

Features of Engorgement of Ixodes ricinus Ticks Infesting the Northern White-Breasted Hedgehog in an Urban Park

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Statistical Analysis
3. Results and Discussion
3.1. Collected Hedgehogs and Ticks
3.2. Abundance of Ticks
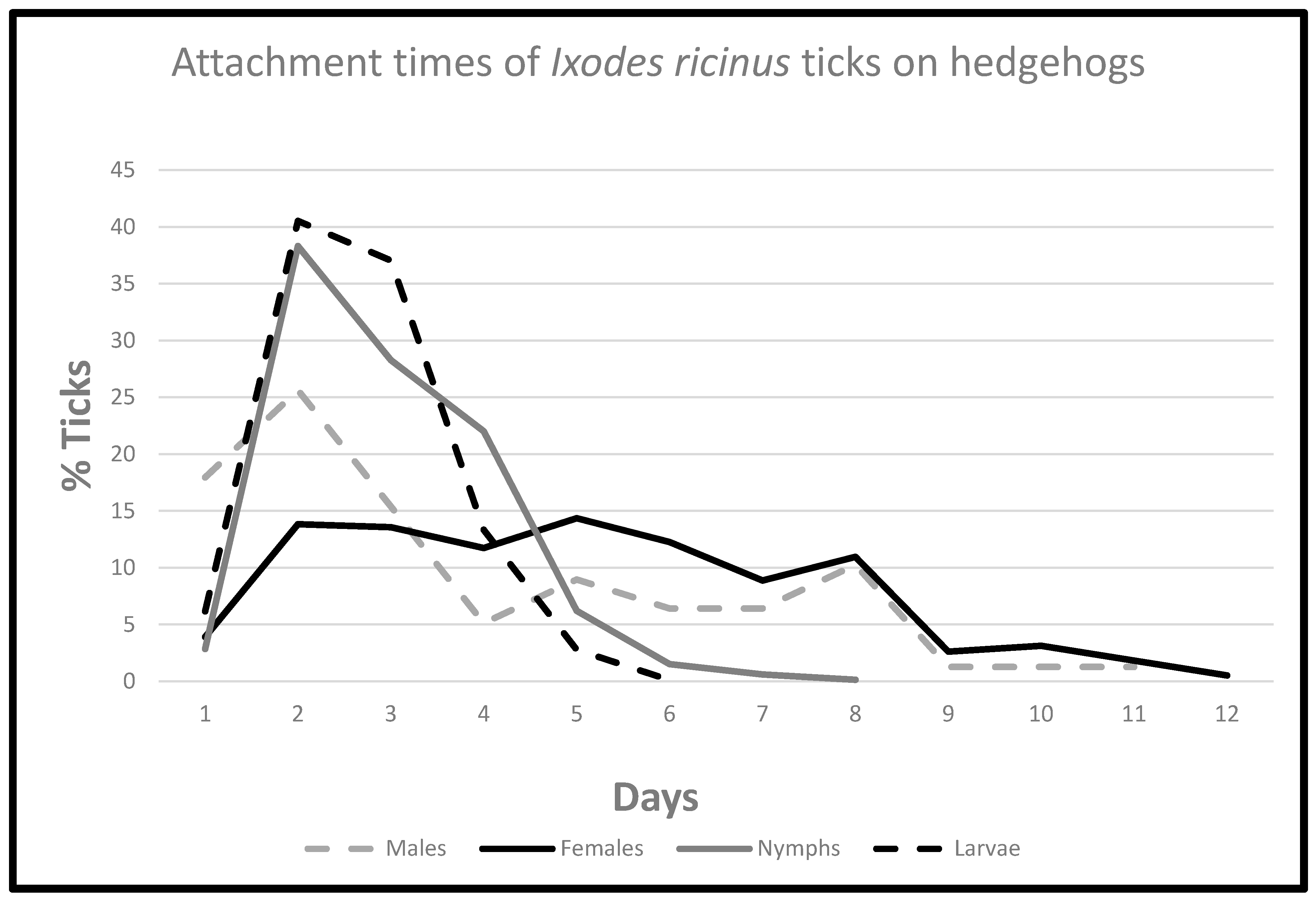
3.3. Time of Engorgement
3.4. Ticks Detached on the First Day
3.5. Sensitivity Analysis
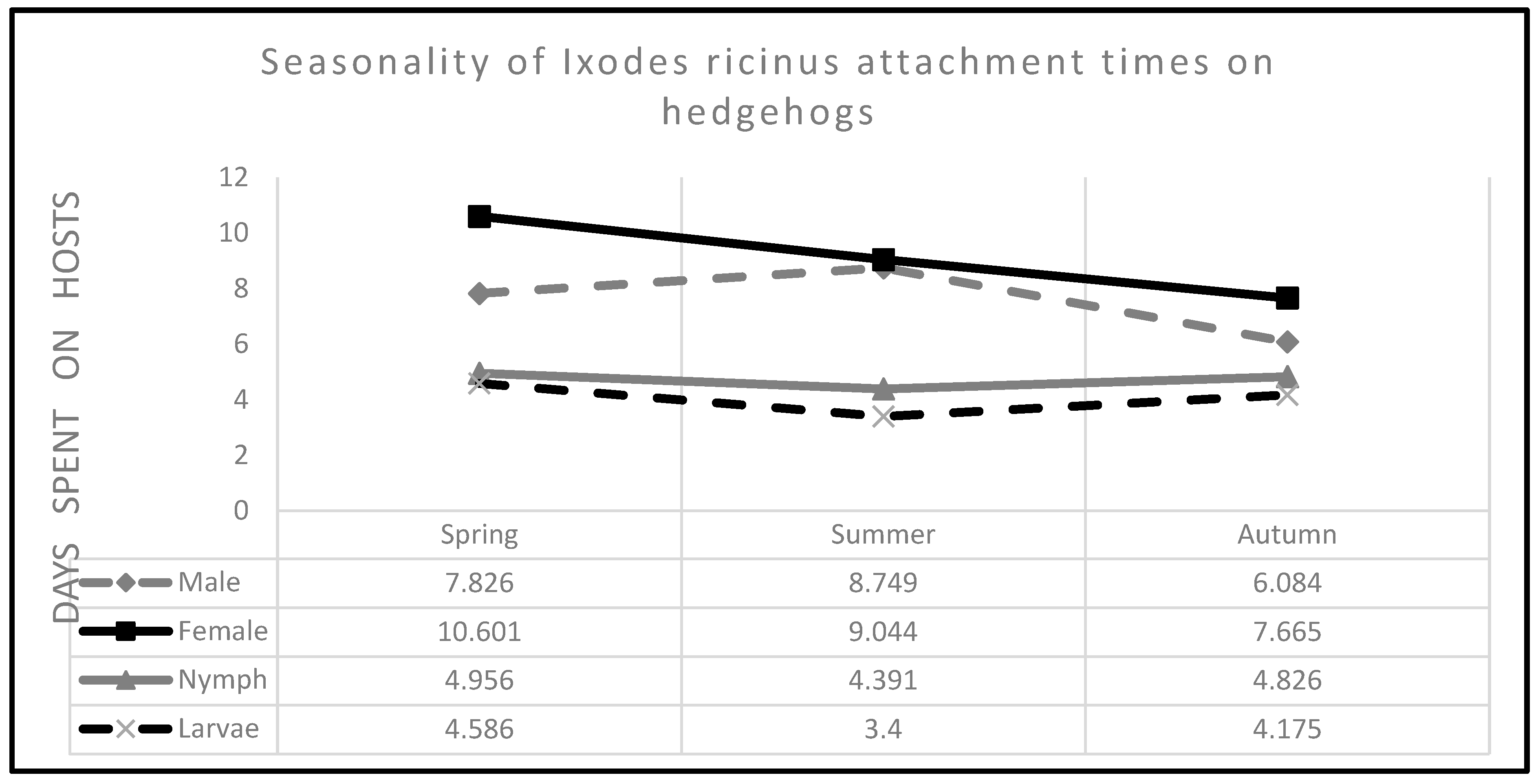
3.6. Seasonality of the Tick Infestation
3.7. Juvenile Hedgehogs
3.8. Additional Observations
3.9. Effect of Weather
3.10. Zoonotic Potential of This Parasite/Host System
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix B
Appendix C
References
- Zöldi, V.; Reicziegel, J.; Egyed, L. Monitoring the diel activity of Ixodes ricinus ticks in Hungary over three seasons. Exp. Appl. Acarol. 2013, 61, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Gern, L.; Rouvinez, E.; Toutoungi, L.N.; Godfroid, E. Transmission cycles of Borrelia burgdorferi sensu lato involving Ixodes ricinus and/or I. hexagonus ticks and the European hedgehog, Erinaceus europaeus, in suburban and urban areas in Switzerland. Folia Parasitol. 1997, 44, 309–314. [Google Scholar]
