
Assessing the Impact of Different Technological Strategies on the Fate of Salmonella in Chicken Dry-Fermented Sausages by Means of Challenge Testing and Predictive Models


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Preparation
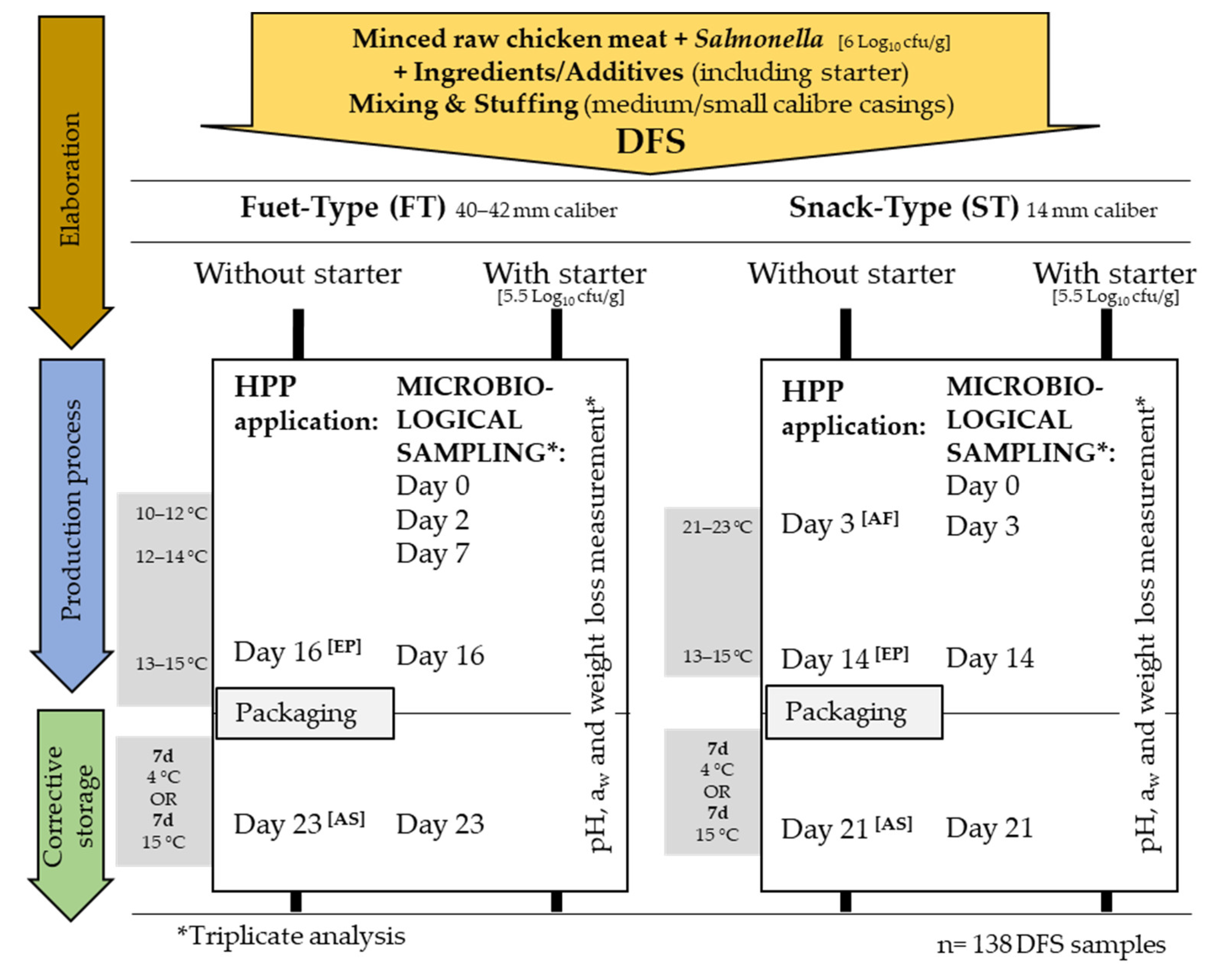
2.2. Chicken-Based DFS Preparation, Processing and Storage Conditions
2.3. High-Pressure Processing
2.4. Microbiological and Physicochemical Determinations
2.5. Statistical Analysis of Analytical Results
2.6. Simulation of the Behaviour of Salmonella
3. Results
3.1. Physicochemical Characteristics of Fuet-Type (FT) and Snack-Type (ST) DFS
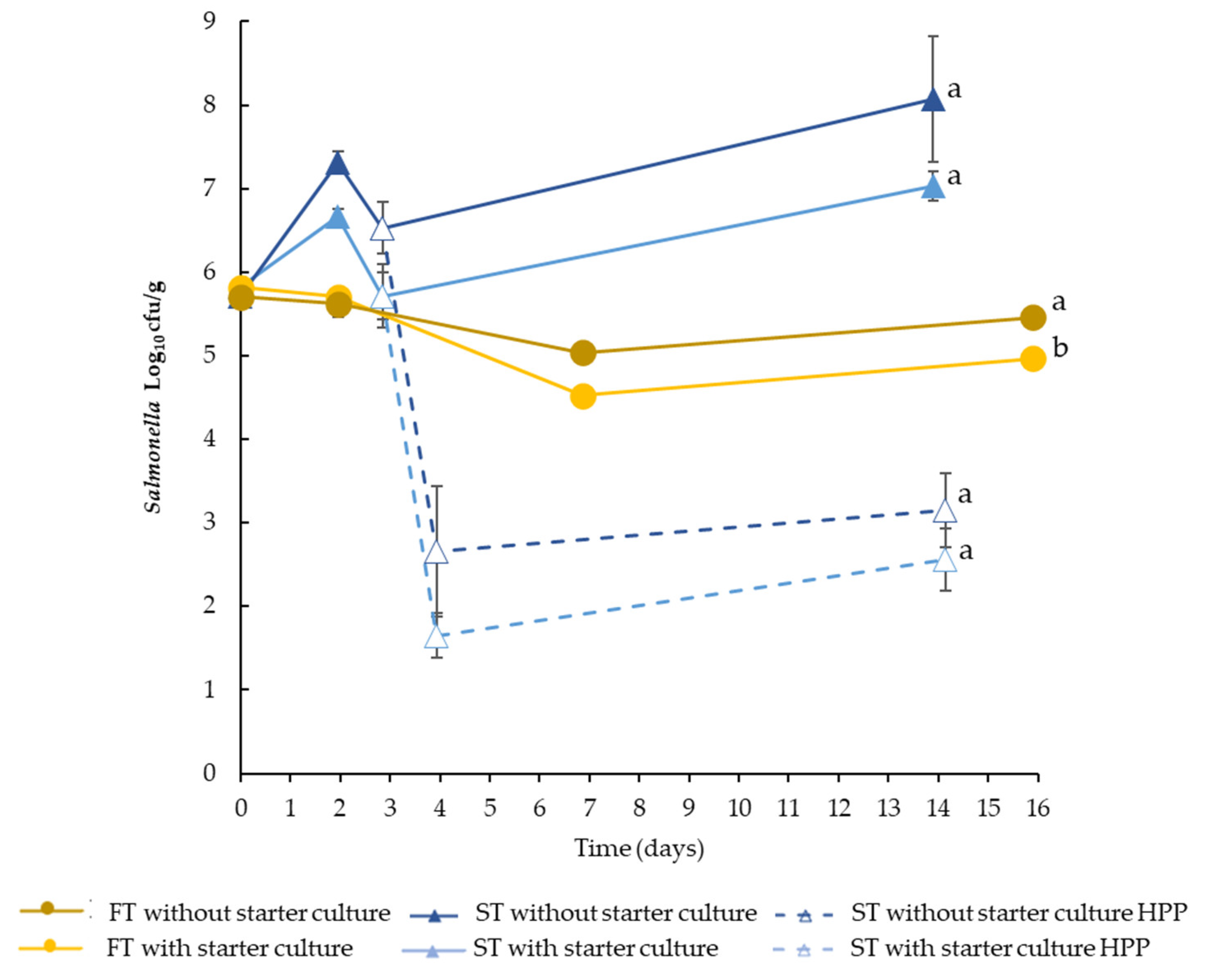
3.2. Behaviour of LAB and Salmonella during the Production Process
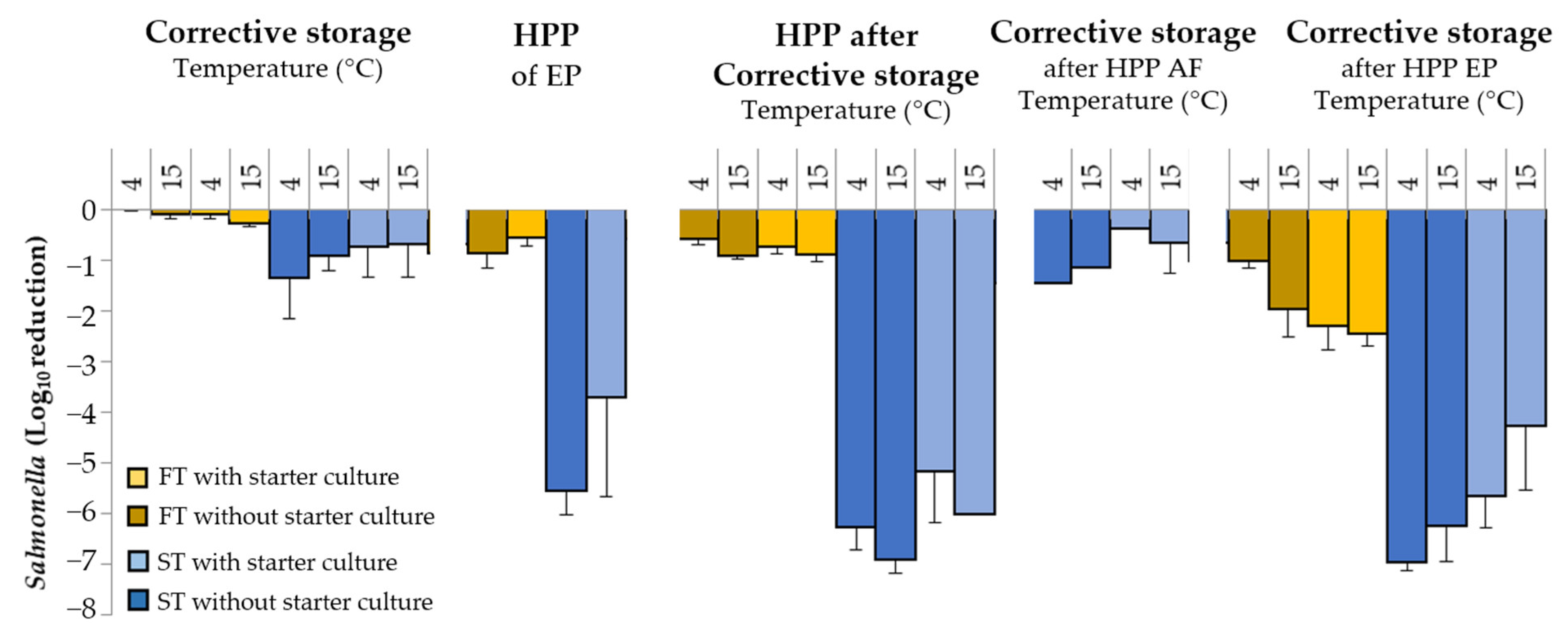
3.3. Impact of Corrective Storage and HPP on Salmonella
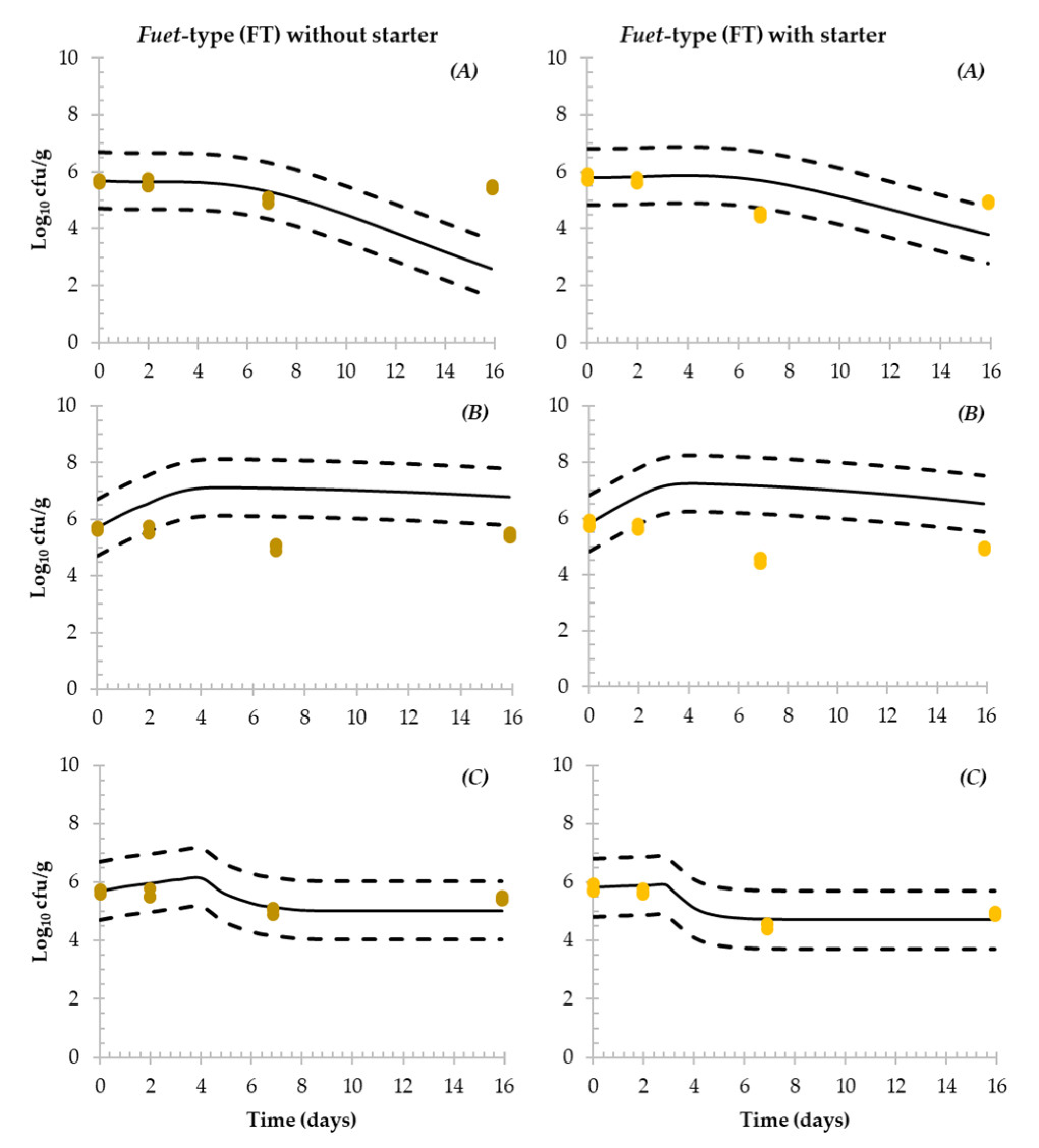
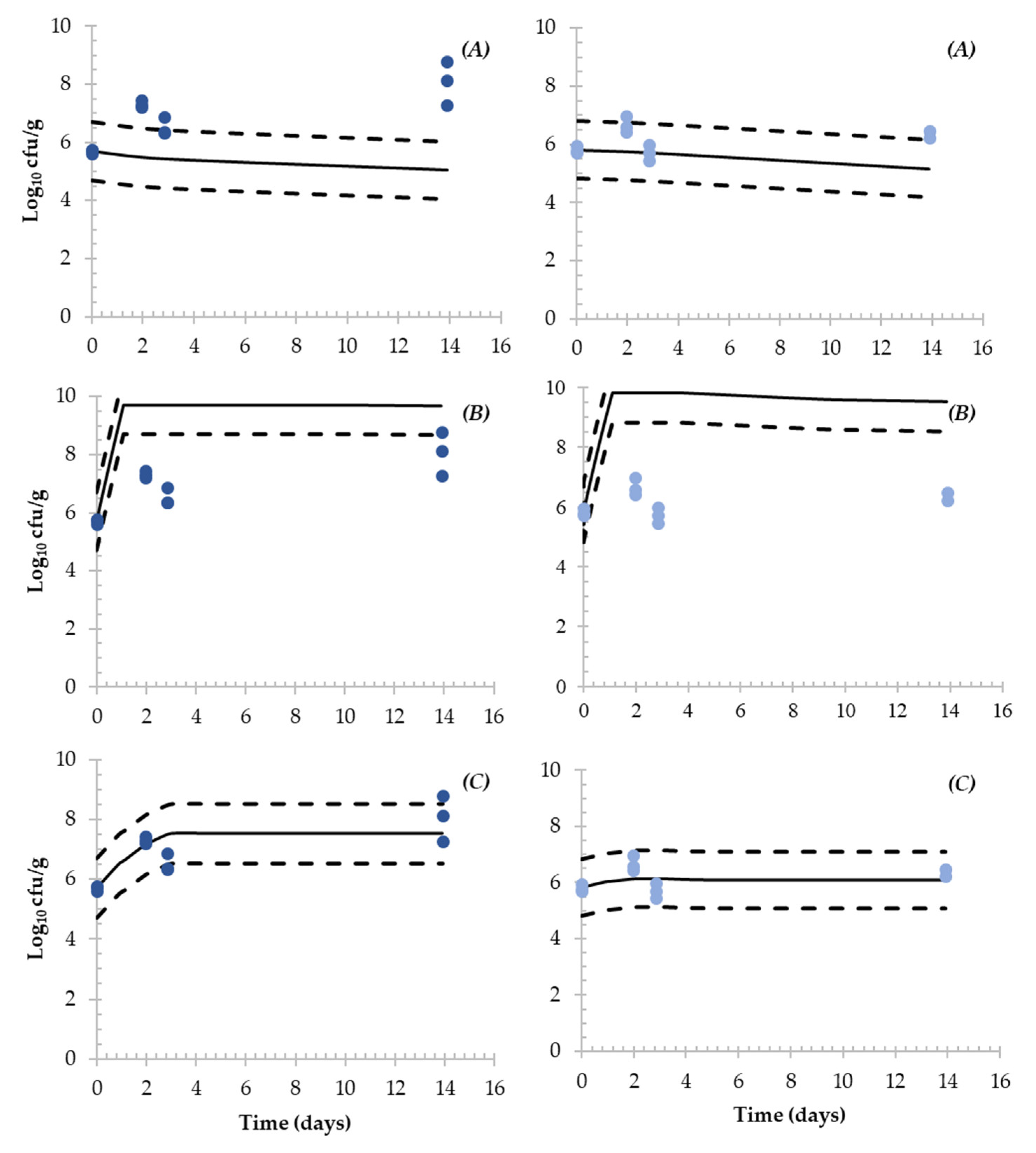
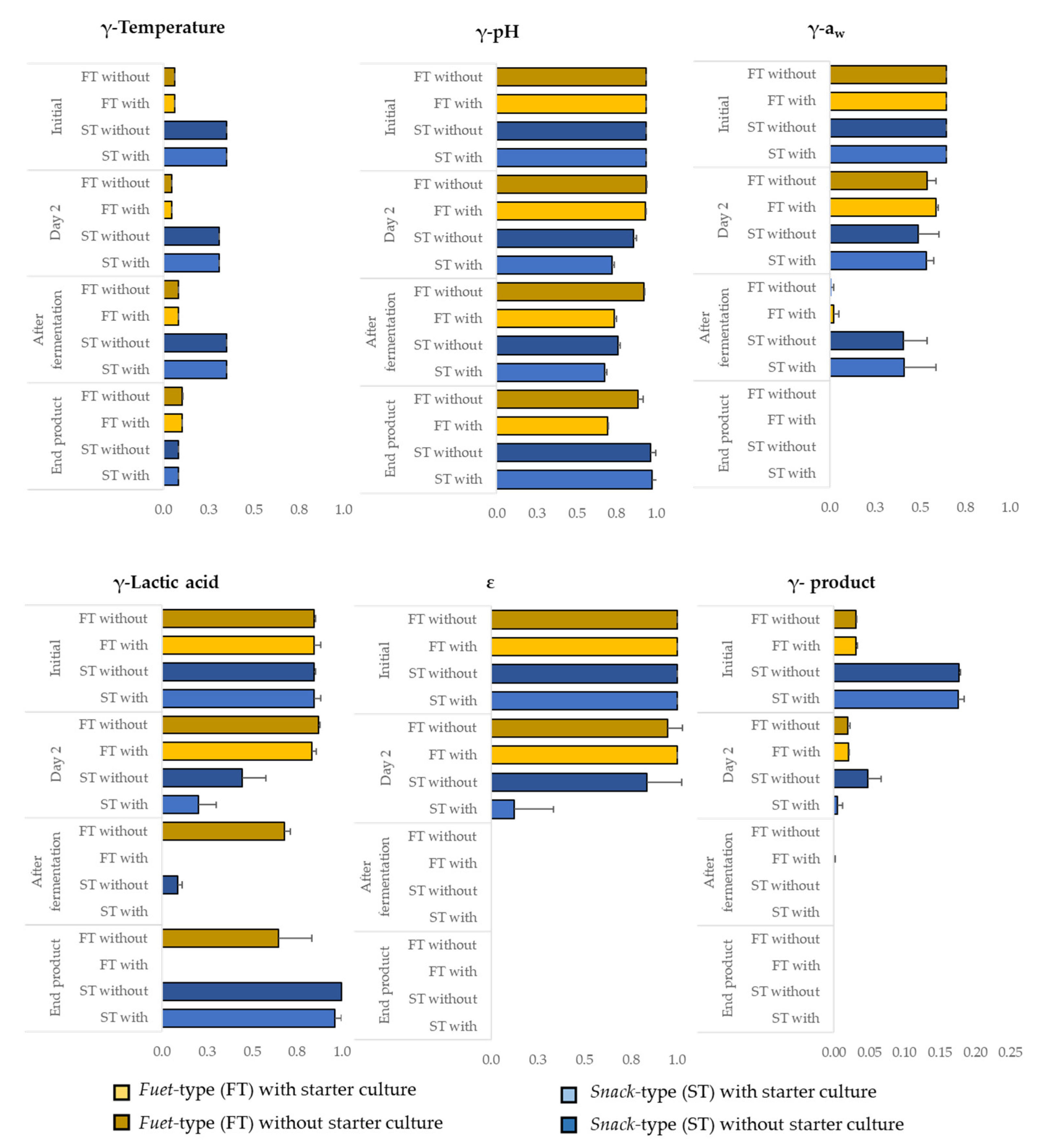
3.4. Simulation of the Fate of Salmonella during the Production of DFS
4. Discussion
4.1. Effect of Formulation and Production Process on Salmonella
4.2. Impact of Corrective Storage and HPP on Salmonella
4.3. Evaluation of Predictive Models Simulating the Fate of Salmonella in DFS
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Würtz, A.M.L.; Jakobsen, M.U.; Bertoia, M.L.; Hou, T.; Schmidt, E.B.; Willett, W.C.; Overvad, K.; Sun, Q.; Manson, J.A.E.; Hu, F.B.; et al. Replacing the Consumption of Red Meat with Other Major Dietary Protein Sources and Risk of Type 2 Diabetes Mellitus: A Prospective Cohort Study. Am. J. Clin. Nutr. 2021, 113, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Leistner, L. Basic Aspects of Food Preservation by Hurdle Technology. Int. J. Food Microbiol. 2000, 55, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Garriga, M.; Aymerich, T. Prevalence of Salmonella spp. and Listeria monocytogenes at Small-Scale Spanish Factories Producing Traditional Fermented Sausages. J. Food Prot. 2011, 74, 812–815. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA) The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [CrossRef]
