
Focus and Insights into the Synthetic Biology-Mediated Chassis of Economically Important Fungi for the Production of High-Value Metabolites



Abstract
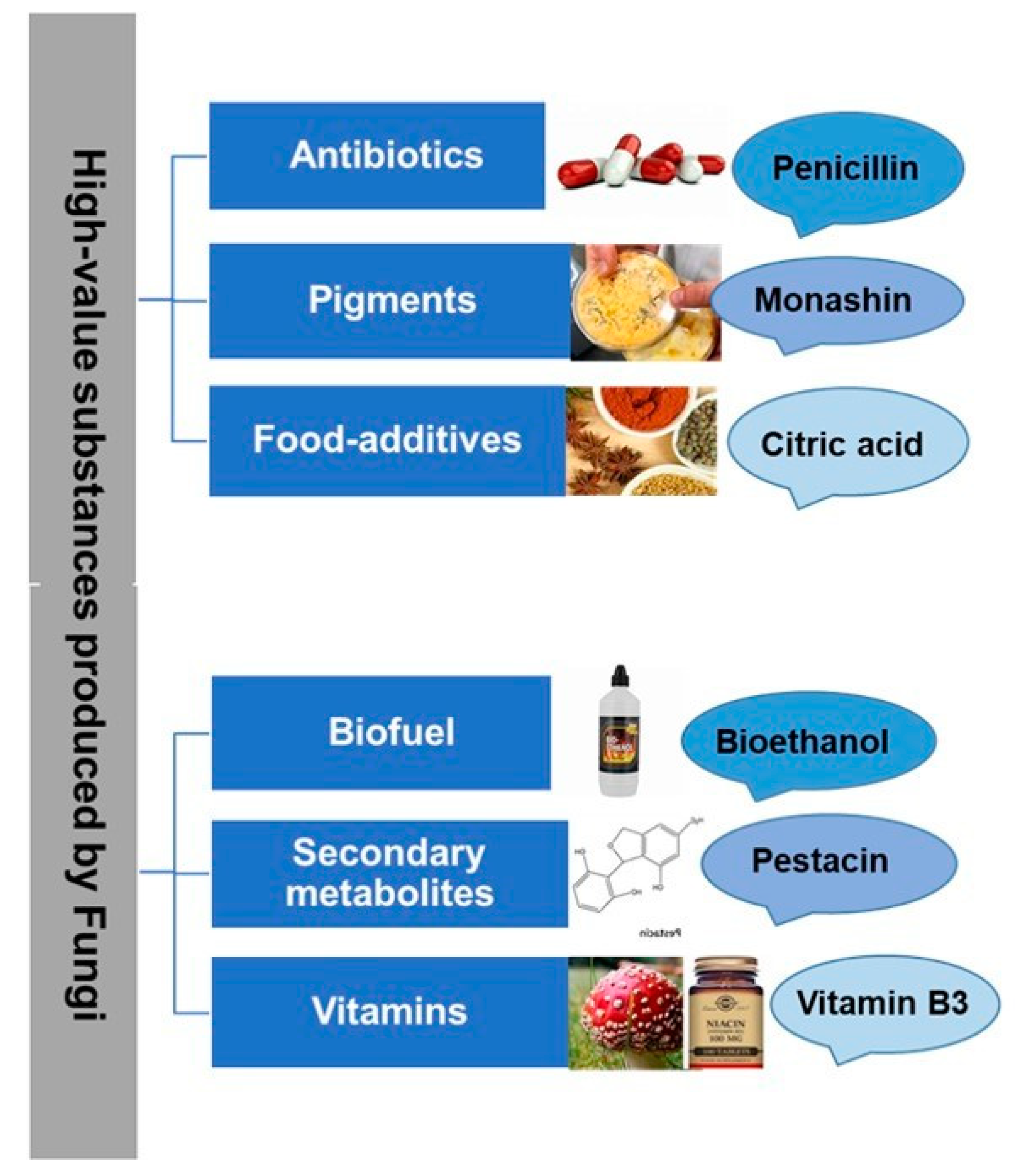
:1. Introduction
2. Nanotechnology and Fungal Biotechnology

{kind=link}
{kind=link}
| Nanoparticle | Fungal Species | Socio-Economic Application | Reference |
|---|---|---|---|
| Biomedical and therapeutic applications | |||
| AgNP | Trichoderma viride | Combination with antibiotics produces a synergistic effect | [61] |
| AuNP | Helmithosporum solani | Anticancer drug | [62] |
| AgNP | Aspergillus fumigatus | Antiviral against HIV-1 | [63] |
| TiO2 | Aspergillus flavus | Antimicrobial function | [64] |
| AuNP | Candida albicans | Liver cancer detection | [65] |
| Agricultural applications | |||
| AuNP | Rhizopus oryzae | Pesticides | [66] |
| Ca3P2O8 NP | Aspergillus tubingensis | Agriculture | [67] |
| AuNP | Fusarium semitectum | Optoelectronics | [68] |
| FeCl3 | Aspergillus oryzae | Agriculture | [69] |
| AgNP | Rhizopus stolonifer | ----- | [70] |
| TiO2 | Aspergillus flavus TFR7 | Plant nutrient | [71] |
3. Fungi—An Emerging Biological Platform for Metabolic Engineering
| Fungal Strain | High-Value Compounds | Strategy for Metabolic Engineering | Research Outcome | Reference |
|---|---|---|---|---|
| S. cerevisiae | Naringenin (Flavanones) | Genetic manipulation for SmCHS2 expression in a heterologous system | Increased production of naringenin (648.63 mg/L) | [83] |
| S. cerevisiae | Rubusoside and Rebaudiosides | Construction of de novo Rubusoside biosynthetic pathway, removal of rate-limiting steps, metabolic model-based prediction of engineerable targets | Improved production of rubusoside (1368.6 mg/L) and rebaudioside (132.7 mg/L) | [84] |
| S. cerevisiae | Lycopene (carotenoid) | Genome engineering for lycopene pathway optimization and increase in acetyl-CoA pool | Enhanced production of lycopene (56 mg/g) | [85] |
| S. cerevisiae | Eriodictyol (Flavanones) | A cytochrome P450 F3′H from Gerbera hybrida was functionally expressed in S. cerevisiae | Increased production of eriodictyol (200 mg/L) | [86] |
| S. cerevisiae | Fatty acid ethyl esters (FAEEs) | Genetic manipulation of S. cerevisiae to utilize glycerol as a substrate for ethanol production | Higher yields of fatty acid ethyl esters | [87] |
| S. cerevisiae | Ethanol | T. reesei endoglucanase EGLI and Saccharomycopsis fibuligera (β-glucosidase) Bgl1 was introduced into S. cerevisiae | Enhanced ethanol production | [88] |
| S. cerevisiae | Fatty acid-derived n-butanol (biofuel) | The silencing of ADH1, ADH4, GPD1, and GPD2 genes in fungal strain | Increased production of n-butanol (100 mg/L) | [78] |
| S. cerevisiae | Citrate | Deletion of IDH1 and IDH2 genes via marker-based homologous recombination | Increased production of citrate | [89] |
| S. cerevisiae | Free fatty acids (FFA) | Pathway reconstitution and optimization (synthetic citrate lyase pathway), Heterologous expression of ATP citrate lyase, and malic enzyme | Increased free fatty acid production (10.4 g/L) | [90] |
| S. cerevisiae | Terpenes | Mevalonic acid (MVA) pathway engineering in S. cerevisiae | Enhanced terpene production | [91] |
| S. cerevisiae | Cis, cis-Muconic acid | Pathway engineering (amino acid synthesis) in yeast and conversion of 3-dehydroshikimate (DHS) into cis, cis-muconic acid | Screening for best heterologous genes catalyzing the conversion of DHS to cis–cis muconic acid | [92] |
| S. cerevisiae WRY2 | Fatty acids | ATP citrate lyase was introduced and malate synthase was downregulated in the engineered strain | Increased fatty acid production (460 mg/L) | [93] |
| S. cerevisiae | Rosmarinic acid (hydroxycinnamic acid ester) | Metabolic engineering of S. cerevisiae | High-level production of rosmarinic acid (5.93 mg/L) | [94] |
| S. cerevisiae | p-hydroxybenzoic acid (PHBA) | Fungal engineering for gene deletion (for negative feedback), overexpression of chorismite lyase (from E. coli) | Enhanced production of aromatic compounds | [95] |
| S. cerevisiae | (+)-Valencene and (+)-nootkatone (Sesquiterpenoids) | Combinational engineering comprising promoter change, regulator ROX1 knockout, squalene pathway inhibition, and HMGR overexpression | Enhanced production of β-nootkatol and (+)-nootkatone (170.5 and 45.6 mg/mL) | [96] |
| S. cerevisiae | Fatty acids | Overexpression of ACC1, FAS1, and FAS2 genes in the fungal strain | Enhanced fatty acid production | [97] |
| S. cerevisiae | Chlorogenic acid (phenolic compound) | An optimized de novo biosynthetic pathway for CGA was reconstructed in S. cerevisiae, a multi-module engineering strategy | Increased production of chlorogenic acid (806.8 mg/L) | [98] |
| S. cerevisiae | Naringenin (Flavanones) | RgTAL (encoding tyrosine ammonia lyase) from Rhodotorula glutinosa, Pc4CL (encoding 4-coumaric acid-CoA ligase) from Petroselinum crispum, PhCHS from P. hybrida, and MsCHI from Medicago sativa in S. cerevisiae | Increased production of naringenin (29 mg/L) | [99] |
| A. niger, S. cerevisiae | Cheese | Genetic modification with calf rennet gene (chymosin) | Improvement of cheese products | [100] |
| Y. lipolytica | Taxifolin (flavanonols) | Taxifolin biosynthetic pathway expression in Y. lipolytica | Increased production of taxifolin (48.1 mg/L) | [101] |
| Kluyveromyces lactis | L-ascorbic acid (vitamin C) | Genetic transformation of K. lactis with a plasmid harboring cloned plant genes | Increased production of L-ascorbic acid (30 mg/L) | [102] |
| K. lactis | Bioethanol | Construction and characterization of a null mutant (Δklndi1) in the single gene encoding a mitochondrial alternative internal dehydrogenase | Increased bioethanol production | [103] |
| K. lactis | B-galactosidase | A rational mutagenesis strategy by introducing disulfide bonds in the interface between the enzyme subunits was used | Improvement of β-galactosidase enzyme for high-temperature industrial applications | [104] |
| A. niger | Glucoamylase | glaA gene encoding for glucoamylase was expressed in A. niger under the control of the tunable Tet-on system, deletion of the racA gene in the engineered strain | Enhanced glucoamylase secretion in the engineered strain | [105] |
| A. niger | glucoamylase–glucuronidase (GlaGus) protein | Disruption of ER-associated degradation (ERAD) components, derA, doaA, hrdC, mifA, or mnsA in A. niger | High-viable A. niger strain, enhanced production of (GlaGus) protein | [106] |
