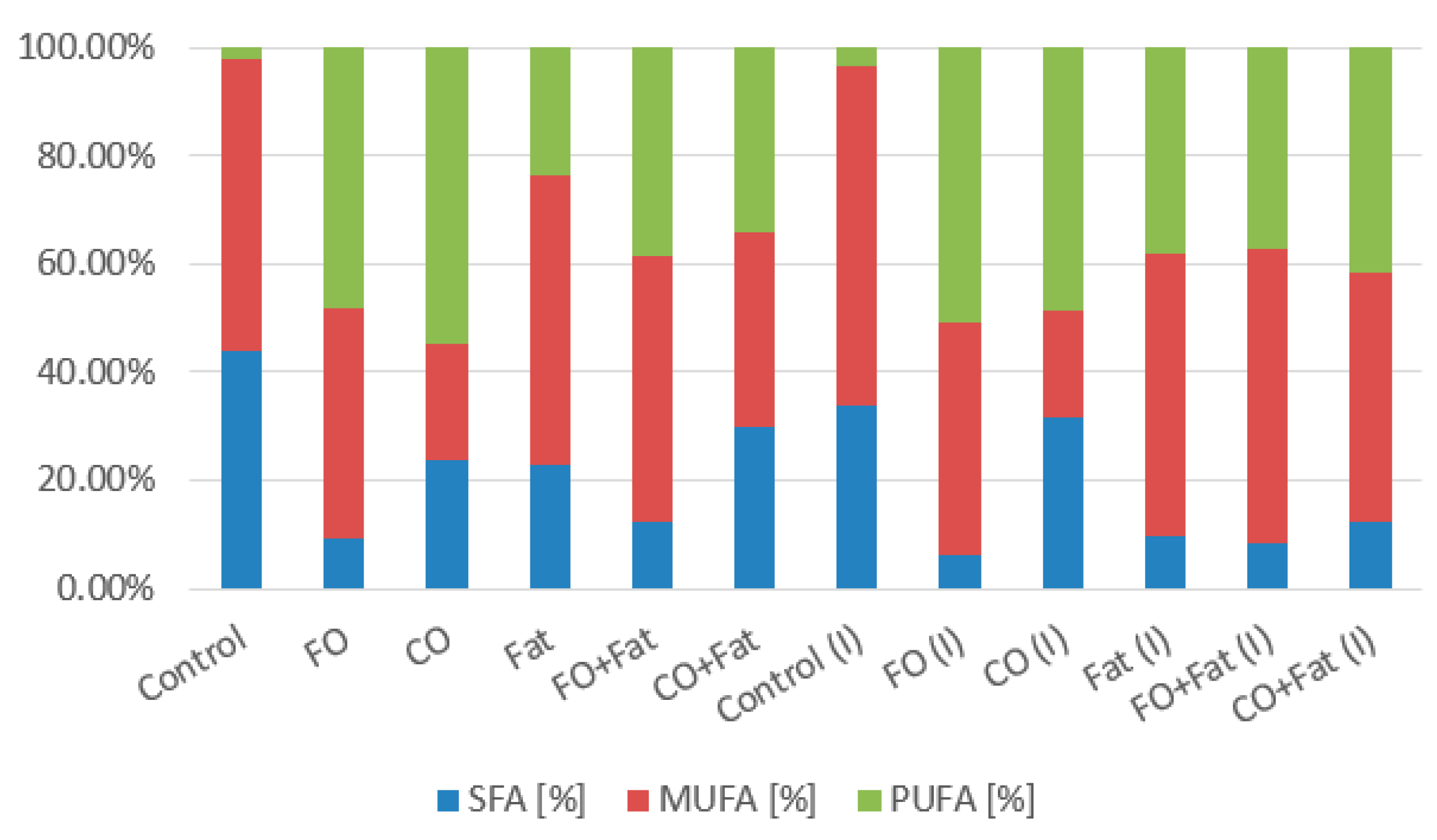
3.1. Phase I and Phase II Screening Cultivation Results
3.1.1. Rhodotorula kratochvilovae CCY 020-002-026 Cultivation
Rhodotorula kratochvilovae CCY 020-002-026 Cultivation—Statistical Analysis
3.1.2. Rhodotorula toruloides CCY 062-002-004 Cultivation
Rhodotorula toruloides CCY 062-002-004 Cultivation—Statistical Analysis
3.1.3. Rhodotorula mucilaginosa CCY 019-004-006 Cultivation
Rhodotorula mucilaginosa CCY 019-004-006 Cultivation—Statistical Analysis
3.1.4. Sporidiobolus pararoseus CCY 019-009-006 Cultivation
Sporidiobolus pararoseus CCY 019-009-006 Cultivation—Statistical Analysis
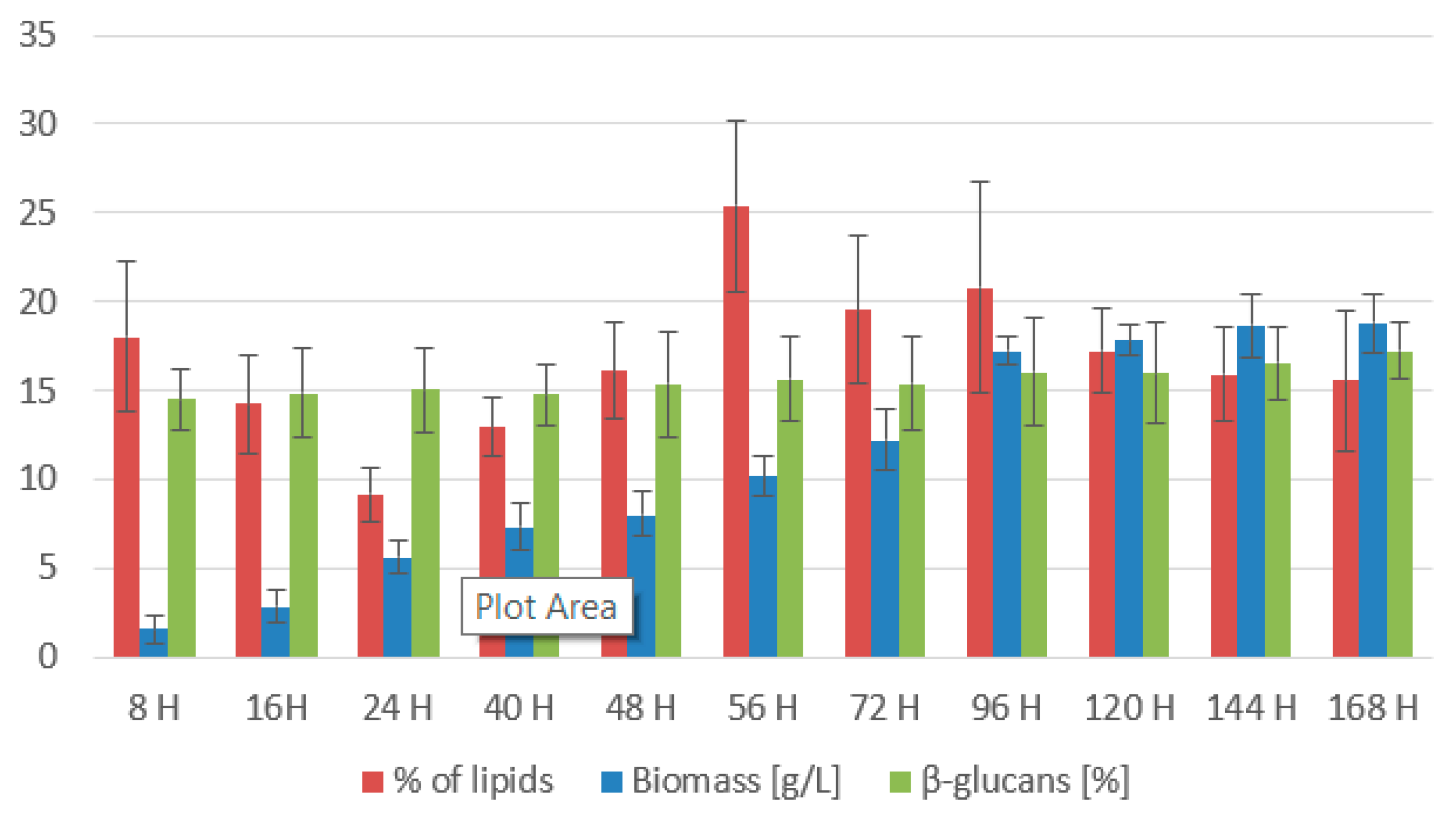
3.2. Bioreactor Cultivation of the Yeasts
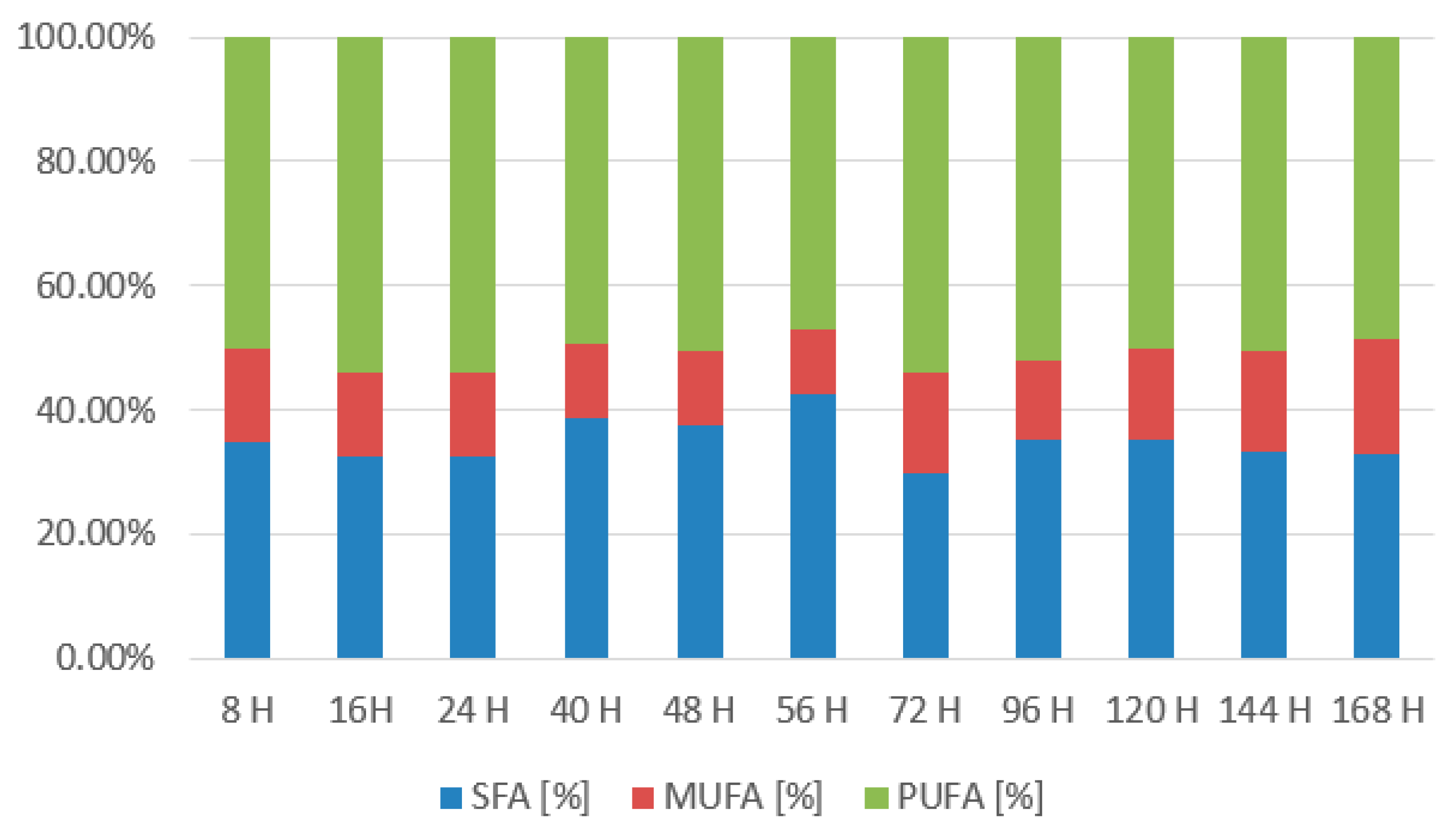
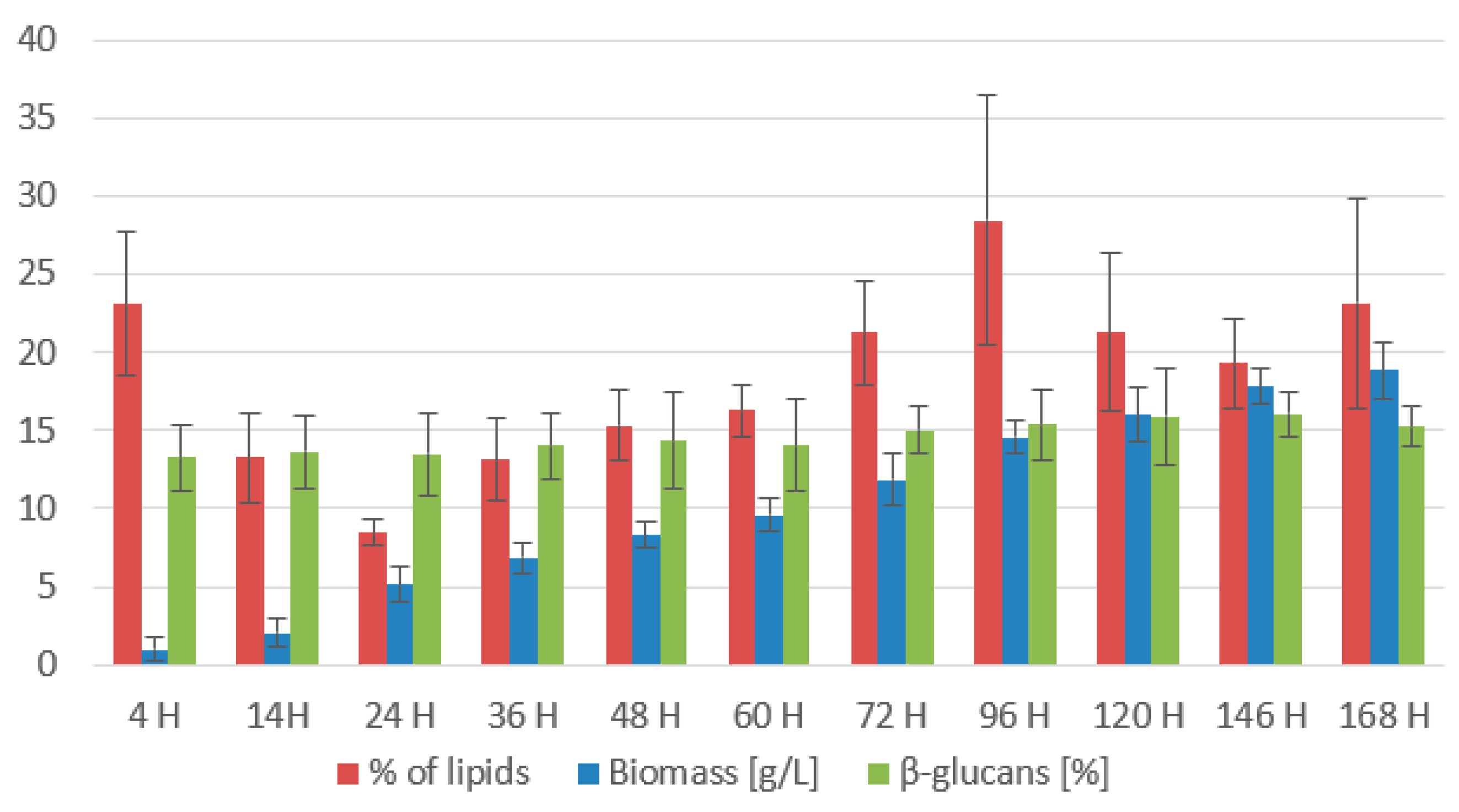
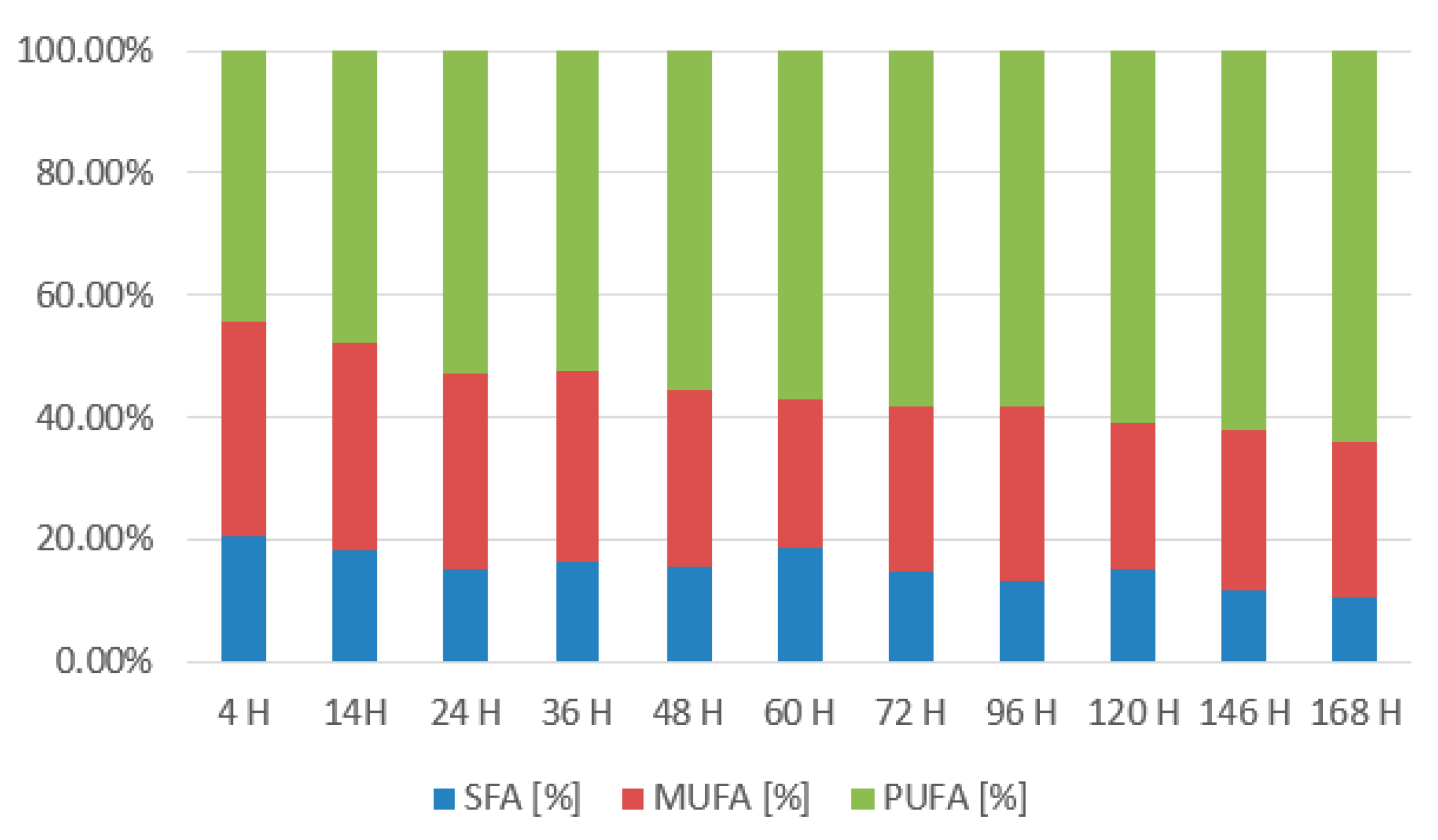
3.2.1. Rhodotorula kratochvilovae CCY 020-002-026 Bioreactor Cultivation
3.2.2. Rhodotorula toruloides CCY 062-002-004 Bioreactor Cultivation
3.2.3. β-Glucan Production Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schilling, C.; Weiss, S.A. Roadmap for Industry to Harness Biotechnology for a More Circular Economy. New Biotechnol. 2021, 60, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Fang, C. Circular economy development in China-current situation, evaluation and policy implications. Environ. Impact Assess. Rev. 2020, 84, 106441. [Google Scholar] [CrossRef]
- Scarlat, N.; Dallemand, J.-F.; Monforti-Ferrario, F.; Nita, V. The role of biomass and bioenergy in a future bioeconomy: Policies and facts. Environ. Dev. 2015, 15, 3–34. [Google Scholar] [CrossRef]
- Velasco-Muñoz, J.F.; Aznar-Sánchez, J.A.; López-Felices, B.; Román-Sánchez, I.M. Circular economy in agriculture. An analysis of the state of research based on the life cycle. Sustain. Prod. Consum. 2022, 34, 257–270. [Google Scholar] [CrossRef]
- Carmona-Cabello, M.; Garcia, I.L.; Leiva-Candia, D.; Dorado, M.P. Valorization of food waste based on its composition through the concept of biorefinery. Curr. Opin. Green Sustain. Chem. 2018, 14, 67–79. [Google Scholar] [CrossRef]
- Total production by all exporting countries. In Historical Data on the Global Coffee; International Coffee Organization: London, UK, 2020.
- Jenkins, R.W.; Stageman, N.E.; Fortune, C.M.; Chuck, C.J. Effect of the Type of Bean, Processing, and Geographical Location on the Biodiesel Produced from Waste Coffee Grounds. Energy Fuels 2014, 28, 1166–1174. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Saez, N.; García, A.T.; Peréz, I.D.; Rebollo-Hernanz, M.; Mesías, M.; Morales, F.J.; Martín-Cabrejas, M.A.; Del Castillo, M.D. Use of spent coffee grounds as food ingredient in bakery products. Food Chem. 2017, 216, 114–122. [Google Scholar] [CrossRef]
