

Use of Different Nutrients to Improve the Fermentation Performances of Lactiplantibacillus pentosus OM13 during the Production of Sevillian Style Green Table Olives

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
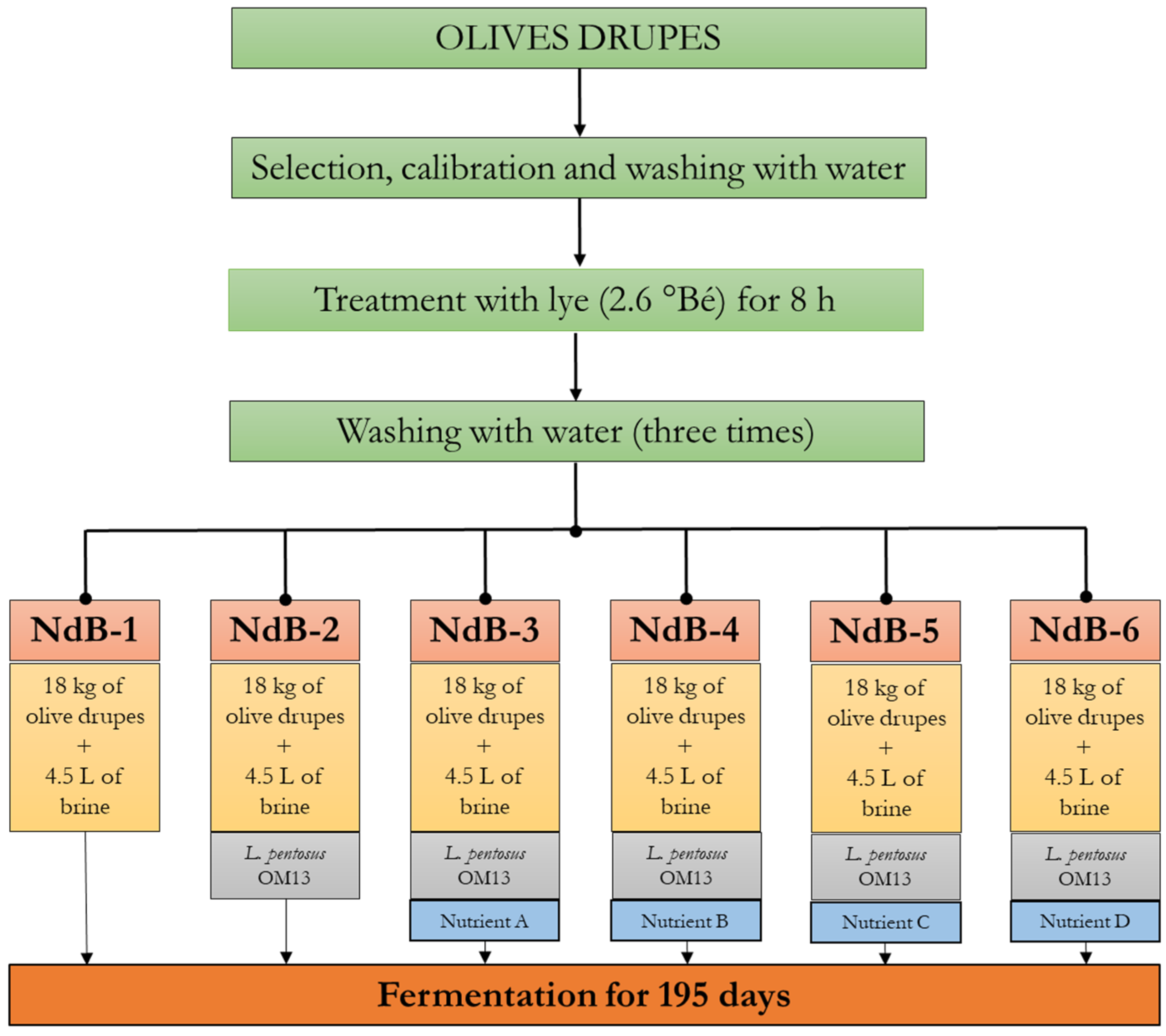
2.1. Experimental Design, Table Olive Manufacturing and Sample Collection
2.2. Physico-Chemical and Microbiological Analysis
2.3. Dominance of Lactiplantibacillus pentosus Isolates
2.4. Volatile Organic Compound Identification
2.5. Sensory Evaluation
2.6. Statistical Analysis
3. Results
3.1. Brine Acidification
3.2. Dynamics of Microbial Populations
3.2.1. Lactic Acid Bacteria
3.2.2. Yeasts
3.2.3. Enterobacteriaceae
3.2.4. Pseudomonadaceae
3.2.5. Staphylococcaceae
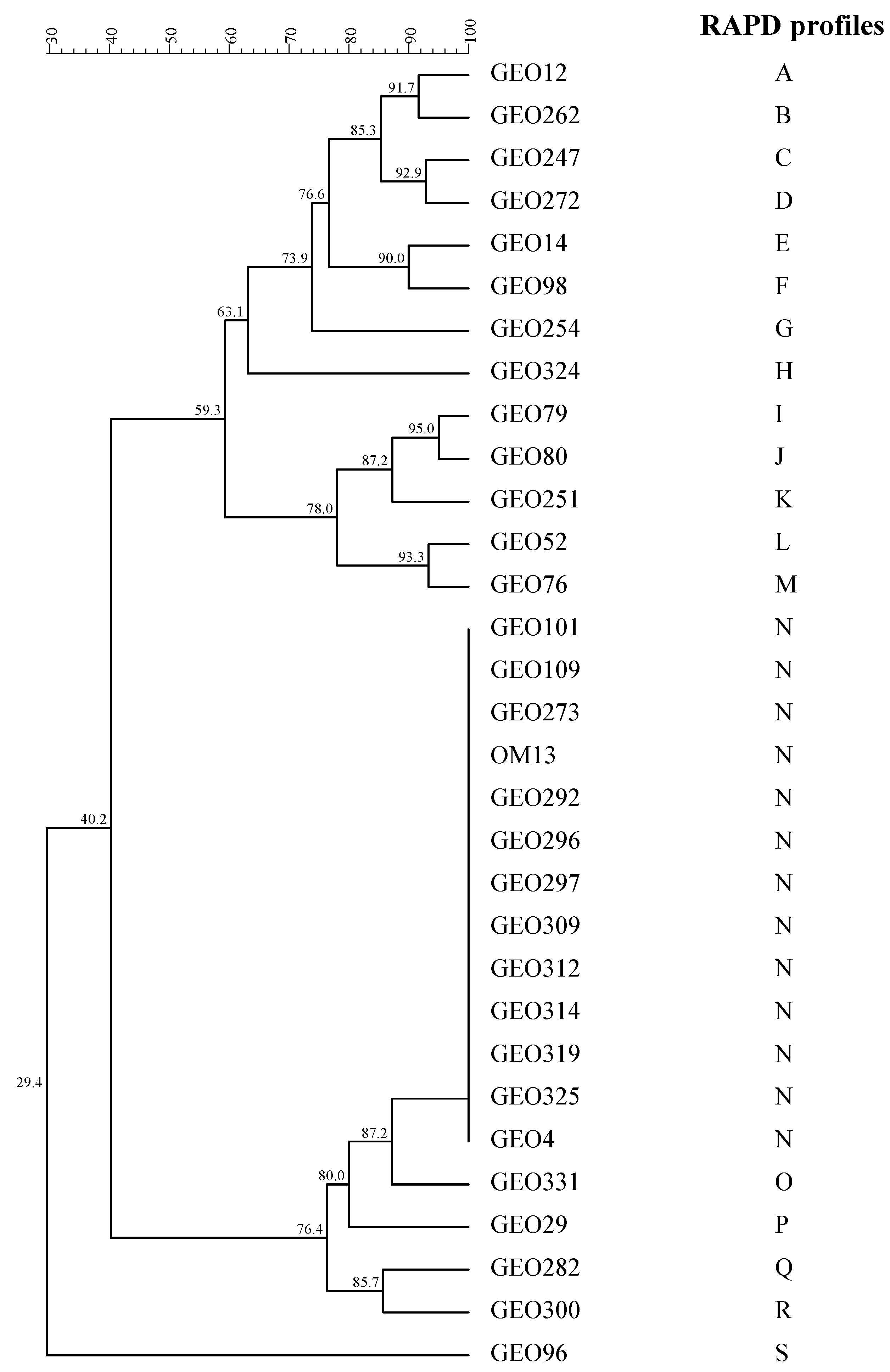
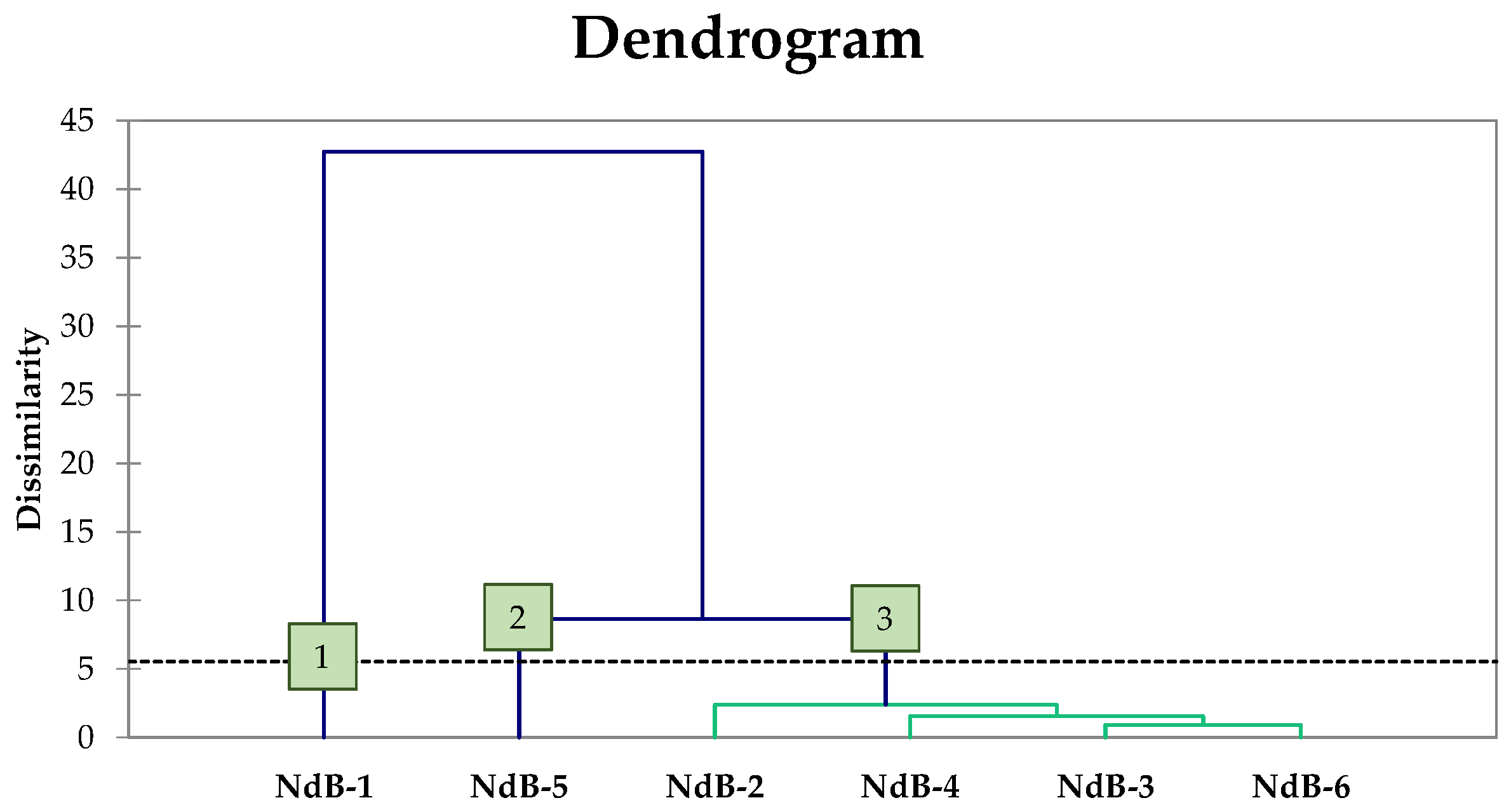
3.3. Genotypic Characterisation of Lactic Acid Bacteria
3.4. Volatile Organic Compound Composition
3.5. Sensory Characterisation of Tables Olives
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of starter cultures to table olive fermentation: An overview on the experimental studies. Front. Microbiol. 2012, 3, 248. [Google Scholar] [CrossRef] [Green Version]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.J.E. Table olive fermentation using starter cultures with multifunctional potential. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef]
- Tofalo, R.; Perpetuini, G.; Schirone, M.; Ciarrocchi, A.; Fasoli, G.; Suzzi, G.; Corsetti, A. Lactobacillus pentosus dominates spontaneous fermentation of Italian table olives. LWT Food Sci. Technol. 2014, 57, 710–717. [Google Scholar] [CrossRef]
- Portilha-Cunha, M.F.; Macedo, A.C.; Malcata, F.X. A review on adventitious lactic acid bacteria from table olives. Foods 2020, 9, 948. [Google Scholar] [CrossRef] [PubMed]
- Lanza, B.; Zago, M.; Di Marco, S.; Di Loreto, G.; Cellini, M.; Tidona, F.; Bonvini, B.; Bacceli, M.; Simone, N. Single and multiple inoculum of Lactiplantibacillus plantarum strains in table olive lab-scale fermentations. Fermentation 2020, 6, 126. [Google Scholar] [CrossRef]
- Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table olives more than a fermented food. Foods 2020, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Benincasa, C.; Muccilli, S.; Amenta, M.; Perri, E.; Romeo, F.V. Phenolic trend and hygienic quality of green table olives fermented with Lactobacillus plantarum starter culture. Food Chem. 2015, 186, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Martorana, A.; Alfonzo, A.; Gaglio, R.; Settanni, L.; Corona, O.; La Croce, F.; Vagnoli, P.; Caruso, T.; Moschetti, N.; Francesca, N. Evaluation of different conditions to enhance the performances of Lactobacillus pentosus OM13 during industrial production of Spanish-style table olives. Food Microbiol. 2017, 61, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bellis, P.; Valerio, F.; Sisto, A.; Lonigro, S.L.; Lavermicocca, P. Probiotic table olives: Microbial populations adhering on olive surface in fermentation sets inoculated with the probiotic strain Lactobacillus paracasei IMPC2.1 in an industrial plant. Int. J. Food Microbiol. 2010, 140, 6–13. [Google Scholar] [CrossRef]