| A. niger | Manganese peroxidase | Heterologous expression of manganese peroxidase (from Phanerochaete chrysosporium) in A. niger, overexpression of calnexin (chaperone) | Increased production of manganese peroxidase | [107] |
| A. niger | Human α1-proteinase inhibitor (α1-PI) (antitrypsin) | Heterologous expression of Human α1-proteinase inhibitor in A. niger, a fusion of α1-PI with glucoamylase G2, transformants screening | biologically active glycosylated r-α1-PI with yields of up to 12 mg/L | [108] |
| A. niger | lignin peroxidase | Heterologous expression of lignin peroxidase (from Phanerochaete chrysosporium) in A. niger under NOS promoter and terminator | Lignin peroxidase production | [109] |
| A. niger | Secondary metabolites (enniatins) | Heterologous expression of nonribosomal peptide synthetase ESYN (from F. oxysporum) under Tet-on hybrid promoter in A. niger | Enhanced production of enniatins | [110] |
| A. oryzae | Cellulase activity | Integration of multiple copies of genes encoding cellulases (cellobiohydrolase, endoglucanase, and β-glucosidase) into the fungal genome | Enhanced activity of cellulases | [111] |
| A. niger | Citric acid | The system consists of two expression modules jointly targeted to a defined genomic locus | Expression of the desired gene and its function | [80] |
| A. niger | Galactaric acid (organic acid) | Single- and multiplexed CRISPR approaches for galactarate overproduction | Higher production of galactarate (12 g/L) | [81] |
| A. niger | Vitamin C | Metabolic engineering of the fungal d-galacturonate pathway | A higher titer of vitamin C (170 mg/L) | [112] |
| A. niger | Human granulocyte colony-stimulating factor (G-CSF) | Fusion of G-CSF behind a KEX2 cleavage site downstream of glucoamylase | High yields of protein G-CSF in A. niger (5–10 mg/L culture medium) | [113] |
| P. pastoris | (+)-nootkatone | Chimeric strain co-expressing the premnaspirodiene oxygenase Hyoscyamus muticus and A. thaliana cytochrome P450 reductase. Intracellular production of (+)-valencene by co-expression of valencene synthase | Enhanced production of (+)-nootkatone (208 mg/L) | [114] |
| P. pastoris | Fatty acid alcohols | Metabolic transformation of the fatty acid cell factory | High-level production of fatty acid derivatives (2.0 g/L) | [115] |
| P. pastoris | Malic acid | Metabolic engineering and redistribution of metabolic flux | High-level production of malic acid (2.79 g/L) | [116] |
| Yarrowia lipolytica | β-Carotene (carotenoid) | Optimization of promoter–gene pairs of heterologous crt pathway | High yields of β-carotene (6.5 g/L) | [117] |
| Trichoderma reesei | Cellulase | Deletion of the small GTPase rac1 in T. reesei | Hyperbranching in T. reesei strain enhanced cellulase production | [118] |
| Neurospora crassa | Cellulase | Disruption of gul-1 decreased culture viscosity gul-1 overexpression increased viscosity | Increased cellulase secretion in the engineered strain | [119] |
| Y. lipolytica | Omega-3 eicosapentaenoic acid | Heterologous expression of Δ-9 elongase, a Δ-8 desaturase, a Δ-5 desaturase and a Δ-17 desaturase, transformation in Y. lipolytica | High yields of eicosapentaenoic (EPA) acid | [82] |
| Mortierella alpina | Oleic and Linoleic acids | Heterologous expression of the D12-desaturase (from Coprinopsis cinerea) in the D6DS activity-defective mutant of M. alpina | Enhanced production of oleic and linoleic acids in the engineered strain | [120] |
| Xanthophyllomyces dendrorhous | Zeaxanthin | Overexpression of β-carotene hydrolase and mutagenesis of astaxanthin synthase | Enhanced production of zeaxanthin (0.5 mg/g) | [121] |
| T. reesei | Cellulase | Overexpression of β-glucosidase in T. reesei under a strong inducible promoter | Enhanced production of cellulase | [122] |
| T. koningii | Cellulase | RNA interference was used to regulate the expression of the cre1 gene | Enhanced production of cellulase | [123] |
| Blakeslea trispora | Lycopene (carotenoid) | Fermentation optimization with lycopene cyclase inhibitor | Enhanced production of lycopene (256 mg/L) | [22] |
| Filamentous fungi | Austinoids (insecticides) | Combinational engineering and rewiring of austinoid pathway | Production of diverse austinoid derivatives | [124] |
4. Fungal Chassis and Production of Functional Foods
4.1. Metabolic Engineering toward Microbial Strain Improvement
4.2. Mutagenesis Approaches in Fungi
4.3. Pathway Engineering in Fungi-Recent Trends and Initiatives
4.4. Precursor Supply Increase
4.5. Downregulation/Deletion of Metabolic Pathways
4.6. Metabolic Pathway Engineering
4.7. CRISPR/Cas Genome Editing in Fungi
4.8. Key Metabolic Engineering Studies in Fungi
5. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Piffieri, F.; Aujard, F. Caloric restriction, longevity, and aging: Recent contributions from human and non-human primate studies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 95, 109702. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; He, S.; Cao, X.; Ye, Y.; Yang, L.; Wang, J.; Liu, H.; Sun, H. Potential prebiotic activities of soybean peptides Maillard reaction products on modulating gut microbiota to alleviate aging-related disorders in D-galactose-induced ICR mice. J. Funct. Foods 2020, 65, 103729. [Google Scholar] [CrossRef]
- Tiwari, P.; Bae, H. Endophytic fungi: Key insights, emerging prospects, and challenges in natural product drug discovery. Microorganisms 2022, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Seogchan, K.; Bae, H. Plant-endophyte associations: Rich yet under-explored sources of novel bioactive molecules and applications. Microbiol. Res. 2023, 266, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P. Sustainable agriculture and nanotechnologies for food and nutraceutical production—An update. In Plant and Nanoparticles; Chen, J., Ed.; Springer Book Series: Berlin/Heidelberg, Germany, 2022; ISBN 978-981-19-2502-3. [Google Scholar]
- Karwehl, S.; Stadler, M. Exploitation of fungal biodiversity for the discovery of novel antibiotics. Curr. Top. Microbiol. Immunol. 2017, 398, 303–338. [Google Scholar]
- Tiwari, P.; Srivastava, Y.; Bajpai, M.; Sharma, A. Bioactive metabolites from natural sources: Prospects and significance in drug discovery and research. Bioingene PSJ 2021, 1, 1–14. [Google Scholar]