- Serra-Castelló, C.; Bover-Cid, S.; Garriga, M.; Beck Hansen, T.; Gunvig, A.; Jofré, A. Risk Management Tool to Define a Corrective Storage to Enhance Salmonella Inactivation in Dry Fermented Sausages. Int. J. Food Microbiol. 2021, 346, 109160. [Google Scholar] [CrossRef]
- da Mutz, Y.S.; Rosario, D.K.A.; Paschoalin, V.M.F.; Conte-Junior, C.A. Salmonella Enterica: A Hidden Risk for Dry-Cured Meat Consumption? Crit. Rev. Food Sci. Nutr. 2020, 60, 976–990. [Google Scholar] [CrossRef]
- Jofré, A.; Aymerich, T.; Garriga, M. Improvement of the Food Safety of Low Acid Fermented Sausages by Enterocins A and B and High Pressure. Food Control 2009, 20, 179–184. [Google Scholar] [CrossRef]
- Meloni, D.; Consolati, S.G.; Mazza, R.; Mureddu, A.; Fois, F.; Piras, F.; Mazzette, R. Presence and Molecular Characterization of the Major Serovars of Listeria monocytogenes in Ten Sardinian Fermented Sausage Processing Plants. Meat Sci. 2014, 97, 443–450. [Google Scholar] [CrossRef]
- Rapid Alert System for Food and Feed (RASFF). NOTIFICATION 2021.3787. Salmonella Food Poisoning—FUET. 2021. Available online: https://webgate.ec.europa.eu/rasff-window/screen/notification/488132 (accessed on 24 January 2023).
- Public Health Agency of Canada. Public Health Notice—Outbreak of Salmonella Illnesses Linked to Filicetti Brand Italian Style mild, dry, cured sausage. 23 December 2019. Available online: https://www.canada.ca/en/public-health/services/public-health-notices/2019/outbreak-salmonella-sausages.html (accessed on 24 January 2023).
- Sams, A.R. Poultry Meat Processing, 1st ed.; CRC Press: Boca Ratón, FL, USA, 2001; pp. 1–334. [Google Scholar]