- Ciafardini, G.; Zullo, B.A. Use of selected yeast starter cultures in industrial-scale processing of brined Taggiasca black table olives. Food Microbiol. 2019, 84, 103250. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.J.; Tavares, T.G.; Rocha, J.M.; Malcata, F.X.; Macedo, A.C. Cobrançosa Table Olives: Characterization of Processing Method and Lactic Acid Bacteria Profile throughout Spontaneous Fermentation. Appl. Sci. 2020, 12, 9738. [Google Scholar] [CrossRef]
- Kiai, H.; Raiti, J.; El Abbassi, A.; Hafidi, A. Chemical Profiles of Moroccan Picholine Olives and Its Brines during Spontaneous Fermentation. Int. J. Fruit Sci. 2020, 20, S1297–S1312. [Google Scholar] [CrossRef]
- Romeo, F.V. Microbiological Aspects of Table Olives. In Olive Germplasm: The Olive Cultivation, Table Olive and Olive Oil Industry in Italy; BoD–Books on Demand: Norderstedt, Germany, 2012. [Google Scholar]
- Poiana, M.; Romeo, F.V. Changes in chemical and microbiological parameters of some varieties of Sicily olives during natural fermentation. Grasas Aceites 2006, 57, 402–408. [Google Scholar] [CrossRef]
- Ruiz-Barba, J.L.; Jiménez-Díaz, R. Vitamin and amino acid requirements of Lactobacillus plantarum strains isolated from green olive fermentations. J. Appl. Bacteriol. 1994, 76, 350–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagou, E.Z.; Tassou, C.C.; Katsaboxakis, C.Z. Induced lactic acid fermentation of untreated green olives of the Conservolea cultivar by Lactobacillus pentosus. J. Sci. Food Agric. 2003, 83, 667–674. [Google Scholar] [CrossRef]
- Alfonzo, A.; Martorana, A.; Settanni, L.; Matraxia, M.; Corona, O.; Vagnoli, P.; Caruso, T.; Moschetti, G.; Francesca, N. Approaches to improve the growth of the starter lactic acid bacterium OM13 during the early stages of green Spanish-style table olive production. Grasas Aceites 2018, 69, e265. [Google Scholar] [CrossRef] [Green Version]
- Campaniello, D.; Bevilacqua, A.; D’Amato, D.; Corbo, M.R.; Altieri, C.; Sinigaglia, M. Microbial characterization of table olives processed according to Spanish and natural styles. Food Technol. Biotechnol. 2005, 43, 289–294. [Google Scholar]
- Gregersen, T. Rapid method for distinction of Gram-negative from Gram-positive bacteria. Eur. J. Appl. Microbiol. Biotechnol. 1978, 5, 123–127. [Google Scholar] [CrossRef]
- Martorana, A.; Alfonzo, A.; Settanni, L.; Corona, O.; La Croce, F.; Caruso, T.; Moschetti, G.; Francesca, N. An innovative method to produce green table olives based on “pied de cuve” technology. Food Microbiol. 2015, 50, 126–140. [Google Scholar] [CrossRef] [Green Version]
