
Specialized Metabolites Produced by Phytotopatogen Fungi to Control Weeds and Parasite Plants


Abstract
:1. Introduction
2. Recent Developments for the Purification and Identification of Natural Compounds
3. Fungal Phytotoxins to Biocontrol Weeds
4. Fungal Phytotoxins to Biocontrol Parasitic Plants
5. Commercially Available Bioherbicides
6. Application Methods of Bioherbicides
6.1. Liquid Formulations
6.2. Solid Formulations
7. Future Perspectives
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gladbach, A.; Zhang, Y.; Fantke, P.; Settles, L.; Glas, D. Environmental impact reduction of crop protection-how to model, how to Mmasure, how to achieve? In Proceedings of the SETAC Europe 32nd Annual Meeting: Towards a Reduced Pollution, Copenhagen, Denmark, 15–19 May 2022. [Google Scholar]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Moyo, C.; Harrington, K.C.; Kemp, P.D.; Eerens, J.P.; Ghanizadeh, H. Wiper Application of herbicides to Cirsium arvense. Agronomy 2022, 12, 2262. [Google Scholar] [CrossRef]
- Goldwasser, Y.; Sazo, M.R.M.; Lanini, W.T. Control of field dodder (Cuscuta campestris) parasitizing tomato with ALS-inhibiting herbicides. Weed Technol. 2012, 26, 740–746. [Google Scholar] [CrossRef]
- Cannizzo, F. Briciole di Bellezza: Dialoghi di Speranza per il Futuro del Bel Paese; Mimesis Edizioni: Milano, Italy, 2018. [Google Scholar]
- Wheeler, G.S.; Jones, E.; Dyer, K.; Silverson, N.; Wright, S.A. Quarantine host range and natural history of Gadirtha fusca, a potential biological control agent of Chinese tallowtree (Triadica sebifera) in North America. Entomol. Exp. Appl. 2018, 166, 894–906. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Sung, C.Y.; Li, M.H.; Chu, K.H. Bioretention for stormwater quality improvement in Texas: Removal effectiveness of Escherichia coli. Sep. Purif. Technol. 2012, 84, 120–124. [Google Scholar] [CrossRef]
- Schoenenberger, N.; D’Andrea, L. Surveying the occurrence of subspontaneous glyphosate-tolerant genetically engineered Brassica napus L. (Brassicaceae) along Swiss railways. Environ. Sci. Eur. 2012, 24, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Evidente, A.; Capretti, P.; Giordano, F.; Surico, G. Identification and phytotoxicity of 3-nitropropanoic acid produced in vitro by Melanconis thelebola. Experientia 1992, 48, 1169–1172. [Google Scholar] [CrossRef]
- Gazoulis, I.; Kanatas, P.; Papastylianou, P.; Tataridas, A.; Alexopoulou, E.; Travlos, I. Weed management practices to improve establishment of selected lignocellulosic crops. Energie 2021, 14, 2478. [Google Scholar] [CrossRef]
- Evidente, A.; Abouzeid, M.A. Characterization of phytotoxins from phytopathogenic fungi and their potential use as herbicides in integrated crop management. In Handbook of Sustainable Weed Management; Sing, H.P., Batish, D.R., Kohli, R.K., Eds.; The Haworth Press: New York, NY, USA, 2006; pp. 507–532. [Google Scholar]
- Evidente, A.; Andolfi, A.; Cimmino, A. Fungal phytotoxins for control of Cirsium arvense and Sonchus arvensis. Pest Technol. 2011, 5, 1–17. [Google Scholar]
- Joel, D.M.; Hershenhorn, J.; Eizenberg, H.; Aly, R.; Ejeta, G.; Rich, P.J.; Ransom, J.K.; Sauerborn, J.; Rubiales, D. Biology and management of weedy root parasites. Hortic. Rev. 2007, 33, 267–349. [Google Scholar]
- Evidente, A.; Abouzeid, M.A.; Andolfi, A.; Cimmino, A.J. Recent achievements in the bio-control of Orobanche infesting important crops in the Mediterranean basin. Agric. Sci. Technol. 2011, 1, 461–483. [Google Scholar]
- Rimando, A.M.; Duke, O.S. Natural products for pest management. In Natural Products for Pest Management; ACS Symposium Series 927; Rimando, A.M., Duke, O.S., Eds.; American Chemica Society: Washington, DC, USA, 2006; pp. 2–23. [Google Scholar]
- Loddo, D.; McElroy, J.S.; Giannini, V. Problems and perspectives in weed management. Ital. J. Agron. 2021, 16, 1854. [Google Scholar] [CrossRef]
- Koivunen, M.E.; Coat, J.R.; Duke, O.S.; Beck, J.J. Pest Management with natural products. In Pest Management with Natural Products; ACS Symposium Series 1141; Beck, J.J., Coat, J.R., Duke, O.S., Koivunen, M.E., Eds.; American Chemica Society: Washington, DC, USA, 2013; pp. 1–4. [Google Scholar]
- Ballio, A.; Graniti, A. Phytotoxins and their involvement in plant disease. Experientia 1991, 47, 751–826. [Google Scholar]
- García-Pajón, C.M.; Collado, I.G. Secondary metabolites isolated from Colletotrichum species. Nat. Pod. Rep. 2003, 20, 426–431. [Google Scholar] [CrossRef]
- Strange, R.N. Phytotoxins produced by microbial plant pathogens. Nat. Prod. Rep. 2007, 24, 127–144. [Google Scholar] [CrossRef]
- Berestetskiy, O. Metabolite profiles and biological activity of extracts from Alternaria sonchi S-102 Culture grown by different fermentation methods. Appl. Biochem. Microbiol. 2008, 44, 443–465. [Google Scholar] [CrossRef]
- Möbius, N.; Hertweck, C. Fungal phytotoxins as mediators of virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Varejão, E.V.; Demuner, A.J.; Barbosa, L.C.; Barreto, R.W. The search for new natural herbicides–Strategic approaches for discovering fungal phytotoxins. Crop Prot. 2013, 48, 41–50. [Google Scholar] [CrossRef]
- Schueffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef]
- Jones, R.W.; Hancock, J.G. Soilborne fungi for biological control of weeds. In Microbes and Microbial Products as Herbicides; ACS Symposium Series 439; Honcock, J.G., Ed.; American Chemica Society: Washington, DC, USA, 1990; pp. 276–286. [Google Scholar]
- Abbas, H.K.; Duke, S.O. Phytotoxins from plant pathogens as potential herbicides. J. Toxicol. Toxin Rev. 1995, 14, 523–543. [Google Scholar] [CrossRef]
- El-Sayed, W. Biological control of weeds with pathogens: Current status and future trends/Biologische Schadpflanzenbekämpfung mit Pathogenen: Aktueller Status und Trends von morgen. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 2005, 112, 209–221. [Google Scholar]
- Hoagland, R.E. Microbial allelochemicals and pathogens as bioherbicidal agents. Weed Technol. 2001, 15, 835–857. [Google Scholar] [CrossRef]
- Chakraborty, A.; Ray, P. Mycoherbicides for the Noxious Meddlesome: Can Colletotrichum be a Budding Candidate? Front. Microbiol. 2021, 12, 754048. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, R.E.; Boyette, C.D.; Weaver, M.A.; Abbas, H.K. Bioherbicides: Research and risks. Toxin Rev. 2007, 26, 313–342. [Google Scholar] [CrossRef]
- .Strobel, G.; Kenfield, D.; Bunkers, G.; Sugawara, F.; Clardy, J. Phytotoxins as potential herbicides. Experientia 1991, 47, 819–826. [Google Scholar] [CrossRef]
- Singh, A.K.; Pandey, A.K. Fungal metabolites as a natural source of herbicide: A novel approach of weed management. J. Appl. Nat. Sci. 2019, 11, 158–163. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Evidente, A. Fungal phytotoxins with potential herbicidal activity to control Chenopodium album. Nat. Prod. Commun. 2015, 10, 1121. [Google Scholar] [CrossRef] [Green Version]
- Macías-Rubalcava, M.L.; Garrido-Santos, M.Y. Phytotoxic compounds from endophytic fungi. Appl. Microbiol. Biotechnol. 2022, 106, 931–950. [Google Scholar] [CrossRef]
- Vurro, M.; Boari, A.; Evdente, A.; Andolfi, A.; Zermane, N. Natural metabolites for parasitic weed management. Pest Manag. Sci. 2009, 65, 566–571. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Abouzeid, M.; Evidente, A. Polyphenols as fungal phytotoxins, seed germination stimulants and phytoalexins. Phytochem. Rev. 2013, 12, 653–672. [Google Scholar] [CrossRef]
- Hostettmann, K.; Hostettmann, M.; Marston, A. Preparative Chromatography Techniques. In Applications in Natural Product Isolation, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Tringali, C. Bioactive Compounds from Natural Sources: Isolation, Characterization and Biological Properties; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Spano, M.; Andreani, S.; Naubron, J.V.; Mannina, L.; Pricl, S.; Muselli, A.; Tintaru, A. Smart IM-MS and NMR study of natural diastereomers: The study case of the essential oil from Senecio transiens. Anal. Bioanal. Chem. 2022, 414, 6695–6705. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, C.L.; Trimpin, S. Ion Mobility Spectrometry-Mass Spectrometry: Theory and Applications; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds-Distribution and Biology; The University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Iwasaki, S.; Muro, H.; Nozoe, S.; Okuda, S.; Sato, Z. Isolation of 3, 4-dihydro-3, 4, 8-trihydroxy-1 (2H)-naphthalenone and tenuazonic acid from Pyricularia oryzae cavara. Tetrahedron Lett. 1972, 13, 13–16. [Google Scholar] [CrossRef]
- Tarr, S.A.J. Diseases of Sorghum, Sudan Grass and Broom Corn; Commonwealth Mycological Institute: Kew, UK, 1962. [Google Scholar]
- Mikami, Y.; Nishijima, Y.; Iimura, H.; Suzuki, A.; Tamura, S. Chemical studies on brown-spot disease of tobacco plants part I. Tenuazonic acid as a vivotoxin of Alternaria longipes. Agric. Biol. Chem. 1971, 35, 611–618. [Google Scholar]
- Robeson, D.J.; Strobel, G.A. Monocerin, a phytotoxin from Exserohilum turcicum (≡ Drechslera turcica). Agric. Biol. Chem. 1982, 46, 2681–2683. [Google Scholar]