- Toldrá, F.; Hui, Y.H.; Astiasarán, I.; Sebranek, J.G.; Talon, R. Handbook of Fermented Meat and Poultry, 2nd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 1–499. [Google Scholar] [CrossRef]
- Parry-Hanson Kunadu, A.; Otwey, R.Y.; Mosi, L. Microbiological Quality and Salmonella Prevalence, Serovar Distribution and Antimicrobial Resistance Associated with Informal Raw Chicken Processing in Accra, Ghana. Food Control 2020, 118, 107440. [Google Scholar] [CrossRef]
- Regalado-Pineda, I.; Rodarte-Medina, R.; Resendiz-Nava, C.; Sainz-Garcia, C.; Castañeda-Serrano, P.; Nava, G. Three-Year Longitudinal Study: Prevalence of Salmonella Enterica in Chicken Meat Is Higher in Supermarkets than Wet Markets from Mexico. Foods 2020, 9, 264. [Google Scholar] [CrossRef]
- Marcos, B.; Aymerich, T.; Garriga, M. Evaluation of High Pressure Processing as an Additional Hurdle to Control Listeria monocytogenes and Salmonella Enterica in Low-Acid Fermented Sausages. J. Food Sci. 2005, 70, 339–344. [Google Scholar] [CrossRef]
- Heir, E.; Holck, A.L.; Omer, M.K.; Alvseike, O.; Høy, M.; Måge, I.; Axelsson, L. Reduction of Verotoxigenic Escherichia coli by Process and Recipe Optimisation in Dry-Fermented Sausages. Int. J. Food Microbiol. 2010, 141, 195–202. [Google Scholar] [CrossRef]
- Mataragas, M.; Bellio, A.; Rovetto, F.; Astegiano, S.; Greci, C.; Hertel, C.; Decastelli, L.; Cocolin, L. Quantification of Persistence of the Food-Borne Pathogens Listeria monocytogenes and Salmonella Enterica during Manufacture of Italian Fermented Sausages. Food Control 2015, 47, 552–559. [Google Scholar] [CrossRef]
- Drosinos, E.H.; Mataragas, M.; Vesković-Moračanin, S.; Gasparik-Reichardt, J.; Hadžiosmanović, M.; Alagić, D. Quantifying Nonthermal Inactivation of Listeria monocytogenes in European Fermented Sausages Using Bacteriocinogenic Lactic Acid Bacteria or Their Bacteriocins: A Case Study for Risk Assessment. J. Food Prot. 2006, 69, 2648–2663. [Google Scholar] [CrossRef]