- Ruzauskas, M.; Couto, N.; Kerziene, S.; Siugzdiniene, R.; Klimiene, I.; Virgailis, M.; Pomba, C. Prevalence, species distribution and antimicrobial resistance patterns of methicillin-resistant staphylococci in Lithuanian pet animals. Acta Vet. Scand. 2015, 57, 27. [Google Scholar] [CrossRef] [Green Version]
- Rossetti, L.; Giraffa, G. Rapid identification of dairy lactic acid bacteria by M13-generated, RAPD-PCR fingerprint databases. J. Microbiol. Methods 2005, 63, 135–144. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA gene sequence analysis and multiplex PCR assay with recA gene-derived primers. Appl. Environ. Microb. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawliszyn, J. Solid Phase Microextraction: Theory and Practice; Wiley, VCH: New York, NY, USA, 1997. [Google Scholar]
- Corona, O.; Liguori, L.; Albanese, D.; Di Matteo, M.; Cinquanta, L.; Russo, P. Quality and volatile compounds in red wine atdifferent degrees of dealcoholization by membrane process. Eur. Food Res. Technol. 2019, 245, 2601–2611. [Google Scholar] [CrossRef]
- ISO 13299:2016; Sensory Analysis. Methodology. General Guidance for Establishing a Sensory Profile. ISO: Genéve, Switzerland, 2016.
- Aponte, M.; Ventorino, V.; Blaiotta, G.; Volpe, G.; Farina, V.; Avellone, G.; Lanza, C.M.; Moschetti, G. Study of green Sicilian table olive fermentations through microbiological, chemical and sensory analysis. Food Microbiol. 2010, 27, 162–170. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G.; La Croce, F.; Mazzaglia, A.; Farina, V.; Settanni, L.; Moschetti, G. Use of selected autochthonous lactic acid bacteria for Spanish-style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Martorana, A.; Alfonzo, A.; Settanni, L.; Corona, O.; La Croce, F.; Caruso, T.; Moschetti, G.; Francesca, N. Effect of the mechanical harvest of drupes on the quality characteristics of green fermented table olives. J. Sci. Food Agric. 2016, 96, 2004–2017. [Google Scholar] [CrossRef] [Green Version]
- International Olive Oil Council (IOC). Method for the Sensory Analysis of Table Olives; International Olive Oil Council: Madrid, Spain, 2011. [Google Scholar]
- Gaglio, R.; Alfonzo, A.; Francesca, N.; Corona, O.; Di Gerlando, R.; Columba, P.; Moschetti, G. Production of the Sicilian distillate “Spiritu re fascitrari” from honey by-products: An interesting source of yeast diversity. Int. J. Food Microbiol. 2017, 261, 62–72. [Google Scholar] [CrossRef]
- Tofalo, R.; Schirone, M.; Perpetuini, G.; Angelozzi, G.; Suzzi, G.; Corsetti, A. Microbiological and chemical profiles of naturally fermented table olives and brines from different Italian cultivars. Antonie Van Leeuwenhoek 2012, 102, 121–131. [Google Scholar] [CrossRef]
- Chakraborty, R.; Ganguly, R.; Hore, P.; Nath, S.; Sarkar, K. Maltodextrin: A prebiotic of choice for Lactobacillus plantarum, but not for Lactobacillus casei in combination with antibiotics. Int. J. Probiotics Prebiotics 2018, 13, 19. [Google Scholar]