- Robeson, D.; Strobel, G.; Matusumoto, G.K.; Fisher, E.L.; Chen, M.H.; Clardy, J. Alteichin: An unusual phytotoxin from Alternaria eichorniae, a fungal pathogen of water hyacinth. Experientia 1984, 40, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Charudattan, R.; Rao, K.V. Bostrycin and 4-deoxybostrycin: Two nonspecific phytotoxins produced by Alternaria eichhorniae. Appl. Environ. Microbiol. 1982, 43, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, F.; Strobel, G.; Fisher, L.E.; Van Duyne, G.D.; Clardy, J. Bipolaroxin, a selective phytotoxin produced by Bipolaris cynodontis. Proc. Natl. Acad. Sci. USA 1985, 82, 8291–8294. [Google Scholar] [CrossRef] [Green Version]
- Stierle, A.C.; Cardellina, J.H.; Strobel, G.A. Maculosin, a host-specific phytotoxin for spotted knapweed from Alternaria alternata. Proc. Natl. Acad. Sci. USA 1988, 85, 8008–8011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.; Cranston, R. An economic evaluation of control methods for diffuse and spotted knapweed in western Canada. Can. J. Plant Sci. 1979, 59, 375–382. [Google Scholar] [CrossRef]
- Puopolo, G.; Cimmino, A.; Palmieri, M.C.; Giovannini, O.; Evidente, A.; Pertot, I. Lysobacter capsici AZ78 produces cyclo (l-Pro-l-Tyr), a 2, 5-diketopiperazine with toxic activity against sporangia of Phytophthora infestans and Plasmopara viticola. J. Appl. Microbiol. 2014, 117, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Fry, W. Phytophthora infestans: The plant (and R gene) destroyer. Mol. Plant. Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Cimmino, A.; Puopolo, G.; Perazzolli, M.; Andolfi, A.; Melck, D.; Pertot, I.; Evidente, A. Cyclo (L-pro-L-tyr), the fungicide isolated from Lysobacter capsici AZ78: A structure–activity relationship study. Chem. Heterocycl. Compounds 2014, 50, 290–295. [Google Scholar] [CrossRef]
- Hassall, K.A. Biochemistry and Uses of Pesticides; Verlag Chemie: Weinheim, Germany, 1990; p. 58. [Google Scholar]
- Bräse, S.; Gil, C.; Knepper, K.; Zimmermann, V. Organic azides: An exploding diversity of a unique class of compounds. Angew. Chem. Int. Ed. 2005, 4, 5188–5240. [Google Scholar] [CrossRef]
- Sharma, K.; Butz, A.F.; Finckh, M.R. Effects of host and pathogen genotypes on inducibility of resistance in tomato (Solanum lycopersicum) to Phytophthora infestans. Plant Pathol. 2010, 59, 1062–1071. [Google Scholar] [CrossRef]
- Cimmino, A.; Bejarano, A.; Masi, M.; Puopolo, G.; Evidente, A. Isolation of 2, 5-diketopiperazines from Lysobacter capsici AZ78 with activity against Rhodococcus fascians. Nat. Prod. Res. 2021, 35, 4969–4977. [Google Scholar] [CrossRef]
- Kusumi, T.; Ohtani, I.; Nishiyama, K.; Kakisawa, H. Caryoynencins, potent antibiotics from a plant pathogen Pseudomonas caryophylli. Tetrahedron Lett. 1987, 28, 3981–3984. [Google Scholar] [CrossRef]
- Young, J.M.; Fletcher, M.J. Pseudomonas syringae pv. panici (Elliott 1923) Young, Dye & Wilkie 1978 is a doubtful name. Australas. Plant Pathol. 1994, 23, 66–68. [Google Scholar]
- Mapuranga, N. A new race of Pseudomonas syringae pv. tabaci on tobacco in Zimbabwe. Plant Dis. 1998, 82, 1404. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Barranquero, J.A.; Cazorla, F.M.; de Vicente, A. Pseudomonas syringae pv. syringae associated with mango trees, a particular pathogen within the “hodgepodge” of the Pseudomonas syringae complex. Front. Plant Sci. 2019, 10, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.M. Pseudomonas syringae pv. japonica (Mukoo 1955) Dye et al. 1980 is a junior synonym of Ps. syringae pv. syringae van Hall 1902. Lett. Appl. Microbiol. 1992, 15, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.D.; Dias-Neto, J.J.; Santos, M.D.; Formento, A.N.; Bizerra, L.V.; Fonseca, M.E.N.; Boiteux, L.S.; Café-Filho, A.C. Current status of soybean anthracnose associated with Colletotrichum truncatum in Brazil and Argentina. Plants 2019, 8, 459. [Google Scholar] [CrossRef] [Green Version]
- Castaldi, S.; Cimmino, A.; Masi, M.; Evidente, A. Bacterial lipodepsipeptides and some of their derivatives and cyclic dipeptides as potential agents for biocontrol of pathogenic bacteria and fungi of agrarian plants. J. Agric. Food Chem. 2022, 70, 4591–4598. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.C.; Caddlina, J.H.; Strobel, G.A. Phytotoxins from Alternaria alternata, a pathogen of spotted knapweed. J. Nat. Prod. 1989, 52, 42–47. [Google Scholar] [CrossRef]
- Okuno, T.; Natsume, I.; Sawai, K.; Sawamura, K.; Furusaki, A.; Matsumoto, T. Structure of antifungal and phytotoxic pigments produced by Alternaria sp. Tetrahedron Lett. 1983, 24, 5653–5656. [Google Scholar] [CrossRef]
- Stack, M.E.; Mazzola, E.P.; Page, S.W.; Pohland, A.E.; Highet, R.J.; Tempesta, M.S.; Corley, D.G. Mutagenic perylenequinone metabolites of Alternaria alternata: Altertoxins I, II, and III. J. Nat. Prod. 1986, 49, 866–871. [Google Scholar] [CrossRef]
- Jones, R.W.; Lanini, W.T.; Hancock, J.G. Plant growth response to the phytotoxin viridiol produced by the fungus Gliocladium virens. Weed Sci. 1988, 36, 683–687. [Google Scholar] [CrossRef]
- Bell, M.R.; Johnson, J.R.; Wildi, B.S.; Woodward, R.B. The structure of gliotoxin. J. Am. Chem. Soc. 1958, 80, 1001. [Google Scholar] [CrossRef]
- Stipanovic, R.D.; Howell, C.R. The structure of gliovirin, a new antibiotic from Gliocladium virens. J. Antibiot. 1982, 35, 1326–1330. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Tuzi, A.; Linaldeddu, B.T.; Montecchio, L.; Maddau, L.; Evidente, A. Hyfraxinic acid, a phytotoxic tetrasubstituted octanoic acid, produced by the ash (Fraxinus excelsior L.) pathogen Hymenoscyphus fraxineus together with viridiol and some of its analogues. J. Agric. Food Chem. 2019, 67, 13617–13623. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.K.; Kenfield, D.; Strobel, G.A.; Hess, W.M. Ascochyta cypericola sp. nov. causing leaf blight of purple nutsedge (Cyperus rotundus). Can. J. Bot. 1991, 69, 797–802. [Google Scholar] [CrossRef]
- Stierle, A.; Upadhyay, R.; Strobel, G. Cyperine, a phytotoxin produced by Ascochyta cypericola, a fungal pathogen of Cyperus rotundus. Phytochemistry 1991, 30, 2191–2192. [Google Scholar] [CrossRef]
- Venkatasubbaiah, P.; Dyke, C.V.; Chilton, W.S. Phytotoxic metabolites of Phoma sorghina, a new foliar pathogen of pokeweed. Mycologia 1992, 84, 715–723. [Google Scholar] [CrossRef]
- Madrewesi, J.N.C. White leaf spot disease of fluted pumpkin (Telairia occidentalis) in Nigeria. Niger. J. Plant Prot. 1977, 3, 122–128. [Google Scholar]
- Saksena, A.K.; Jain, S.K.; Saksena, S.B. On the occurrence of three new fungal diseases of cultivated plants in India. Proc. Natl. Acad. Sci. India Sect. B Bioi. Sci. 1982, 52, 79–80. [Google Scholar]
- Shivapuri, A.; Mishra, A. Occurence of Phoma sorghina on phalsa. Indian J. Mycol. Plant Pathol. 1982, 12, 13. [Google Scholar]
- Abdel-Rahim, A.M. Phoma sorghina causing a leaf spot of water hyacinth in the Sudan. Plant Pathol. 1984, 33, 429. [Google Scholar] [CrossRef]
- Kumar, P.; Seth, J.N. A new leaf spot disease of winged bean caused by Phoma sorghina. Int. J. Trop. Plant Dis. 1985, 3, 87–88. [Google Scholar]
- Prabhu, A.S.; Bedendo, I.P. Glume blight of rice in Brazil: Etiology, varietal reactions and loss estimates. Trop. Pest Manag. 1988, 34, 85–88. [Google Scholar] [CrossRef]
- Venkatasubbaiah, P.; Chilton, W.S. Phytotoxins of Ascochyta hyalospora, causal agent of lambsquarters leaf spot. J. Nat. Prod. 1992, 55, 461–467. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Vurro, M.; Bottalico, A. Ascosalitoxin, a phytotoxic trisubstituted salicylic aldehyde from Ascochyta pisi. Phytochemistry 1993, 34, 995–998. [Google Scholar] [CrossRef]
- Kempenar, C. Studies on Biological Control of Chenopodium album by Ascochyta caulin. Ph.D. Thesis, University of Wageningen, Wageningen, The Netherlands, 1995. [Google Scholar]
- Netland, J.; Dutton, L.C.; Greaves, M.P.; Baldwin, M.; Vurro, M.; Evidente, A.; Einhorn, G.; Scheepens, P.C.; French, L.W. Biological control of Chenopodium album L. in Europe. BioControl 2001, 46, 175–196. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Cutignano, A.; Taglialatela-Scafati, O.; Vurro, M.; Zonno, M.C.; Motta, A. Ascaulitoxin, a phytotoxic bis-amino acid N-glucoside from Ascochyta caulina. Phytochemistry 1998, 48, 1131–1137. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Vurro, M.; Zonno, M.C.; Motta, A. Trans-4-aminoproline, a phytotoxic metabolite with herbicidal activity produced by Ascochyta caulina. Phytochemistry 2000, 53, 231–237. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Vurro, M.; Zonno, M.C. Determination of Ascochyta caulina phytotoxins by high-performance anion exchange chromatography and pulsed amperometric detection. Phytochem. Anal. 2001, 12, 383–387. [Google Scholar] [CrossRef]
- Bassarello, C.; Bifulco, G.; Evidente, A.; Riccio, R.; Gomez-Paloma, L. Stereochemical studies on ascaulitoxin: A J-based NMR configurational analysis of a nitrogen substituted system. Tetrahedron Lett. 2001, 42, 8611–8613. [Google Scholar] [CrossRef]
- Avolio, F.; Andolfi, A.; Zonno, M.C.; Boari, A.; Cimmino, A.; Vurro, M.; Evidente, A. Large-scale production and purification of Ascochyta caulina phytotoxins and a new HPLC method for their analysis. Chromatographia 2011, 74, 633–638. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.O.; Evidente, A.; Fiore, M.; Rimando, A.M.; Dayan, F.E.; Vurro, M.; Christiansen, N.; Looser, R.; Hutzler, J.; Grossmann, K. Effects of the aglycone of ascaulitoxin on amino acid metabolism in Lemna paucicostata. Pestic. Biochem. Physiol. 2011, 100, 41–50. [Google Scholar] [CrossRef]