- Jofré, A.; Serra, X. Processing of Meat Products Utilizing High Pressure. In High Pressure Processing of Food–Principles, Technology and Applications; Balasubramaniam, V., Barbosa-Cánovas, G.V., Lelieveld, H.L., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Bover-Cid, S.; Belletti, N.; Aymerich, T.; Garriga, M. Modeling the Protective Effect of aw and Fat Content on the High Pressure Resistance of Listeria monocytogenes in Dry-Cured Ham. Food Res. Int. 2015, 75, 194–199. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Belletti, N.; Garriga, M.; Aymerich, T. Model for Listeria monocytogenes Inactivation on Dry-Cured Ham by High Hydrostatic Pressure Processing. Food Microbiol. 2011, 28, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Walls, I.; Scott, V.N. Use of Predictive Microbiology in Microbial Food Safety Risk Assessment. Int. J. Food Microbiol. 1997, 36, 97–102. [Google Scholar] [CrossRef]
- Fakruddin, M.; Mazumdar, R.M.; Mannan, K.S. Bin Predictive Microbiology: Modeling Microbial Responses in Food. Ceylon J. Sci. Biological Sci. 2012, 40, 121. [Google Scholar] [CrossRef]
- Guillier, L. Predictive Microbiology Models and Operational Readiness. Procedia Food Sci. 2016, 7, 133–136. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Garriga, M. Microbiología Predictiva: Herramienta de Soporte Para La Gestión de La Seguridad y La Calidad Alimentaria. Eurocarne 2008, 18, 104–113. [Google Scholar]
- Stollewerk, K.; Jofré, A.; Comaposada, J.; Arnau, J.; Garriga, M. The Impact of Fast Drying (QDS Process(R)) and High Pressure on Food Safety of NaCl-free Processed Dry Fermented Sausages. Innov. Food Sci. Emerg. Technol. 2012, 16, 89–95. [Google Scholar] [CrossRef]