- Chan, M.Z.A.; Toh, M.; Liu, S.Q. Growth, survival, and metabolic activities of probiotic Lactobacillus spp. in fermented coffee brews supplemented with glucose and inactivated yeast derivatives. Food Res. Int. 2020, 137, 109746. [Google Scholar] [CrossRef]
- Toh, M.; Liu, S.Q. Influence of commercial inactivated yeast derivatives on the survival of probiotic bacterium Lactobacillus rhamnosus HN001 in an acidic environment. AMB Express 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pozo-Bayón, M.Á.; Andújar-Ortiz, I.; Moreno-Arribas, M.V. Scientific evidences beyond the application of inactive dry yeast preparations in winemaking. Food Res. Int. 2009, 42, 754–761. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K.; Stoyancheva, G. Starch-modifying enzymes of lactic acid bacteria–structures, properties, and applications. Starch-Stärke 2013, 65, 34–47. [Google Scholar] [CrossRef]
- Lanza, B. Abnormal fermentations in table-olive processing: Microbial origin and sensory evaluation. Front. Microbiol. 2013, 4, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista-Gallego, J.; Rodríguez-Gómez, F.; Barrio, E.; Querol, A.; Garrido-Fernández, A.; Arroyo-López, F.N. Exploring the yeast biodiversity of green table olive industrial fermentations for technological applications. Int. J. Food Microbiol. 2011, 147, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, R.; Fernández, A.; Martín-Tornero, E.; Meléndez, F.; Lozano, J.; Martín-Vertedor, D. Application of Digital Olfaction for Table Olive Industry. Sensors 2022, 22, 5702. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, L.; Cano-Lamadrid, M.; Carbonell-Barrachina, Á.A.; Sendra, E.; Hernández, F. Volatile composition, sensory profile and consumer acceptability of hydrosostainable table olives. Foods 2019, 8, 470. [Google Scholar] [CrossRef] [Green Version]
- Pino, A.; De Angelis, M.; Todaro, A.; Van Hoorde, K.; Randazzo, C.L.; Caggia, C. Fermentation of Nocellara Etnea table olives by functional starter cultures at different low salt concentrations. Front. Microbiol. 2018, 9, 1125. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, C.L.; Todaro, A.; Pino, A.; Pitino, I.; Corona, O.; Caggia, C. Microbiota and metabolome during controlled and spontaneous fermentation of Nocellara Etnea table olives. Food Microbiol. 2017, 65, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Sansone-Land, A.; Takeoka, G.R.; Shoemaker, C.F. Volatile constituents of commercial imported and domestic black-ripe table olives (Olea europaea). Food Chem. 2014, 149, 285–295. [Google Scholar] [CrossRef]
- Lee, S.M.; Kitsawad, K.; Sigal, A.; Flynn, D.; Guinard, J.X. Sensory properties and consumer acceptance of imported and domestic sliced black ripe olives. J. Food Sci. 2012, 77, S439–S448. [Google Scholar] [CrossRef]
