

Effect of Methionine on Gene Expression in Komagataella phaffii Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media and Cultivation Conditions
2.2. Yeast and Bacterial Strains
2.3. Molecular Methods
2.4. Enzymatic Assays
2.5. RNA-Sequencing
2.6. Bioinformatic Analysis
2.7. Gene Nomenclature
3. Results
3.1. Addition of Methionine Does Not Enhance Growth of K. phaffii X-33 Strain
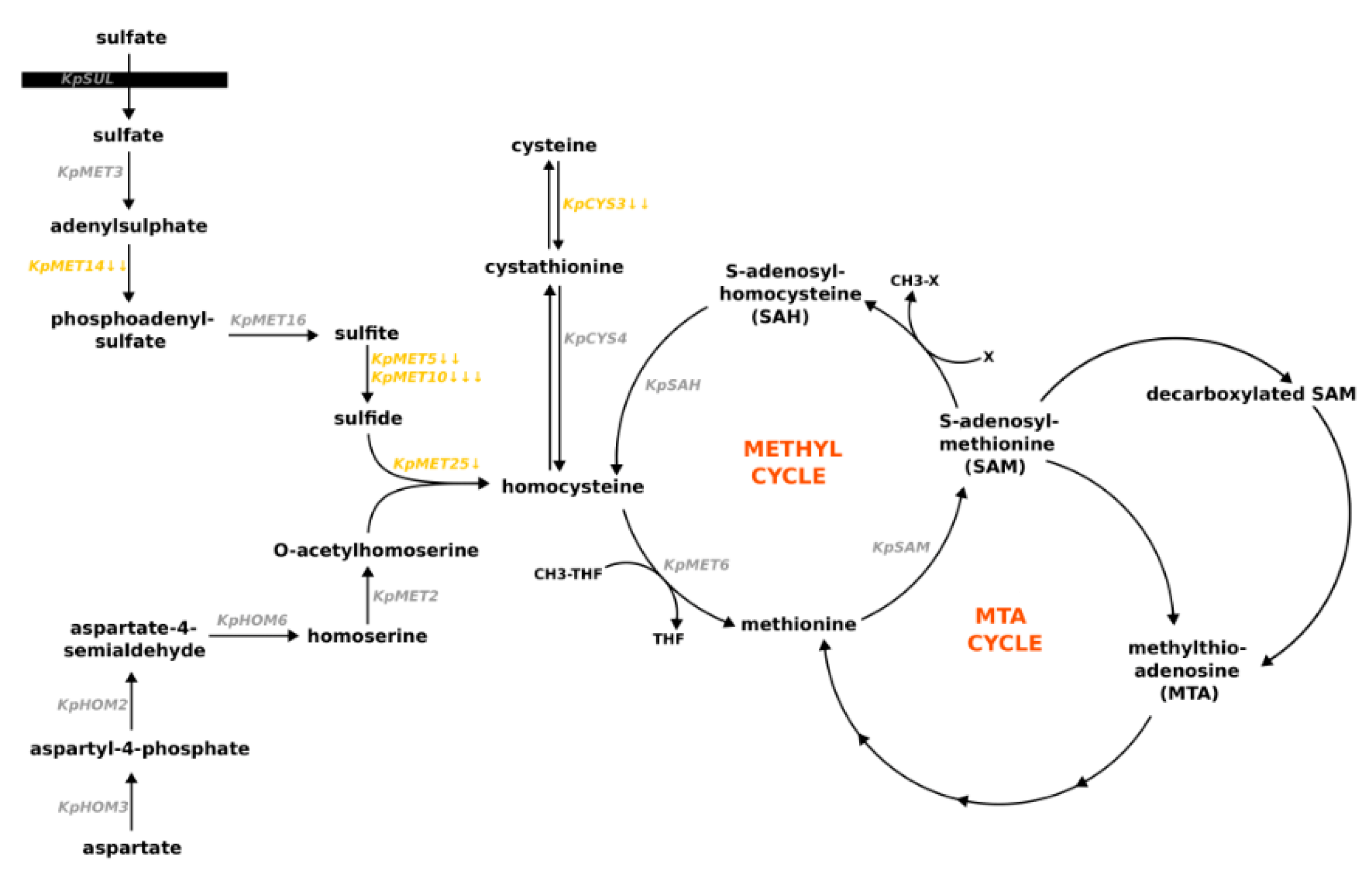
3.2. Methionine Influences the Expression of Genes Involved in Its Synthesis and Degradation in K. phaffii Cells