- Baggio, L.M.; Panagio, L.A.; Gasparin, F.G.M.; Sartori, D.; Celligoi, M.A.P.C.; Baldo, C. Production of fibrinogenolytic and fibrinolytic enzymes by a strain of Penicillium sp. isolated from contaminated soil with industrial eluent. Acta Sci. Health Sci. 2019, 41, 40606. [Google Scholar] [CrossRef]
- Machida, M.; Yamada, O.; Gomi, K. Genomics of Aspergillus oryzae: Learning from the history of Koji mold and exploration of its future. DNA Res. 2008, 15, 173–183. [Google Scholar] [CrossRef]
- Skellam, E. Strategies for engineering natural product biosynthesis in Fungi. Trends Biotechnol 2019, 37, 416–427. [Google Scholar] [CrossRef]
- Takahashi, J.A.; Barbosa, B.V.R.; Martins, B.de.A.; Guirlanda, C.P.; Moura, M.A.F. Use of the versatility of fungal metabolism to meet modern demands for healthy aging, functional foods, and sustainability. J. Fungi 2020, 6, 223. [Google Scholar] [CrossRef]
- Bills, G.F.; Gloer, J.B. Biologically active secondary metabolites from the fungi. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, E.J.; Delange, J. Fungal protein—What is it and what is the health evidence? A systematic review focusing on mycoprotein. Front. Sustain. Food Syst. 2021, 5, 581682. [Google Scholar] [CrossRef]
- Hashempour-Baltork, F.; Khosravi-Darani, K.; Hosseini, H.; Farshi, P.; Reihani, S.F.S. Mycoproteins as safe meat substitutes. J. Clean. Prod. 2020, 253, 119958. [Google Scholar] [CrossRef]
- Kewuyemi, Y.O.; Kesa, H.; Adebo, O.A. Trends in functional food development with three-dimensional (3D) food printing technology: Prospects for value-added traditionally processed food products. Crit. Rev. Food Sci. Nutr. 2022, 62, 7866–7904. [Google Scholar] [CrossRef] [PubMed]
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef]
- Browne, A.G.P.; Fisher, M.C.; Henk, D.A. Species-specific PCR to describe local-scale distributions of four cryptic species in the Penicillium chrysogenum complex. Fungal Ecol. 2013, 6, 419–429. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, J.; Huang, Y.; Xin, Q.; Wang, Z. Diversifying of the chemical structure of native Monascus pigments. Front. Microbiol. 2018, 9, 3143. [Google Scholar] [CrossRef]
- Shi, T.-Q.; Liu, G.N.; Ji, R.Y.; Shi, K.; Song, P.; Ren, L.J.; Huang, H.; Ji, X.J. CRISPR/Cas9-based genome editing of the filamentous fungi: The state of the art. Appl. Microbiol. Biotechnol. 2017, 101, 7435–7443. [Google Scholar] [CrossRef]
- Dutta, B.; Lahiri, D.; Nag, M.; Ghosh, S.; Dey, A.; Ray, R.R. Fungi in pharmaceuticals and production of antibiotics. In Applied Mycology. Fungal Biology; Shukla, A.C., Ed.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Meyer, V. Metabolic engineering of Filamentous fungi. In Metabolic Engineering: Concepts and Applications, 1st ed.; Lee, S.Y., Nielsen, J., Stephanopoulos, S., Eds.; WILEY-VCH GmbH: Weinheim, Germany, 2021. [Google Scholar]
- Mantzouridou, F.T.; Naziri, E. Scale translation from shaken to diffused bubble aerated systems for lycopene production by Blakeslea trispora under stimulated conditions. Appl. Microbiol. Biotechnol. 2017, 101, 1845–1856. [Google Scholar] [CrossRef]
- Saha, B.C.; Kennedy, G.J. Efficient itaconic acid production by Aspergillus terreus: Overcoming the strong inhibitory effect of manganese. Biotechnol. Prog. 2020, 36, e2939. [Google Scholar] [CrossRef]
- Patil, R.H.; Krishnan, P.; Maheshwari, V.L. Production of lovastatin by wild strains of Aspergillus terreus. Nat. Prod. Commun. 2011, 6, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Men, P.; Tang, S.; Lu, X. Aspergillus terreus as an industrial filamentous fungus for pharmaceutical biotechnology. Curr. Opin. Biotechnol. 2021, 69, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, Z.; Liu, W.; Ke, X.; Tian, X.; Chu, J. Cephalosporin C biosynthesis and fermentation in Acremonium chrysogenum. Appl. Microbiol. Biotechnol. 2022, 106, 6413–6426. [Google Scholar] [CrossRef]
- Kato, T.; Azegami, J.; Yokomori, A.; Dohra, H.; Enshasy, H.A.; Park, E.Y. Genomic analysis of a riboflavin-overproducing Ashbya gossypii mutant isolated by disparity mutagenesis. BMC Genom. 2020, 21, 319. [Google Scholar] [CrossRef] [PubMed]
- Jesus, L.F.M.C. Produção de-Galactosidase Por Fungos Filamentosos: Screening, Purificação e Caracterização Bioquímica. Master’s Thesis, Universidade Estadual Paulista “Julio Mesquita Filho”, UNESP, São Paulo, Brazil, 2020. [Google Scholar]
- Terabayashi, Y.; Sano, M.; Yamane, N.; Marui, J.; Tamano, K.; Sagara, J.; Dohmoto, M.; Oda, K.; Ohshima, E.; Tachibana, K.; et al. Identification and characterization of genes responsible for the biosynthesis of kojic acid, an industrially important compound from Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 953–961. [Google Scholar] [CrossRef]
- Huang, Z.; Brennan, C.S.; Zheng, H.; Mohan, M.S.; Stipkovits, L.; Liu, W.; Kulasiri, D.; Guan, W.; Zhao, H.; Liu, J. The effects of fungal lipase-treated milk lipids on bread making. LWT 2020, 128, 109455. [Google Scholar] [CrossRef]
- Hjortmo, S.B.; Hellström, A.M.; Andlid, T.A. Production of folates by yeasts in Tanzanian fermented togwa. FEMS Yeast Res. 2008, 8, 781–787. [Google Scholar] [CrossRef]
- Mamani, L.D.G.; Magalhães, A.I., Jr.; Ruan, Z.; de Carvalho, J.C.; Soccol, C.R. Industrial production, patent landscape, and market trends of arachidonic acid-rich oil of Mortierella alpina. Biotech. Res. Innov. 2019, 3, 103–119. [Google Scholar] [CrossRef]
- Anand, S.; Singh, K.S.; Aggarwal, D. Expanding avenues for probiotic yeast. In Microbial Cell Factories, 1st ed.; Sharma, D., Saharan, B.S., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 125–141. [Google Scholar]
- Mallikarjuna, N.; Yellamma, K. Genetic and metabolic engineering of microorganisms for the production of various food products. Recent Dev. Appl. Microbiol. Biochem. 2019, 60, 167–182. [Google Scholar]
- Tiwari, P.; Khare, T.; Shriram, V.; Bae, H.; Kumar, V. Exploring synthetic biology strategies for producing potent antimicrobial phytochemicals. Biotechnol. Adv. 2021, 48, 107729. [Google Scholar] [CrossRef]
- Tiwari, P.; Srivastava, Y.; Bae, H. Trends of pharmaceutical design of Endophytes as anti-infective. Curr. Top. Med. Chem. 2021, 21, 1572–1586. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P. Endophytes: Types, Potential Uses and Mechanisms of Action; Nova publishers: Hauppauge, NY, USA, 2022; ISBN 979-8-88697-205-4. [Google Scholar]
- U.S. Department of Agriculture and U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed. 2020. Available online: DietaryGuidelines.gov (accessed on 26 August 2021).