- Serra-Castelló, C.; Possas, A.; Jofré, A.; Garriga, M.; Bover-Cid, S. High-Pressure Processing Inactivation of Salmonella in Raw Pet Food for Dog is Enhanced by Acidulation with Lactic Acid. Anim. Feed Sci. Technol. 2022, 290, 115347. [Google Scholar] [CrossRef]
- Hugas, M.; Garriga, M.; Aymerich, M.T.; Monfort, J.M. Inhibition of Listeria in Dry Fermented Sausages by the Bacteriocinogenic Lactobacillus sake CTC494. J. Appl. Bacteriol. 1995, 79, 322–330. [Google Scholar] [CrossRef]
- ISO 15214:1998; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Mesophilic Lactic acid Bacteria—Colony-Count Technique at 30 Degrees C. International Organization for Standardization: Geneva, Switzerland, 1998.
- Hwang, C.A.; Porto-Fett, A.C.S.; Juneja, V.K.; Ingham, S.C.; Ingham, B.H.; Luchansky, J.B. Modeling the Survival of Escherichia Coli O157:H7, Listeria monocytogenes, and Salmonella Typhimurium during Fermentation, Drying, and Storage of Soudjouk-Style Fermented Sausage. Int. J. Food Microbiol. 2009, 129, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Pin, C.; Avendaño-Perez, G.; Cosciani-Cunico, E.; Gómez, N.; Gounadakic, A.; Nychas, G.J.; Skandamis, P.; Barker, G. Modelling Salmonella Concentration throughout the Pork Supply Chain by Considering Growth and Survival in Fluctuating Conditions of Temperature, pH and aw. Int. J. Food Microbiol. 2011, 145, S96–S102. [Google Scholar] [CrossRef]
- Coroller, L.; Jeuge, S.; Couvert, O.; Christieans, S.; Ellouze, M. Extending the Gamma Concept to Non-Thermal Inactivation: A Dynamic Model to Predict the Fate of Salmonella during the Dried Sausages Process. Food Microbiol. 2015, 45, 266–275. [Google Scholar] [CrossRef]
- Rubio, R.; Jofré, A.; Aymerich, T.; Guàrdia, M.D.; Garriga, M. Nutritionally Enhanced Fermented Sausages as a Vehicle for Potential Probiotic Lactobacilli Delivery. Meat Sci. 2014, 96, 937–942. [Google Scholar] [CrossRef]