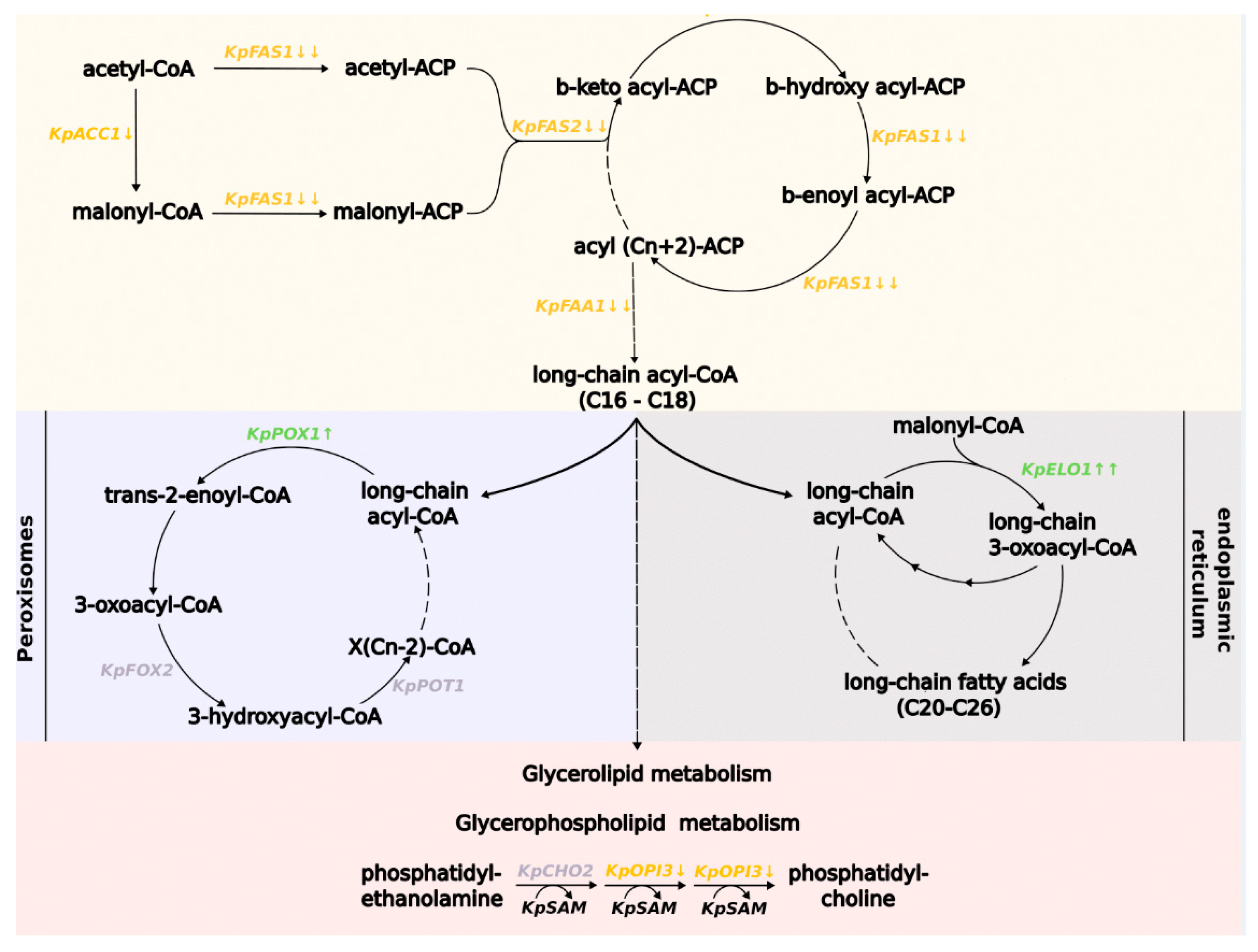
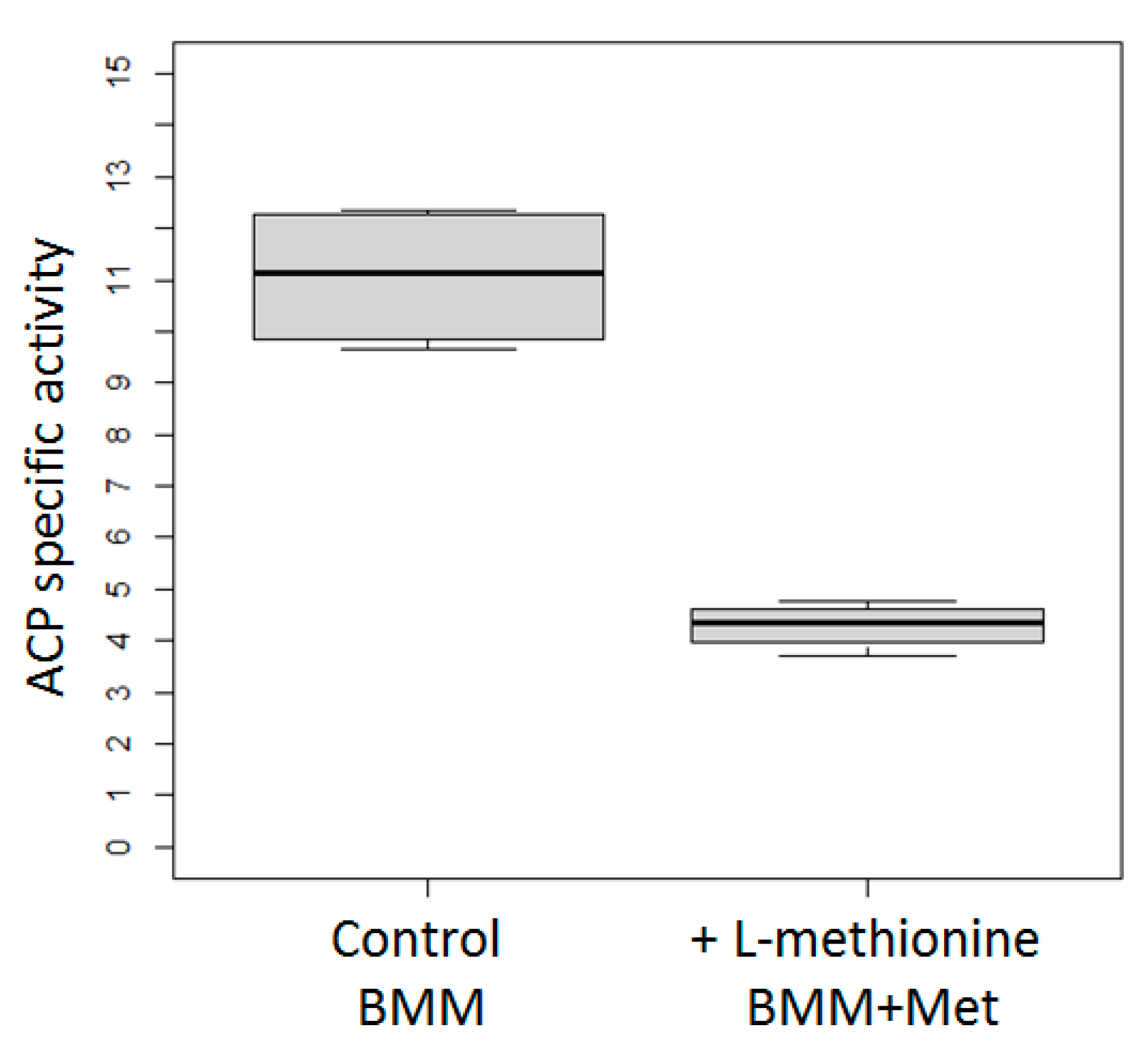
3.3. Methionine Influences the Expression of Genes Involved in Fatty Acid Metabolism in K. phaffii Cells
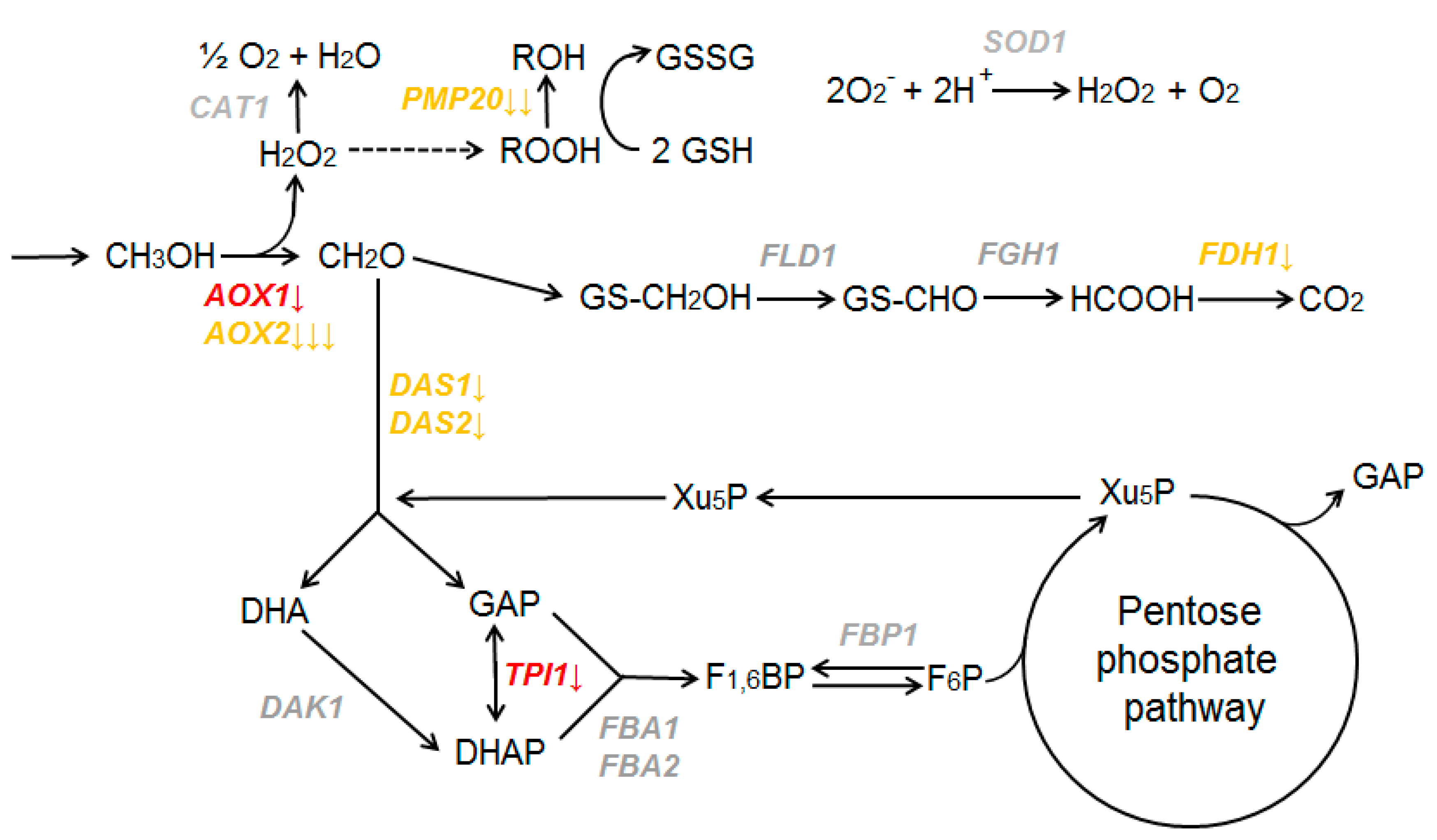
3.4. Methionine Influences the Expression of Genes Involved in Methanol Utilization in K. phaffii Cells
3.5. AOX1 Gene Promoter Is Repressed in Medium with Methionine
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Athmaram, T.N.; Singh, A.K.; Saraswat, S.; Srivastava, S.; Misra, P.; Kameswara