- Vega Oliveros, C. Comparación de la Producción de Metabolitos Secundarios Bioactivos con dos Fuentes de Carbono en la Fermentación Líquida de Una Especie de Pleurotus y su uso Potencial en un Alimento de Tipo Funcional (Thesis). Facultad de Ciencias Agragrias. Posgrado en Ciencia y Tecnología de Alimentos. Ph.D. Thesis, Universidad Nacional de Colombia, Bogotá, DC, Colombia, 2016. [Google Scholar]
- Souza Filho, P.F.; Andersson, D.; Ferreira, J.A.; Taherzadeh, M.J. Mycoprotein: Environmental impact and health aspects. World J. Microbiol. Biotechnol. 2019, 35, 147. [Google Scholar] [CrossRef] [PubMed]
- Barzee, T.J.; Cao, L.; Pan, Z.; Zhang, R. Fungi for future foods. J. Future Foods 2021, 1, 25–37. [Google Scholar] [CrossRef]
- Ojwach, J.; Adetunji, A.I.; Mutanda, T.; Mukaratirwa, S. Oligosaccharides production from coprophilous fungi: An emerging functional food with potential health-promoting properties. Biotechnol. Rep. 2022, 33, e00702. [Google Scholar] [CrossRef]
- Fu, L.; Wang, Y.; Wang, J.; Yang, Y.; Hao, L. Evaluation of the antioxidant activity of extracellular polysaccharides from Morchella esculenta. Food Funct. 2013, 4, 871–879. [Google Scholar] [CrossRef]
- Jeong, S.C.; Koyyalamudi, S.R.; Jeong, Y.T.; Song, C.H.; Pang, G. Macrophage immunomodulating and antitumor activities of polysaccharides isolated from Agaricus bisporus white button mushrooms. J. Med. Food 2012, 15, 58–65. [Google Scholar] [CrossRef]
- Wen, L.; Gao, Q.; Ma, C.W.; Ge, Y.; You, L.; Liu, R.H.; Fu, X.; Liu, D. Effect of polysaccharides from Tremella fuciformis on UV-induced photoaging. J. Func. Foods. 2016, 20, 400–441. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Zhang, T.H. Immunomodulatory activities of polysaccharides from Ganoderma on immune effector cells. Food Chem. 2020, 340, 127933. [Google Scholar] [CrossRef]
- GNC Ganoderma Capsules. Available online: http://www.vitagou.com/gnc/161.html (accessed on 23 February 2023).
- Drugs.com. Lentinan. Available online: https://www.drugs.com/npp/lentinan.html (accessed on 23 February 2023).
- Hanqi Edible Mushroom Products. Available online: https://www.meipian.cn/1ez6jqn1 (accessed on 23 February 2023).
- Wang, L.; Li, C.; Ren, L.; Guo, H.; Li, Y. Production of pork sausages using Pleaurotus eryngii with different treatments as replacements for pork back fat. J. Food Sci. 2019, 84, 3091–3098. [Google Scholar] [CrossRef]
- Jo, K.; Lee, S.; Jo, C.; Jeon, H.J.; Choe, J.H.; Choi, Y.S.; Jung, S. Utility of winter mushroom treated by atmospheric non-thermal plasma as an alternative for synthetic nitrite and phosphate in ground ham. Meat Sci. 2020, 166, 108151. [Google Scholar] [CrossRef]
- Lu, X.; Brennan, M.A.; Serventi, L.; Liu, J.; Guan, W.; Brennan, C.S. Addition of mushroom powder to pasta enhances the antioxidant content and modulates the predictive glycaemic response of pasta. Food Chem. 2018, 264, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Steudler, S.; Werner, A.; Walther, T. It is the Mix that matters: Substrate-specific enzyme production from filamentous fungi and bacteria through solid-state fermentation. Adv. Biochem. Eng. Biotechnol. 2019, 169, 51–81. [Google Scholar]
- Hussain, H.; Nazir, M.; Saleem, M.; Al-Harrasi, A.; Green, I.R. Fruitful decade of fungal metabolites as anti-diabetic agents from 2010 to 2019: Emphasis on α-glucosidase inhibitors. Phytochem. Rev. 2021, 20, 145–179. [Google Scholar] [CrossRef]
- Meyer, V.; Fiedler, M.; Nitsche, B.; King, R. The cell factory Aspergillus enters the big data era: Opportunities and challenges for optimizing product formation. Adv. Biochem. Eng. Biotechnol. 2015, 149, 91–132. [Google Scholar]
- Abe, K.; Gomi, K.; Hasegawa, F.; Machida, M. Impact of Aspergillus oryzae genomics on industrial production of metabolites. Mycopathologia 2006, 162, 143–153. [Google Scholar] [CrossRef]
- Jin, F.J.; Hu, S.; Wang, B.T.; Jin, L. Advances in genetic engineering technology and its application in the industrial fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 644404. [Google Scholar] [CrossRef]
- Jiang, C.; Lv, G.; Tu, Y.; Cheng, X.; Duan, Y.; Zeng, B.; He, B. Applications of CRISPR/Cas9 in the synthesis of secondary metabolites in filamentous fungi. Front. Microbiol. 2021, 12, 638096. [Google Scholar] [CrossRef]
- Srivastava, A.K. The role of fungus in bioactive compound production and nanotechnology. In Role of Plant Growth Promoting Microorganisms in Sustainable Agriculture and Nanotechnology; Woodhead Publishing, Elsevier: Cambridge, UK, 2019; pp. 145–162. [Google Scholar]
- Adebayo, E.A.; Azeez, M.A.; Alao, M.B.; Oke, A.M.; Aina, D.A. Fungi as a veritable tool in current advances in nanobiotechnology. Heliyon 2021, 7, 2405–8440. [Google Scholar] [CrossRef] [PubMed]
- Fayaz, A.M.; Balaji, K.; Girilal, M.; Yadav, R.; Kalaichelvan, P.T.; Venketesan, R. Biogenic synthesis of silver nanoparticles and their synergistic effect with antibiotics: A study against gram-positive and gram-negative bacteria. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 103–109. [Google Scholar] [CrossRef]
- Kumar, S.A.; Peter, Y.A.; Nadeau, J.L. Facile biosynthesis, separation, and conjugation of gold nanoparticles to doxorubicin. Nanotechnology 2008, 19, 495101. [Google Scholar] [CrossRef] [PubMed]
- Alani, F.; Moo-Young, M.; Anderson, W. Biosynthesis of silver nanoparticles by a new strain of Streptomyces sp. compared with Aspergillus fumigatus. World J. Microbiol. Biotechnol. 2012, 28, 1081–1086. [Google Scholar] [CrossRef]
- Rajakumar, G.; Rahuman, A.; Roopan, S.M.; Khanna, V.G.; Elango, G.; Kamaraj, C.; Zahir, A.A.; Velayutham, K. Fungus-mediated biosynthesis and characterization of TiO2 nanoparticles and their activity against pathogenic bacteria. Spectrochim. Acta Mol. Biomol. Spectrosc. 2012, 91, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Chuhan, A.; Zubair, S.; Tufail, S.; Sherwani, A.; Sajid, M.; Raman, S.C.; Azam, A.; Owais, M. Fungus-mediated biological synthesis of gold nanoparticles: Potential in detection of liver cancer. Int. J. Nanomed. 2011, 6, 2305–2319. [Google Scholar]
- Das, S.K.; Das, A.R.; Guha, A.K. Gold nanoparticles: Microbial synthesis and application in water hygiene management. Langmuir 2009, 25, 8192–8199. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Raliya, R.; Rathore, I. Microbial synthesis of phosphorous nanoparticle from tri-calcium phosphate using Aspergillus tubingensis TFR-5. J. Bionanoscience 2012, 6, 84–89. [Google Scholar] [CrossRef]
- Sawle, B.D.; Salimath, B.; Deshpande, R.; Bedre, M.D.; Prabhakar, B.K.; Venkataraman, A. Biosynthesis and stabilization of Au and Au-Ag alloy nanoparticles by fungus. Fusarium semitectum. Sci. Technol. Adv. Mater. 2008, 9, 035012. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Raliya, R. Rapid, low-cost, and eco-friendly approach for iron nanoparticle synthesis using Aspergillus oryzae TFR9. J. Nanoparticles 2013, 141274. [Google Scholar] [CrossRef]
- Binupriya, A.R.; Sathishkumar, M.; Yun, S.I. Biocrystallization of silver and gold ions by inactive cell filtrate of Rhizopus stolonifer. Colloids Surf. B Biointerfaces 2010, 79, 531–534. [Google Scholar] [CrossRef]
- Raliya, R.; Biswas, P.; Tarafdar, J.C. TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.). Biotechnol. For. Rep. 2015, 5, 22–26. [Google Scholar] [CrossRef]
- Nature. Available online: https://www.nature.com/subjects/metabolic-engineering (accessed on 24 September 2019).