- Eshkol, N.; Sendovski, M.; Bahalul, M.; Katz-Ezov, T.; Kashi, Y.A.; Fishman, A. Production of 2-phenylethanol from L-phenylalanine by a stress tolerant Saccharomyces cerevisiae strain. J. Appl. Microbiol. 2009, 106, 534–542. [Google Scholar] [CrossRef]
- Cortés-Delgado, A.; Sánchez, A.H.; de Castro, A.; López-López, A.; Beato, V.M.; Montaño, A. Volatile profile of Spanish-style green table olives prepared from different cultivars grown at different locations. Food Res. Int. 2016, 83, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, E.; Aydeniz, B. Sensory evaluation and consumer perception of some commercial green table olives. Br. Food J. 2012, 114, 1085–1094. [Google Scholar] [CrossRef]
- López-López, A.; Moreno-Baquero, J.M.; Rodríguez-Gómez, F.; García-García, P.; Garrido-Fernández, A. Sensory assessment by consumers of traditional and potentially probiotic green Spanish-style table olives. Front. Nutr. 2018, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (g/Kg) | Nutrient A | Nutrient B | Nutrient C | Nutrient D |
|---|---|---|---|---|
| Starch | 550 | 0 | 0 | 0 |
| Sugars (glucose) | 200 | 400 | 400 | 400 |
| Maltodextrine | 0 | 500 | 500 | 500 |
| Inactivated yeast | 0 | 0 | 100 | 0 |
| Inactivated yeast rich in amino acids | 0 | 100 | 0 | 0 |
| Inactivated yeast rich in mannoproteins | 0 | 0 | 0 | 100 |
| Salt (NaCl) | 250 | 0 | 0 | 0 |
| Fermentation Days | Trials | Statistical Significance | |||||
|---|---|---|---|---|---|---|---|
| NdB-1 | NdB-2 | NdB-3 | NdB-4 | NdB-5 | NdB-6 | ||
| 0 | 8.50 ± 0.02 a | 8.50 ± 0.01 a | 8.50 ± 0.02 a | 8.50 ± 0.03 a | 8.50 ± 0.02 a | 8.50 ± 0.03 a | n.s. |
| 3 | 7.14 ± 0.06 a | 6.47 ± 0.01 b | 6.05 ± 0.12 c | 6.03 ± 0.05 c | 6.01 ± 0.12 c | 6.04 ± 0.13 c | *** |
| 6 | 6.77 ± 0.02 a | 6.02 ± 0.01 b | 5.71 ± 0.01 c | 5.68 ± 0.03 c | 5.70 ± 0.07 c | 5.66 ± 0.01 c | *** |
| 9 | 6.68 ± 0.03 a | 5.64 ± 0.02 b | 5.44 ± 0.03 c | 5.04 ± 0.03 d | 5.04 ± 0.11 d | 4.96 ± 0.13 d | *** |
| 15 | 6.29 ± 0.12 a | 5.11 ± 0.07 b | 5.00 ± 0.14 bc | 4.79 ± 0.14 d | 4.65 ± 0.09 d | 4.82 ± 0.04 cd | *** |
| 35 | 5.82 ± 0.02 a | 4.98 ± 0.06 b | 4.87 ± 0.06 b | 4.58 ± 0.11 c | 4.48 ± 0.12 c | 4.51 ± 0.02 c | *** |
| 65 | 5.49 ± 0.01 a | 4.68 ± 0.04 c | 4.79 ± 0.05 b | 4.51 ± 0.05 d | 4.39 ± 0.04 e | 4.48 ± 0.02 d | *** |
| 143 | 5.28 ± 0.12 a | 4.73 ± 0.13 b | 4.65 ± 0.06 b | 4.47 ± 0.05 c | 4.37 ± 0.05 c | 4.48 ± 0.01 c | *** |
| 195 | 4.93 ± 0.08 a | 4.54 ± 0.07 b | 4.54 ± 0.04 b | 4.47 ± 0.10 bc | 4.35 ± 0.05 c | 4.48 ± 0.09 bc | *** |
| Chemical Compounds | Trials | S.s 1 | |||||
|---|---|---|---|---|---|---|---|
| NdB-1 | Ndb-2 | NdB-3 | NdB-4 | NdB-5 | NdB-6 | ||
| Σ Acids | 2793.08 ± 302.52 b | 1337.90 ± 160.82 c | 1455.16 ± 166.83 c | 2407.99 ± 184.84 b | 2082.21 ± 161.32 d | 3973.71 ± 599.66 a | *** |
| Acetic acid | 1997.51 ± 249.70 b | 105.61 ± 9.37 d | 967.73 ± 107.37 c | 1891.65 ± 129.84 b | 1483.32 ± 97.81 b | 3190.57 ± 524.38 a | *** |