- Pfäffle, M.; Petney, T.; Skuballa, J.; Taraschewski, H. Comparative population dynamics of a generalist (Ixodes ricinus) and specialist tick (I. hexagonus) species from European hedgehogs. Exp. Appl. Acarol. 2011, 54, 151–164. [Google Scholar] [CrossRef]
- Földvári, G.; Rigó, K.; Jablonszky, M.; Biró, N.; Majoros, G.; Molnár, V.; Tóth, M. Ticks and the city: Ectoparasites of the Northern white-breasted hedgehog (Erinaceus roumanicus) in an urban park. Ticks Tick Borne Dis. 2011, 2, 231–234. [Google Scholar] [CrossRef] [PubMed]
- South, K.E.; Haynes, K.; Jackson, A.C. Hibernation patterns of the European hedgehog, Erinaceus europaeus, at a Cornish rescue centre. Animals 2020, 10, 1418. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: www.R-project.org/ (accessed on 23 March 2023).
- Keiding, N.; Moeschberger, M. Independent delayed entry. In Survival Analysis: State of the Art; Springer: Dordrecht, The Netherlands, 1992; pp. 309–326. [Google Scholar]
- Keiding, N.; Fine, J.P.; Hansen, O.H.; Slama, R. Accelerated failure time regression for backward recurrence times and current durations. Stat. Probab. Lett. 2011, 81, 724–729. [Google Scholar] [CrossRef] [Green Version]
- van Es, B.; Klaassen, C.A.; Oudshoorn, K. Survival analysis under cross-sectional sampling: Length bias and multiplicative censoring. J. Stat. Plan. Inference 2000, 91, 295–312. [Google Scholar] [CrossRef]
- Kalbfleisch, J.; Prentice, R. The Statistical Analysis of Failure Time Data, 2nd ed.; Wiley: New York, NY, USA, 2002. [Google Scholar]
- Stacy, E.W. A generalization of the gamma distribution. Ann. Math. Stat. 1962, 33, 1187–1192. [Google Scholar] [CrossRef]
- Yamaguchi, K. Accelerated failure-time mover-stayer regression models for the analysis of last-episode data. Soc. Methodol. 2003, 33, 81–110. [Google Scholar] [CrossRef]
- Liang, K.Y.; Zeger, S.L. Longitudinal data analysis using generalized linear models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.2-10. 2021. Available online: CRAN.R-project.org/package=survival (accessed on 23 March 2023).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R package geepack for generalized estimating equations. J. Stat. Softw. 2006, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5.3. 2020. Available online: CRAN.R-project.org/package=emmeans (accessed on 23 March 2023).