- Sharma, A.; Mistry, V.; Kumar, V.; Tiwari, P. Production of effective Phyto-antimicrobials via metabolic engineering strategies. Curr. Top. Med. Chem. 2022, 22, 1068–1092. [Google Scholar]
- Podolsky, I.; Seppala, S.; Lankiewicz, T.; Brown, J.; Swift, C.; O’Malley, M. Harnessing nature’s anaerobes for biotechnology and bioprocessing. Annu. Rev. Chem. Biomol. Eng. 2019, 10, 105–128. [Google Scholar] [CrossRef]
- Seppala, S.; Wilken, S.E.; Knop, D.; Solomon, K.V.; O’Malley, M.A. The importance of sourcing enzymes from non-conventional fungi for metabolic engineering and biomass breakdown. Metab. Eng. 2017, 44, 45–49. [Google Scholar] [CrossRef] [PubMed]
- de Backe, M.D.; van Dijck, P. Progress in functional genomics approaches to antifungal drug target discovery. Trends Microbiol. 2003, 11, 470–478. [Google Scholar] [CrossRef]
- Villena Gretty, K.; Kitazono, A.A.; Hernández-Macedo, M.L. Bioengineering fungi and yeast for the production of enzymes, metabolites, and value-added compounds. In Fungal Biotechnology and Bioengineering; Hesham, A.E.L., Ed.; Fungal Biology; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Lian, J.; Si, T.; Nair, N.U.; Zhao, H. Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. In Proceedings of the Food, Pharmaceutical, and Bioengineering Division 2014-Core Programming Area at the 2014 AIChE An-nual Meeting, Atlanta, GA, USA, 16–21 November 2014. [Google Scholar]
- Vieira Gomes, A.M.; Souza Carmo, T.; Silva Carvalho, L.; Mendonca Bahia, F.; Parachin, N.S. Comparison of yeasts as hosts for recombinant protein production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Wanka, F.; van Gent, J.; Arentshorst, M.; van den Hondel, C.A.; Ram, A.F. Fungal gene expression on demand: An inducible, tunable, and metabolism-independent expression system for Aspergillus niger. Appl. Environ. Microbiol. 2011, 77, 2975–2983. [Google Scholar] [CrossRef]
- Kuivanen, J.; Wang, Y.J.; Richard, P. Engineering Aspergillus niger for galactaric acid production: Elimination of galactaric acid catabolism by using RNA sequencing and CRISPR/Cas9. Microb. Cell Fact. 2016, 15, 210. [Google Scholar] [CrossRef]
- Xue, Z.; Sharpe, P.L.; Hong, S.P.; Yadav, N.S.; Xie, D.; Short, D.R.; Damude, H.G.; Rupert, R.A.; Seip, J.E.; Wang, J.; et al. Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat. Biotechnol. 2013, 31, 8. [Google Scholar] [CrossRef]
- Gao, S.; Lyu, Y.; Zeng, W.; Du, G.; Zhou, J.; Chen, J. Efficient biosynthesis of (2S)-Naringenin from p-Coumaric acid in Saccharomyces cerevisiae. J. Agric. Food Chem. 2019, 68, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, X.; Zhang, C.; Zhou, X.; Xu, X.; Han, L.; Lv, X.; Liu, Y.; Liu, S.; Li, J.; et al. De novo biosynthesis of rubusoside and rebaudiosides in engineered yeasts. Nat. Commun. 2022, 13, 3040. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, W.; Wang, Y.; Liu, H.; Li, X.; Yuan, Y. Lycopene overproduction in Saccharomyces cerevisiae through combining pathway engineering with host engineering. Microb. Cell Fact. 2016, 15, 113. [Google Scholar] [CrossRef] [PubMed]
- Amor, I.L.-B.; Hehn, A.; Guedone, E.; Ghedira, K.; Engasser, J.-M.; Chekir-Ghedrira, L. Mohamed Ghoul. 2010. Biotransformation of naringenin to eriodictyol by Saccharomyces cerevisiae functionally expressing flavonoid 3′ hydroxylase. Nat. Prod. Commun. 2010, 5, 1893–1898. [Google Scholar] [PubMed]
- Thompson, R.A.; Trinh, C.T. Enhancing fatty acid ethyl ester production in Saccharomyces cerevisiae through metabolic engineering and medium optimization. Biotechnol. Bioeng. 2014, 111, 2200–2208. [Google Scholar] [CrossRef]
- Den Haan, R.; Rose, S.H.; Lynd, L.R.; van Zyl, W.H. Hydrolysis and fermentation of amorphous cellulose by recombinant Saccharomyces cerevisiae. Metab. Eng. 2007, 9, 87–94. [Google Scholar] [CrossRef]
- Tang, X.; Feng, H.; Chen, W.N. Metabolic engineering for enhanced fatty acids synthesis in Saccharomyces cerevisiae. Metab. Eng. 2013, 16, 95–102. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Qin, J.; Siewers, V.; Nielsen, J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 2016, 7, 11709. [Google Scholar] [CrossRef] [PubMed]
- Paramasivan, K.; Mutturi, S. Progress in terpene synthesis strategies through the engineering of Saccharomyces cerevisiae. Crit. Rev. Biotechnol. 2017, 37, 974–989. [Google Scholar] [CrossRef]
- Weber, C.; Brückner, C.; Weinreb, S.; Lehr, C.; Essl, C.; Boles, E. Biosynthesis of cis, cis-muconic acid and its aromatic precursors, catechol and protocatechuic acid, from renewable feedstocks by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 8421–8430. [Google Scholar] [CrossRef]
- Ghosh, A.; Ando, D.; Gin, J.; Runguphan, W.; Denby, C.; Wang, G.; Baidoo, E.E.K.; Shymansky, C.; Keasling, J.D.; Martín, H.G. 13C metabolic flux analysis for systematic metabolic engineering of S. cerevisiae for overproduction of fatty acids. Front. Bioeng. Biotechnol. 2016, 4, 76. [Google Scholar] [CrossRef]
- Babaei, M.; Gheorghe, M.B.Z.; Chen, X.; Christensen, H.B.; Kristensen, M.; Nielsen, J.; Borodina, I. Metabolic engineering of Saccharomyces cerevisiae for rosmarinic acid production. ACS Synth. Biol. 2020, 9, 1978–1988. [Google Scholar] [CrossRef] [PubMed]
- Krömer, J.O.; Nunez-Bernal, D.; Averesch, N.J.H.; Hampe, J.; Varela, J.; Varela, C. Production of aromatics in Saccharomyces cerevisiae-a feasibility study. J. Biotechnol. 2013, 163, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, X.; Cha, Y.; Li, W.; Zhu, C.; Zhu, M.; Li, S.; Zhuo, M.; Huang, S.; Li, J. Stepwise engineering of Saccharomyces cerevisiae to produce (+)-valencene and its related sesquiterpenes. RSC Adv. 2019, 9, 30171–30181. [Google Scholar] [CrossRef] [PubMed]
- Runguphan, W.; Keasling, J.D. Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid-derived biofuels and chemicals. Metab. Eng. 2014, 21, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Lian, J.; Tu, S.; Xie, L.; Li, J.; Zhang, F.; Linhardt, R.J.; Huang, H.; Zhong, W. Metabolic engineering of Saccharomyces cerevisiae for high-level production of chlorogenic acid from glucose. ACS Synth. Biol. 2022, 11, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.N.S.; Koffas, M.A.G.; Stephanopoulos, G. Optimization of a heterologous pathway for the production of flavonoids from glucose. Metab. Eng. 2011, 13, 392–400. [Google Scholar] [CrossRef]
- Pai, J.S. Applications of microorganisms in food biotechnology. Indian J. Biotechnol. 2003, 2, 382–386. [Google Scholar]
- Lv, Y.; Edwards, H.; Zhou, J.; Xu, P. Combining 26s rDNA and the Cre-loxP system for iterative gene integration and efficient marker curation in Yarrowia lipolytica. ACS Synth. Biol. 2019, 8, 568–576. [Google Scholar] [CrossRef]
- Rosa, J.C.C.; Colombo, L.T.; Alvim, M.C.T.; Avonce, N.; Dijck, P.V.; Passos, F.M.L. Metabolic engineering of Kluyveromyces lactis for L-ascorbic acid (vitamin C) biosynthesis. Microb. Cell Fact. 2013, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- González-Siso, M.I.; Touriño, A.; Vizoso, Á.; Pereira-Rodríguez, Á.; Rodríguez-Belmonte, E.; Becerra, M.; Cerdán, M.E. Improved bioethanol production in an engineered Kluyveromyces lactis strain shifted from respiratory to fermentative metabolism by deletion of NDI1. Microb. Biotechnol. 2015, 8, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Rico-Díaz, A.; Álvarez-Cao, M.E.; Escuder-Rodríguez, J.J.; González-Siso, M.I.; Cerdán, M.E.; Becerra, M. Rational mutagenesis by engineering disulfide bonds improves Kluyveromyces lactis beta-galactosidase for high-temperature industrial applications. Sci. Rep. 2017, 7, 45535. [Google Scholar] [CrossRef]
- Fiedler, M.R.; Barthel, L.; Kubisch, C.; Nai, C.; Meyer, V. Construction of an improved Aspergillus niger platform for enhanced glucoamylase secretion. Microb. Cell Fact. 2018, 17, 95. [Google Scholar] [CrossRef]
- Carvalho, N.D.; Arentshorst, M.; Kooistra, R.; Stam, H.; Sagt, C.M.; van den Hondel, C.A.; Ram, A.F. Effects of a defective ERAD pathway on growth and heterologous protein production in Aspergillus niger. Appl. Microbiol. Biotechnol. 2011, 89, 357–373. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Jeenes, D.; Archer, D.B.; van den Hondel, C.A.M.J.J.; Punt, P.J. Calnexin overexpression increases manganese peroxidase production in Aspergillus niger. Appl. Environ. Microbiol. 2002, 68, 846–851. [Google Scholar] [CrossRef]