Rao, M.; Gopalan, N.; Rao, P.V.L. A Simple Pichia pastoris Fermentation and Downstream Processing Strategy for Making Recombinant Pandemic Swine Origin Influenza a Virus Hemagglutinin Protein. J. Ind. Microbiol. Biotechnol. 2013, 40, 245–255. [Google Scholar] [CrossRef]
- Liu, W.-C.; Gong, T.; Wang, Q.-H.; Liang, X.; Chen, J.-J.; Zhu, P. Scaling-up Fermentation of Pichia pastoris to Demonstration-Scale Using New Methanol-Feeding Strategy and Increased Air Pressure Instead of Pure Oxygen Supplement. Sci. Rep. 2016, 6, 18439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-C.; Inwood, S.; Gong, T.; Sharma, A.; Yu, L.-Y.; Zhu, P. Fed-Batch High-Cell-Density Fermentation Strategies for Pichia pastoris Growth and Production. Crit. Rev. Biotechnol. 2019, 39, 258–271. [Google Scholar] [CrossRef]
- Balamurugan, V.; Reddy, G.R.; Suryanarayana, V.V.S. Pichia pastoris: A Notable Heterologous Expression System for the Production of Foreign Proteins—Vaccines. 1 April 2007. Available online: http://nopr.niscpr.res.in/handle/123456789/3021 (accessed on 28 February 2023).