- Fumagalli, P.; Andolfi, A.; Avolio, F.; Boari, A.; Cimmino, A.; Finizio, A. Ecotoxicological characterization of a mycoherbicide mixture from the fungus Ascochyta caulina. Pest Manag. Sci. 2013, 69, 850–856. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Avolio, F.; Santini, A.; Tuzi, A.; Berestetskyi, A.; Vurro, M.; Evidente, A. Chenopodolin: A phytotoxic unrearranged ent-pimaradiene diterpene produced by Phoma chenopodiicola, a fungal pathogen for Chenopodium album biocontrol. J. Nat. Prod. 2013, 76, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Avolio, F.; Berestetskiy, A.; Vurro, M.; Evidente, A. Chenopodolans A–C: Phytotoxic furopyrans produced by Phoma chenopodiicola, a fungal pathogen of Chenopodium album. Phytochemistry 2013, 96, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Evidente, M.; Cimmino, A.; Zonno, M.C.; Masi, M.; Berestetskyi, A.; Santoro, E.; Superchi, S.; Vurro, M.; Evidente, A. Phytotoxins produced by Phoma chenopodiicola, a fungal pathogen of Chenopodium album. Phytochemistry 2015, 117, 482–488. [Google Scholar] [CrossRef]
- Evidente, M.; Cimmino, A.; Zonno, M.C.; Masi, M.; Santoro, E.; Vergura, S.; Berestetskyi, A.; Superchi, S.; Vurro, M.; Evidente, A. Chenopodolans E and F, two new furopyrans produced by Phoma chenopodiicola and absolute configuration determination of chenopodolan B. Tetrahedron 2016, 72, 8502–8507. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Ogilvie, K.K.; Watson, A.K. Phytotoxic metabolites of Phomopsis convolvulus, a host-specific pathogen of field bindweed. Can. J. Chem. 1992, 70, 2276–2284. [Google Scholar] [CrossRef] [Green Version]
- Kenfield, D.; Bunkers, G.; Wu, Y.H.; Strobel, G.; Sugawara, F.; Hallock, Y.; Clardy, J. Gigantenone, a novel sesquiterpene phytohormone mimic. Experientia 1989, 45, 900–902. [Google Scholar] [CrossRef]
- Masi, M.; Sautua, F.; Zatout, R.; Castaldi, S.; Arrico, L.; Isticato, R.; Pescitelli, G.; Carmona, M.A.; Evidente, A. Phaseocyclopentenones A and B, phytotoxic penta-and tetrasubstituted cyclopentenones produced by Macrophomina phaseolina, the causal agent of charcoal rot of soybean in Argentina. J. Nat. Prod. 2021, 84, 459–465. [Google Scholar] [CrossRef]
- Sugama, K.; Hayashi, K.; Nakagawa, T.; Mitsuhashi, H.; Yoshida, N. Sesquiterpenoids from Petasites fragrans. Phytochemistry 1983, 22, 1619–1622. [Google Scholar] [CrossRef]
- Bottalico, A.; Frisullo, S.; Iacobellis, N.S.; Capasso, R.; Corrado, E.; Randazzo, G. Occurrence of phomenone in Tomato plants infected by Phoma destructiva Plowr. Phytopathol. Mediterr. 1983, 22, 116–119. [Google Scholar]
- Sugawara, F.; Strobel, G.; Yoshida, S. Phytoactive eremophilanes produced by the weed pathogen Drechslera gigantea. Biosci. Biotechnol. Biochem. 1993, 57, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Vurro, M.; Fracchiolla, M.; Charudattan, R. Herbicidal potential of ophiobolins produced by Drechslera gigantea. J. Agric. Food Chem. 2006, 54, 1779–1783. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T. Cotylenines, leaf growth substances produced by fungus. Part I. Isolation and characterization of cotylenins. Agric. Biol. Chem. 1971, 35, 1415–1418. [Google Scholar]
- Canonica, L.; Fiecchi, A.; Galli Kienle, M.; Scala, A. The costitution of cochliobolin. Tetrahedron Lett. 1966, 7, 1211–1218. [Google Scholar] [CrossRef]
- Nozoe, S.; Hirai, K.; Tsuda, K. The structure of zizanin-A and -B, C25-terpenoids isolated from Helminthosporium zizaniae. Tetrahedron Lett. 1966, 7, 2211–2216. [Google Scholar] [CrossRef]
- Canonica, L.; Fiecchi, A.; Galli Kienle, M.; Scala, A. Isolation and constitution of cochliobolin. Tetrahedron Lett. 1966, 7, 1329–1333. [Google Scholar] [CrossRef]
- Itai, A.; Nozoe, S.; Tsuda, K.; Okuda, S.; Iitaka, Y.; Makayama, Y. The structure of cephalonic acid, a pentaprenyl terpenoid. Tetrahedron Lett. 1967, 42, 4111–4112. [Google Scholar] [CrossRef]
- Nozoe, S.; Itai, A.; Tsuda, K.; Okuda, S. Chemical transformation of cephalonic acid. Tetrahedron Lett. 1967, 8, 4113–4117. [Google Scholar] [CrossRef] [PubMed]
- Nozoe, S.; Morisaki, M.; Fukushima, K.; Okuda, S. The isolation of an acyclic C25-isoprenoid alcohol, geranylnerolidol, and a new ophiobolin. Tetrahedron Lett. 1968, 9, 4457–4458. [Google Scholar] [CrossRef]
- Au, T.K.; Chick, W.S.H.; Leung, P.C. The biology of ophiobolins. Life Sci. 2000, 67, 733–742. [Google Scholar] [CrossRef]
- Tian, W.; Deng, Z.; Hong, K. The biological activities of sesterterpenoid-type ophiobolins. Mar. Drugs 2017, 15, 229. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Dasari, R.; Evidente, A.; Mathieu, V.; Kornienko, A. Chemistry and biology of ophiobolin A and its congeners. Bioorg. Med. Chem. Lett. 2019, 29, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Reisenauer, K.N.; Masi, M.; Evidente, A.; Taube, J.H.; Romo, D. Pharmacophore-directed retrosynthesis applied to ophiobolin A: Simplified bicyclic derivatives displaying anticancer activity. Org. Lett. 2020, 22, 8307–8312. [Google Scholar] [CrossRef] [PubMed]
- Reisenauer, K.N.; Tao, Y.; Das, P.; Song, S.; Svatek, H.; Patel, S.D.; Mikhail, S.; Ingros, A.; Sheesley, P.; Masi, M.; et al. Epithelial-mesenchymal transition sensitizes breast cancer cells to cell death via the fungus-derived sesterterpenoid ophiobolin A. Sci. Rep. 2021, 11, 10652. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Vurro, M.; Fracchiolla, M.; Charudattan, R.; Motta, A. Ophiobolin E and 8-epi-ophiobolin J produced by Drechslera gigantea, a potential mycoherbicide of weedy grasses. Phytochemistry 2006, 67, 2281–2287. [Google Scholar] [CrossRef]
- Zatout, R.; Masi, M.; Sangermano, F.; Vurro, M.; Zonno, M.C.; Santoro, E.; Calabrò, V.; Superchi, S.; Evidente, A. Drophiobiolins A and B, bioactive ophiobolan sestertepenoids produced by Dreschslera gigantea. J. Nat. Prod. 2020, 83, 3387–3396. [Google Scholar] [CrossRef]
- Abbas, H.K.; Vesonder, R.F.; Boyette, C.; Peterson, S.W. Phytotoxicity of AAL-toxin and other compounds produced by Alternaria alternata to jimsonweed (Datura stramonium). Can. J. Bot. 1993, 71, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Boyette, C.D. Evaluation of Alternaria crassa for biological control of jimsonweed: Host range and virulence. Plant Sci. 1986, 45, 223–228. [Google Scholar] [CrossRef]
- Brice, V.R.; Stack, M.E.; Mislivec, P.B. Incidence of toxic Alternaria species in small grains from the U.S.A. J. Food Sci. 1984, 49, 1626–1627. [Google Scholar] [CrossRef]
- Harvan, D.J.; Pero, R.W. The structure and toxicity of Alternaria metabolites. In Mycotoxins and Other Fungal Related Food Problems; Advances in Chemistry; Rodricks, J.V., Ed.; American Chemical Society: Washington, DC, USA, 1976; Volume 149, pp. 344–355. [Google Scholar]
- Hori, M.; Kakiki, K.; Misato, T. Mechanism of polyoxin-resistance in Alternaria kikuchiarza. J. Pestic. Sci. 1976, 1, 31–40. [Google Scholar] [CrossRef] [Green Version]
- King, A.D., Jr.; Schade, J.E. Alternaria toxins and their importance in food. J. Food Prot. 1984, 47, 886–901. [Google Scholar] [CrossRef]
- Kohmoto, K.; Scheffer, R.P.; Whiteside, J.O. Host-selective toxins from Alternaria citri. Phytopathology 1979, 69, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Kohmoto, K.; Verma, V.S.; Nishirnara, S.; Tagami, M.; Scheffer, R.P. New outbreak of Alternaria stem canker of tomato in Japan and production of host-selective toxins by the causal fungus. J. Fac. Agric. Tottori Univ. 1982, 17, 1–8. [Google Scholar]
- Logrieco, A.; Bottalico, A.; Solfrizzo, M.; Mule, G. Incidence of Alternaria species in grains from Mediterranean countries and their ability to produce mycotoxins. Mycologia 1990, 82, 501–505. [Google Scholar] [CrossRef]
- Templeton, G.E. Alternanria toxins related to pathogenesis in plants. In Microbial Toxins; Kadis, S., Ciegler, A., Eds.; Academic Press: New York, NY, USA, 1972; pp. 169–172. [Google Scholar]
- Templeton, G.E.; Grable, C.I.; Fulton, N.D.; Bollenbacher, K. Factors affecting the amount and pattern of chlorosis caused by metabolite of Alternaria renuis. Phytopathology 1967, 57, 516–518. [Google Scholar]
- Tiejin, W.; Ceponis, M. Nature and extent of losses in fresh repacked tomatoes. Phytopathology 1982, 72, 266–267. [Google Scholar]
- Walker, H.L.; Riley, J.A. Evaluation of Alternaria cassiae for the biocontrol of sicklepod (Cassia obrnsifolia). Weed Sci. 1982, 30, 651–654. [Google Scholar] [CrossRef]
- Walker, H.L.; Sciumbato, G.L. Evaluation of Alterrlaria macrospora as potential biocontrol agent for spurred anoda (Atzoda cristata): Host range studies. Weed Sci. 1979, 27, 612–614. [Google Scholar] [CrossRef]
- Tanaka, M.; Ohra, J.; Tsujino, Y.; Sawaji, Y.; Fujimori, T. Phytotoxin produced by Nimbya scirpicola. Biosci. Biotechnol. Biochem. 1994, 58, 565–566. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Matsutani, S.; Sugita, K.; Yoshida, H.; Hayashi, F.; Tcrui, Y.; Nakai, H.; Uotani, N.; Kawamura, Y.; Matsumoto, K.; et al. Depudecin: A novel compound inducing the flat phenotype of NIH3T3 cells doubly transformed by ras-and src-oncogene, produced by Alternaria brassicicola. J. Antibiot. 1992, 45, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Ohra, J.; Morita, K.; Tsujino, Y.; Tazaki, H.; Fujimori, T.; Goering, M.; Evans, S.; Zorner, P. Production of the phytotoxic metabolite, ferricrocin, by the fungus Colletotrichum gloeosporioides. Biosci. Biotechnol. Biochem. 1995, 59, 113–114. [Google Scholar] [CrossRef] [Green Version]