- Karnaukhova, E.; Ophir, Y.; Trinh, L.; Dalal, N.; Punt, P.J.; Golding, B.; Shiloach, J. Expression of human α1-proteinase inhibitor in Aspergillus niger. Microb Cell Fact. 2007, 6, 34. [Google Scholar] [CrossRef]
- Aifa, M.S.; Sayadi, S.; Gargouri, A. Heterologous expression of lignin peroxidase of Phanerochaete chrysosporium in Aspergillus niger. Biotechnol. Lett. 1999, 21, 849–853. [Google Scholar] [CrossRef]
- Richter, L.; Wanka, F.; Boecker, S.; Storm, D.; Kurt, T.; Vural, O.; Süßmuth, R.; Meyer, V. Engineering of Aspergillus niger for the production of secondary metabolites. Fungal Biol. Biotechnol. 2014, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Wakai, S.; Nakashima, N.; Ogino, C.; Tsutsumi, H.; Hata, Y.; Kondo, A. Modified expression of multi-cellulases in a filamentous fungus Aspergillus oryzae. Bioresour. Technol. 2019, 276, 146–153. [Google Scholar] [CrossRef]
- Kuivanen, J.; Penttilä, M.; Richard, P. Metabolic engineering of the fungal d-galacturonate pathway for l-ascorbic acid production. Microb. Cell Fact. 2015, 14, 2. [Google Scholar] [CrossRef]
- Kraševec, N.; Milunović, T.; Lasnik, M.A.; Lukančič, I.; Komel, R.; Porekar, V.G. Human granulocyte colony-stimulating factor (G-CSF) produced in the filamentous fungus Aspergillus niger. Acta Chim. Slov. 2014, 61, 709–717. [Google Scholar]
- Wriessnegger, T.; Augustin, P.; Engleder, M.; Leitner, E.; Müller, M.; Kaluzna, I.; Schürmann, M.; Mink, D.; Zellnig, G.; Schwab, H.; et al. Production of the sesquiterpenoid (+)-nootkatone by metabolic engineering of Pichia pastoris. Metab. Eng. 2014, 24, 18–29. [Google Scholar] [CrossRef]
- Cai, P.; Wu, X.; Deng, J.; Gao, L.; Shen, Y.; Yao, L.; Zhou, Y.J. Methanol biotransformation toward high-level production of fatty acid derivatives by engineering the industrial yeast Pichia pastoris. Proc. Natl. Acad. Sci. USA 2022, 119, e2201711119. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Dai, Z.; Peng, W.; Zhang, S.; Zhou, J.; Ma, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Metabolic engineering of Pichia pastoris for malic acid production from methanol. Biotechnol. Bioeng. 2021, 1, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Larroude, M.; Celinska, E.; Back, A.; Thomas, S.; Nicaud, J.M.; Ledesma-Amaro, R. A synthetic biology approach to transform Yarrowia lipolytica into a competitive biotechnological producer of b-carotene. Biotechnol. Bioeng. 2018, 115, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Fitz, E.; Gamauf, C.; Seiboth, B.; Wanka, F. Deletion of the small GTPase rac1 in Trichoderma reesei provokes hyperbranching and impacts growth and cellulase production. Fungal Biol. Biotechnol. 2019, 6, 16. [Google Scholar] [CrossRef]
- Lin, L.; Sun, Z.; Li, J.; Chen, Y.; Liu, Q.; Sun, W.; Tian, C. Disruption of gul-1 decreased the culture viscosity and improved protein secretion in the filamentous fungus Neurospora crassa. Microb. Cell Fact. 2018, 17, 96. [Google Scholar] [CrossRef]
- Sakamoto, T.; Sakuradani, E.; Okuda, T.; Kikukawa, H.; Ando, A.; Kishino, S.; Izumi, Y.; Bamba, T.; Shima, J.; Ogawa, J. Metabolic engineering of oleaginous fungus Mortierella alpina for high production of oleic and linoleic acids. Bioresour. Technol. 2017, 247, 1610–1615. [Google Scholar] [CrossRef]
- Pollmann, H.; Breitenbach, J.; Sandmann, G. Engineering of the carotenoid pathway in Xanthophyllomyces dendrorhous leading to the synthesis of zeaxanthin. Appl. Microbiol. Biotechnol. 2017, 101, 103–111. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, X.Y.; Xiong, L.; Mehmood, M.A.; Zhao, X.; Bai, F. On-site cellulase production and efficient saccharification of corn Stover employing cbh2 overexpressing Trichoderma reesei with novel induction system. Bioresour. Technol. 2017, 238, 643–649. [Google Scholar] [CrossRef]
- Wang, S.; Liu, G.; Wang, J.; Yu, J.; Huang, B.; Xing, M. Enhancing cellulase production in Trichoderma reesei RUT C30 through combined manipulation of activating and repressing genes. J. Ind. Microbiol. Biotechnol. 2013, 40, 633–641. [Google Scholar] [CrossRef]
- Mattern, D.J.; Valiante, V.; Horn, F.; Petzke, L.; Brakhage, A.A. Rewiring of the Austinoid biosynthetic pathway in filamentous fungi. ACS Chem. Biol. 2017, 12, 2927–2933. [Google Scholar] [CrossRef]
- Prielhofer, R.; Barrero, J.J.; Steuer, S.; Gassler, T.; Zahrl, R.; Baumann, K.; Sauer, M.; Mattanovich, D.; Gasser, B.; Marx, H. GoldenPiCS: A Golden Gate-derived modular cloning system for applied synthetic biology in the yeast Pichia pastoris. BMC Syst. Biol. 2017, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Rodicio, R.; Heinisch, J.J. Yeast on the milky way: Genetics, physiology, and biotechnology of Kluyveromyces lactis. Yeast 2013, 30, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and its multiple applications in the biotechnological industry. Sci. World J. 2014, 2014, 476207. [Google Scholar] [CrossRef] [PubMed]
- Vickers, C.E.; Williams, T.C.; Peng, B.; Cherry, J. Recent advances in synthetic biology for engineering isoprenoid production in yeast. Curr. Opin. Chem. Biol. 2017, 40, 47–56. [Google Scholar] [CrossRef]
- Dilworth, M.V.; Piel, M.S.; Bettaney, K.E.; Ma, P.; Luo, J.; Sharples, D.; Poyner, D.R.; Gross, S.R.; Moncoq, K.; Henderson, P.J.F.; et al. Microbial expression systems for membrane proteins. Methods 2018, 147, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, M.; Nielsen, J. Exploring the potential of Saccharomyces cerevisiae for biopharmaceutical protein production. Curr. Opin. Biotechnol. 2017, 48, 77–84. [Google Scholar] [CrossRef]
- Tiwari, P.; Bajpai, M.; Singh, L.K.; Mishra, S.; Yadav, A.N. Phytohormones producing fungal communities: Metabolic engineering for abiotic stress tolerance in plants. In Agriculturally Important Fungi for Sustainable Agriculture; Gupta, V.J., Tuohy, M., Eds.; Springer publication: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Duina, A.A.; Miller, M.E.; Keeney, J.B. Budding yeast for budding geneticists: A primer on the Saccharomyces cerevisiae model system. Genetics 2014, 197, 33–48. [Google Scholar] [CrossRef]
- Stovicek, V.; Holkenbrink, C.; Borodina, I. CRISPR/Cas system for yeast genome engineering: Advances and applications. FEMS Yeast Res. 2017, 17, 1–16. [Google Scholar] [CrossRef]
- Schwartz, C.; Wheeldon, I. CRISPR-Cas9-mediated genome editing and transcriptional control in Yarrowia lipolytica. Methods Mol. Biol. 2018, 1772, 327–345. [Google Scholar]
- Meyer, V. Genetic engineering of filamentous fungi—Progress, obstacles, and future trends. Biotechnol. Adv. 2008, 26, 177–185. [Google Scholar] [CrossRef]
- Kluge, J.; Terfehr, D.; Kück, U. Inducible promoters and functional genomic approaches for the genetic engineering of filamentous fungi. Appl. Microbiol. Biotechnol. 2018, 102, 6357–6372. [Google Scholar] [CrossRef]
- Schuetze, T.; Meyer, V. Polycistronic gene expression in Aspergillus niger. Microb. Cell Fact. 2017, 16, 162. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Pandit, S.G.; Puttananjaiah, M.H.; Harohally, N.V.; Dhale, M.A. Structural and functional characterization of new pigment molecule monashin from Monascus purpureus CFR410-11. Process. Biochem. 2019, 82, 173–178. [Google Scholar] [CrossRef]
- Shi, Y.C.; Pan, T.M.; Liao, V.H.C. Monascin from Monascus-fermented products reduces oxidative stress and amyloid-beta toxicity via DAF-16/FOXO in Caenorhabditis elegans. J. Agric. Food. Chem. 2016, 64, 7114–7120. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Cheng, M.J.; Wu, M.D.; Chen, J.J.; Chen, Y.L.; Chang, H.S.; Chen, K.P. Secondary metabolites from the fermented rice of the fungus Monascus purpureus and their bioactivities. Nat. Prod. Res. 2019, 33, 3541–3550. [Google Scholar] [CrossRef]
- American Chemical Society (ACS). Dyes, Pigments, and Inks. 2020. Available online: https://www.acs.org/content/acs/en/careers/college-to-career/chemistry-careers/dyes-pigments-ink.html (accessed on 8 September 2020).