- Zagorec, M.; Champomier-Vergès, M.C. Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products. Microorganisms 2017, 5, 56. [Google Scholar] [CrossRef]
- Sunesen, L.O.; Stahnke, L.H. Mould Starter Cultures for Dry Sausages—Selection, Application and Effects. Meat Sci. 2003, 65, 935–948. [Google Scholar] [CrossRef]
- Bruna, J.M.; Ordóñez, J.A.; Fernández, M.; Herranz, B.; De La Hoz, L. Microbial and Physico-Chemical Changes during the Ripening of Dry Fermented Sausages Superficially Inoculated with or Having Added an Intracellular Cell-Free Extract of Penicillium Aurantiogriseum. Meat Sci. 2001, 59, 87–96. [Google Scholar] [CrossRef]
- Garriga, M.; Marcos, B.; Martín, B.; Veciana-Nogués, M.T.; Bover-Cid, S.; Hugas, M.; Aymerich, T. Starter Cultures and High-Pressure Processing to Improve the Hygiene and Safety of Slightly Fermented Sausages. J. Food Prot. 2005, 68, 2341–2348. [Google Scholar] [CrossRef]
- Werlang, G.O.; Vieira, T.R.; Cardoso, M.; de Costa, E.F. Application of a Predictive Microbiological Model for Estimation of Salmonella Behavior throughout the Manufacturing Process of Salami in Environmental Conditions of Small-Scale Brazilian Manufacturers. Microb. Risk Anal. 2021, 19, 100177. [Google Scholar] [CrossRef]
- Bonilauri, P.; Grisenti, M.S.; Daminelli, P.; Merialdi, G.; Ramini, M.; Bardasi, L.; Taddei, R.; Cosciani-Cunico, E.; Dalzini, E.; Frustoli, M.A.; et al. Reduction of Salmonella spp. Populations in Italian Salami during Production Process and High Pressure Processing Treatment: Validation of Processes to Export to the U.S. Meat Sci. 2019, 157, 107869. [Google Scholar] [CrossRef] [PubMed]
- Kurrat, J.; Perkins, L. Microbiological Safety of Raw-Fermented Sausages with Glucono-Delta-Lactone. Available online: https://www.jungbunzlauer.com/fileadmin/content/_PDF/PRINT_PROJECTS/Article_facts/JBL_AR_GdL_in_meat_2019-037.pdf (accessed on 27 November 2022).