| Butanoic acid | 63.44 ± 3.81 b | 443.75 ± 68.23 a | 66.71 ± 11.13 b | 54.89 ± 3.08 b | 61.28 ± 4.02 b | 56.15 ± 5.59 b | *** |
| Hexanoic acid | 126.42 ± 0.91 c | 459.20 ± 65.82 a | 167.78 ± 13.09 c | 173.10 ± 21.52 c | 219.32 ± 19.09 b | 259.41 ± 27.46 b | *** |
| Pentanoic acid | 0.00 ± 0.00 b | 196.17 ± 0.15 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | *** |
| Propionic acid | 605.75 ± 48.10 a | 133.17 ± 17.25 e | 252.94 ± 35.23 d | 288.35 ± 30.41 d | 318.29 ± 40.40 c | 467.57 ± 42.23 b | *** |
| Σ Alcohols | 2021.64 ± 164.88 b | 1428.48 ± 88.08 c | 1510.95 ± 118.47 c | 1879.68 ± 201.88 b | 360.31 ± 30.15 d | 2425.59 ± 192.48 a | *** |
| 1-Butanol | 0.00 ± 0.00 c | 56.00 ± 6.39 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 34.19 ± 5.75 b | 0.00 ± 0.00 c | *** |
| 3-Methyl-1-butanol | 75.82 ± 11.94 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 46.09 ± 8.55 b | 72.86 ± 3.65 a | 43.51 ± 1.31 b | *** |
| 1-Hexanol | 95.36 ± 17.71 ab | 94.44 ± 5.57 ab | 28.60 ± 4.87 c | 104.89 ± 12.25 a | 0.00 ± 0.00 d | 75.34 ± 12.42 ab | *** |
| 1-Octanol | 113.27 ± 9.56 a | 71.76 ± 10.90 b | 74.49 ± 2.42 b | 91.79 ± 7.25 b | 78.83 ± 6.95 b | 115.48 ± 5.66 a | *** |
| 2-Butanol | 684.42 ± 62.44 a | 146.18 ± 19.72 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | *** |
| 3-Hexen-1-ol | 408.24 ± 11.29 a | 288.72 ± 36.59 b | 306.72 ± 11.03 b | 337.28 ± 29.69 b | 125.15 ± 11.75 c | 315.00 ± 34.98 b | *** |
| Benzyl alcohol | 644.53 ± 51.94 a | 307.50 ± 2.19 c | 403.18 ± 71.03 bc | 487.31 ± 54.81 b | 0.00 ± 0.00 d | 686.69 ± 53.32 a | *** |
| Phenylethyl alcohol | 0.00 ± 0.00 e | 463.88 ± 6.72 d | 697.96 ± 29.11 c | 812.33 ± 89.34 b | 49.28 ± 2.05 e | 1189.57 ± 84.79 a | *** |
| Σ Aldehydes | 422.53 ± 28.03 b | 346.26 ± 46.32 b | 243.96 ± 33.00 b | 301.59 ± 36.31 b | 2959.21 ± 372.18 a | 292.78 ± 28.99 b | *** |
| Benzaldehyde | 232.00 ± 3.29 b | 132.91 ± 23.60 b | 134.78 ± 21.89 b | 150.83 ± 19.53 b | 2200.07 ± 307.83 a | 145.73 ± 15.71 b | *** |
| 2,5-Dimethylbenzaldehyde | 73.25 ± 8.85 b | 31.09 ± 1.69 cd | 41.93 ± 5.17 c | 22.15 ± 2.15 d | 119.39 ± 8.88 a | 0.00 ± 0.00 e | *** |
| Benzaldehyde-3-ethyl | 52.92 ± 9.22 a | 27.96 ± 3.11 b | 30.23 ± 2.58 b | 26.73 ± 3.95 b | 18.88 ± 0.64 b | 19.50 ± 2.44 b | ** |
| Isophthaldehyde | 0.00 ± 0.00 b | 28.20 ± 0.65 b | 37.03 ± 3.36 b | 23.52 ± 0.77 b | 265.11 ± 36.69 a | 19.78 ± 1.52 b | *** |
| Nonal | 0.00 ± 0.00 d | 84.89 ± 15.27 a | 0.00 ± 0.00 d | 25.33 ± 2.03 c | 0.00 ± 0.00 d | 41.69 ± 1.30 b | *** |
| Octanal | 50.45 ± 4.01 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 335.42 ± 16.11 a | 0.00 ± 0.00 c | *** |
| Phenylacetaldehyde | 13.92 ± 2.67 d | 41.22 ± 2.01 c | 0.00 ± 0.00 e | 53.03 ± 7.88 b | 20.36 ± 2.02 d | 66.09 ± 8.03 a | *** |
| Σ Aromatic hydrocarbons | 173.47 ± 13.22 a | 136.45 ± 13.04 b | 38.11 ± 3.54 d | 105.19 ± 12.80 c | 141.73 ± 12.54 ab | 155.92 ± 10.55 ab | *** |