- Pickens, L.B.; Tang, Y.; Chooi, Y.-H. Metabolic engineering for the production of Natural products. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 211–236. [Google Scholar] [CrossRef]
- Parekh, S. Strain improvement. In The Desk Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 960–973. [Google Scholar]
- Rokem, J.S.; Lantz, A.E.; Nielsen, J. Systems biology of antibiotic production by microorganisms. Nat. Prod. Rep. 2007, 24, 1262–1287. [Google Scholar] [CrossRef]
- Parekh, S.; Vinci, V.A.; Strobel, R.J. Improvement of microbial strains and fermentation processes. Appl. Microbiol. Biotechnol. 2000, 54, 287–301. [Google Scholar] [CrossRef]
- Li, X.-H.; Yang, H.-J.; Roy, B.; Park, E.Y.; Jiang, L.-J.; Wang, D.; Miao, Y.-G. Enhanced cellulase production of the Trichoderma viride mutated by microwave and ultraviolet. Microbiol. Res. 2010, 165, 190–198. [Google Scholar] [CrossRef]
- Peterson, R.; Nevalainen, H. Trichoderma reesei RUT-C30–thirty years of strain improvement. Microbiology 2012, 158, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Ziemons, S.; Koutsantas, K.; Becker, K.; Dahlmann, T.; Kück, U. Penicillin production in industrial strain Penicillium chrysogenum P2niaD18 is not dependent on the copy number of biosynthesis genes. BMC Biotechnol. 2017, 17, 1. [Google Scholar] [CrossRef]
- Zhgun, A.A. Random mutagenesis of filamentous fungi strains for high-yield production of secondary metabolites: The role of polyamines. In Genotoxicity and Mutagenicity—Mechanisms and Test Methods; Soloneski, S., Larramendy, M.L., Eds.; Intech Open: London, UK, 2020. [Google Scholar] [CrossRef]
- Yang, T.; Li, L.; Wang, B.; Tian, J.; Shi, F.; Zhang, S.; Wu, Z. Isolation, mutagenesis, and organic acid secretion of a highly efficient Phosphate-solubilizing fungus. Front. Microbiol. 2022, 13, 793122. [Google Scholar] [CrossRef]
- Smith, D.J.; Bull, J.H.; Edwards, J.; Turner, G. Amplification of the isopenicillin N synthetase gene in a strain of Penicillium chrysogenum producing high levels of penicillin. Mol. Gen. Genet. 1989, 216, 492–497. [Google Scholar] [CrossRef]
- Keasling, J.D. Gene-expression tools for the metabolic engineering of bacteria. Trends Biotechnol. 1999, 17, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Leonard, E.; Ajikumar, P.K.; Thayer, K.; Xiao, W.H.; Mo, J.D.; Tidor, B.; Stephanopoulos, G.; Prather, K.L.J. Combining metabolic and protein engineering of a terpenoid biosynthetic pathway for overproduction and selectivity control. Proc. Natl. Acad. Sci. USA 2010, 107, 13654–13659. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Turner, G. δ-(L-α-Aminoadipyl)-L-cysteinyl-D-valine synthetase is a rate-limiting enzyme for Penicillin production in Aspergillus nidulans. Mol. Gen. Genet. 1996, 253, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Cañón, J.M.; Peñalva, M.A. Overexpression of two Penicillin structural genes in Aspergillus nidulans. Mol. Gen. Genet. 1995, 246, 110–118. [Google Scholar] [CrossRef]
- Malla, S.; Niraula, N.P.; Liou, K.; Sohng, J.K. Enhancement of doxorubicin production by expression of structural sugar biosynthesis and glycosyltransferase genes in Streptomyces peucetius. J. Biosci. Bioeng. 2009, 108, 92–98. [Google Scholar] [CrossRef]
- Malla, S.; Niraula, N.P.; Liou, K.; Sohng, J.K. Self-resistance mechanism in Streptomyces peucetius: Overexpression of drrA, drrB and drrC for doxorubicin enhancement. Microbiol. Res. 2010, 165, 259–267. [Google Scholar] [CrossRef]
- Yihua, C.; Smanski, M.J.; Shen, B. Improvement of secondary metabolite production in Streptomyces by manipulating pathway regulation. Appl. Microbiol. Biotechnol. 2010, 86, 19–25. [Google Scholar]
- Yihua, C.; Wendt-Pienkowski, E.; Shen, B. Identification and utility of FdmR1 as a Streptomyces antibiotic regulatory protein activator for fredericamycin production in Streptomyces griseus ATCC 49344 and heterologous hosts. J. Bacteriol. 2008, 190, 5587–5596. [Google Scholar]
- Lombo, F.; Brana, A.F.; Mendez, C.; Salas, J.A. The Mithramycin gene cluster of Streptomyces argillaceus contains a positive regulatory gene and two repeated DNA sequences that are located at both ends of the cluster. J. Bacteriol. 1999, 181, 642–647. [Google Scholar] [CrossRef]
- Ryu, Y.-G.; Butler, M.J.; Chater, K.; Lee, K.J. Engineering of primary carbohydrate metabolism for increased production of actinorhodin in Streptomyces coelicolor. Appl. Environ. Microbiol. 2006, 72, 7132–7139. [Google Scholar] [CrossRef]
- Zha, W.; Rubin-Pitel, S.B.; Shao, Z.; Zhao, H. Improving cellular malonyl-CoA level in Escherichia coli via metabolic engineering. Metab. Eng. 2009, 11, 192–198. [Google Scholar] [CrossRef]
- Kajiwara, S.; Fraser, P.D.; Kondo, K.; Misawa, N. Expression of an exogenous isopentenyl diphosphate isomerase gene enhances isoprenoid biosynthesis in Escherichia coli. Biochem. J. 1997, 324, 421–426. [Google Scholar] [CrossRef]
- Kim, S.W.; Keasling, J.D. Metabolic engineering of the non-mevalonate isopentenyl diphosphate synthesis pathway in Escherichia coli enhances lycopene production. Biotechnol. Bioeng. 2001, 72, 408–415. [Google Scholar] [CrossRef]
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for the production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef]
- Thykaer, J.; Nielsen, J.; Wohlleben, W.; Weber, T.; Gutknecht, M.; Lantz, A.E.; Stegmann, E. Increased glycopeptide production after overexpression of shikimate pathway genes being part of the Balhimycin biosynthetic gene cluster. Metab. Eng. 2010, 12, 455–461. [Google Scholar] [CrossRef]
- Lomovskaya, N.; Otten, S.L.; Doi-Katayama, Y.; Fonstein, L.; Liu, X.-C.; Takatsu, T.; Inventi-Solari, A.; Filippini, S.; Torti, F.; Colombo, A.L.; et al. Doxorubicin overproduction in Streptomyces peucetius: Cloning and characterization of the dnrU ketoreductase and dnrV genes and the doxA cytochrome P-450 hydroxylase gene. J. Bacteriol. 1999, 181, 305–318. [Google Scholar] [CrossRef]
- Chemler, J.A.; Fowler, Z.L.; McHugh, K.P.; Koffas, M.A.G. Improving NADPH availability for natural product biosynthesis in Escherichia coli by metabolic engineering. Metab. Eng. 2010, 12, 96–104. [Google Scholar] [CrossRef]