- Farr, D.F.; Bills, G.F.; Chamuris, G.P.; Rossman, A.Y. Fungi on Plants and Plant Products in the United States; APS Press: St. Paul, MN, USA, 1989; p. 638. [Google Scholar]
- Keller-Schierlein, W.; Deer, A. Stoffwechselprodukte von Mikroorganismen. 45. Mitteilung. Über die Konstitution von Ferrirubin, Ferrirhodin und Ferrichrom A. Helv. Chim. Acta 1963, 46, 1907–1929. [Google Scholar] [CrossRef]
- Neilands, J.B. Hydroxamic Acids in Nature: Sophisticated ligands play a role in iron metabolism and possibly in other processes in microorganisms. Science 1967, 156, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Barnes, L.; Eng-Wilmot, D.L.; Helm, D.V.D. Ferricrocin (C29H44FeN9O13. 7H2O), an iron (III)-binding peptide from Aspergillus versicolor. Acta Cryst. 1984, 40, 922–926. [Google Scholar]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Andolfi, A.; Bottalico, A.; Vurro, M.; Zonno, M.C. Putaminoxin, a phytotoxic nonenolide from Phoma putaminum. Phytochemistry 1995, 40, 1637–1641. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Andolfi, A.; Vurro, M.; Zonno, M.C. Putaminoxins B and C from Phoma putaminum. Phytochemistry 1997, 44, 1041–1045. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Andolfi, A.; Vurro, M.; Zonno, M.C. Putaminoxins D and E from Phoma putaminum. Phytochemistry 1998, 48, 941–945. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Vurro, M.; Botralico, A. Pinolidoxin, a phytotoxic nonenolide from Ascochyta pinodes. Phytochemistry 1993, 34, 999–1003. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Abouzeid, M.A.; Lanzetta, R.; Vurro, M.; Bottalico, A. Three new toxic pinolidoxins from Ascochyta pinodes. J. Nat. Prod. 1993, 56, 1937–1943. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Andolfi, A.; Vurro, M.; Zonno, M.C. Structure–activity relationship studies of putaminoxins and pinolidoxins: Phytotoxic nonenolides produced by phytopathogenic Phoma and Ascochyta species. Nat. Toxins 1998, 6, 183–188. [Google Scholar] [CrossRef]
- Rivero-Cruz, J.F.; García-Aguirre, G.; Cerda-García-Rojas, C.M.; Mata, R. Conformational behavior and absolute stereostructure of two phytotoxic nonenolides from the fungus Phoma herbarum. Tetrahedron 2000, 56, 5337–5344. [Google Scholar] [CrossRef]
- Rivero-Cruz, J.F.; Macías, M.; Cerda-García-Rojas, C.M.; Mata, R. A New Phytotoxic Nonenolide from Phoma herbarum. J. Nat. Prod. 2003, 66, 511–514. [Google Scholar] [CrossRef]
- Seo, C.O.; Oh, H.C.; Lee, H.B.; Kim, J.K.; Kong, I.S.; Ahn, S.C. Hexaketides from phytopathogenic fungus Paraphaeosphaeria recurvifoliae. Bull. Korean Chem. Soc. 2007, 28, 1803–1806. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Fondevilla, S.; Abouzeid, M.A.; Rubiales, D.; Evidente, A. Pinolide, a new nonenolide produced by Didymella pinodes, the causal agent of Ascochyta blight on Pisum sativum. J. Agric. Food Chem. 2012, 60, 5273–5278. [Google Scholar] [CrossRef] [Green Version]
- Yuzikhin, O.; Mitina, G.; Berestetskiy, A. Herbicidal potential of stagonolide, a new phytotoxic nonenolide from Stagonospora cirsii. J. Agric. Food Chem. 2007, 55, 7707–7711. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Cimmino, A.; Berestetskiy, A.; Mitina, G.; Andolfi, A.; Motta, A. Stagonolides B−F, nonenolides produced by Stagonospora cirsii, a potential mycoherbicide of Cirsium arvense. J. Nat. Prod. 2008, 71, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Cimmino, A.; Berestetskiy, A.; Andolfi, A.; Motta, A. Stagonolides G–I and modiolide A, nonenolides produced by Stagonospora cirsii, a potential mycoherbicide for Cirsium arvense. J. Nat. Prod. 2008, 71, 1897–1901. [Google Scholar] [CrossRef] [PubMed]
- Dalinova, A.; Dubovik, V.; Chisty, L.; Kochura, D.; Ivanov, A.; Smirnov, S.; Petrova, M.; Zolotarev, A.; Berestetskiy, A. Stagonolides J and K and Stagochromene A, two new natural substituted nonenolides and a new disubstituted chromene-4,5-dione isolated from Stagonospora cirsii S-47 proposed for the biocontrol of Sonchus arvensis. J. Agric. Food Chem. 2019, 67, 13040–13050. [Google Scholar] [CrossRef]
- Mikhailova, N.F.; Tarasov, A.V. On the character of Cirsium arvense (Asteraceae) thickets. Bot. J. 1989, 74, 509–514. (In Russian) [Google Scholar]
- Donald, W.W. Management and control of Canada thistle (Cirsium arrvense). Rev. Weed Sci. 1990, 5, 193–250. [Google Scholar]
- Dalinova, A.; Fedorov, A.; Dubovik, V.; Voitsekhovskaja, O.; Tyutereva, E.; Smirnov, S.; Kochura, D.; Chisty, L.; Senderskiy, I.; Berestetskiy, A. Structure–activity relationship of phytotoxic natural 10-membered lactones and their semisynthetic derivatives. J. Fungi 2021, 7, 829. [Google Scholar] [CrossRef]
- Imaizumi, S.; Tokumasu, S.; Nitta, N.; Fujimori, T.; Yamada, M. A new disease of paddy weed Eleocharis kuroguwai caused by Dendryphiella sp. Ann. Phytopath. Soc. Jpn. 1992, 58, 158. [Google Scholar]
- Tanaka, M.; Ohra, J.; Tsujtno, Y.; Fujimori, T.; Ago, H.; Tsuge, H. Dendryol A, B, C, and D, four new compounds produced by a weed pathogenic fungus Dendryphiella sp. Z. Für Nat. C 1995, 50, 751–756. [Google Scholar] [CrossRef]
- Abbas, H.K.; Duke, S.O.; Merrill Jr, A.H.; Wang, E.; Shier, W.T. Phytotoxicity of australifungin, AAL-toxins and fumonisin B1 to Lemna pausicostata. Phytochemistry 1998, 47, 1509–1514. [Google Scholar] [CrossRef]
- Vurro, M.; Evidente, A.; Andolfi, A.; Zonno, M.C.; Giordano, F.; Motta, A. Brefeldin A and α, β-dehydrocurvularin, two phytotoxins from Alternaria zinniae, a biocontrol agent of Xanthium occidentale. Plant Sci. 1998, 138, 67–79. [Google Scholar] [CrossRef]
- Tietjen, K.G.; Hunkler, D.; Matern, U. Differential response of cultured parsley cells to elicitor from twonon-pathogenic strains of fungi. Eur. J. Biochem. 1983, 131, 401–407. [Google Scholar] [CrossRef]
- Schmitzer, P.R.; Graupner, P.R.; Chapin, E.L.; Fields, S.C.; Gilbert, J.R.; Gray, J.A.; Peacock, C.L.; Gerwick, B.C. Ribofuranosyl triazolone: A natural product herbicide with activity on adenylosuccinate synthetase following phosphorylation. J. Nat. Prod. 2000, 63, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Haines, D.R.; Leonard, N.J.; Wiemer, D.F. Syntheses and structure assignments of six azolinone ribonucleosides. J. Org. Chem. 1982, 47, 474–482. [Google Scholar] [CrossRef]
- Nakajima, M.; Itoi, K.; Takamatsu, Y.; Kinoshita, T.; Okazaki, T.; Kawakubo, K.; Masahiro Shindo, M.; Honma, T.; Tohjigamori, M.; Haneshi, T. Hydantocidin: A new compound with herbicidal activity from Streptomyces hygroscopicus. J. Antibiot. 1991, 44, 293–300. [Google Scholar] [CrossRef]
- Hoagland, R.E. The Genus Streptomyces: A Rich Source of Novel Phytotoxins; Ecology of Desert Environment; Scientific Publishers: Jodhpur, India, 2001; pp. 139–169. [Google Scholar]
- Harir, M.; Bendif, H.; Bellahcene, M.; Fortas, Z.; Pogni, R. Streptomyces secondary metabolites. Basic Biol. Appl. Actinobacteria 2018, 6, 99–122. [Google Scholar]
- Haneishi, T.; Nakajima, M.; Torikata, A.; Okazaki, T.; Tohjigamori, M.; Kawakubo, K. New Agrochemicail Named “Substance” no. 51262”, Its Preparation and Use. European Patent EP0232572A2, 19 August 1992. [Google Scholar]
- Pachlatko, J.P.; Zahner, H. New Streptomyces Hygroscopicus TU-2474—For production of the Herbicide Hydantocidin and the Bactericide Homomycin. German Patent DE 4129616 A1, Application 1991-09-06 Filed by Ciba Geigy AG, 3 December 1992. [Google Scholar]
- Chemla, P. Stereoselective synthesis of (+)-hydantocidin. Tetrahedron Lett. 1993, 34, 7391–7394. [Google Scholar] [CrossRef]
- Heim, D.R.; Cseke, C.; Gerwick, B.C.; Murdoch, M.G.; Green, S.B. Hydantocidin: A possible proherbicide inhibiting purine biosynthesis at the site of adenylosuccinate synthetase. Pest Biochem. Phys. 1995, 53, 138–145. [Google Scholar] [CrossRef]
- Macías, M.; Ulloa, M.; Gamboa, A.; Mata, R. Phytotoxic compounds from the new coprophilous fungus Guanomyces polythrix. J. Nat. Prod. 2000, 63, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubbaiah, P.; Baudoin, A.B.A.M.; Chilton, W.S. Leaf spot of hemp dogbane caused by Stagonospora apocyni, and its phytotoxins. J. Phytopathol. 1992, 135, 309–316. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Vurro, M.; Zonno, M.C.; Motta, A. Cytochalasins Z1, Z2 and Z3, three 24-oxa [14] cytochalasans produced by Pyrenophora semeniperda. Phytochemistry 2002, 60, 45–53. [Google Scholar] [CrossRef]
- Medd, R.H. A review of the world distribution and host range of Pyrenophora semeniperda. Rev. Plant Pathol. 1992, 71, 891–901. [Google Scholar]
- Neergard, P. Seed Pathology; The Macmillan Press: London, UK, 1979. [Google Scholar]
- Capasso, R.; Evidente, A.; Randazzo, G.; Ritieni, A.; Bottalico, A.; Vurro, M.; Logrieco, A. Isolation of cytochalasins A and B from Ascochyta heteromorpha. J. Nat. Prod. 1987, 50, 989–990. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Berestetskiy, A.; Evidente, A. Production of phytotoxins by Phoma exigua var. exigua, a potential mycoherbicide against perennial thistles. J. Agric. Food Chem. 2008, 56, 6304–6309. [Google Scholar]
- Evidente, A.; Cimmino, A.; Andolfi, A.; Berestetskiy, A.; Motta, A. Phomachalasins A–D, 26-oxa [16] and [15] cytochalasans produced by Phoma exigua var. exigua, a potential mycoherbicide for Cirsium arvense biocontrol. Tetrahedron 2011, 67, 1557–1563. [Google Scholar]