- Süss, J. Epidemiology and ecology of TBE relevant to the production of effective vaccines. Vaccine 2003, 21, S19–S35. [Google Scholar] [CrossRef]
- Gray, J.S. The development and seasonal activity of the tick Ixodes ricinus: A vector of Lyme borreliosis. Rev. Med. Vet. Entomol. 1991, 79, 323–333. [Google Scholar]
- Széll, Z.; Sréter-Lancz, Z.; Márialigeti, K.; Sréter, T. Temporal distribution of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna in Hungary. Vet. Parasitol. 2006, 141, 377–379. [Google Scholar] [CrossRef]
- Egyed, L.; Élő, P.; Sréter-Lancz, Z.; Széll, Z.; Balogh, Z.; Sréter, T. Seasonal activity and tick-borne pathogen infection rates of Ixodes ricinus ticks in Hungary. Ticks Tick Borne Dis. 2012, 3, 90–94. [Google Scholar] [CrossRef]
- Tälleklint, L.; Jaenson, T.G. Infestation of mammals by Ixodes ricinus ticks (Acari: Ixodidae) in south-central Sweden. Exp. Appl. Acarol. 1997, 21, 755–771. [Google Scholar] [CrossRef]
- Zöldi, V.; Papp, T.; Rigó, K.; Farkas, J.; Egyed, L. A four-year-long study of a natural tick-borne encephalitis virus focus in Hungary 2010–2013. Ecohealth 2015, 12, 174–182. [Google Scholar] [CrossRef]
- Alasmari, S.; Wall, R. Determining the total energy budget of the tick Ixodes ricinus. Exp. Appl. Acarol. 2020, 80, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Sixl, W.; Kock, M.; Withalm, H.; Stunzner, D. Serological investigations of the hedgehog (Erinaceus europaeus) in Styria. 2. Report. Geogr. Med. Suppl. 1989, 2, 105–108. [Google Scholar] [PubMed]
- Riley, Y.P.; Chomel, B.B. Hedgehog zoonoses. Emerg. Infect. Dis. 2005, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Babudieri, B.; Farina, R. The Leptospirae of the Italian hedgehog. Pathol. Microbiol. 1964, 27, 103–116. [Google Scholar] [CrossRef]
- Földvári, G.; Jahfari, S.; Rigó, K.; Jablonszky, M.; Szekeres, S.; Majoros, G.; Tóth, M.; Molnár, V.; Coipan, E.C.; Sprong, H. Candidatus Neoehrlichia mikurensis and Anaplasma phagocytophilum in urban hedgehogs. Emerg. Infect. Dis. 2014, 20, 496–498. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, A.D.; Thomas, W.E. Paramyxoviruses of the morbilli group in the wild hedgehog Erinaceus europaeus. Br. J. Exp. Pathol. 1981, 62, 79–86. [Google Scholar] [PubMed]
- Faragó, Z. Rabid hedgehog in inner-city area of Budapest. Orv. Hetil. 1997, 138, 2231–2232. [Google Scholar]
- McLauchlan, J.D.; Henderson, W.M. The occurrence of foot-and-mouth disease in the hedgehog under natural conditions. Epidemiol. Infect. 1947, 45, 474–479. [Google Scholar] [CrossRef] [Green Version]
- Kozuch, O.; Nosek, J.; Ernek, E.; Lichard, M.; Albrecht, P. Persistence of tick-borne encephalitis virus in hibernating hedgehogs and dormice. Acta Virol. 1963, 7, 430–433. [Google Scholar]
- Schönbächler, K.; Hatt, J.; Silaghi, C.; Merz, N.; Fraefel, C.; Bachofen, C. Confirmation of Tick-borne encephalitis virus in an European hedgehog (Erinaceus europaeus). Schweiz. Arch. Für Tierheilkd. 2019, 161, 23–31. [Google Scholar] [CrossRef]
- Skuballa, J.; Oehme, R.; Hartelt, K.; Petney, T.; Bucher, T.; Kimmig, P.; Taraschewski, H. European hedgehogs as hosts for Borrelia spp. Germany. Emerg. Infect. Dis. 2007, 13, 952–953. [Google Scholar] [CrossRef]