- Girotto, F.; Pivato, A.; Cossu, R.; Nkeng, G.E.; Lavagnolo, M.C. The broad spectrum of possibilities for spent coffee grounds valorisation. J. Mater. Cycles Waste Manag. 2018, 20, 695–701. [Google Scholar] [CrossRef]
- Kumar, L.R.; Kaur, R.; Tygi, R.D.; Drogui, P. Identifying economical route for crude glycerol valorisation: Biodiesel versus polyhydroxy-butyrate (PHB). Bioresour. Technol. 2021, 323, 124565. [Google Scholar] [CrossRef]
- Santos, C.; Fonseca, J.; Aaires, A.; Coutinho, J.; Trindade, H. Effect of different rates of spent coffee grounds (SCG) on composting process, gaseous emissions and quality of end-product. Waste Manag. 2017, 59, 37–47. [Google Scholar] [CrossRef]
- Limousy, L.; Jeguirim, M.; Dutournié, P.; Kraiem, N.; Lajili, M.; Said, R. Gaseous products and particulate matter emissions of biomass residential boiler fired with spent coffee grounds pellets. Fuel 2013, 107, 323–329. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Kucera, D.; Petrik, S.; Marova, I. Biotechnological conversion of spent coffee grounds into polyhydroxyalkanoates and carotenoids. New Biotechnol. 2015, 32, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Aksu, Z.; Eren, A.T. Carotenoids production by the yeast Rhodotorula mucilaginosa: Use of agricultural wastes as a carbon source. Process Biochem. 2005, 40, 2985–2991. [Google Scholar] [CrossRef]
- Kamilah, H.; Tsuge, T.; Yang, T.A.; Sudesh, K. Waste cooking oil as substrate for biosynthesis of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3-hydroxyhexanoate): Turning waste into a value-added product. Malays. J. Microbiol. 2013, 9, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Marova, I.; Certik, M.; Breierova, E. Production of Enriched Biomass by Carotenogenic Yeasts—Application of Whole-Cell Yeast Biomass to Production of Pigments and Other Lipid Compounds. In Biomass—Detection, Production and Usage; InTech: London, UK, 2011; ISBN 978-953-307-492-4. [Google Scholar]
- Szotkowski, M.; Byrtusova, D.; Haronikova, A.; Vysoka, M.; Rapta, M.; Shapaval, V.; Marova, I. Study of metabolic adaptation of red yeast to waste animal fat substrate. Microorganisms 2019, 7, 578. [Google Scholar] [CrossRef] [Green Version]
- Byrtusova, D.; Shapaval, V.; Holub, J.; Simansky, S.; Rapta, M.; Szotkowski, M.; Kohler, A.; Marova, I. Revealing the Potential of Lipid and β-Glucans Coproduction in Basidiomycetes Yeast. Microorganisms 2020, 8, 1034. [Google Scholar] [CrossRef]
- Walker, G.M. Yeasts. In Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 478–491. [Google Scholar]