- Bottalico, A.; Capasso, R.; Evidente, A.; Randazzo, G.; Vurro, M. Cytochalasins: Structure–activity relationships. Phytochemistry 1990, 29, 93–96. [Google Scholar] [CrossRef]
- Capasso, R.; Evidente, A.; Vurro, M. Cytochalasin from Phoma exigua var. heteromorpha. Phytochemistry 1991, 30, 3945–3950. [Google Scholar] [CrossRef]
- Vurro, M.; Bottalico, A.; Capasso, R.; Evidente, A. Cytochalasins from phytopathogenic Ascochyta and Phoma species. In Toxins in Plant Disease Development and Evolving Biotechnology; Upadhyay, R.K., Mukerji, K.G., Eds.; Oxford & IBH Publishing Co. PVT. Ltd.: New Delhi, India, 1997; pp. 127–147. [Google Scholar]
- Stahlman, P.W.; Miller, S.D. Downy brome (Bromus tectorum) interference and economic thresholds in winter wheat (Triticum aestivum). Weed Sci. 1990, 38, 224–228. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; Di Tomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Meyer, S.; Clement, S.; Andolfi, A.; Cimmino, A.; Evidente, A. Spirostaphylotrichin W, a spirocyclic γ-lactam isolated from liquid culture of Pyrenophora semeniperda, a potential mycoherbicide for cheatgrass (Bromus tectorum) biocontrol. Tetrahedron 2014, 70, 1497–1501. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Cimmino, A.; Andolfi, A.; Evidente, A. Pyrenophoric acid, a phytotoxic sesquiterpenoid penta-2, 4-dienoic acid produced by a potential mycoherbicide, Pyrenophora semeniperda. J. Nat. Prod. 2014, 77, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Meyer, S.; Cimmino, A.; Clement, S.; Black, B.; Evidente, A. Pyrenophoric acids B and C, two new phytotoxic sesquiterpenoids produced by Pyrenophora semeniperda. J. Agric. Food Chem. 2014, 62, 10304–10311. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; Masi, M.; Cimmino, A.; Clement, S.; Fernández, M.A.; Antoni, R.; Meyer, S.; Rodriguez, P.L.; Evidente, A. The fungal sesquiterpenoid pyrenophoric acid B uses the plant ABA biosynthetic pathway to inhibit seed germination. J. Exp. Bot. 2019, 70, 5487–5494. [Google Scholar] [CrossRef] [Green Version]
- Graupner, P.R.; Carr, A.; Clancy, E.; Gilbert, J.; Bailey, K.L.; Derby, J.A.; Gerwick, B.C. The macrocidins: Novel cyclic tetramic acids with herbicidal activity produced by Phoma macrostoma. J. Nat. Prod. 2003, 66, 1558–1561. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Abouzeid, M.A.; Vurro, M.; Zonno, M.C.; Motta, A. Ascosonchine, the enol tautomer of 4-pyridylpyruvic acid with herbicidal activity produced by Ascochyta sonchi. Phytochemistry 2004, 65, 475–480. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Vurro, M.; Fracchiolla, M.; Zonno, M.C.; Motta, A. Drazepinone, a trisubstituted tetrahydronaphthofuroazepinone with herbicidal activity produced by Drechslera siccans. Phytochemistry 2005, 66, 715–721. [Google Scholar] [CrossRef]
- Evidente, A.; Cimmino, A.; Andolfi, A.; Vurro, M.; Zonno, M.C.; Motta, A. Phyllostoxin and phyllostin, bioactive metabolites produced by Phyllosticta cirsii, a potential mycoherbicide for Cirsium arvense biocontrol. J. Agric. Food Chem. 2008, 56, 884–888. [Google Scholar] [CrossRef]
- Ayer, W.A.; Fukazawa, Y.; Orszanska, H. Scytolide, a new shikimate metabolite from the fungus Scytalidium uredinicola. Nat. Prod. Lett. 1993, 2, 77–82. [Google Scholar] [CrossRef]
- Irvine, N.M.; Yerkes, C.N.; Graupner, P.R.; Roberts, R.E.; Hahn, D.R.; Pearce, C.; Gerwick, B.C. Synthesis and characterization of synthetic analogs of cinnacidin, a novel phytotoxin from Nectria sp. Pest Manag. Sci. 2008, 64, 891–899. [Google Scholar] [CrossRef]
- Han, H.S.; Koh, Y.J.; Hur, J.S.; Jung, J.S. Identification and characterization of coronatine-producing Pseudomonas syringae pv. actinidiae. J. Microbiol. Biotechnol. 2003, 13, 110–118. [Google Scholar]
- Osbourn, A.E.; Lanzotti, V. Plant-Derived Natural Products; Springer: Dordrecht, The Netherlands, 2009; pp. 361–384. [Google Scholar]
- Evidente, A.; Berestetskiy, A.; Cimmino, A.; Tuzi, A.; Superchi, S.; Melck, D.; Andolfi, A. Papyracillic acid, a phytotoxic 1,6-dioxaspiro [4, 4] nonene produced by Ascochyta agropyrina var. nana, a potential mycoherbicide for Elytrigia repens biocontrol. J. Agric. Food Chem. 2009, 57, 11168–11173. [Google Scholar] [CrossRef]
- Shan, R.; Heidren, A.; Stadler, M.; Sterner, O. The reactivity of fungal papyracillic acid. Tetrahedron 1997, 53, 6209–6213. [Google Scholar] [CrossRef]
- Shan, R.; Heidren, A.; Stadler, M.; Sterner, O. Papyracillic acid, a new penicillic acid analogue from the ascomycete Lachnum papyraceum. Tetrahedron 1996, 52, 10249–102543. [Google Scholar] [CrossRef]
- Andolfi, A.; Cimmino, A.; Vurro, M.; Berestetskiy, A.; Troise, C.; Zonno, M.C.; Mtta, A.; Evidente, A. Agropyrenol and agropyrenal, phytotoxins from Ascochyta agropyrina var. nana, a fungal pathogen of Elitrigia repens. Phytochemistry 2012, 79, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Zonno, M.C.; Andolfi, A.; Troise, C.; Motta, A.; Vurro, M.; Evidente, A. Agropyrenol, a phytotoxic fungal metabolite, and its derivatives: A structure–activity relationship study. J. Agric. Food Chem. 2013, 61, 1779–1783. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Troise, C.; Santini, A.; Tuzi, A.; Vurro, M.; Ash, G.; Evidente, A. Phomentrioloxin: A phytotoxic pentasubstituted geranylcyclohexentriol produced by Phomopsis sp., a potential mycoherbicide for Carthamus lanatus biocontrol. J. Nat. Prod. 2012, 75, 1130–1137. [Google Scholar] [CrossRef]
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and Phomentrioloxins B and C produced by Diaporthe gulyae, a potential mycoherbicide for saffron thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef]
- Varejão, E.V.V.; Demuner, A.J.; Barbosa, L.C.D.A.; Barreto, R.W. Phytotoxic effects of metabolites from Alternaria euphorbiicola against its host plant Euphorbia heterophylla. Quím. Nova 2013, 36, 1004–1007. [Google Scholar] [CrossRef] [Green Version]
- Webster, T.M.; Nichols, R.L. Changes in the prevalence of weed species in the major agronomic crops of the Southern United States: 1994/1995 to 2008/2009. Weed Sci. 2012, 60, 145–157. [Google Scholar] [CrossRef]
- Sica, V.P.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Darveaux, B.A.; Pearce, C.J.; Oberlies, N.H. Optimizing production and evaluating biosynthesis in situ of a herbicidal compound, mevalocidin, from Coniolariella sp. J. Ind. Microbiol. Biotechnol. 2016, 43, 1149–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerwick, B.C.; Brewster, W.K.; de Boer, G.J.; Fields, S.C.; Graupner, P.R.; Hahn, D.R.; Pearce, C.J.; Schmitzer, P.R.; Webster, J.D. Mevalocidin: A novel, phloem mobile phytotoxin from Fusarium DA056446 and Rosellinia DA092917. J. Chem. Ecol. 2013, 39, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Meyer, S.; Clement, S.; Cimmino, A.; Cristofaro, M.; Evidente, A. Cochliotoxin, a dihydropyranopyran-4, 5-dione, and its analogues produced by Cochliobolus australiensis display phytotoxic activity against buffelgrass (Cenchrus ciliaris). J. Nat. Prod. 2017, 80, 1241–1247. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Clement, S.; Pescitelli, G.; Cimmino, A.; Cristofaro, M.; Evidente, A. Chloromonilinic acids C and D, phytotoxic tetrasubstituted 3-chromanonacrylic acids isolated from Cochliobolus australiensis with potential herbicidal activity against buffelgrass (Cenchrus ciliaris). J. Nat. Prod. 2017, 80, 2771–2777. [Google Scholar] [CrossRef]
- Sassa, T.; Kachi, H.; Nukina, M. Chloromonilicin, a new antifungal metabolite produced by Monilinia fructicola. J. Antibiot. 1985, 38, 439–441. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, A.; Pescitelli, G.; Berestetskiy, A.; Dalinova, A.; Krivorotov, D.; Tuzi, A.; Evidente, A. Biological evaluation and determination of the absolute configuration of chloromonilicin, a strong antimicrobial metabolite isolated from Alternaria sonchi. J. Antibiot. 2016, 69, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Meyer, S.; Górecki, M.; Mandoli, A.; Di Bari, L.; Pescitelli, G.; Cimmino, A.; Cristofaro, M.; Clement, S.; Evidente, A. Pyriculins A and B, two monosubstituted hex-4-ene-2,3-diols and other phytotoxic metabolites produced by Pyricularia grisea isolated from buffelgrass (Cenchrus ciliaris). Chirality 2017, 29, 726–736. [Google Scholar] [CrossRef]
- Masi, M.; Santoro, E.; Clement, S.; Meyer, S.; Scafato, P.; Superchi, S.; Evidente, A. Further secondary metabolites produced by the fungus Pyricularia grisea isolated from buffelgrass (Cenchrus ciliaris). Chirality 2020, 32, 1234–1242. [Google Scholar] [CrossRef]
- Meepagala, K.M.; Clausen, B.M.; Johnson, R.D.; Wedge, D.E.; Duke, S.O. A phytotoxic and antifungal metabolite (pyrichalasin H) from a fungus infecting Brachiaria eruciformis (signal grass). J. Agric. Chem. Environ. 2019, 8, 115–118. [Google Scholar]
- Waller, J.M.; Lenne, J.M.; Waller, S.J. Plant Pathologist’s Pocketbook, 3rd ed.; CABI: Wallingford, UK, 2012; pp. 4–18. [Google Scholar]
- Damm, U.; O’connell, R.J.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum destructivum species complex-hemibiotrophic pathogens of forage and field crops. Stud. Mycol. 2014, 79, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Cimmino, A.; Boari, A.; Tuzi, A.; Zonno, M.C.; Baroncelli, R.; Vurro, M.; Evidente, A. Colletochlorins E and F, new phytotoxic tetrasubstituted pyran-2-one and dihydrobenzofuran, isolated from Colletotrichum higginsianum with potential herbicidal activity. J. Agric. Food Chem. 2017, 65, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Cimmino, A.; Boari, A.; Zonno, M.C.; Górecki, M.; Pescitelli, G.; Tuzi, A.; Vurro, M.; Evidente, A. Colletopyrandione, a new phytotoxic tetrasubstituted indolylidenepyran-2,4-dione, and colletochlorins G and H, new tetrasubstituted chroman-and isochroman-3,5-diols isolated from Colletotrichum higginsianum. Tetrahedron 2017, 73, 6644–6650. [Google Scholar] [CrossRef]