- Jacobs, P.P.; Geysens, S.; Vervecken, W.; Contreras, R.; Callewaert, N. Engineering Complex-Type N-Glycosylation in Pichia pastoris Using GlycoSwitch Technology. Nat. Protoc. 2009, 4, 58–70. [Google Scholar] [CrossRef]
- Li, P.; Anumanthan, A.; Gao, X.-G.; Ilangovan, K.; Suzara, V.V.; Düzgüneş, N.; Renugopalakrishnan, V. Expression of Recombinant Proteins in Pichia pastoris. Appl. Biochem. Biotechnol. 2007, 142, 105–124. [Google Scholar] [CrossRef]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein Expression in Pichia pastoris: Recent Achievements and Perspectives for Heterologous Protein Production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghban, R.; Farajnia, S.; Ghasemi, Y.; Mortazavi, M.; Zarghami, N.; Samadi, N. New Developments in Pichia pastoris Expression System, Review and Update. Curr. Pharm. Biotechnol. 2020, 19, 451–467. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A Highly Successful Expression System for Optimal Synthesis of Heterologous Proteins. J. Cell Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, S.; Wang, Y. Recent Advances in the Production of Recombinant Subunit Vaccines in Pichia pastoris. Bioengineered 2016, 7, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahic, M.; Veide, A.; Charoenrat, T.; Teeri, T.; Enfors, S.-O. Process Technology for Production and Recovery of Heterologous Proteins with Pichia pastoris. Biotechnol. Prog. 2006, 22, 1465–1473. [Google Scholar] [CrossRef]
- Byrne, B. Pichia pastoris as an Expression Host for Membrane Protein Structural Biology. Curr. Opin. Struct. Biol. 2015, 32, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Hirz, M.; Richter, G.; Leitner, E.; Wriessnegger, T.; Pichler, H. A Novel Cholesterol-Producing Pichia pastoris Strain Is an Ideal Host for Functional Expression of Human Na,K-ATPase A3β1 Isoform. Appl. Microbiol. Biotechnol. 2013, 97, 9465–9478. [Google Scholar] [CrossRef]
- Tanaka, K.; Ishii, Y.; Ogawa, J.; Shima, J. Enhancement of Acetic Acid Tolerance in Saccharomyces cerevisiae by Overexpression of the HAA1 Gene, Encoding a Transcriptional Activator. Appl. Environ. Microbiol. 2012, 78, 8161–8163. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, C.V.G.C.; Serra, L.A.; Pacheco, T.F.; Ferreira, L.M.M.; Brandão, L.T.D.; Freitas, M.N.d.M.; Trichez, D.; Almeida, J.R.M.d. Advances in Komagataella phaffii Engineering for the Production of Renewable Chemicals and Proteins. Fermentation 2022, 8, 575. [Google Scholar] [CrossRef]
- Bernauer, L.; Radkohl, A.; Lehmayer, L.G.K.; Emmerstorfer-Augustin, A. Komagataella phaffii as Emerging Model Organism in Fundamental Research. Front. Microbiol. 2021, 11, 607028. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.M.; Madden, K.R.; Barringer, K.J.; Thill, G.P.; Stillman, C.A. Functional Characterization of the Two Alcohol Oxidase Genes from the Yeast Pichia pastoris. Mol. Cell Biol. 1989, 9, 1316–1323. [Google Scholar] [CrossRef]
- Rußmayer, H.; Buchetics, M.; Gruber, C.; Valli, M.; Grillitsch, K.; Modarres, G.; Guerrasio, R.; Klavins, K.; Neubauer, S.; Drexler, H.; et al. Systems-Level Organization of Yeast Methylotrophic Lifestyle. BMC Biol. 2015, 13, 80. [Google Scholar] [CrossRef] [Green Version]
- Berrios, J.; Theron, C.W.; Steels, S.; Ponce, B.; Velastegui, E.; Bustos, C.; Altamirano, C.; Fickers, P. Role of Dissimilative Pathway of Komagataella phaffii (Pichia pastoris): Formaldehyde Toxicity and Energy Metabolism. Microorganisms 2022, 10, 1466. [Google Scholar] [CrossRef] [PubMed]
- Tschopp, J.F.; Brust, P.F.; Cregg, J.M.; Stillman, C.A.; Gingeras, T.R. Expression of the LacZ Gene from Two Methanol-Regulated Promoters in Pichia pastoris. Nucleic Acids Res. 1987, 15, 3859–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SEllis, S.B.; Brust, P.F.; Koutz, P.J.; Waters, A.F.; Harpold, M.M.; Gingeras, T.R. Isolation of alcohol oxidase and two other methanol regulatable genes from the yeast Pichia pastoris. Mol. Cell Biol. 1985, 5, 1111–1121. [Google Scholar] [CrossRef]