- Mannazu, I.; Landolfo, S.; Da Silva, T.L.; Buzzini, P. Red yeasts and carotenoid production: Outlining a future for non-conventional yeasts of biotechnological interest. World J. Microbiol. Biotechnol. 2015, 31, 1665–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szotkowski, M.; Holub, J.; Šimanský, S.; Hubačová, K.; Hladká, D.; Němcová, A.; Marová, I. Production of Enriched Sporidiobolus sp. Yeast Biomass Cultivated on Mixed Coffee Hydrolyzate and Fat/Oil Waste Materials. Microorganisms 2021, 9, 1848. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Chen, Q.; Wei, C.; Hu, R.; Long, Y.; Zong, Y.; Chu, Z. Comparison of the effect of extraction methods on the quality of green coffee oil from Arabica coffee beans: Lipid yield, fatty acid composition, bioactive components, and antioxidant activity. Ultrason. Sonochemistry 2021, 74, 105578. [Google Scholar] [CrossRef] [PubMed]
- Loyao, A.S., Jr.; Villasica, S.L.G.; Dela Peña, P.L.L.; Go, A.W. Extraction of lipids from spent coffee grounds with non-polar renewable solvents as alternative. Ind. Crops Prod. 2018, 119, 152–161. [Google Scholar] [CrossRef]
- Passadis, K.; Fragoulis, V.; Stoumpou, V.; Novakovic, J.; Barampouti, E.M.; Mai, S.; Moustakas, K.; Malamis, D.; Loizidou, M. Study of Valorisation Routes of Spent Coffee Grounds. Waste Biomass Valor. 2020, 11, 5295–5306. [Google Scholar] [CrossRef]
- Bautista, L.F.; Vicente, G.; Rodríguez, R.; Pacheco, M. Optimisation of FAME production from waste cooking oil for biodiesel use. Biomass Bioen. 2009, 33, 862–872. [Google Scholar] [CrossRef]
- Paul, S.; Mittal, G.S. Dynamics of fat/oil degradation during frying based on optical properties. J. Food Eng. 1996, 30, 389–403. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Dimou, A.; Fakas, S.; Diamantopoulou, P.; Philippoussis, A.; Galiotou-Panayotou, M.; Aggelis, G. Biotechnological conversion of waste cooking olive oil into lipid-rich biomass using Aspergillus and Penicillium strains. J. Appl. Microbiol. 2011, 110, 1138–1150. [Google Scholar] [CrossRef] [PubMed]
- Spalvins, K.; Geiba, Z.; Kusnere, Z.; Blumberga, D. Waste cooking oil as substrate for single cell protein production by yeast Yarrowia lipolytica. Environ. Clim. Technol. 2020, 24, 457–469. [Google Scholar] [CrossRef]
- Petrik, S.; Hároniková, A.; Márová, I.; Kostovová, I.; Breierová, E. Production of biomass, carotenoids and other lipid metabolites by several red yeast strains cultivated on waste glycerol from biofuel production-a comparative screening study. Ann. Microbiol. 2013, 63, 1537–1551. [Google Scholar] [CrossRef]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol metabolism and transport in yeast and fungi: Established knowledge and ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [CrossRef] [Green Version]