- Masi, M.; Zonno, M.C.; Cimmino, A.; Reveglia, P.; Berestetskiy, A.; Boari, A.; Vurro, M.; Evidente, A. On the metabolites produced by Colletotrichum gloeosporioides a fungus proposed for the Ambrosia artemisiifolia biocontrol; spectroscopic data and absolute configuration assignment of colletochlorin A. Nat. Prod. Res. 2018, 2, 1537–1547. [Google Scholar] [CrossRef]
- Marsico, G.; Pignataro, B.A.; Masi, M.; Evidente, A.; Casella, F.; Zonno, M.C.; Takd, J.-H.; Bloomiquist, J.R.; Stefano, S.; Scafato, P. Asymmetric synthesis and structure-activity studies of the fungal metabolites colletorin A, colletochlorin A and their halogenates analogues. Tetrahedron 2018, 74, 3912–3923. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, M.; Tian, Y.; Zhao, P.; Niu, Y.; Liao, M. Dirhamnolipid produced by the pathogenic fungus Colletotrichum gloeosporioides BWH-1 and its herbicidal activity. Molecules 2019, 4, 2969. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.J.; Yang, X.; Wang, X.R.; Zeng, Y.S.; Liao, M.D.; Chen, C.J.; Jia, D.M. First report of anthracnose disease on young stems of Bawanghua (Hylocereus undatus) caused by Colletotrichum gloeosporioides in China. Plant Dis. 2014, 98, 991. [Google Scholar] [CrossRef]
- Poluektova, E.; Tokarev, Y.; Sokornova, S.; Chisty, L.; Evidente, A.; Berestetskiy, A. Curvulin and phaeosphaeride A from Paraphoma sp. VIZR 1.46 isolated from Cirsium arvense as potential herbicides. Molecules 2018, 23, 2795. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Meyer, S.; Górecki, M.; Pescitelli, G.; Clement, S.; Cimmino, A.; Evidente, A. Phytotoxic activity of metabolites isolated from Rutstroemia sp., the causal agent of bleach blonde syndrome on cheatgrass (Bromus tectorum). Molecules 2018, 23, 1734. [Google Scholar] [CrossRef] [Green Version]
- Dalinova, A.; Chisty, L.; Kochura, D.; Garnyuk, V.; Petrova, M.; Prokofieva, D.; Darya Prokofieva, D.; Yurchenko, A.; Dubovik, V.; Ivanov, A.; et al. Isolation and bioactivity of secondary metabolites from solid culture of the fungus, Alternaria sonchi. Biomolecules 2020, 10, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, M.; Boari, A.; Sautua, F.; Carmona, M.A.; Vurro, M.; Evidente, A. Araufuranone: A new phytotoxic tetrasubstituted dihydrofuro [3, 2-b] furan-2 (5 H)-one isolated from Ascochyta araujiae. Biomolecules 2022, 12, 1274. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.R.; Press, M.C. The physiology and biochemistry of parasitic angiosperms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 127–151. [Google Scholar] [CrossRef]
- Cook, C.E.; Whichard, L.; Turner, B.; Wall, M.E.; Egley, G.H. Germination of witchweed (Striga lutea): Isolation and properties of a potent stimulant. Science 1966, 154, 1189–1190. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.E.; Whichard, L.P.; Wall, M.E.; Egley, G.H.; Coggon, P.; Luhan, P.A.; McPhail, A.T. Germination stimulants. II. The structure of strigol a potent seed germination stimulant for witchweed (Striga lutea Lour.). J. Am. Chem. Soc. 1972, 94, 6198–6199. [Google Scholar] [CrossRef]
- Joel, D.M.; Gressel, J.; Musselman, L.J. Parasitic Orobanchaceae. Parasitic Mechanisms and Control Strategies; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Capasso, R.; Evidente, A.; Cutignano, A.; Vurro, M.; Zonno, M.C.; Bottalico, A. Fusaric and 9, 10-dehydrofusaric acids and their methyl esters from Fusarium nygamai. Phytochemistry 1996, 41, 1035–1039. [Google Scholar] [CrossRef]
- Abbasher, A.A.; Sauerborn, J. Fusarium nygamai, a potential bioherbicide for Striga hermonthica control in sorghum. Biocontrol 1992, 2, 291–296. [Google Scholar] [CrossRef]
- Dor, E.; Hershenhorn, J.; Andolfi, A.; Cimmino, A.; Evidente, A. Fusarium verticillioides as a new pathogen of the parasitic weed Orobanche spp. Phytoparasitica 2009, 37, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Marra, M.; Camoni, L.; Visconti, S.; Fiorillo, A.; Evidente, A. The surprising story of fusicoccin: A wilt-inducing phytotoxin, a tool in plant physiology and a 14-3-3-targeted drug. Biomolecules 2021, 11, 1393. [Google Scholar] [CrossRef]
- Sassa, T.; Togashi, M.; Kitaguchi, T. The structures of cotylenins A, B, C, D and E. Agric. Biol. Chem. 1975, 39, 1735–1744. [Google Scholar]
- Sassa, T.; Neguro, T.; Ueki, H. Production and characterization of a new fungal metabolite, cotylenol. Agric. Biol. Chem. 1972, 36, 2281–2285. [Google Scholar] [CrossRef]
- Yoneyama, K.; Takeuchi, Y.; Ogasawara, M.; Konnai, M.; Sugimoto, Y.; Sassa, T. Cotylenins and fusicoccins stimulate seed germination of Striga hermonthica (Del.) Benth and Orobanche minor Smith. J. Agric. Food Chem. 1998, 46, 1583–1586. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Fiore, M.; Boari, A.; Vurro, M. Stimulation of Orobanche ramosa seed germination by fusicoccin derivatives: A structure–activity relationship study. Phytochemistry 2006, 67, 19–26. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Andolfi, A.; Cimmino, A.; Rubiales, D.; Evidente, A. Stimulation of seed germination of Orobanche species by ophiobolin A and fusicoccin derivatives. J. Agric. Food Chem. 2008, 56, 8343–8347. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.J.; Cox, R.H. Handobook of Toxic Metabolites; Academic Press: New York, NY, USA, 1981; pp. 152–263. [Google Scholar]
- Zonno, M.C.; Vurro, M. Inhibition of germination of Orobanche ramosa seeds by Fusarium toxins. Phytoparasitica 2002, 30, 519–524. [Google Scholar] [CrossRef]
- Andolfi, A.; Boari, A.; Evidente, A.; Vurro, M. Metabolites inhibiting germination of Orobanche ramosa seeds produced by Myrothecium verrucaria and Fusarium compactum. J. Agric. Food Chem. 2005, 53, 1598–1603. [Google Scholar] [CrossRef]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Scanu, B.; Evidente, A.; Cimmino, A. Bioactive metabolites from pathogenic and endophytic fungi of forest trees. Curr. Med. Chem. 2018, 25, 208–252. [Google Scholar] [CrossRef]
- Evidente, A.; Randazzo, G.; Iacobellis, N.S.; Bottalico, A. Structure of cavoxin, a new phytotoxin from Phoma cava and cavoxone, its related chroman-4-one. J. Nat. Prod. 1985, 48, 916–923. [Google Scholar] [CrossRef]
- Cimmino, A.; Fernandez-Aparicio, M.; Andolfi, A.; Basso, S.; Rubiales, D.; Evidente, A. Effect of fungal and plant metabolites on broomrapes (Orobanche and Phelipanche spp.) seed germination and radicle growth. J. Agric. Food Chem. 2014, 62, 10485–10492. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Masi, M.; Maddau, L.; Cimmino, A.; Evidente, M.; Rubiales, D.; Evidente, A. Induction of haustorium development by sphaeropsidones in radicles of the parasitic weeds Striga and Orobanche. A structure–activity relationship study. J. Agric. Food Chem. 2016, 64, 5188–5196. [Google Scholar] [CrossRef]
- Triolet, M.; Guillemin, J.P.; Andre, O.; Steinberg, C. Fungal-based bioherbicides for weed control: A myth or a reality? Weed Res. 2020, 60, 60–77. [Google Scholar] [CrossRef]
- Roberts, J.; Florentine, S.; Fernando, W.D.; Tennakoon, K.U. Achievements, developments and future challenges in the field of bioherbicides for weed control: A global review. Plants 2022, 11, 2242. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Company | Microorganism Formulated | Weed 1 | Colture 2 |
|---|---|---|---|---|
| Collego | TUCO, Amarillo, TX, USA | Colletotrichum gleosporioides f. sp. aeschynomene | Aeschynomene virginica, Aeschynomene indica and Sesbania exaltata | Rice, soybean |
| De Vine | Abott Laboratories, Chicago, IL, USA | Phytophthora palmivora f. sp. aeschinomene | Morrenia odorata | Citrus groves |
| BIOMAL | Novartis Biomedical research, Dorval, QC, Canada | Colletotrichum gleosporioides f. sp. malvae | Malva pusilla | Different |
| Stompout | CAB International, Wallingford, UK | Cylindrobasidium leave | Acacia spp. | Natural enviroments |
| Dr. Biosedge | Bicosis (Adavanced Biocontrol System) Co., Ltd., Geochang-gun Gyeongsangnam-Do, Korea | Puccinia canaliculata | Cyperus esculentus | Different |
| Myco.Tech | MycoTechnology, Inc., Aurora, CO, USA | Chondostereum purpureum | Trees | Forests |
| Camperico | Japan Tobacco, Inc., Toranomon, Minato-Ku, Tokyo, Japan | Xanthomonas campestris subsp. poae | Poa annua | Golf courses |
| SolviNix | BIOPRODEX, INC., Gainesville, FL, USA | Tobacco mild green mosaic virus | Solanum viarum | Perennial grass pastures |
| Sarritor | McGill Spin-off Anomera Inc, Montreal, QC, Canada | Sclerotim minor | Traxonum officinale | Not reported |
| Phytotoxin | Weed | Fungus | Biological Activity | References |
|---|---|---|---|---|
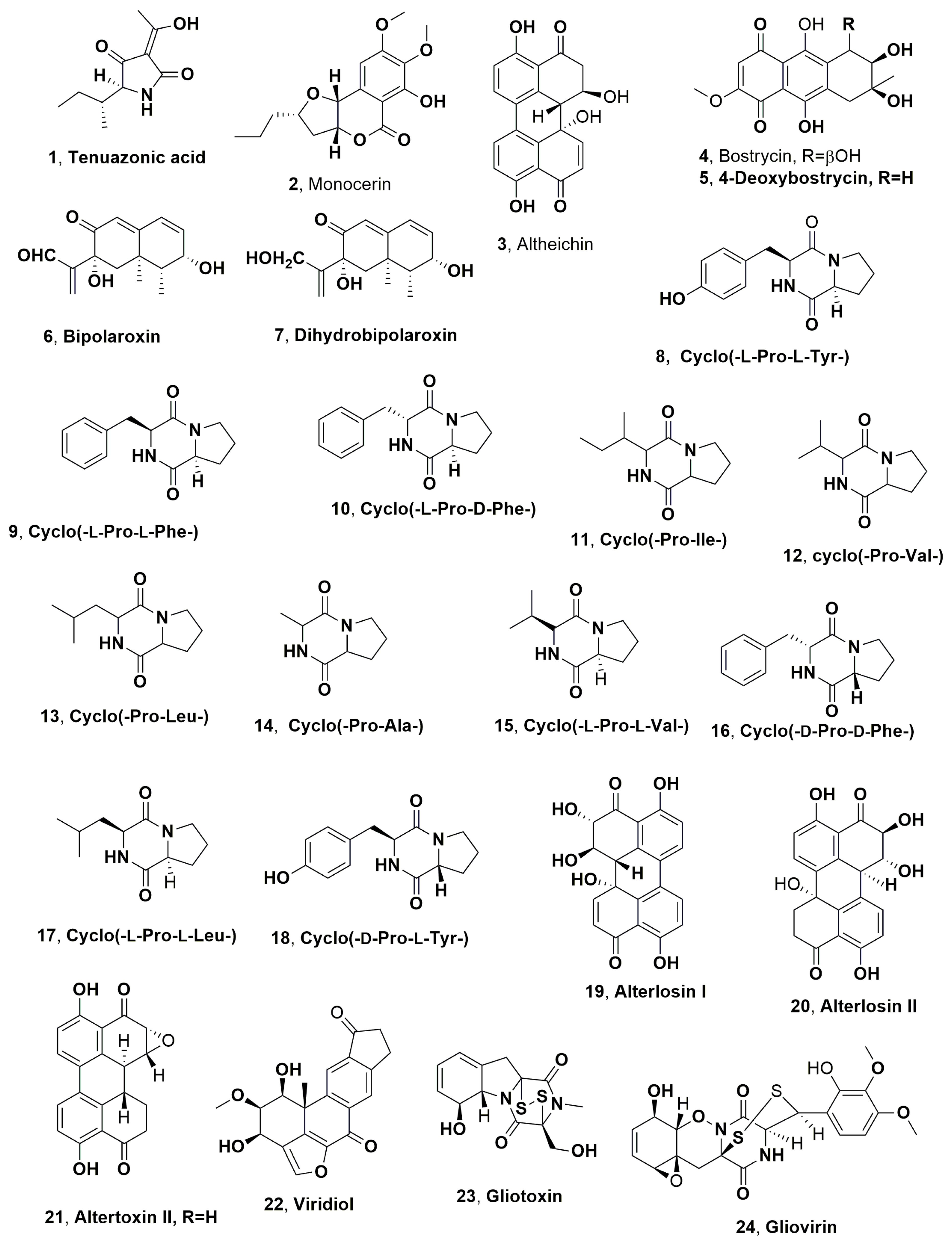
| Tuenazoic acid (1) | Johnsongrass (Sorghum halepense) L. | Alternaria alternata Alternaria longipes | Phytotoxic activity on tobacco leaves | [44] [46] |
| Monocerin (2) | “ | Exserohilum turcicum | Phytotoxic activity on the host plant, tomato and Cirsium arvenses | [47] |