- Kalender, Ö.; Çalık, P. Transcriptional Regulatory Proteins in Central Carbon Metabolism of Pichia pastoris and Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2020, 104, 7273–7311. [Google Scholar] [CrossRef]
- Rumjantsev, A.M.; Padkina, M.V.; Sambuk, E.V. Effect of Nitrogen Source on Gene Expression of First Steps of Methanol Utilization Pathway in Pichia pastoris. Russ. J. Genet. 2013, 49, 394–400. [Google Scholar] [CrossRef]
- Rumjantsev, A.M.; Bondareva, O.V.; Padkina, M.V.; Sambuk, E.V. Effect of Nitrogen Source and Inorganic Phosphate Concentration on Methanol Utilization and PEX Genes Expression in Pichia pastoris. Sci. World J. 2014, 743615. [Google Scholar] [CrossRef] [Green Version]
- Sahu, U.; Rangarajan, P.N. Methanol Expression Regulator 1 (Mxr1p) Is Essential for the Utilization of Amino Acids as the Sole Source of Carbon by the Methylotrophic Yeast, Pichia pastoris. J. Biol. Chem. 2016, 291, 20588–20601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumyantsev, A.M.; Soloviev, G.A.; Slepchenkov, A.V.; Sambuk, E.V. Effects of Deletions in Pichia pastoris RTG Genes on Phenotype and AOX1 Expression. Adv. Microbiol. 2018, 8, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Rumyantsev, A.; Sidorin, A.; Volkov, A.; Al Shanaa, O.; Sambuk, E.; Padkina, M. Transcriptome Analysis Unveils the Effects of Proline on Gene Expression in the Yeast Komagataella phaffii. Microorganisms 2021, 10, 67. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Che, Y.; Yang, D.; Wang, Q.; Yang, H.; Boutet, J.; Huet, R.; Yin, S. Production of L-Methionine from 3-Methylthiopropionaldehyde and O-Acetylhomoserine by Catalysis of the Yeast O-Acetylhomoserine Sulfhydrylase. J Agric. Food. Chem. 2021, 69, 7932–7937. [Google Scholar] [CrossRef] [PubMed]
- Rajakumar, S.; Suriyagandhi, V.; Nachiappan, V. Impairment of MET Transcriptional Activators, MET4 and MET31 Induced Lipid Accumulation in Saccharomyces cerevisiae. FEMS Yeast Res. 2020, 20, foaa039. [Google Scholar] [CrossRef] [PubMed]
- Walvekar, A.S.; Kadamur, G.; Sreedharan, S.; Gupta, R.; Srinivasan, R.; Laxman, S. Methylated PP2A stabilizes Gcn4 to enable a methionine-induced anabolic program. J. Biol. Chem. 2020, 295, 18390–18405. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. Independent and Additive Effects of Glutamic Acid and Methionine on Yeast Longevity. PLoS ONE 2013, 8, e79319. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.; Vowinckel, J.; Keller, M.A.; Ralser, M. Methionine Metabolism Alters Oxidative Stress Resistance via the Pentose Phosphate Pathway. Antioxid. Redox Sign. 2016, 24, 543–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.-A.A.; Ren, W.; Yin, Y. The Role of Methionine on Metabolism, Oxidative Stress, and Diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Ouyang, Q.; Li, H.; Zheng, J. A Global Characterization of the Translational and Transcriptional Programs Induced by Methionine Restriction through Ribosome Profiling and RNA-Seq. BMC Genom. 2017, 18, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walvekar, A.S.; Srinivasan, R.; Gupta, R.; Laxman, S. Methionine Coordinates a Hierarchically Organized Anabolic Program Enabling Proliferation. Mol. Biol. Cell 2018, 29, 3183–3200. [Google Scholar] [CrossRef]
- Wiame, J.M.; Grenson, M.; Arst, H.N. Nitrogen Catabolite Repression in Yeasts and Filamentous Fungi. Adv. Microb. Physiol. 1985, 26, 1–88. [Google Scholar] [CrossRef]
- Thomas, D.; Surdin-Kerjan, Y. Metabolism of Sulfur Amino Acids in Saccharomyces cerevisiae. Microbiol Mol. Biol. Rev. 1997, 61, 503–532. [Google Scholar] [CrossRef]