- Juarez, G.F.Y.; Pabilona, K.B.C.; Manlangit, K.B.L.; Go, A.W. Direct Dilute Acid Hydrolysis of Spent Coffee Grounds: A New Approach in Sugar and Lipid Recovery. Waste Biomass Valorization 2018, 9, 235–246. [Google Scholar] [CrossRef]
- Szotkowski, M.; Holub, J.; Simansky, S.; Hubacova, K.; Sikorova, P.; Marinicova, V.; Nemcova, A.; Marova, I. Bioreactor co-cultivation of high lipid and carotenoid producing yeast Rhodotorula kratochvilovae and several microalgae under stress. Microorganisms 2021, 9, 1160. [Google Scholar] [CrossRef] [PubMed]
- Byrtusova, D.; Szotkowski, M.; Kurowska, K.; Shapaval, V.; Marova, I. Rhodotorula kratochvilovae CCY 20-2-26—The source of multifunctional metabolites. Microorganisms 2021, 9, 1280. [Google Scholar] [CrossRef]
- Donzella, S.; Serra, I.; Fumagalli, A.; Pellegrino, L.; Mosconi, G.; Scalzo, R.L.; Compagno, C. Recycling industrial food wastes for lipid production by oleaginous yeasts Rhodosporidiobolus azoricus and Cutaneotrichosporon oleaginosum. Biotechnol. Biofuels Bioprod. 2022, 15, 51. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Alotaibi, M.K.H.; Li, L.; Abomohra, A.E.-F. Enhanced waste glycerol recycling by yeast for efficient biodiesel production: Towards waste biorefinery. Biomass Bioenergy 2022, 159, 106410. [Google Scholar] [CrossRef]
- Nemcova, A.; Szotkowski, M.; Samek, O.; Caganova, L.; Sipiczki, M.; Marova, I. Use of Waste Substrates for the Lipid Production by Yeasts of the Genus Metschnikowia—Screening Study. Microorganisms 2021, 9, 2295. [Google Scholar] [CrossRef] [PubMed]
- Drzymala, K.; Mironczuk, A.M.; Pietrzak, W.; Dobrowolski, A. Rye and Oat Agricultural Wastes as Substrate Candidates for Biomass Production of the Non-Conventional Yeast Yarrowia lipolytica. Sustainability 2020, 12, 7704. [Google Scholar] [CrossRef]
- Rubio, F.T.V.; Haminiuk, C.W.I.; Martelli-Tosi, M.; da Silva, M.P.; Makimori, G.Y.F.; Favaro-Trindade, C.S. Utilization of grape pomaces and brewery waste Saccharomyces cerevisiae for the production of bio-based microencapsulated pigments. Food Res. Int. 2020, 136, 109470. [Google Scholar] [CrossRef]
- Holub, J.; Szotkowski, M.; Chujanov, O.; Spackova, D.; Sniegonova, P.; Marova, I. Production of enriched biomass by carotenogenic yeasts cultivated on by-roducts of poultry processing—A screening study. Microorganisms 2022, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Linder, T. Making the case for edible microorganisms as an integral part of a more sustainable and resilient food production system. Food Secur. 2019, 11, 265–278. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Media Type | Explanation |
|---|---|
| Control | Control glycerol media |
| CO | Media with a waste coffee oil as sole carbon source |
| FO | Media with a waste frying oil as sole carbon source |