- Komatsu, M.; Uchiyama, T.; Omura, S.; Cane, D.E.; Ikeda, H. Genome-minimized Streptomyces host for the heterologous expression of secondary metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 2646–2651. [Google Scholar] [CrossRef]
- Hopwood, D.; Malpartida, F.; Kieser, H.M.; Ikeda, H.; Duncan, J.; Fujii, I.; Rudd, B.A.M.; Floss, H.; Ōmura, S. Production of ‘hybrid’ antibiotics by genetic engineering. Nature 1985, 314, 642–644. [Google Scholar] [CrossRef] [PubMed]
- de Boer, A.L.; Schmidt-Dannert, C. Recent efforts in engineering microbial cells to produce new chemical compounds. Curr. Opin. Chem. Biol. 2003, 7, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Punt, P.J.; van Biezen, N.; Conesa, A.; Albers, A.; Mangnus, J.; van den Hondel, C. Filamentous fungi as cell factories for heterologous protein production. Trends Biotechnol. 2002, 20, 200–206. [Google Scholar] [CrossRef]
- Mackenzie, D.A.; Wongwathanarat, P.; Carter, A.T.; Archer, D.B. Isolation and use of a homologous histone H4 promoter and a ribosomal DNA region in a transformation vector for the oil-producing fungus Mortierella alpine. Appl. Environ. Microbiol. 2000, 66, 4655–4661. [Google Scholar] [CrossRef] [PubMed]
- Royer, J.C.; Moyer, D.L.; Yoder, W.; Shuster, J.R. Patent Application WO 96/00787; World Intellectual Property Organization: Geneva, Switzerland, 1996. [Google Scholar]
- Berka, R.M.; Feng, X.U.; Thompsom, S.A. World Patent Application WO 95/15391; World Intellectual Property Organization: Geneva, Switzerland, 1995. [Google Scholar]
- Emalfarb, M.A.; Punt, P.J.; van Zeijl, C.; van den Hondel, C. World Patent Application WO 00/20555; World Intellectual Property Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Chen, B.-X.; Wei, T.; Ye, Z.-W.; Yun, F.; Kang, L.-Z.; Tang, H.-B.; Guo, L.-Q.; Lin, J.-F. Efficient CRISPR-Cas9 gene disruption system in edible-medicinal mushroom Cordyceps militaris. Front. Microbiol. 2018, 9, 1157. [Google Scholar] [CrossRef]
- Chen, B.-X.; Xue, L.-N.; Wei, T.; Ye, Z.-W.; Li, X.-H.; Guo, L.-Q.; Lin, J.-F. Enhancement of Ergothioneine production by discovering and regulating its metabolic pathway in Cordyceps militaris. Microb. Cell Fact. 2022, 21, 169. [Google Scholar] [CrossRef]
- Wang, H.N.; Ke, X.; Zhou, J.P.; Liu, Z.Q.; Zheng, Y.G. Recent advances in metabolic regulation and bioengineering of gibberellic acid biosynthesis in Fusarium fujikuroi. World J. Microbiol. Biotechnol. 2022, 38, 131. [Google Scholar] [CrossRef]
- Iyyappan, J.; Bharathiraja, B.; Baskar, G.; Kamalanaban, E. Process optimization and kinetic analysis of malic acid production from crude glycerol using Aspergillus niger. Bioresour. Technol. 2019, 281, 18–25. [Google Scholar] [CrossRef]
- Kallscheuer, N. Engineered microorganisms for the production of food additives approved by the European Union-A systematic analysis. Front. Microbiol. 2018, 9, 1746. [Google Scholar] [CrossRef]
- Tramontin, L.R.R.; Kildegaard, K.R.; Sudarsan, S.; Borodina, I. Enhancement of astaxanthin biosynthesis in oleaginous yeast Yarrowia lipolytica via the microalgal pathway. Microorganisms 2019, 7, 472. [Google Scholar] [CrossRef]
- Sun, X.; Su, X. Harnessing the knowledge of protein secretion for enhanced protein production in filamentous fungi. World J. Microbiol. Biotech. 2019, 35, 54. [Google Scholar] [CrossRef] [PubMed]
- Chankhamjon, P.; Boettger-Schmidt, D.; Scherlach, K.; Urbansky, B.; Lackner, G.; Kalb, D.; Dahse, H.M.; Hoffmeister, D.; Hertweck, C. Biosynthesis of the halogenated mycotoxin aspirochlorine in koji mold involves a cryptic amino acid conversion. Angew. Chem. Int. Ed. Engl. 2014, 53, 13409–13413. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Yan, D.; Zhang, T.; Guo, Y.; Liu, Y.; Zou, Y.; Tang, M.; Liu, B.; Wu, Q.; Yu, S.; et al. A cascade of redox reactions generates complexity in the biosynthesis of the protein phosphatase-2 inhibitor rubratoxin A. Angew. Chem. Int. Ed. Engl. 2017, 56, 4782–4786. [Google Scholar] [CrossRef]
- Newman, A.G.; Townsend, C.A. Molecular characterization of the cercosporin biosynthetic pathway in the fungal plant pathogen Cercospora nicotianae. J. Am. Chem. Soc. 2016, 138, 4219–4228. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, R.; Li, X.; Fu, D.; Zhu, B.; Tian, H.; Luo, Y.; Zhu, H. Multiplexed CRISPR/Cas9-mediated metabolic engineering of γ-aminobutyric acid levels in Solanum lycopersicum. Plant Biotechnol. J. 2018, 16, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Gouka, R.J.; Punt, P.J.; Van Den Hondel, C.A.M.J.J. Efficient production of secreted proteins by Aspergillus: Progress, limitations, and prospects. Appl. Microbiol. Biotechnol. 1997, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wakai, S.; Arazoe, T.; Ogino, C.; Kondo, A. Future insights in fungal metabolic engineering. Bioresour. Technol. 2017, 245, 1314–1326. [Google Scholar] [CrossRef]
- Sakai, K.; Kinoshita, H.; Nihira, T. Heterologous expression system in Aspergillus oryzae for fungal biosynthetic gene clusters of secondary metabolites. Appl. Microbiol. Biotechnol. 2012, 93, 2011–2022. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, P.; Dufossé, L. Focus and Insights into the Synthetic Biology-Mediated Chassis of Economically Important Fungi for the Production of High-Value Metabolites. Microorganisms 2023, 11, 1141. https://doi.org/10.3390/microorganisms11051141
Tiwari P, Dufossé L. Focus and Insights into the Synthetic Biology-Mediated Chassis of Economically Important Fungi for the Production of High-Value Metabolites. Microorganisms. 2023; 11(5):1141. https://doi.org/10.3390/microorganisms11051141
Chicago/Turabian StyleTiwari, Pragya, and Laurent Dufossé. 2023. "Focus and Insights into the Synthetic Biology-Mediated Chassis of Economically Important Fungi for the Production of High-Value Metabolites" Microorganisms 11, no. 5: 1141. https://doi.org/10.3390/microorganisms11051141
APA StyleTiwari, P., & Dufossé, L. (2023). Focus and Insights into the Synthetic Biology-Mediated Chassis of Economically Important Fungi for the Production of High-Value Metabolites. Microorganisms, 11(5), 1141. https://doi.org/10.3390/microorganisms11051141