- Cherest, H.; Eichler, F.; de Robichon-Szulmajster, H. Genetic and Regulatory Aspects of Methionine Biosynthesis in Saccharomyces cerevisiae. J. Bacteriol. 1969, 97, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Linder, T. ATP Sulfurylase Is Essential for the Utilization of Sulfamate as a Sulfur Source in the Yeast Komagataella pastoris (Syn. Pichia pastoris). Curr. Microbiol. 2017, 74, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.; Barbey, R.; Surdin-Kerjan, Y. Gene-Enzyme Relationship in the Sulfate Assimilation Pathway of Saccharomyces cerevisiae. Study of the 3’-Phosphoadenylylsulfate Reductase Structural Gene. J. Biol. Chem. 1990, 265, 15518–15524. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Kajihata, S.; Matsuda, F.; Shimizu, H. (13)C-Metabolic Flux Analysis in S-Adenosyl-L-Methionine Production by Saccharomyces cerevisiae. J. Biosci. Bioeng. 2015, 120, 532–538. [Google Scholar] [CrossRef]
- Stoner, G.L.; Eisenberg, M.A. Biosynthesis of 7, 8-Diaminopelargonic Acid from 7-Keto-8-Aminopelargonic Acid and S-Adenosyl-L-Methionine. The Kinetics of the Reaction. J. Biol. Chem. 1975, 250, 4037–4043. [Google Scholar] [CrossRef] [PubMed]
- Slany, R.K.; Bösl, M.; Crain, P.F.; Kersten, H. A New Function of S-Adenosylmethionine: The Ribosyl Moiety of AdoMet Is the Precursor of the Cyclopentenediol Moiety of the TRNA Wobble Base Queuine. Biochemistry 1993, 32, 7811–7817. [Google Scholar] [CrossRef]
- Gupta, R.; Laxman, S. TRNA Wobble-Uridine Modifications as Amino Acid Sensors and Regulators of Cellular Metabolic State. Curr. Genet. 2020, 66, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Wu, S.; Letchworth, G.J. High efficiency transformation by electroporation of Pichia pastoris pretreated with lithium acetate and dithiothreitol. BioTechniques 2004, 36, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Padkina, M.V.; Krasnopevtseva, N.G.; Petrashen, M.G. Genetic and biochemical studies of acid phosphatases of Saccharomyces cerevisiae. Properties of acid phosphatases from different strains (Russian). Genetika 1974, 10, 100–110. [Google Scholar]
- Leggett, R.M.; Ramirez-Gonzalez, R.H.; Clavijo, B.J.; Ewaite, D.; Davey, R.P. Sequencing quality assessment tools to enable data-driven informatics for high throughput genomics. Front. Genet. 2013, 4, 288. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2212. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 28 February 2023).
- Vogl, T.; Sturmberger, L.; Kickenweiz, T.; Wasmayer, R.; Schmid, C.; Hatzl, A.M.; Gerstmann, M.A.; Pitzer, J.; Wagner, M.; Thallinger, G.G.; et al. A Toolbox of Diverse Promoters Related to Methanol Utilization: Functionally Verified Parts for Heterologous Pathway Expression in Pichia pastoris. ACS Synth. Biol. 2016, 5, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Dey, T.; Krishna Rao, K.; Khatun, J.; Rangarajan, P.N. The Nuclear Transcription Factor Rtg1p Functions as a Cytosolic, Post-Transcriptional Regulator in the Methylotrophic Yeast Pichia pastoris. J. Biol. Chem. 2018, 293, 16647–16660. [Google Scholar] [CrossRef] [Green Version]
- Solà, A.; Maaheimo, H.; Ylönen, K.; Ferrer, P.; Szyperski, T. Amino Acid Biosynthesis and Metabolic Flux Profiling of Pichia pastoris. Eur. J. Biochem. 2004, 271, 2462–2470. [Google Scholar] [CrossRef]
- Ferrer, P.; Albiol, J. 13C-Based Metabolic Flux Analysis of Recombinant Pichia pastoris. Methods Mol. Biol. 2014, 1191, 291–313. [Google Scholar] [CrossRef]
- Chan, S.Y.; Appling, D.R. Regulation of S-Adenosylmethionine Levels in Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 43051–43059. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhu, Z.; Nielsen, J.; Siewers, V. Engineering Saccharomyces cerevisiae Cells for Production of Fatty Acid-Derived Biofuels and Chemicals. Open. Biol. 2019, 9, 190049. [Google Scholar] [CrossRef] [Green Version]
- Tehlivets, O.; Scheuringer, K.; Kohlwein, S.D. Fatty Acid Synthesis and Elongation in Yeast. Biochim. Biophys. Acta 2007, 1771, 255–270. [Google Scholar] [CrossRef]
- Johansson, P.; Wiltschi, B.; Kumari, P.; Kessler, B.; Vonrhein, C.; Vonck, J.; Oesterhelt, D.; Grininger, M. Inhibition of the fungal fatty acid synthase type I multienzyme complex. Proc. Natl. Acad. Sci. USA 2008, 105, 12803–12808. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Sutter, B.M.; Wang, Y.; Kuang, Z.; Tu, B.P. A Metabolic Function for Phospholipid and Histone Methylation. Mol. Cell 2017, 66, 180–193.e8. [Google Scholar] [CrossRef] [Green Version]