| Fat | Media with a mixed waste animal fat as sole carbon source |
| CO+Fat | Media with a combination coffee oil:animal fat 50:50 as carbon source |
| FO+Fat | Media with a combination frying oil:animal fat 50:50 as carbon source |
| FO+Gly10 | Media with a combination frying oil:glycerol 90:10 as a carbon source |
| CO+Gly10 | Media with a combination coffee oil:glycerol 90:10 as a carbon source |
| Fat+Gly10 | Media with a combination animal fat:glycerol 90:10 as a carbon source |
| FO+Gly25 | Media with a combination frying oil:glycerol 75:25 as a carbon source |
| CO+Gly25 | Media with a combination coffee oil:glycerol 75:25 as a carbon source |
| Fat+Gly25 | Media with a combination animal fat:glycerol 75:25 as a carbon source |
| (I) | Lipase induction media—yeast inoculum culture was cultivated with the addition of oil to induce the lipase production |
| Parameters | Values |
|---|---|
| Media volume | 5.25 L |
| Stirring | 300–800 rpm—regulated by oxygen consumption |
| pH | 6.5 |
| pO2 | 30% |
| Temperature | 25 °C |
| Aeration | 4 L per minute |
| Illumination | 200 μmol·m2·s−1 of photons |
| Inoculation ratio | 1:10 |
| Lipid waste feeding | regulated by oxygen consumption |
| Sample Name | Betacarotene | Torularhodin | Torulene | Total Carotenoids | Ubiquinone | Ergosterol |
|---|---|---|---|---|---|---|
| Control | 0.269 ± 0.019 | 1.352 ± 0.098 | 0 ± 0 | 1.705 ± 0.121 | 4.940 ± 0.531 | 3.239 ± 0.294 |
| F.O. | 0.013 ± 0.001 | 0.077 ± 0.005 | 0 ± 0 | 0.081 ± 0.006 | 7.427 ± 0.672 | 2.987 ± 0.291 |
| C.O. | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.035 ± 0.002 | 3.555 ± 0.274 | 4.531 ± 0.385 |
| Fat | 0.170 ± 0.012 | 2.354 ± 0.208 | 0.113 ± 0.008 | 2.744 ± 0.216 | 5.426 ± 0.392 | 3.621 ± 0.312 |
| FO+Fat | 0.124 ± 0.009 | 1.496 ± 0.107 | 0.118 ± 0.008 | 1.897 ± 0.143 | 5.797 ± 0.675 | 3.607 ± 0.378 |
| CO+Fat | 0.132 ± 0.009 | 2.916 ± 0.215 | 0.114 ± 0.008 | 3.212 ± 0.288 | 4.438 ± 0.371 | 3.985 ± 0.432 |
| Control (I) | 0.229 ± 0.016 | 1.479 ± 0.122 | 0.126 ± 0.009 | 1.922 ± 0.170 | 5.940 ± 0.716 | 3.457 ± 0.351 |
| F.O. (I) | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.209 ± 0.015 | 2.401 ± 0.175 | 2.846 ± 0.255 |
| C.O. (I) | 0.110 ± 0.008 | 2.813 ± 0.199 | 0.116 ± 0.008 | 3.058 ± 0.215 | 3.481 ± 0.257 | 4.462 ± 0.478 |
| Fat (I) | 0.098 ± 0.007 | 2.557 ± 0.197 | 0.139 ± 0.010 | 4.930 ± 0.560 | 5.260 ± 0.547 | 3.728 ± 0.322 |
| FO+Fat (I) | 0.141 ± 0.010 | 1.986 ± 0.142 | 0.150 ± 0.011 | 2.471 ± 0.232 | 7.127 ± 0.714 | 3.980 ± 0.359 |
| CO+Fat (I) | 0.120 ± 0.008 | 2.997 ± 0.282 | 0.120 ± 0.009 | 3.334 ± 0.273 | 5.504 ± 0.570 | 3.991 ± 0.437 |
| Sample Name | Betacarotene | Torularhodin | Torulene | Total Carotenoids | Ubiquinone | Ergosterol |