| Alpha-cubebene | 125.17 ± 8.62 a | 71.34 ± 6.82 b | 38.11 ± 3.54 c | 66.68 ± 7.20 b | 40.95 ± 2.48 c | 70.50 ± 3.86 b | *** |
| Styrene | 48.30 ± 4.60 d | 65.11 ± 6.22 c | 0.00 ± 0.00 e | 38.50 ± 5.60 d | 100.78 ± 10.06 a | 85.42 ± 6.69 b | *** |
| Σ Esters | 1233.08 ± 89.05 c | 6389.72 ± 453.45 a | 296.99 ± 37.22 d | 362.92 ± 54.56 d | 2209.58 ± 252.43 b | 472.18 ± 53.72 d | *** |
| Butyrolactone | 29.23 ± 3.46 b | 30.38 ± 3.01 b | 37.72 ± 4.06 b | 36.78 ± 3.80 b | 451.76 ± 50.78 a | 59.12 ± 6.54 b | *** |
| cis-3-Hexenylacetate | 106.30 ± 26.90 abc | 133.41 ± 13.45 ab | 91.06 ± 6.68 bc | 158.16 ± 32.39 a | 70.69 ± 4.93 c | 160.68 ± 12.41 a | *** |
| Ethyl cyclohexanecaboxlate | 0.00 ± 0.00 b | 3036.13 ± 289.05 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 30.04 ± 7.04 b | 19.50 ± 1.54 b | *** |
| Ethyl dihydrocinnamate | 989.23 ± 49.23 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 860.63 ± 95.32 b | 0.00 ± 0.00 c | *** |
| Ethyl hexanoate | 0.00 ± 0.00 b | 1076.68 ± 65.44 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 22.47 ± 2.95 b | 0.00 ± 0.00 b | *** |
| Ethyl pentanoate | 0.00 ± 0.00 c | 395.66 ± 34.21 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 39.70 ± 3.62 b | 0.00 ± 0.00 c | *** |
| Methyl cyclohexanoate | 0.00 ± 0.00 b | 1226.60 ± 10.51 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | *** |
| Methyl hexanoate | 0.00 ± 0.00 b | 346.67 ± 26.92 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | *** |
| Methyl hydrocinnamate | 22.69 ± 1.19 b | 14.79 ± 1.54 c | 15.12 ± 1.52 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 41.82 ± 2.59 a | *** |
| Octyl acetate | 31.85 ± 5.03 c | 16.24 ± 1.44 c | 18.80 ± 1.46 c | 30.74 ± 3.35 c | 191.47 ± 23.87 a | 108.99 ± 15.34 b | *** |
| Phenylmethyl acetate | 53.77 ± 3.23 bc | 0.00 ± 0.00 c | 92.74 ± 19.24 b | 77.72 ± 10.13 b | 503.10 ± 62.05 a | 82.07 ± 15.31 b | *** |
| Propyl butyrate | 0.00 ± 0.00 d | 113.16 ± 7.90 a | 41.56 ± 4.26 c | 59.53 ± 4.91 b | 39.73 ± 1.87 c | 0.00 ± 0.00 d | *** |
| Σ Ketones | 202.83 ± 39.42 b | 180.45 ± 11.68 b | 142.60 ± 11.33 b | 138.91 ± 21.79 b | 3782.91 ± 421.32 a | 160.74 ± 10.46 b | *** |
| 4-Ethylacetophenone | 202.83 ± 39.42 b | 180.45 ± 11.68 b | 142.60 ± 11.33 b | 138.91 ± 21.79 b | 3782.91 ± 421.32 a | 160.74 ± 10.46 b | *** |
| Σ Phenols | 4430.94 ± 300.82 b | 3052.54 ± 181.17 d | 3191.54 ± 162.06 cd | 3703.29 ± 342.62 c | 264.68 ± 38.61 e | 7300.78 ± 306.86 a | *** |
| Creosol | 4217.22 ± 278.65 b | 2302.65 ± 107.69 d | 3000.63 ± 138.77 c | 3106.17 ± 286.19 c | 23.16 ± 1.23 e | 6176.69 ± 230.55 a | *** |
| Guaiacol | 80.82 ± 14.33 d | 272.30 ± 21.75 c | 99.11 ± 16.68 d | 486.15 ± 45.37 b | 62.36 ± 9.15 d | 900.64 ± 40.43 a | *** |
| Phenol | 52.68 ± 4.12 b | 31.73 ± 4.32 b | 39.48 ± 1.51 b | 56.07 ± 7.04 b | 133.57 ± 24.88 a | 111.63 ± 22.60 a | *** |
| p-Creosol | 80.22 ± 3.72 bc | 445.86 ± 47.40 a | 52.33 ± 5.11 c | 54.90 ± 4.02 c | 45.59 ± 3.35 c | 111.82 ± 13.28 b | *** |