- Câmara, A.d.A.; Maréchal, P.-A.; Tourdot-Maréchal, R.; Husson, F. Oxidative Stress Resistance during Dehydration of Three Non-Saccharomyces Wine Yeast Strains. Food Res. Int. 2019, 123, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Carman, G.M.; Han, G.-S. Regulation of Phospholipid Synthesis in the Yeast Saccharomyces cerevisiae. Annu. Rev. Biochem. 2011, 80, 859–883. [Google Scholar] [CrossRef] [PubMed]
- Sadhu, M.J.; Moresco, J.J.; Zimmer, A.D.; Yates, J.R.; Rine, J. Multiple Inputs Control Sulfur-Containing Amino Acid Synthesis in Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 1653–1665. [Google Scholar] [CrossRef]
- Shi, L.; Wang, X.; Wang, J.; Zhang, P.; Qi, F.; Cai, M.; Zhang, Y.; Zhou, X. Transcriptome analysis of ∆mig1∆mig2 mutant reveals their roles in methanol catabolism, peroxisome biogenesis and autophagy in methylotrophic yeast Pichia pastoris. Genes Genom. 2018, 40, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.T.; Khalawan, S.A.; Curran, B.P. Subtle Alterations in Growth Medium Composition Can Dramatically Alter the Percentage of Unsaturated Fatty Acids in the Yeast Saccharomyces cerevisiae. Yeast 2001, 18, 81–88. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, Y.; Miao, H.; Tang, X.; Xu, B.; Wu, Q.; Mu, Y.; Huang, Z. Transcriptomic Analysis of Pichia pastoris (Komagataella phaffii) GS115 During Heterologous Protein Production Using a High-Cell-Density Fed-Batch Cultivation Strategy. Front. Microbiol. 2020, 11, 463. [Google Scholar] [CrossRef]
- Shirra, M.K.; Patton-Vogt, J.; Ulrich, A.; Liuta-Tehlivets, O.; Kohlwein, S.D.; Henry, S.A.; Arndt, K.M. Inhibition of Acetyl Coenzyme A Carboxylase Activity Restores Expression of the INO1 Gene in a Snf1 Mutant Strain of Saccharomyces cerevisiae. Mol. Cell Biol. 2001, 21, 5710–5722. [Google Scholar] [CrossRef] [Green Version]
- Mertz, J.E.; Spence, K.D. Methionine Adenosyltransferase and Ethionine Resistance in Saccharomyces cerevisiae. J. Bacteriol. 1972, 111, 778–783. [Google Scholar] [CrossRef] [Green Version]
- Farfán, M.J.; Aparicio, L.; Calderón, I.L. Threonine Overproduction in Yeast Strains Carrying the HOM3-R2 Mutant Allele under the Control of Different Inducible Promoters. Appl. Environ. Microbiol. 1999, 65, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Schotte, P.; Dewerte, I.; De Groeve, M.; De Keyser, S.; De Brabandere, V.; Stanssens, P. Pichia pastoris MutS Strains Are Prone to Misincorporation of O-Methyl-l-Homoserine at Methionine Residues When Methanol Is Used as the Sole Carbon Source. Microb. Cell Fact. 2016, 15, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ianshina, T.; Sidorin, A.; Petrova, K.; Shubert, M.; Makeeva, A.; Sambuk, E.; Govdi, A.; Rumyantsev, A.; Padkina, M. Effect of Methionine on Gene Expression in Komagataella phaffii Cells. Microorganisms 2023, 11, 877. https://doi.org/10.3390/microorganisms11040877
Ianshina T, Sidorin A, Petrova K, Shubert M, Makeeva A, Sambuk E, Govdi A, Rumyantsev A, Padkina M. Effect of Methionine on Gene Expression in Komagataella phaffii Cells. Microorganisms. 2023; 11(4):877. https://doi.org/10.3390/microorganisms11040877
Chicago/Turabian StyleIanshina, Tatiana, Anton Sidorin, Kristina Petrova, Maria Shubert, Anastasiya Makeeva, Elena Sambuk, Anastasiya Govdi, Andrey Rumyantsev, and Marina Padkina. 2023. "Effect of Methionine on Gene Expression in Komagataella phaffii Cells" Microorganisms 11, no. 4: 877. https://doi.org/10.3390/microorganisms11040877
APA StyleIanshina, T., Sidorin, A., Petrova, K., Shubert, M., Makeeva, A., Sambuk, E., Govdi, A., Rumyantsev, A., & Padkina, M. (2023). Effect of Methionine on Gene Expression in Komagataella phaffii Cells. Microorganisms, 11(4), 877. https://doi.org/10.3390/microorganisms11040877