|---|---|---|---|---|---|---|
| Control | 0.216 ± 0.016 | 1.357 ± 0.097 | 0.110 ± 0.008 | 4.420 ± 0.396 | 1.719 ± 0.139 | 2.787 ± 0.243 |
| FO+G10 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.202 ± 0.014 | 4.754 ± 0.360 | 3.594 ± 0.323 |
| FO+G25 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.014 ± 0.001 | 6.808 ± 0.858 | 3.336 ± 0.240 |
| CO+G10 | 0.090 ± 0.006 | 1.191 ± 0.088 | 0 ± 0 | 1.933 ± 0.171 | 0.72 ± 0.0530 | 3.771 ± 0.393 |
| CO+G25 | 0.205 ± 0.015 | 1.880 ± 0.165 | 0.114 ± 0.008 | 2.352 ± 0.171 | 8.486 ± 0.908 | 3.303 ± 0.319 |
| Fat+G10 | 0.133 ± 0.009 | 2.275 ± 0.210 | 0.125 ± 0.009 | 2.567 ± 0.216 | 4.462 ± 0.390 | 3.381 ± 0.241 |
| Fat+G25 | 0.220 ± 0.016 | 1.480 ± 0.115 | 0.109 ± 0.008 | 1.912 ± 0.167 | 4.738 ± 0.443 | 3.804 ± 0.380 |
| FO+Fat+G10 | 0.238 ± 0.017 | 2.186 ± 0.190 | 0.122 ± 0.009 | 2.808 ± 0.233 | 9.262 ± 1.427 | 4.318 ± 0.321 |
| FO+Fat+G25 | 0.173 ± 0.012 | 1.180 ± 0.084 | 0 ± 0 | 1.468 ± 0.118 | 8.021 ± 1.090 | 3.417 ± 0.306 |
| CO+Fat+G10 | 0.217 ± 0.015 | 2.241 ± 0.179 | 0.123 ± 0.009 | 2.681 ± 0.205 | 6.547 ± 0.684 | 3.801 ± 0.402 |
| CO+Fat+G25 | 0.296 ± 0.021 | 2.539 ± 0.210 | 0.109 ± 0.008 | 3.058 ± 0.256 | 7.381 ± 0.590 | 3.894 ± 0.360 |
| Sample Name | Betacarotene | Torularhodin | Torulene | Total Carotenoids | Ubiquinone | Ergosterol |
|---|---|---|---|---|---|---|
| Control | 0.262 ± 0.019 | 2.745 ± 0.209 | 0.210 ± 0.015 | 4.961 ± 0.436 | 3.371 ± 0.308 | 1.881 ± 0.144 |
| FO+G10 | 0.138 ± 0.010 | 2.692 ± 0.223 | 0.136 ± 0.010 | 2.948 ± 0.233 | 2.024 ± 0.155 | 6.059 ± 0.602 |
| FO+G25 | 0.315 ± 0.022 | 5.604 ± 0.686 | 0.147 ± 0.010 | 6.091 ± 0.720 | 2.705 ± 0.224 | 4.667 ± 0.503 |
| CO+G10 | 0.200 ± 0.014 | 4.182 ± 0.320 | 0.152 ± 0.011 | 4.544 ± 0.522 | 3.741 ± 0.323 | 4.464 ± 0.401 |
| CO+G25 | 0.365 ± 0.026 | 5.395 ± 0.591 | 0.158 ± 0.011 | 5.957 ± 0.755 | 2.56 ± 0.206 | 5.528 ± 0.599 |
| Fat+G10 | 0.409 ± 0.030 | 7.213 ± 1.001 | 0 ± 0 | 7.904 ± 1.016 | 4.785 ± 0.336 | 7.758 ± 0.850 |
| Fat+G25 | 0.351 ± 0.025 | 5.380 ± 0.431 | 0 ± 0 | 6.109 ± 0.577 | 5.142 ± 0.515 | 6.195 ± 0.588 |
| FO+Fat+G10 | 0.481 ± 0.035 | 7.806 ± 0.806 | 0.119 ± 0.008 | 8.611 ± 0.896 | 4.971 ± 0.376 | 7.134 ± 0.941 |
| FO+Fat+G25 | 0.746 ± 0.058 | 10.232 ± 1.369 | 0.139 ± 0.010 | 11.256 ± 0.997 | 3.964 ± 0.394 | 6.854 ± 0.751 |
| CO+Fat+G10 | 0.385 ± 0.028 | 6.455 ± 0.754 | 0.106 ± 0.007 | 7.084 ± 0.933 | 10.224 ± 1.665 | 7.114 ± 0.647 |
| CO+Fat+G25 | 0.637 ± 0.046 | 8.480 ± 0.802 | 0.129 ± 0.009 | 10.912 ± 1.164 | 9.532 ± 1.289 | 7.739 ± 1.076 |
| Sample Name | Betacarotene | Torularhodin | Torulene | Total Carotenoids | Ubiquinone | Ergosterol |
|---|---|---|---|---|---|---|