| Altheichin (3) | Water hyacinth (Eichornia crassipes) | Alternaria eichorniae Alternaria alternata | Phytotoxic activity on the host plant water hyacinth (Eichornia crassipes) and non-host plant Pigment | [48] [69] |
| Bostrycin (4) | “ | Alternaria eichorniae | Phytotoxc activity on the host plant water hyacinth (Eichornia crassipes) and some agrarian and weedy plants | [49] |
| 4-Deoxybostrycin (5) | “ | “ | “ | “ |
| Bipolaroxin (6) | Bermuda grass (Cynodon dactylon L.) | Bipolaris cynodontis | Specific activity against the two host plants Bermuda grass and johnson grass | [50] |
| Dihydrobipolaroxin (7) | “ | “ | No toxicity | “ |
| Cyclo(-L-Pro-L-Tyr-) (8) | Knapweed, (Centaurea maculosa) | Alternaria alternata | Phytotoxic activity against the host knapweed, Antifungal Antibiotic | [51] [53,55] [60] |
| Cyclo(-L-Pro-L-Phe-) (9) | “ | “ | Phytotoxic activity against the host knapweed | [51] |
| cyclo(-L-Pro-D-Phe-) (10) | “ | “ | No toxic on host plant | “ |
| cyclo(-Pro-Hle-) (11) | “ | “ | “ | “ |
| cyclo(-Pro-Val-) (12) | “ | “ | “ | “ |
| cyclo(-Pro-Leu-) (13) | “ | “ | “ | “ |
| cyclo(-Pro-Ala-) (14) | “ | “ | “ | “ |
| cyclo(-L-Pro-L-Val-) (15) | - | Lysobacter capsici | Antifungal activity Antibiotic activity | [60] |
| cyclo(-D-Pro-D-Phe-) (16) | “ | “ | Antifungal activity | “ |
| cyclo(-L-Pro-L-Leu-) (17) | “ | “ | “ | “ |
| cyclo(-D-Pro-L-Tyr-) (18) | “ | “ | “ | “ |
| Alterlosin I (19) | Knapweed (Centaurea maculosa) | Alternaria alternata | Phytotoxic on knapweed | [68] |
| Alterlosin II (20) | “ | “ | “ | “ |
| Altertoxin III (21) | “ | “ | Phytotoxic on lettuge Mutagenic. | [69] [70] |
| Viridiol (22) | Several spp. | Gliocladium virens Hymenoscyphus fraxineus | Phytotoxic activity on weeds Phytotoxic activity on some forest plant | [71] [74] |
| Gliotoxin (23) | Several spp. | Gliocladium virens | Antibiotic, Antifungal | [71,72] |
| Gliovirin (24) | “ | “ | “ | [71,73] |
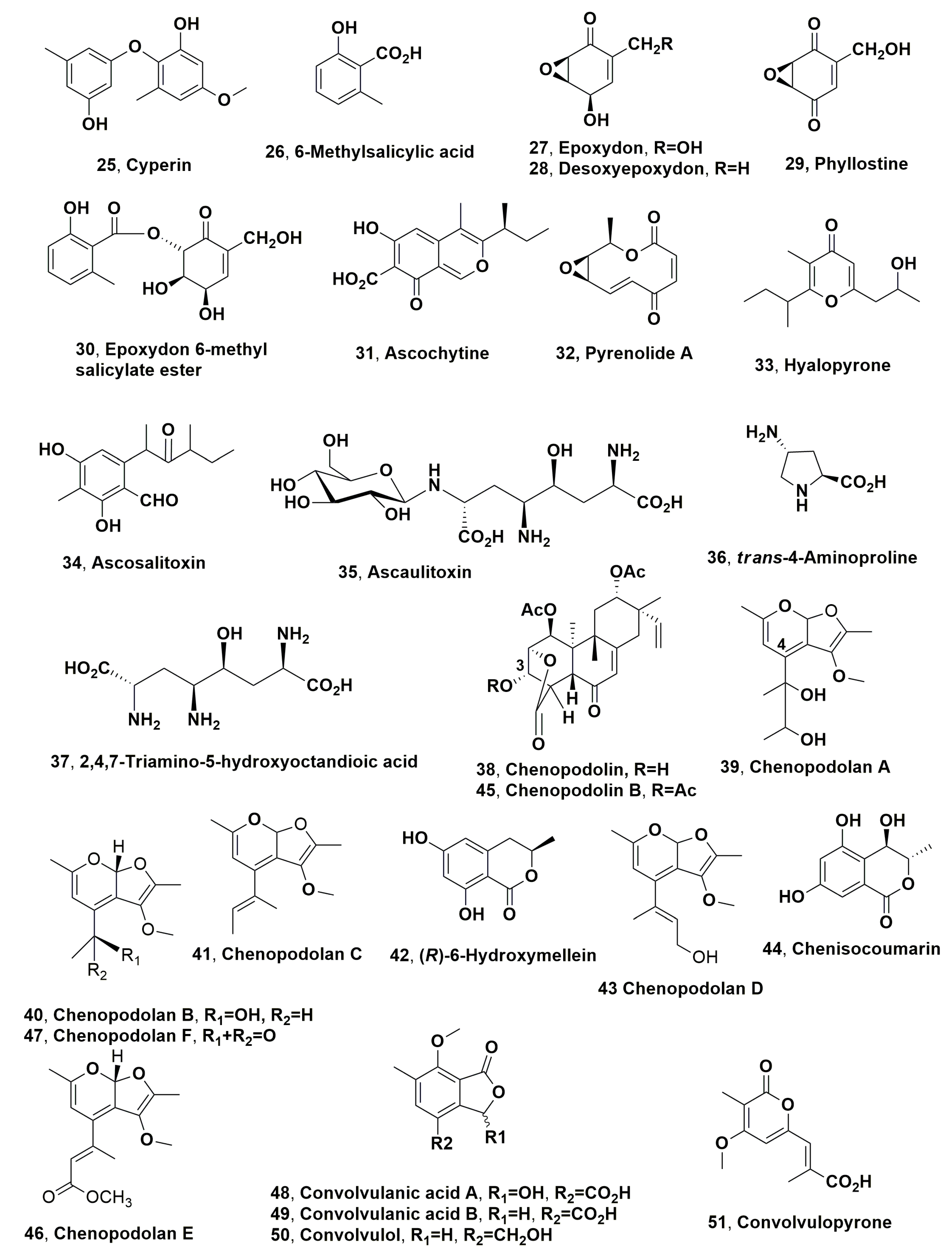
| Cyperine (25) | Purple nutsedge (Cyperus rotundus L.) | Ascochyta cypericola | Phytotoxic activity on Cyperus spp. | [76] |
| 6-Methylsalicylic acid (26) | Pockeweed (Phytolacca americana L.) | Phoma sorghina | Phytotoxic activity on the host pokeweed and 8 other weeds | [77] |
| Epoxydon (27) | “ | “ | “ | “ |
| Desoxyepoxydon (28) | “ | “ | “ | “ |
| Phyllostine (29) | “ | “ | “ | “ |
| Epoxydon 6-methylsalicylate ester (30) | “ | “ | “ | “ |
| Ascochytine (31) | Lambsquarters or ft hen (Chenopodium album L.) | Ascochyta hyalospora | Phytotoxic activity on the host lambsquarters and 8 other weeds | [84] |
| Pyrenolide A (32) | “ | “ | “ | |
| Hyalopyrone (33) | “ | “ | “ | “ |
| Ascosalitoxin (34) | Pea and bean (Pisum sativum and Phaseolus vulgaris) | Ascochyta pisi | Phytotoxic activity on the host plants | [85] |
| Ascaulitoxin (35) | “ | Ascochyta caulina | Phytotoxic activity on the host lambsquarters | [88] |
| trans-4-Aminoproline (36) | “ | “ | “ | [89] |
| 2,4,7-Triamino-5-hydroxyoctandioic acid (37) | “ | “ | “ | [90] |
| Chenopodolin (38) | “ | Phoma chenopodiicola | Phytotoxic activity on Mercurialis annua, Cirsium arvense and Setaria viride | [95] |
| Chenopodolan A (39) | “ | “ | Phytotoxic activity on Sonchus oleraceus, M. annua and C. album | [96] |
| Chenopodolan B (40) | “ | “ | “ | “ |
| Chenopodolan C (41) | “ | “ | No phytoxicity | “ |
| (R)-6-Hydroxymellein (42) | “ | “ | No phytoxicity | “ |
| Chenopodolan D (43) | “ | “ | Phytotoxic activity on Sonchus arvensis, Urtica dioica and Parietaria officinalis | [97] |
| Chenisocoumarin (44) | “ | “ | No phytotoxic | “ |
| Chenopodolin B (45) | “ | “ | Phytotoxic activity on S. arvensis, U. dioica and P. officinalis | “ |
| Chenopodolan E (46) | “ | “ | Zootoxic acrivity | [98] |
| Chenopodolan F (47) | “ | “ | Phytotoxic activity on S. arvensis | “ |
| Convolvulanic acid A (48) | Bindweed (Corzvolvulus arvensis) | Phomopsis convolvulus | Phytotoxic activity against bindweed (Corzvolvulus arvensis) | [99] |
| Convolvulanic acid B (49) | “ | “ | “ | “ |
| Convolvulol (50) | “ | “ | “ | “ |
| Convolvulopyrone (51) | “ | “ | “ | “ |
| Ergosterol (52) | “ | “ | No toxicity | “ |
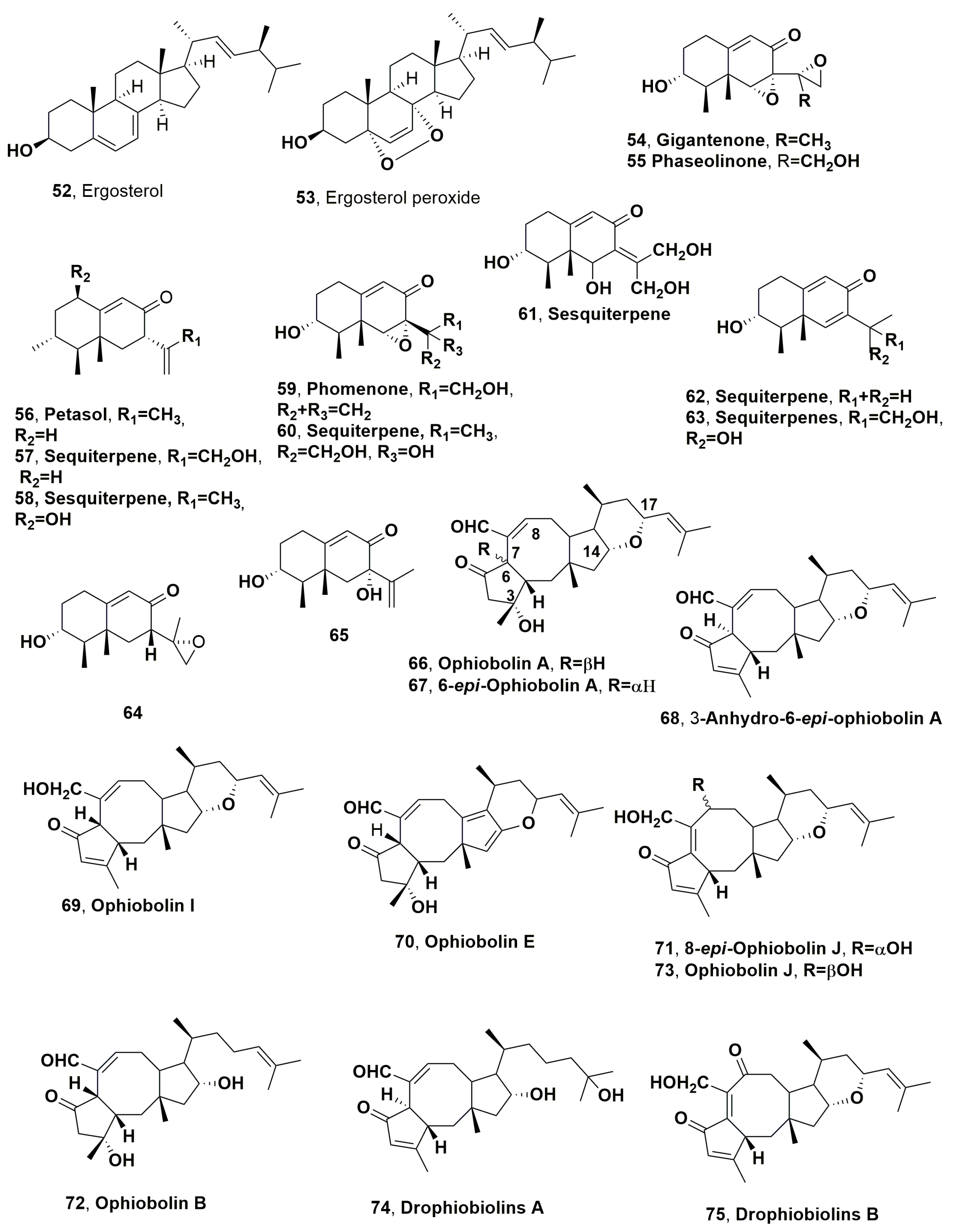
| Ergosterol peroxide (53) | “ | “ | “ | “ |
| Gigantenone (54) | Several grasses as weed cabgrass (Digitaria spp.), quackgrass (Agropyron repens) and Bermuda grass (Cynodon dactylon) | Drechslera gigantea | Phytotoxic activity on monocot species | [100] |
| Phaseolinone (55) | “ | “ | “ | “ |
| Petasol (56) | “ | “ | “ | “ |
| Sesquiterpenes (57) | “ | “ | Not Toxic | “ |
| Sequiterpene(58) | “ | “ | Phytotoxic activity on dicot species | “ |
| Phomenone (59) | “ | “ | Phytotoxic activity on monocot species | “ |
| Sesquiterpenes (60) | “ | “ | Phytotoxic activity on dicot species | “ |
| Sesquiterpenes (61) | “ | “ | “ | “ |
| Sesquiterpenes (62) | “ | “ | Not Toxic | “ |
| Sesquiterpenes (63) | “ | “ | “ | “ |
| Sesquiterpenes (64) | “ | “ | “ | “ |
| Sesquiterpenes (65) | “ | “ | “ | “ |
| Ohpiobolin A (66) | “ | “ | Phytotoxic activity against some monocot grasses and dicot weeds | [105] |
| 6-epi-Ophiobolin A (67) | “ | “ | Reduced phytotoxicity against some monocot grasses and dicot weeds | “ |
| 3-Anhydro-6-epi-ophiobolin A (68) | “ | “ | “ | “ |
| Ophiobolin I (69) | “ | “ | No Toxicity | “ |
| ophiobolin E (70) | “ | “ | “ | [118] |
| 8-epi-Ophiobolin J (71) | “ | “ | “ | “ |
| Ophiobolins B (72) | “ | “ | Phytotoxic activity against some weeds | “ |
| Ophiobolin J (73) | “ | “ | Reduce or no toxicity against some weeds | “ |
| Drophiobiolins A (74) | “ | “ | Phytotoxic activity on host, other weed and agrarian plants Cytotoxic | [119] |
| Drophiobiolins B (75) | “ | “ | “ | “ |
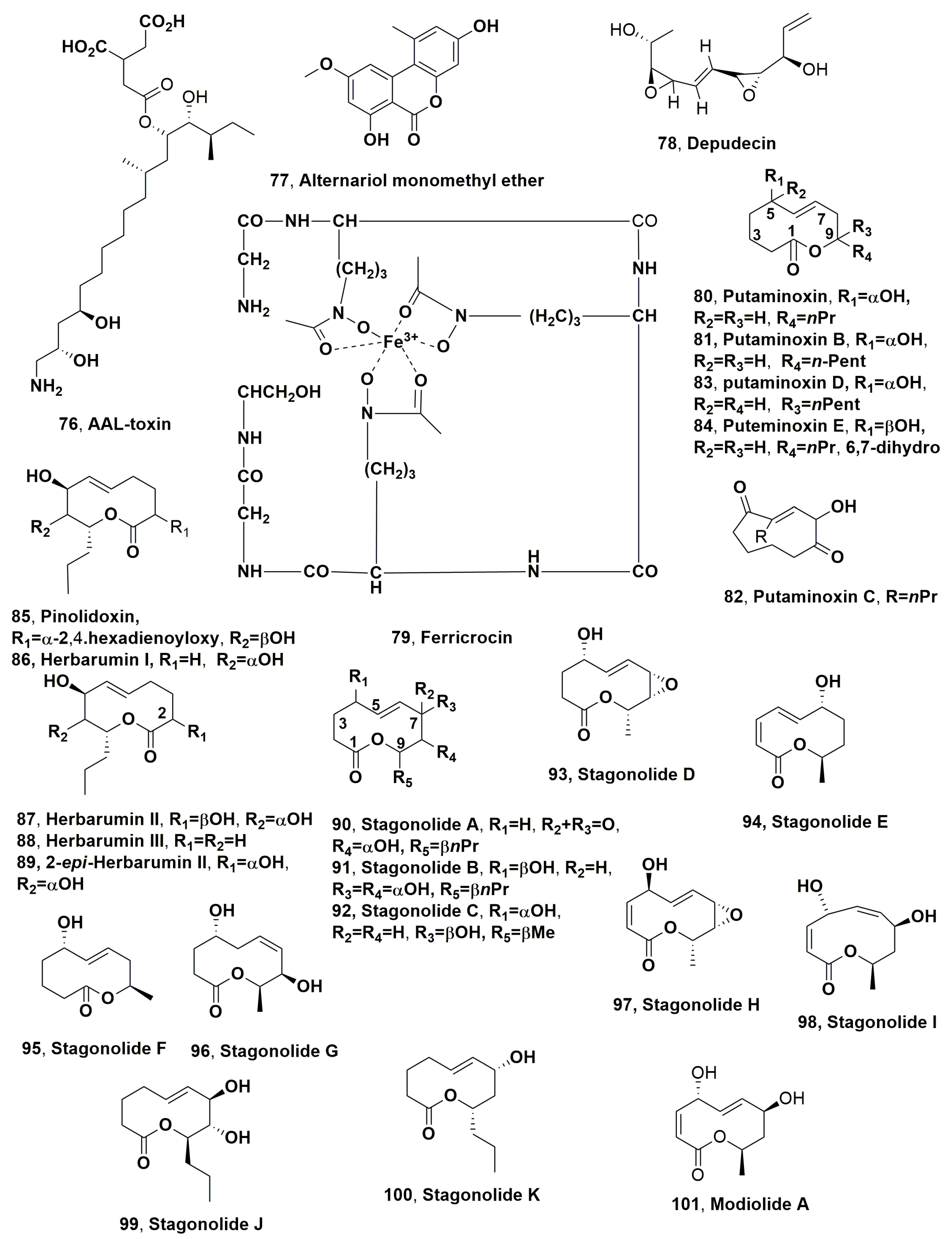
| AAL-toxin (76) | Jimsonweed (Datura stramonium) | Alternaria alternata | Phytotoxic activity on jimsonweed | [120] |
| Alternariol monomethyl ether (77) | “ | “ | No toxicity | “ |
| Depudecin (78) | Kuroguwai (Eleocharis kuroguwai) | Nimbya scirpicola Alternaria brassicola | Phytotoxic activity on host weed and some crops Anticancer | [134] [135] |
| Ferrocinin (79) | blackberry (Rubus spp.) | Colletotrichum gloeosporioides | Phytotoxic activity on some weeds | [136] |
| Putaminoxin (80) | Annual flaebane (Erigeron annuus) | Phoma putaminum | Phytotoxic activity on the host annual fleabane, some other weeds and cultivated plants | [141] |
| Putaminoxin B (81) | “ | “ | No toxicity | [142] |
| Putaminoxin C (82) | “ | “ | Phytotoxic activity on annual dog’s mercury, globe artichoke, tomato. | “ |
| Putaminoxin D (83) | “ | “ | No toxicity | [143] |
| Putaminoxin E (84) | “ | “ | “ | “ |
| Pinolidoxin (85) | Pisum sativum L. | Dydimella pinodes | Phytotoxic activity on some weed and crop plants | [144] |
| Herbarumin I (86) | Prine’s father (Amaranthus hypochondriacus) | Phoma herbarum | Phytotoxic activity against Amaranthus hypochondriacus | [147] |
| Herbarumin II (87) | “ | “ | “ | “ |
| Herbarumin III (88) | “ | “ | “ | [148] |
| 2-epi-Herbarumin II (89) | Pendulus yucca (Yucca recurvifolici) | Paraphaeosphaeria recurvifoliae | Inhibition of murine tyrosinase | [149] |
| Stagonolide A (90) | Cirsium arvense (Canada thisle) | Stagonospora cirsii | Phytotoxic activity against the host plant C. arvense, and several other weeds and cultivated plants | [151] |
| Stagonolide B (91) | “ | “ | No phytotoxicity | [152] |
| Stagonolide C (92) | “ | “ | “ | “ |
| Stagonolide D (93) | “ | “ | “ | “ |
| Stagonolide E (94) | “ | “ | “ | “ |
| Stagonolide F (95) | “ | “ | “ | “ |
| Stagonolide G (96) | “ | “ | “ | [153] |
| Stagonolide H (97) | “ | “ | Strong phytotoxicity against the host plants | “ |
| Stagonolide I (98) | “ | “ | Moderate phytotoxicity against the host plants | “ |
| Stagonolide J (99) | “ | “ | No toxicity | [154] |
| Stagonolide K (100) | “ | “ | Phytotoxicity against C. arvense and S. arvensis | “ |
| Modiolide A (101) | “ | “ | Moderate phytotoxicity against the host plants | [153] |
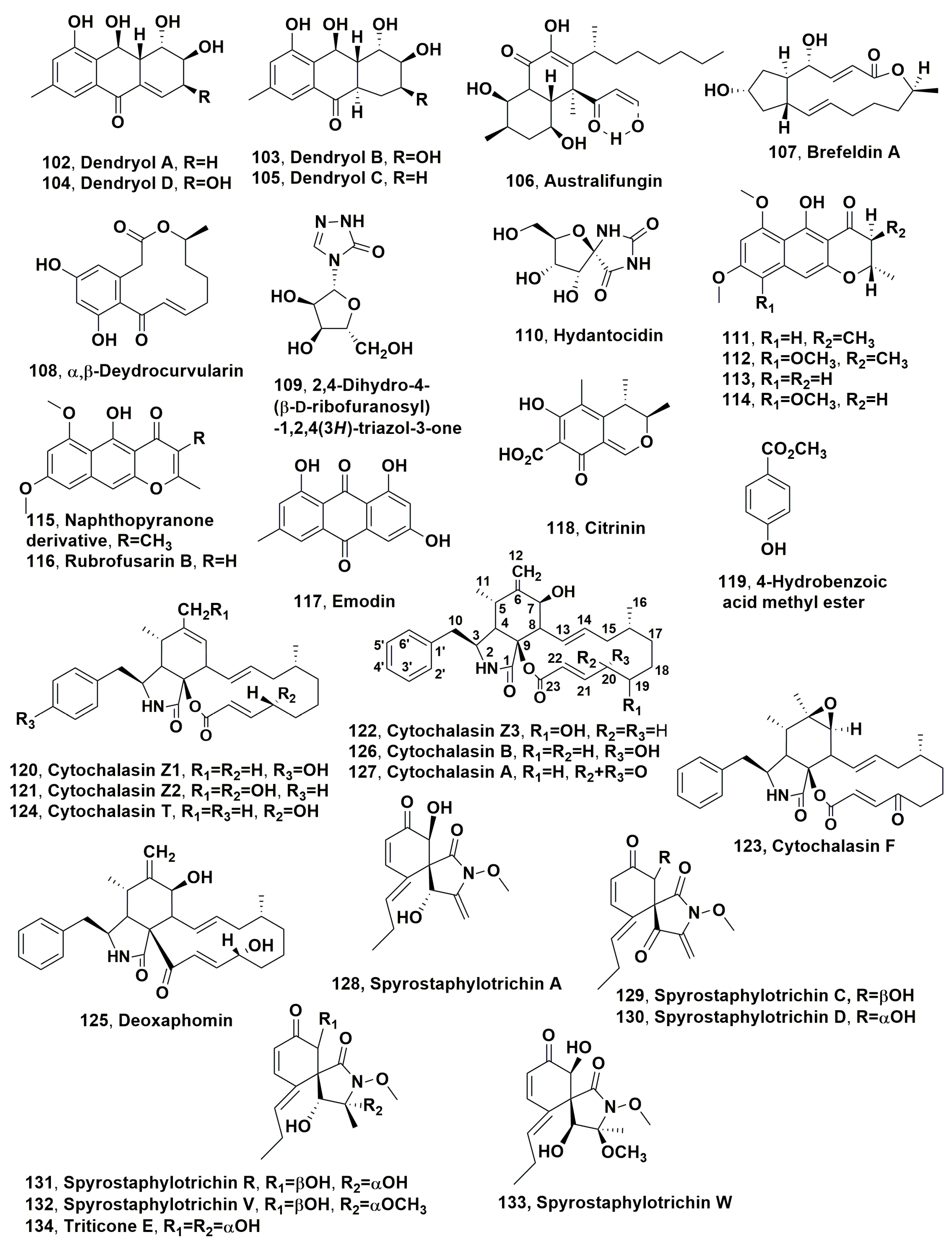
| Dendryol A (102) | Kuroguwai (Eleocharis kuroguwai) | Dendryphiella sp. | Phytotoxic activity against barnyardgrass | [159] |
| Dendryol B (103) | “ | “ | “ | “ |
| Dendryol C (104) | “ | “ | “ | “ |
| Dendryol D (105) | “ | “ | “ | “ |
| Australifungin (106) | No infected weed | Sporormiella australis | Phytotoxicity against duckweed (Lemna pausicostata) | [160] |
| Brefeldin (107) | Alternaria zinniae | Xanthium occidentale. | Phytotoxicity against the host plant and other weeds | [161] |
| α,β-Dehydrocurvularin (108) | “ | “ | “ | “ |
| 2,4-Dihydro-4-(β-D-ribofuranosyl)-1,2,4(3H)-triazol-3-one (109) | No infected weed | Actinomadura sp. | Phytotoxic activity on several weeds | [163,168] |
| Hydantocidin (110) | “ | Streptomyces hygroscopicus | “ | [165,168] |
| Naphthopyranone derivatives (111) | “ | Guanomyces polythrix | Phytotoxic activity against Amaranthus hypochondriacus and Echinochloa crusgalli | [173] |
| Naphthopyranone derivatives (112) | “ | “ | “ | “ |
| Naphthopyranone derivatives (113) | “ | “ | “ | “ |
| Naphthopyranone derivatives (114) | “ | “ | “ | “ |
| Naphthopyranone derivatives (115) | “ | “ | “ | “ |
| Rubrofusarin B (116) | “ | “ | “ | “ |
| Emodin (117) | “ | “ | “ | “ |
| Citrinin (118) | “ hemp dogbane (Apocynum cannabinum L.). | “ Stagonospora apocyni | “ Toxicity on several weeds | “ [174] |