- Leistner, L. Update on Hurdle Technology for Mild and Effective Preservation of Foods. In Handbook of Food Preservation; CRC press: Boca Raton, FL, USA, 2007; pp. 895–904. [Google Scholar]
- ICMSF. Microorganisms in Foods 5. In Microbiol. Specif. food Pathog. London Blackie Acad; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1996. [Google Scholar]
- ANSES. Salmonella spp. Available online: https://www.anses.fr/en/system/files/MIC2011sa0057FiEN.pdf (accessed on 10 November 2022).
- Ferrer-Bustins, N.; Serra-Castelló, C.; Cortada, M.; Cabedo, L.; Bover-Cid, S.; Jofré, A. Salmonella Prevalence in Dry-Fermented Sausages from Catalan Producers and Meat Suppliers, towards Legislation Compliance. In Proceedings of the 7th International Congress of Meat Science and Technology, ICoMST, Krakow, Poland, 23–27 August 2021. [Google Scholar]
- Ihnot, A.M.; Roering, A.M.; Wierzba, R.K.; Faith, N.G.; Luchansky, J.B. Behavior of Salmonella Typhimurium DT104 during the Manufacture and Storage of Pepperoni. Int. J. Food Microbiol. 1998, 40, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Nissen, H.; Holck, A. Survival of Escherichia coli O157:H7, Listeria monocytogenes and Salmonella Kentucky in Norwegian Fermented, Dry Sausage. Food Microbiol. 1998, 15, 273–279. [Google Scholar] [CrossRef]
- Faith, N.G.; Wierzba, R.K.; Ihnot, A.M.; Roering, A.M.; Lorang, T.D.; Kaspar, C.W.; Luchansky, J.B. Survival of Escherichia coli O157:H7 in Full- and Reduced-Fat Pepperoni after Manufacture of Sticks, Storage of Slices at 4°C or 21°C under Air and Vacuum, and Baking of Slices on Frozen Pizza at 135, 191 and 246 °C. J. Food Prot. 1998, 61, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Santillana-Farakos, S.M.; Frank, J.F.; Schaffner, D.W. Modeling the Influence of Temperature, Water Activity and Water Mobility on the Persistence of Salmonella in Low-Moisture Foods. Int. J. Food Microbiol. 2013, 166, 280–293. [Google Scholar] [CrossRef]
- Hereu, A.; Bover-cid, S.; Garriga, M.; Aymerich, T. High Hydrostatic Pressure and Biopreservation of Dry-Cured Ham to Meet the Food Safety Objectives for Listeria monocytogenes . Int. J. Food Microbiol. 2012, 154, 107–112. [Google Scholar] [CrossRef]
- Balamurugan, S.; Gemmell, C.; Lau, A.T.Y.; Arvaj, L.; Strange, P.; Gao, A.; Barbut, S. High Pressure Processing during Drying of Fermented Sausages Can Enhance Safety and Reduce Time Required to Produce a Dry Fermented Product. Food Control 2020, 113, 107224. [Google Scholar] [CrossRef]
- Georget, E.; Sevenich, R.; Reineke, K.; Mathys, A.; Heinz, V.; Callanan, M.; Rauh, C.; Knorr, D. Inactivation of Microorganisms by High Isostatic Pressure Processing in Complex Matrices: A Review. Innov. Food Sci. Emerg. Technol. 2015, 27, 1–14. [Google Scholar] [CrossRef]
- Hayman, M.M.; Anantheswaran, R.C.; Knabel, S.J. The Effects of Growth Temperature and Growth Phase on the Inactivation of Listeria monocytogenes in Whole Milk Subject to High Pressure Processing. Int. J. Food Microbiol. 2007, 115, 220–226. [Google Scholar] [CrossRef] [PubMed]
- McClements, J.; Patterson, M.; Linton, M. The Effect of Growth Stage and Growth Temperature on High Hydrostatic Pressure Inactivation of Some Psychrotrophic Bacteria in Milk. J. Food Prot. 2001, 64, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Casadei, M.A.; Mañas, P.; Niven, G.; Needs, E.; Mackey, B.M. Role of Membrane Fluidity in Pressure Resistance of Escherichia coli NCTC 8164. Appl. Environ. Microbiol. 2002, 68, 5965–5972. [Google Scholar] [CrossRef]
- Jofré, A.; Aymerich, T.; Bover-Cid, S.; Garriga, M. Inactivation and Recovery of Listeria monocytogenes, Salmonella Enterica and Staphylococcus aureus after High Hydrostatic Pressure Treatments up to 900 MPa. Int. Microbiol. 2010, 13, 105–112. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Days) | Fuet-Type | Snack-Type | |||