| Control | 0.467 ± 0.035 | 2.650 ± 0.234 | 0.118 ± 0.008 | 6.484 ± 0.651 | 5.479 ± 0.595 | 2.318 ± 0.213 |
| F.O. | 0.087 ± 0.006 | 0.745 ± 0.054 | 0 ± 0 | 0.843 ± 0.064 | 8.091 ± 0.802 | 4.703 ± 0.530 |
| C.O. | 0.446 ± 0.032 | 6.091 ± 0.617 | 0.144 ± 0.010 | 6.735 ± 0.549 | 2.175 ± 0.192 | 7.080 ± 0.776 |
| Fat | 0.225 ± 0.016 | 4.017 ± 0.293 | 0.132 ± 0.009 | 4.423 ± 0.382 | 2.409 ± 0.186 | 7.097 ± 0.766 |
| FO+Fat | 0.198 ± 0.014 | 3.745 ± 0.382 | 0.096 ± 0.007 | 4.101 ± 0.349 | 3.051 ± 0.282 | 7.488 ± 0.880 |
| CO+Fat | 0.356 ± 0.026 | 6.526 ± 0.534 | 0.131 ± 0.009 | 7.597 ± 0.987 | 2.881 ± 0.266 | 7.720 ± 0.566 |
| Control (I) | 0.602 ± 0.044 | 3.339 ± 0.333 | 0.142 ± 0.010 | 4.382 ± 0.336 | 4.391 ± 0.388 | 2.987 ± 0.273 |
| F.O. (I) | 0.001 ± 0 | 0.903 ± 0.069 | 0.099 ± 0.007 | 1.367 ± 0.097 | 5.653 ± 0.640 | 5.066 ± 0.539 |
| C.O. (I) | 0.408 ± 0.029 | 6.377 ± 0.656 | 0.161 ± 0.011 | 6.966 ± 0.940 | 4.606 ± 0.477 | 6.558 ± 0.861 |
| Fat (I) | 0.424 ± 0.030 | 8.688 ± 1.332 | 0.120 ± 0.008 | 9.323 ± 0.663 | 4.917 ± 0.350 | 8.644 ± 0.744 |
| FO+Fat (I) | 0.357 ± 0.026 | 6.144 ± 0.587 | 0.136 ± 0.010 | 7.026 ± 0.888 | 3.661 ± 0.272 | 8.023 ± 0.933 |
| CO+Fat (I) | 0.415 ± 0.030 | 7.526 ± 0.577 | 0.138 ± 0.012 | 8.105 ± 1.011 | 2.940 ± 0.282 | 8.393 ± 1.197 |
| Sample Name | Betacarotene | Torularhodin | Torulene | Total Carotenoids | Ubiquinone | Ergosterol | Tocopherol |
|---|---|---|---|---|---|---|---|
| 8 H | 0.245 ± 0.018 | 10.313 ± 1.144 | 0.115 ± 0.008 | 10.714 ± 1.63 | 5.135 ± 0.443 | 9.064 ± 0.919 | 0.105 ± 0.085 |
| 16H | 0.186 ± 0.013 | 4.340 ± 0.460 | 0.141 ± 0.010 | 4.754 ± 0.375 | 3.122 ± 0.222 | 5.503 ± 0.598 | 0.142 ± 0.103 |
| 24 H | 0.204 ± 0.014 | 5.918 ± 0.641 | 0.155 ± 0.011 | 6.330 ± 0.577 | 12.137 ± 1.255 | 6.430 ± 0.589 | 0.245 ± 0.153 |
| 40 H | 0.176 ± 0.013 | 5.355 ± 0.547 | 0.102 ± 0.007 | 5.648 ± 0.540 | 4.747 ± 0.419 | 6.520 ± 0.521 | 2.416 ± 0.421 |
| 48 H | 0.232 ± 0.016 | 7.238 ± 0.948 | 0.105 ± 0.007 | 7.614 ± 0.719 | 14.225 ± 2.380 | 7.432 ± 0.937 | 3.042 ± 0.503 |
| 56 H | 0.242 ± 0.017 | 7.648 ± 0.943 | 0.121 ± 0.009 | 8.046 ± 0.836 | 13.306 ± 1.788 | 9.165 ± 1.085 | 3.412 ± 0.548 |
| 72 H | 0.176 ± 0.013 | 6.804 ± 0.882 | 0.108 ± 0.008 | 7.107 ± 0.518 | 4.572 ± 0.513 | 7.705 ± 0.687 | 5.487 ± 0.603 |
| 96 H | 0.143 ± 0.010 | 1.592 ± 0.119 | 0.129 ± 0.009 | 8.597 ± 0.191 | 6.071 ± 0.749 | 6.573 ± 0.761 | 8.412 ± 0.640 |
| 120 H | 0.210 ± 0.015 | 9.386 ± 0.786 | 0.124 ± 0.009 | 9.752 ± 0.774 | 11.762 ± 1.647 | 9.359 ± 0.963 | 9.031 ± 0.584 |
| 144 H | 0.258 ± 0.018 | 10.349 ± 1.647 | 0.111 ± 0.008 | 10.757 ± 1.104 | 2.835 ± 0.269 | 9.783 ± 1.198 | 8.716 ± 0.706 |
| 168 H | 0.212 ± 0.015 | 8.646 ± 1.293 | 0.123 ± 0.009 | 9.007 ± 0.846 | 12.806 ± 1.276 | 8.522 ± 1.011 | 9.203 ± 0.489 |