- Quentin, J.C.; Seguignes, M. Life cycle of Gongylonema mucronatum Seurat, 1916, parasite of the African hedge-hog (author’s transl). Ann. Parasitol. Hum. Comp. 1979, 54, 637–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sormunen, J.J.; Kulha, N.; Klemola, T.; Mäkelä, S.; Vesilahti, E.M.; Vesterinen, E.J. Enhanced threat of tick-borne infections within cities? Assessing public health risks due to ticks in urban green spaces in Helsinki, Finland. Zoonoses Public Health 2020, 67, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Makenov, M.; Karan, L.; Shashina, N.; Akhmetshina, M.; Zhurenkova, O.; Kholodilov, I.; Karganova, G.; Smirnova, N.; Grigoreva, Y.; Yankovskaya, Y.; et al. First detection of tick-borne encephalitis virus in Ixodes ricinus ticks and their rodent hosts in Moscow, Russia. Ticks Tick Borne Dis. 2019, 10, 101265. [Google Scholar] [CrossRef] [PubMed]
- Egyed, L.; Nagy, A.; Lakos, A.; Zöldi, V.; Lang, Z. Tick-borne encephalitis epidemic in Hungary 1951–2021: The story and lessons learned. Zoonoses Public Health 2023, 70, 81–92. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Hedgehogs | Ixodes ricinus | |||||||
|---|---|---|---|---|---|---|---|---|
| M | F | All | M | F | N | L | All | |
| Spring | 7 | 11 | 18 | 49 | 213 | 1600 | 300 | 2162 |
| Summer | 11 | 16 | 27 | 13 | 95 | 804 | 1058 | 1970 |
| Autumn | 7 | 5 | 12 | 13 | 67 | 203 | 299 | 582 |
| Total | 25 | 32 | 57 | 75 | 375 | 2607 | 1657 | 4714 |
| % | 44 | 56 | 100 | 1.6 | 8 | 55.3 | 35.1 | 100 |
| Ticks | GEE Modelled Tick Number | Observed Mean Tick Number per Host | |||||
|---|---|---|---|---|---|---|---|
| Tick No. | 95% CI | Tick No. | SD | Tick Abundance | |||
| Spring | |||||||
| Males | 2.78 | 1.26 | 6.14 | 2.78 | 3.44 | 50 | |
| Females | 12.00 | 4.57 | 31.49 | 12.00 | 18.08 | 216 | |
| Nymphs | 86.22 | 53.78 | 138.22 | 86.22 | 63.55 | 1552 | |
| Larvae | 16.89 | 7.85 | 36.35 | 16.89 | 20.22 | 304 | |
| Summer | |||||||
| Males | 0.52 | 0.21 | 1.29 | 0.52 | 0.89 | 14 | |
| Females | 3.81 | 2.01 | 7.23 | 3.81 | 4.62 | 103 | |
| Nymphs | 31.96 | 15.45 | 66.12 | 31.96 | 44.01 | 863 | |
| Larvae | 43.85 | 25.41 | 75.66 | 43.85 | 45.31 | 1184 | |
| Autumn | |||||||
| Males | 1.25 | 0.56 | 2.77 | 1.25 | 1.29 | 15 | |
| Females | 5.08 | 2.80 | 9.22 | 5.08 | 3.92 | 61 | |
| Nymphs | 12.33 | 6.93 | 21.94 | 12.33 | 9.20 | 148 | |
| Larvae | 15.08 | 6.45 | 35.29 | 15.08 | 16.59 | 181 | |
| Seasons | Tick Stages | Estimate | Lower | Upper | p Values |
|---|---|---|---|---|---|
| Spring | Female/Male | 4.32 | 3.21 | 5.82 | <0.001 |
| Spring | Larva/Male | 6.08 | 2.54 | 14.55 | <0.001 |
| Spring | Nymph/Male | 31.04 | 14.55 | 66.22 | <0.001 |
| S | Female/Larva | 0.71 | 0.26 | 1.91 | 0.790 |
| Spring | Nymph/Larva | 5.11 | 2.28 | 11.45 | <0.001 |
| Spring | Female/nymph | 0.14 | 0.05 | 0.35 | <0.001 |
| Summer | Female/Male | 7.36 | 4.03 | 13.43 | <0.001 |
| Summer | Larva/Male | 84.57 | 33.30 | 214.80 | <0.001 |
| Summer | Nymph/Male | 61.64 | 32.90 | 115.49 | <0.001 |
| Summer | Female/Larva | 0.09 | 0.04 | 0.18 | <0.001 |
| Summer | Nymph/Larva | 0.73 | 0.35 | 1.52 | 0.680 |
| Summer | Female/nymph | 0.12 | 0.06 | 0.23 | <0.001 |
| Autumn | Female/Male | 4.07 | 2.35 | 7.03 | <0.001 |