|---|---|---|---|---|---|
| without Starter | with Starter | without Starter | with Starter | ||
| pH | 0 | 6.11 ± 0.00 a | 6.11 ± 0.00 a | 6.11 ± 0.00 a | 6.11 ± 0.00 a |
| 2 | 6.14 ± 0.02 a | 6.09 ± 0.04 a | 5.69 ± 0.09 a | 5.20 ± 0.04 b | |
| 3 | - | - | 5.32 ± 0.05 a | 5.05 ± 0.04 b | |
| 7 | 6.04 ± 0.03 a | 5.24 ± 0.04 b | - | - | |
| 14 | - | - | 7.63 ± 0.33 a | 7.29 ± 0.02 a | |
| 16 | 5.84 ± 0.16 a | 5.11 ± 0.01 b | - | - | |
| aw | 0 | 0.979 ± 0.000 a | 0.979 ± 0.000 a | 0.979 ± 0.000 a | 0.979 ± 0.000 a |
| 2 | 0.974 ± 0.000 a | 0.977 ± 0.001 a | 0.972 ± 0.005 a | 0.974 ± 0.000 a | |
| 3 | - | - | 0.969 ± 0.006 a | 0.969 ± 0.008 a | |
| 7 | 0.945 ± 0.007 a | 0.944 ± 0.009 a | - | - | |
| 14 | - | - | 0.926 ± 0.009 a | 0.915 ± 0.013 a | |
| 16 | 0.876 ± 0.001 a | 0.864 ± 0.012 a | - | - | |
| Lactic acid (g/100 g) | 0 | 0.79 ± 0.02 a | 0.81 ± 0.10 a | 0.79 ± 0.02 a | 0.81 ± 0.01 a |
| 2 | 0.71 ± 0.01 a | 0.83 ± 0.04 a | 1.26 ± 0.18 a | 1.82 ± 0.04 b | |
| 3 | - | - | 2.13 ± 0.02 a | 2.19 ± 0.09 a | |
| 7 | 1.47 ± 0.09 a | 2.67 ± 0.21 b | - | - | |
| 14 | - | - | 0.54 ± 0.24 a | 1.38 ± 0.62 a | |
| 16 | 1.07 ± 0.34 a | 3.49 ± 0.31 b | - | - | |
| Weight loss (%) | 0 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 2 | 11.83 ± 2.02 a | 12.79 ± 2.73 a | 10.19 ± 6.91 a | 12.79 ± 5.66 a | |
| 3 | - | - | 18.09 ± 6.47 a | 20.87 ± 6.74 a | |
| 7 | 35.38 ± 4.20 a | 38.60 ± 1.91 a | - | - | |
| 14 | - | - | 49.41 ± 1.07 a | 48.81 ± 0.99 a | |
| 16 | 53.26 ± 0.73 a | 53.99 ± 0.27 a | - | - | |
| Time (Day) | Fuet-Type | Time | Snack-Type | ||||||
|---|---|---|---|---|---|---|---|---|---|
| without Starter | with Starter | (Day) | without Starter | with Starter | |||||
| Std | HPP | Std | HPP | Std | HPP | Std | HPP | ||
| 0 | 4.11 ± 0.03 | - | 5.67 ± 0.17 | - | 0 | 4.11 ± 0.03 | - | 5.67 ± 0.17 | - |
| 2 | 5.00 ± 0.05 | - | 8.41 ± 0.10 | - | 2 | 8.42 ± 0.43 | - | 8.68 ± 0.06 | - |
| 7 | 5.22 ± 0.38 | - | 8.17 ± 0.10 | - | 3 | 8.18 ± 0.22 a | 3.46 ± 0.47 b | 8.42 ± 0.08 a | 3.97 ± 1.16 b |
| 16 | 7.38 ± 0.46 a | 7.26 ± 0.19 a | 9.54 ± 0.08 a | 8.51 ± 0.07 b | 14 | 8.24 ± 0.44 a | 5.83 ± 0.40 b | 8.61 ± 0.05 a | 7.29 ± 0.43 b |
| 23 | 7.39 ± 0.24 a | 7.55 ± 0.29 a | 8.53 ± 0.11 a | 8.57 ± 0.07 a | 21 | 8.19 ± 0.38 a | 5.94 ± 0.21 b | 8.53 ± 0.09 a | 5.03 ± 0.95 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Austrich-Comas, A.; Jofré, A.; Gou, P.; Bover-Cid, S. Assessing the Impact of Different Technological Strategies on the Fate of Salmonella in Chicken Dry-Fermented Sausages by Means of Challenge Testing and Predictive Models. Microorganisms 2023, 11, 432. https://doi.org/10.3390/microorganisms11020432
Austrich-Comas A, Jofré A, Gou P, Bover-Cid S. Assessing the Impact of Different Technological Strategies on the Fate of Salmonella in Chicken Dry-Fermented Sausages by Means of Challenge Testing and Predictive Models. Microorganisms. 2023; 11(2):432. https://doi.org/10.3390/microorganisms11020432
Chicago/Turabian StyleAustrich-Comas, Anna, Anna Jofré, Pere Gou, and Sara Bover-Cid. 2023. "Assessing the Impact of Different Technological Strategies on the Fate of Salmonella in Chicken Dry-Fermented Sausages by Means of Challenge Testing and Predictive Models" Microorganisms 11, no. 2: 432. https://doi.org/10.3390/microorganisms11020432
APA StyleAustrich-Comas, A., Jofré, A., Gou, P., & Bover-Cid, S. (2023). Assessing the Impact of Different Technological Strategies on the Fate of Salmonella in Chicken Dry-Fermented Sausages by Means of Challenge Testing and Predictive Models. Microorganisms, 11(2), 432. https://doi.org/10.3390/microorganisms11020432