| 4-Hydroxybenzoic acid methyl ester (119) | No infected weed | Guanomyces polythrix | Phytotoxic activity against Amaranthus hypochondriacus and Echinochloa crusgalli | [173] |
| Cytochalasin Z1 (120) | Annual grasses (Bromus spp.) | Pyrenophora semeniperda | Phytotoxicity on wheat and tomato | [175] |
| Cytochalasin Z2 (121) | “ | “ | “ | “ |
| Cytochalasin Z3 (122) | “ | “ | “ | “ |
| Cytochalasins F (123) | “ | “ | “ | “ |
| Cytochalsin T (124) | “ | “ | “ | “ |
| Deoxaphomin (125) | “ | “ | “ | “ |
| Cytochalasins B (126) | “ | “ | “ | “ |
| Cytochalasin A (127) | Oleander nerium L. | Phoma exigua var. heromorpha | Not tested on weed | [178] |
| Spirostaphylotrichin A (128) | Cheatgrass (Bromus tectorum) | Pyrenophora semeniperda | Phytotoxicity on cheatgrass and non-host plants | [185] |
| Spirostaphylotrichin C (129) | “ | “ | “ | “ |
| Spirostaphylotrichin D (130) | “ | “ | “ | “ |
| Spirostaphylotrichin R (131) | “ | “ | Non-toxic on cheatgrass | “ |
| Spirostaphylotrichin V (132) | “ | “ | Phytotoxicity on cheatgrass | “ |
| Spirostaphylotrichin W (133) | “ | “ | “ | “ |
| Triticone E (134) | “ | “ | Non-toxic on cheatgrass | “ |
| Pyrenophoric acid (135) | “ | “ | Phytotoxic activity on cheatgrass | [186] |
| Pyrenophoric acid B (136) | “ | “ | “ | [187] |
| Pyrenophoric acid C (137) | “ | “ | “ | “ |
| Abscisic acid (138) | “ | “ | “ | “ |
| Macrocidins A (139) | Canada thistle (Cirsium arvense) | Phoma macrostoma | Phytotoxicity on different weeds | [189] |
| Macrocidins B (140) | “ | “ | “ | “ |
| Ascosonchine (141) | Sowthistle (Socnhus arvensis) | Ascochyta sonchi | Phytotoxicity on several weed and cultivated plants | [190] |
| Drazepinone (142) | Ryegrass (Lolium perenne) | Drechslera siccans | Phytotoxicity on several weeds | [191] |
| Phyllostoxin (143) | Canada thistle Cirsium arvense | Phyllosticta cirsii | Phytotoxicity on host plant | [192] |
| Phyllostin (144) | “ | “ | No toxicity | “ |
| Scytolide (145) | Pine-pine gall rust (Endocronartium harknessii) | Scytalidium uredinicola | Inhibiotion of inhibition of germination of E. harknessii spores | [193] |
| Cinnamicidin (146) | Not identified | Nectria sp. | Phytotoxicity on several weeds | [194] |
| Coronatine (147) | “ | Pseudomonas syringae | “ | [195] |
| Jasmonic acid (148) | “ | Plant hormone and fungal metabolites | “ | [196] |
| Papyracillic acid (149) | Quack grass (Elytrigia repens) | Ascochyta agropyrina var. nana | Phtotoxicity against different weeds | [197] |
| Papyracillic acid methyl acetal (150) | “ | “ | “ | “ |
| Agropyrenol (151) | “ | “ | “ | [200] |
| Agropyrenal (152) | “ | “ | “ | “ |
| Agropyrenone (153) | “ | “ | Not toxic | “ |
| Phomentrioloxin (154) | Saffron twistle (C. arvense) | Phomopsis sp. | Phytotoxicity on several weeds and some cultivated plants | [202] |
| Gulypyrone A (155) | “ | Dyaporthe gulyae | No toxicity | [203] |
| Gulypyrone B (156) | “ | “ | Phytotoxicity on Helianthus annuus plantlets | “ |
| Phomentrioloxin B (157) | “ | “ | Phytotoxicity on several weeds and some cultivated plants | “ |
| Phomentrioloxin C (158) | “ | “ | “ | “ |
| Anhydromevalonolactone (159) | wild poinsettia (Euphorbia heterophylla) | Alternaria euphorbiicola | Phytotoxicity on host plant and other weed | [204] |
| Tyrosol (160) | “ | “ | “ | “ |
| (R)-(–)-Mevalonolactone (161) | “ | “ | “ | “ |
| Cycloglycylprolin (162) | “ | “ | Selective toxicity against the host plant | “ |
| Mevalocidin (163) | No infected weed | Coniolariella sp. | Phytotoxicity against broadleaf and grass species | [206,207] |
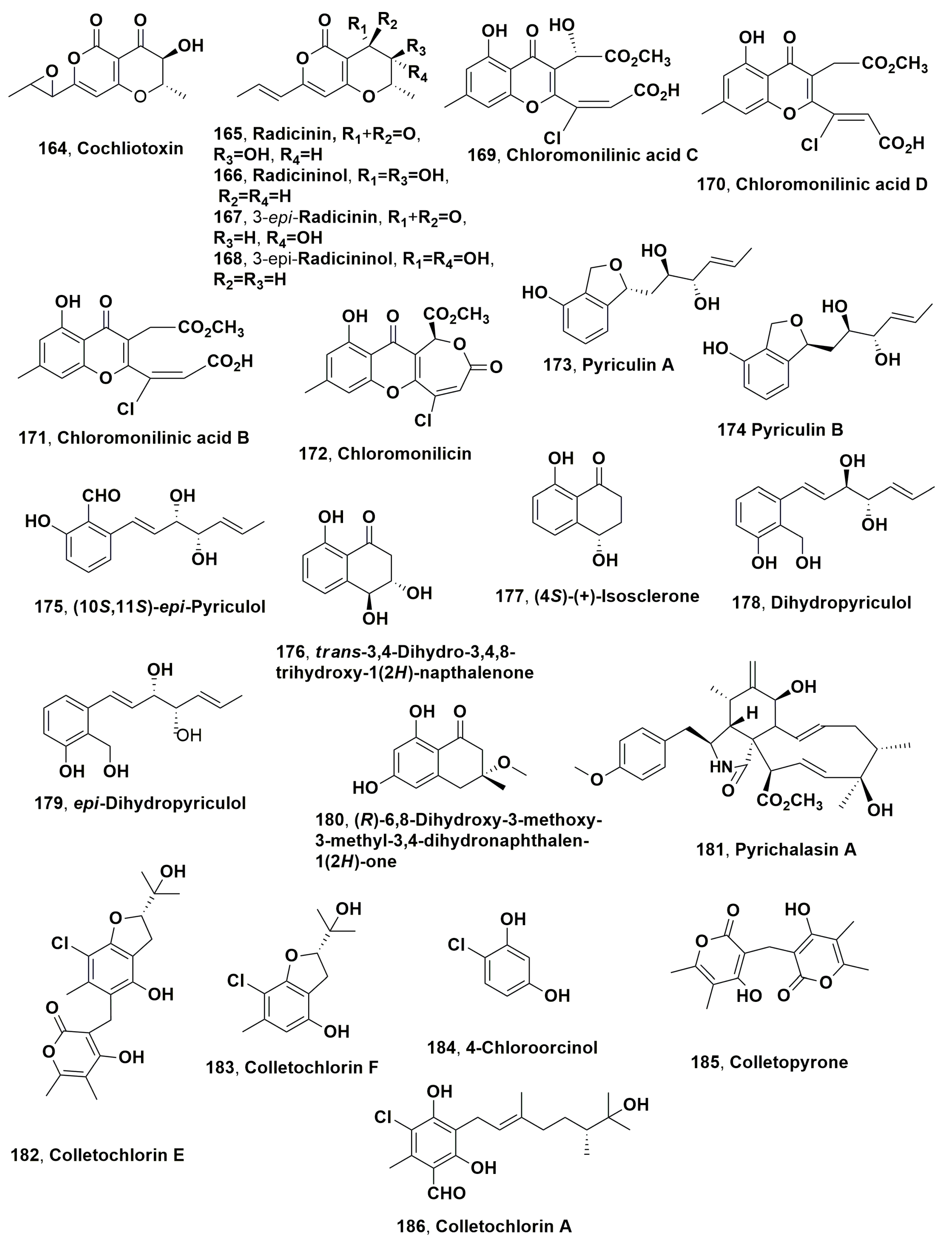
| Cochliotoxin (164) | buffelgrass (Pennisetum ciliare or Cenchrus ciliaris) | Cochliobolus australiensis | Phytotoxic activity on the host plant and other two native weeds | [208] |
| Radicinin (165) | “ | “ | “ | “ |
| Radicinol (166) | “ | “ | Not toxic | “ |
| 3-epi-Radicinin (167) | “ | “ | Phytotoxic activity on the host plant and the other two native weeds | “ |
| 3-epi-Radicinol (168) | “ | “ | Not toxic | “ |
| Chloromonilinic acid C (169) | “ | “ | Phytotoxicity on the host plant | [209] |
| Chloromonilinic acid D (170) | “ | “ | “ | “ |
| Chloromonilinic acid B (171) | “ | “ | “ | “ |
| Chloromonilicin (172) | Sowthistle (Socnhus arvensis) | A. sonchi | No toxicity | [1] |
| Peryculin A (173) | buffelgrass (Pennisetum ciliare or Cenchrus ciliaris) | Perycularia grisea | Phytotoxicity on the host weed | [212] |
| Peryculin B (174) | “ | “ | “ | “ |
| (10S,11S)-(−)-epi-Pyriculol (175) | “ | “ | “ | “ |
| trans-3,4-Dihydro-3,4,8-trihydroxy-1(2H)-napthalenon (176) | “ | “ | “ | “ |
| 4S)-(+)-Isosclerone (177) | “ | “ | “ | “ |
| Dihydropyriculol (178) | “ | “ | No toxicity | [213] |
| epi-Dihydropyriculol (179) | “ | “ | “ | “ |
| 3-Methoxy-6,8-dihydroxy-3-methyl-3,4-dihydroisocoumarin (180) | “ | “ | “ | “ |
| Pyrichalasin H (181) | Signal grass (Brachiaria eruciformis) | “ | Phytotoxicity on weeds and cultivated plants | [214] |
| Colletochlorin E (182) | Brassica sp. | Colletetotrichum higginsianum | Phytotoxicity on weeds, parasitic and cultivated plants | [217] |
| Colletochlorin F (183) | “ | “ | “ | “ |
| 4-Chlororcinol (184) | “ | “ | “ | “ |
| Colletopryrone (185) | “ | “ | “ | “ |
| Colletochlorin A (186) | Ragweed (Ambrosia artemisiifolia) | “ Colletotrichum gloeosporioides | “ Phytotoxicity on the host weed | “ [218] |
| Colletopyrandione (187) | Rape (Brassica sp.) | C. higginsianum | Phytotoxicity on S. arvensis and Helianthus annuus | [218] |
| Colletochlorin G (188) | “ | “ | Not tested | “ |
| Colletochlorin H (189) | “ | “ | “ | “ |
| Orcinol (190) | Ragweed (A. artemisiifolia) | C. gloeosporioides | Phytotoxicity on the host weed | [219] |
| Dirhamnolipid (191) | Bawanghua (Hylocereus undatus) | “ | Phytoxicity against different weeds | [221] |
| Curvulin (192) | Saffron thisle (C. arvense) | Paraphoma sp. | No toxicity | [223] |
| Phaeosphaeride (193) | “ | “ | “ | “ |
| 9-O-Methylfusarubin (194) | Cheatgrass (Bromus tectorum) | Rutstroemia capillus-albis | Phytotoxicity on the host plant | [224] |
| 9-O-Methylbostrycoidin (195) | “ | “ | Weak phytotoxicity on the host plant | “ |
| 5-O-Methylnectriafurone (196) | “ | “ | “ | “ |
| trans-Methyl-p-coumarate (197) | “ | “ | “ | “ |
| Terpestacin (198) | “ | “ | “ | “ |
| 8-Hydroxy-3-methyl-4-chloro-9-oxo-9H-xanthene-1-carboxylate (199) | Canada Thisle (S. arvensis) | Alternaria sonchi | Phytotoxicity on S. arvensis and E. repens | [225] |
| Chloromoniliphenone (200) | “ | “ | “ | “ |
| Araufuranone (201) | White blade flower (Araujia hortorum) | Ascochyta araujiae | Phytotoxicity on different weeds | [226] |
| Neovasinin and (202) | “ | “ | “ | “ |
| 2,4-Dihydroxy-6-hydoxy Methylbenzaldehyde (203) | “ | “ | “ | “ |
| Phytotoxin | Weed | Fungus | Biological Activity | References |
|---|---|---|---|---|
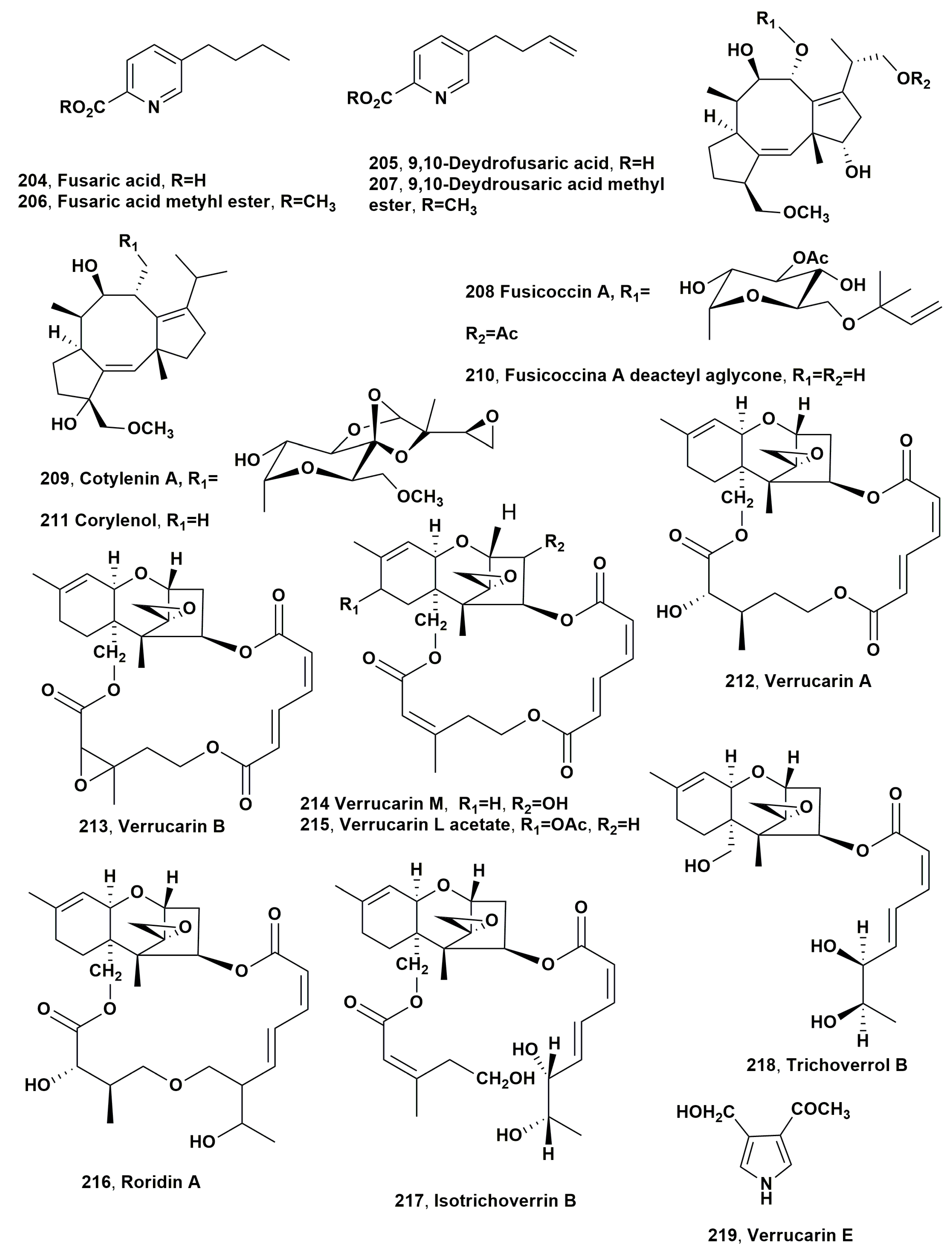
| Fusaric acid (204) | Purple witchweed (Striga hermonthica) Broomrape (Orobanche cumana) | Fusarium nygamai Fusarium verticilloides | Phytotoxicity on tomato and inhibition of Striga hermonthica seeds germination Phytotoxicity against P. aegyptiaca, O. ramosa and O. cumana | [231] [232] |
| 9,10-Deydrofusaric acid (205) | “ | “ | “ | “ |
| Fusaric acid methyl ester (206) | “ | “ | “ | “ |
| 9,10-Deydrofusaric methyl ester (207) | “ | “ | “ | “ |
| Fusicocin (208) | No infected weed | Phomopsis amygdaly | Stimulation of seeds germination of S. hermonthica and O. minor | [237] |
| Cotylenin A (209) | “ | Cladosporium sp. 501-7W | “ | “ |
| Fusicoccin deacetyl aglycone (210) | “ | P. amygdaly | “ | “ |
| Cotylenol (211) | “ | Cladosporium sp. 501-7W | “ | “ |
| Verrucarin A (212) | Broomrape (Pelipanche ramosa) | Myrothecium verrucaria | Inhibition of P. ramosa seed germination | [242] |
| Verrucarin B (213) | “ | “ | “ | “ |
| Verrucarin M (214) | “ | “ | “ | “ |
| Verrucarin L acetate (215) | “ | “ | “ | “ |
| Roridin A (216) | “ | “ | “ | “ |
| Isotrichoverrin B (217) | “ | “ | “ | “ |
| Trichoverrol B (218) | “ | “ | “ | “ |
| Verrucarin E (219) | “ | “ | Not toxic | “ |
| Neosolanial monoactate (220) | “ | Fusarium compactum | Inhibition of P. ramosa seed germination | “ |
| Cyclopaldic acid (221) | No infected weed | Diplodia cupressi | Phytotoxicity on O. crenata, O. cumana, O. minor, and P. ramosa | [243] |
| Sphaeropsidin A (222) | “ | “ | “ | “ |
| Sphaeropsidone (223) | “ | “ | “ | “ |
| epi-Spheropsidone (224) | “ | “ | “ | “ |
| epi-Epoformin (225) | “ | “ | “ | “ |
| Pinolide (226) | “ | D. pinodes | No toxicity | [150] |
| Cavoxin (227) | “ | Phoma cava | No toxicity | [244] |
| Cavoxone (228) | “ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evidente, A. Specialized Metabolites Produced by Phytotopatogen Fungi to Control Weeds and Parasite Plants. Microorganisms 2023, 11, 843. https://doi.org/10.3390/microorganisms11040843
Evidente A. Specialized Metabolites Produced by Phytotopatogen Fungi to Control Weeds and Parasite Plants. Microorganisms. 2023; 11(4):843. https://doi.org/10.3390/microorganisms11040843
Chicago/Turabian StyleEvidente, Antonio. 2023. "Specialized Metabolites Produced by Phytotopatogen Fungi to Control Weeds and Parasite Plants" Microorganisms 11, no. 4: 843. https://doi.org/10.3390/microorganisms11040843
APA StyleEvidente, A. (2023). Specialized Metabolites Produced by Phytotopatogen Fungi to Control Weeds and Parasite Plants. Microorganisms, 11(4), 843. https://doi.org/10.3390/microorganisms11040843