| Attributes | Trials 1 | |||||||
|---|---|---|---|---|---|---|---|---|
| NdB-1 | NdB-2 | NdB-3 | NdB-4 | NdB-5 | NdB-6 | SEM 2 | S.s 3 | |
| Acid | 2.35 b | 4.48 a | 3.89 a | 4.15 a | 2.37 b | 4.03 a | 0.22 | *** |
| Astringent | 1.48 b | 1.95 a | 1.81 ab | 1.47 b | 1.58 ab | 1.55 ab | 0.05 | ** |
| Bitter | 2.82 a | 3.42 a | 3.15 a | 3.19 a | 2.62 a | 2.98 a | 0.07 | n.s.4 |
| Brightness | 4.20 a | 4.55 a | 4.03 a | 4.11 a | 3.92 a | 4.19 a | 0.05 | n.s.4 |
| Crispness | 4.23 b | 7.18 a | 6.82 a | 6.95 a | 7.00 a | 6.88 a | 0.26 | * |
| Green colour intensity | 6.27 a | 5.25 a | 5.33 a | 5.35 a | 6.17 a | 5.86 a | 0.11 | n.s.4 |
| Green olive aroma | 1.83 b | 5.60 a | 5.15 a | 5.37 a | 6.13 a | 5.42 a | 0.36 | * |
| Juicy | 3.38 c | 4.38 bc | 5.33 ab | 5.49 ab | 6.18 a | 5.17 ab | 0.28 | *** |
| Off-flavours | 4.82 a | 2.19 b | 1.51 b | 1.38 b | 1.13 b | 1.27 b | 0.33 | *** |
| Off-odours | 3.27 a | 1.52 b | 1.44 b | 1.39 b | 1.17 b | 1.33 b | 0.18 | *** |
| Salt | 5.97 a | 5.03 b | 5.81 a | 4.98 b | 6.13 a | 6.46 a | 0.14 | *** |
| Sweet | 1.33 d | 2.68 bc | 2.22 c | 2.06 c | 4.32 a | 2.87 ab | 0.16 | *** |
| Overall acceptability | 3.55 c | 5.55 bc | 5.39 bc | 6.48 ab | 7.80 a | 6.07 ab | 0.30 | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfonzo, A.; Naselli, V.; Gaglio, R.; Settanni, L.; Corona, O.; La Croce, F.; Vagnoli, P.; Krieger-Weber, S.; Francesca, N.; Moschetti, G. Use of Different Nutrients to Improve the Fermentation Performances of Lactiplantibacillus pentosus OM13 during the Production of Sevillian Style Green Table Olives. Microorganisms 2023, 11, 825. https://doi.org/10.3390/microorganisms11040825
Alfonzo A, Naselli V, Gaglio R, Settanni L, Corona O, La Croce F, Vagnoli P, Krieger-Weber S, Francesca N, Moschetti G. Use of Different Nutrients to Improve the Fermentation Performances of Lactiplantibacillus pentosus OM13 during the Production of Sevillian Style Green Table Olives. Microorganisms. 2023; 11(4):825. https://doi.org/10.3390/microorganisms11040825
Chicago/Turabian StyleAlfonzo, Antonio, Vincenzo Naselli, Raimondo Gaglio, Luca Settanni, Onofrio Corona, Francesco La Croce, Paola Vagnoli, Sibylle Krieger-Weber, Nicola Francesca, and Giancarlo Moschetti. 2023. "Use of Different Nutrients to Improve the Fermentation Performances of Lactiplantibacillus pentosus OM13 during the Production of Sevillian Style Green Table Olives" Microorganisms 11, no. 4: 825. https://doi.org/10.3390/microorganisms11040825
APA StyleAlfonzo, A., Naselli, V., Gaglio, R., Settanni, L., Corona, O., La Croce, F., Vagnoli, P., Krieger-Weber, S., Francesca, N., & Moschetti, G. (2023). Use of Different Nutrients to Improve the Fermentation Performances of Lactiplantibacillus pentosus OM13 during the Production of Sevillian Style Green Table Olives. Microorganisms, 11(4), 825. https://doi.org/10.3390/microorganisms11040825