| Sample Name | Betacarotene | Torularhodin | Torulene | Total Carotenoids | Ubiquinone | Ergosterol | Tocopherol |
|---|---|---|---|---|---|---|---|
| 4 H | 0.345 ± 0.024 | 0.426 ± 0.031 | 5.125 ± 0.587 | 6.512 ± 0.812 | 7.592 ± 0.873 | 8.042 ± 1.181 | 0.105 ± 0.085 |
| 14H | 0.286 ± 0.020 | 0.314 ± 0.022 | 5.122 ± 0.567 | 6.125 ± 0.436 | 8.195 ± 0.846 | 7.159 ± 0.650 | 0.142 ± 0.103 |
| 24 H | 0.302 ± 0.021 | 0.928 ± 0.071 | 6.152 ± 0.591 | 7.412 ± 0.799 | 5.599 ± 0.545 | 6.865 ± 0.880 | 0.245 ± 0.153 |
| 36 H | 0.284 ± 0.020 | 0.403 ± 0.030 | 6.243 ± 0.805 | 7.648 ± 0.960 | 6.952 ± 0.934 | 7.592 ± 0.711 | 2.416 ± 0.421 |
| 48 H | 0.262 ± 0.018 | 0.215 ± 0.015 | 7.614 ± 0.566 | 8.612 ± 1.196 | 7.020 ± 0.823 | 8.456 ± 1.145 | 3.042 ± 0.503 |
| 60 H | 0.413 ± 0.030 | 0.476 ± 0.033 | 7.046 ± 0.733 | 8.621 ± 0.719 | 6.562 ± 0.529 | 9.295 ± 1.063 | 3.412 ± 0.548 |
| 72 H | 0.125 ± 0.009 | 0.701 ± 0.052 | 7.107 ± 0.797 | 8.462 ± 1.284 | 7.520 ± 0.628 | 8.952 ± 0.941 | 5.487 ± 0.603 |
| 96 H | 0.416 ± 0.029 | 0.562 ± 0.041 | 8.513 ± 1.029 | 10.302 ± 1.699 | 8.195 ± 0.791 | 9.620 ± 0.920 | 8.412 ± 0.640 |
| 120 H | 0.713 ± 0.054 | 0.345 ± 0.024 | 8.925 ± 0.969 | 10.513 ± 1.602 | 10.52 ± 1.587 | 9.880 ± 0.901 | 9.031 ± 0.584 |
| 146 H | 0.842 ± 0.060 | 0.379 ± 0.028 | 8.012 ± 0.581 | 9.715 ± 1.062 | 9.520 ± 1.320 | 10.295 ± 1.66 | 8.716 ± 0.706 |
| 168 H | 0.453 ± 0.032 | 0.418 ± 0.031 | 9.146 ± 1.077 | 10.514 ± 1.425 | 10.195 ± 0.871 | 10.952 ± 1.38 | 9.203 ± 0.489 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szotkowski, M.; Plhalová, Ž.; Sniegoňová, P.; Holub, J.; Chujanov, O.; Špačková, D.; Blažková, J.; Márová, I. Conversion of Mixed Waste Food Substrates by Carotenogenic Yeasts of Rhodotorula sp. Genus. Microorganisms 2023, 11, 1013. https://doi.org/10.3390/microorganisms11041013
Szotkowski M, Plhalová Ž, Sniegoňová P, Holub J, Chujanov O, Špačková D, Blažková J, Márová I. Conversion of Mixed Waste Food Substrates by Carotenogenic Yeasts of Rhodotorula sp. Genus. Microorganisms. 2023; 11(4):1013. https://doi.org/10.3390/microorganisms11041013
Chicago/Turabian StyleSzotkowski, Martin, Žaneta Plhalová, Pavlína Sniegoňová, Jiří Holub, Oleg Chujanov, Dominika Špačková, Jana Blažková, and Ivana Márová. 2023. "Conversion of Mixed Waste Food Substrates by Carotenogenic Yeasts of Rhodotorula sp. Genus" Microorganisms 11, no. 4: 1013. https://doi.org/10.3390/microorganisms11041013
APA StyleSzotkowski, M., Plhalová, Ž., Sniegoňová, P., Holub, J., Chujanov, O., Špačková, D., Blažková, J., & Márová, I. (2023). Conversion of Mixed Waste Food Substrates by Carotenogenic Yeasts of Rhodotorula sp. Genus. Microorganisms, 11(4), 1013. https://doi.org/10.3390/microorganisms11041013