| Autumn | Larva/Male | 12.07 | 9.22 | 15.79 | <0.001 |
| Autumn | Nymph/Male | 9.87 | 5.81 | 16.75 | <0.001 |
| Autumn | Female/Larva | 0.34 | 0.19 | 0.61 | <0.001 |
| Autumn | Nymph/Larva | 0.82 | 0.43 | 1.55 | 0.830 |
| Autumn | Female/nymph | 0.41 | 0.30 | 0.57 | <0.001 |
| Tick stages | |||||
| Males | Summer/Spring | 0.19 | 0.07 | 0.51 | <0.001 |
| Males | Autumn/Spring | 0.45 | 0.18 | 1.15 | 0.115 |
| Males | Autumn/Summer | 2.41 | 0.88 | 6.64 | 0.104 |
| Females | Summer/Spring | 0.32 | 0.12 | 0.84 | 0.015 |
| Females | Autumn/Spring | 0.42 | 0.16 | 1.09 | 0.085 |
| Females | Autumn/Summer | 1.33 | 0.64 | 2.76 | 0.625 |
| Nymphs | Summer/Spring | 0.37 | 0.18 | 0.77 | 0.004 |
| Nymphs | Autumn/Spring | 0.14 | 0.08 | 0.27 | <0.001 |
| Nymphs | Autumn/Summer | 0.39 | 0.18 | 0.84 | 0.011 |
| Larvae | Summer/Spring | 2.60 | 1.18 | 5.71 | 0.013 |
| Larvae | Autumn/Spring | 0.89 | 0.34 | 2.33 | 0.959 |
| Larvae | Autumn/Summer | 0.34 | 0.15 | 0.80 | 0.009 |
| Detached Engorged Ticks (Days) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| Males | 14 | 21 | 12 | 4 | 7 | 6 | 5 | 7 | 1 | 1 | 1 | ||||
| Females | 17 * | 55 | 47 | 46 | 52 | 47 | 41 | 31 | 15 | 18 | 2 | 2 | 4 | 2 | 1 |
| Nymphs | 64 * | 913 | 726 | 618 | 171 | 42 | 18 | 8 | 3 | ||||||
| Larvae | 105 * | 732 | 561 | 217 | 48 | 6 | |||||||||
| Ticks | Season | Modelled Attachment Times | Experienced Attachment Times | ||||
|---|---|---|---|---|---|---|---|
| Days | 95% CI | Days | SD | Tick Abundance | |||
| Spring | |||||||
| Males | 7.826 | 6.486 | 9.442 | 3.42 | 2.625 | 50 | |
| Females | 10.601 | 9.591 | 11.718 | 5.19 | 2.943 | 216 | |
| Nymphs | 4.956 | 4.567 | 5.378 | 2.512 | 1.257 | 1552 | |
| Larvae | 4.586 | 3.977 | 5.263 | 2.717 | 1.059 | 304 | |
| Summer | |||||||
| Males | 8.749 | 7.005 | 10.926 | 3.071 | 3.131 | 14 | |
| Females | 9.044 | 8.281 | 9.877 | 4.267 | 2.72 | 103 | |
| Nymphs | 4.391 | 4.190 | 4.601 | 2.638 | 0.941 | 1184 | |
| Larvae | 3.40 | 3.207 | 3.605 | 1.933 | 0.784 | 863 | |
| Autumn | |||||||
| Males | 6.084 | 5.17 | 7.16 | 3.10 | 1.882 | 15 | |
| Females | 7.665 | 6.90 | 8.515 | 3.992 | 2.18 | 61 | |
| Nymphs | 4.826 | 4.406 | 5.286 | 2.75 | 1.183 | 148 | |
| Larvae | 4.175 | 3.855 | 4.522 | 2.467 | 0.924 | 181 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egyed, L.; Nagy, D.; Lang, Z. Features of Engorgement of Ixodes ricinus Ticks Infesting the Northern White-Breasted Hedgehog in an Urban Park. Microorganisms 2023, 11, 881. https://doi.org/10.3390/microorganisms11040881
Egyed L, Nagy D, Lang Z. Features of Engorgement of Ixodes ricinus Ticks Infesting the Northern White-Breasted Hedgehog in an Urban Park. Microorganisms. 2023; 11(4):881. https://doi.org/10.3390/microorganisms11040881
Chicago/Turabian StyleEgyed, László, Dávidné Nagy, and Zsolt Lang. 2023. "Features of Engorgement of Ixodes ricinus Ticks Infesting the Northern White-Breasted Hedgehog in an Urban Park" Microorganisms 11, no. 4: 881. https://doi.org/10.3390/microorganisms11040881
APA StyleEgyed, L., Nagy, D., & Lang, Z. (2023). Features of Engorgement of Ixodes ricinus Ticks Infesting the Northern White-Breasted Hedgehog in an Urban Park. Microorganisms, 11(4), 881. https://doi.org/10.3390/microorganisms11040881