
A Review of the Bacterial Phosphoproteomes of Beneficial Microbes


Abstract
:1. Introduction
2. Bacterial Protein Phosphorylation
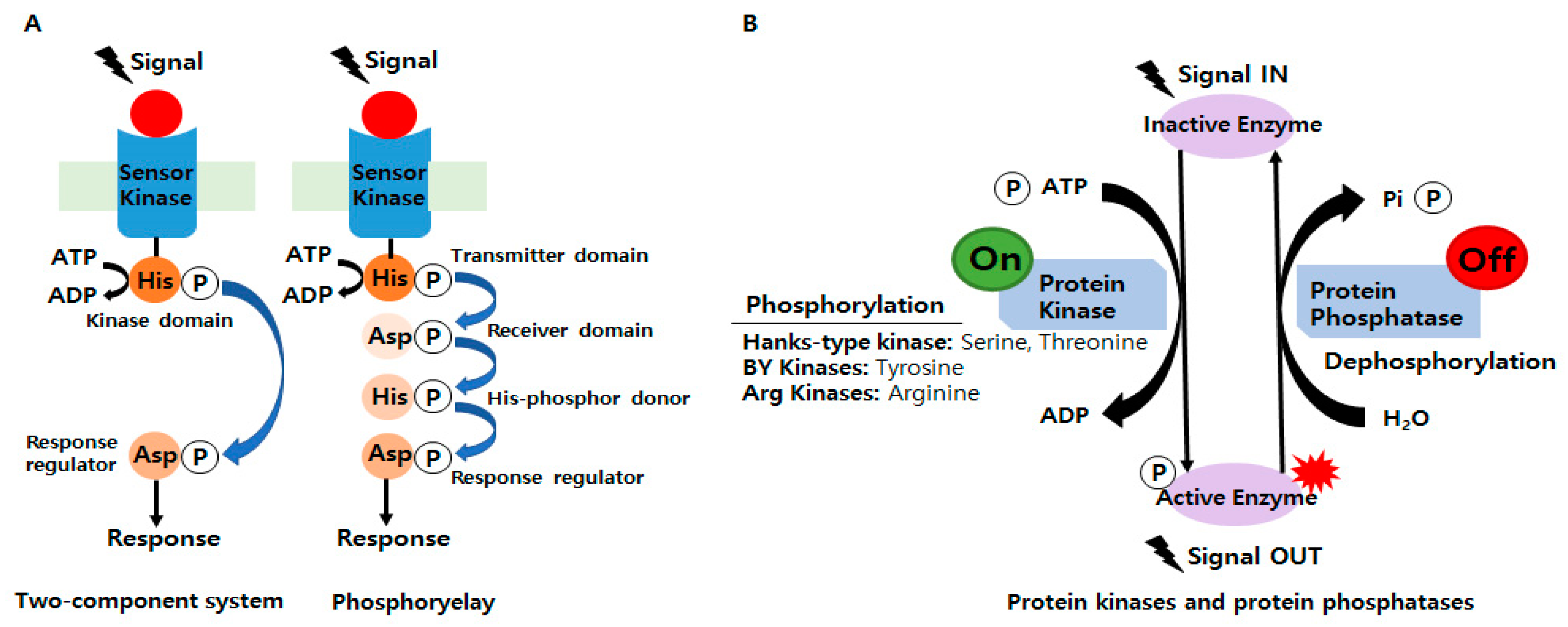
2.1. Two-Component Systems (TCSs)
2.2. Involvements of Ser/Thr/Tyr/Arg Kinases in Bacterial Signaling and Regulation
3. Bacterial Phosphoproteomics
3.1. Phosphoproteome Analysis of Beneficial Microorganisms
3.2. Phylogenetic Diagram of Beneficial Microorganisms
4. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Thingholm, T.E.; Jensen, O.N.; Larsen, M.R. Analytical strategies for phosphoproteomics. Proteomics 2009, 9, 1451–1468. [Google Scholar] [CrossRef] [PubMed]
- Chubukov, V.; Gerosa, L.; Kochanowski, K.; Sauer, U. Coordination of microbial metabolism. Nat. Rev. Microbiol. 2014, 12, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Macek, B.; Mijakovic, I.; Olsen, J.V.; Gnad, F.; Kumar, C.; Jensen, P.R.; Mann, M. The serine/threonine/tyrosine phosphoproteome of the model bacterium Bacillus subtilis. Mol. Cell. Proteom. 2007, 6, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macek, B.; Gnad, F.; Soufi, B.; Kumar, C.; Olsen, J.V.; Mijakovic, I.; Mann, M. Phosphoproteome Analysis of E. coli Reveals Evolutionary Conservation of Bacterial Ser/Thr/Tyr Phosphorylation. Mol. Cell. Proteom. 2008, 7, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Macek, B.; Forchhammer, K.; Hardouin, J.; Weber-Ban, E.; Grangeasse, C.; Mijakovic, I. Protein post-translational modifications in bacteria. Nat. Rev. Microbiol. 2019, 17, 651–664. [Google Scholar] [CrossRef]
- Ramazi, S.; Zahiri, J. Posttranslational modifications in proteins: Resources, tools and prediction methods. Database J. Biol. Databases Curation 2021, 2021, baab012. [Google Scholar] [CrossRef]
- Mijakovic, I.; Grangeasse, C.; Turgay, K. Exploring the diversity of protein modifications: Special bacterial phosphorylation systems. FEMS Microbiol. Rev. 2016, 40, 398–417. [Google Scholar] [CrossRef] [Green Version]
- Bourret, R.B.; Silversmith, R.E. Two-component signal transduction. Curr. Opin. Microbiol. 2010, 13, 113–115. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Jamal, S.B.; Hassan, S.S.; Carvalho, P.; Almeida, S.; Barh, D.; Ghosh, P.; Silva, A.; Castro, T.L.P.; Azevedo, V. Two-Component Signal Transduction Systems of Pathogenic Bacteria As Targets for Antimicrobial Therapy: An Overview. Front. Microbiol. 2017, 8, 1878. [Google Scholar] [CrossRef] [Green Version]
- Mitrophanov, A.Y.; Groisman, E.A. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008, 22, 2601–2611. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, K.; Dworkin, J.; Nagle, E. Identification and Biochemical Characterization of a Novel Protein Phosphatase 2C-Like Ser/Thr Phosphatase in Escherichia coli. J. Bacteriol. 2018, 200, e00225-18, Erratum in J. Bacteriol. 2019, 201, e00648-19. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, H.; Kurushima, J.; Hashimoto, Y.; Tomita, H. Progress Overview of Bacterial Two-Component Regulatory Systems as Potential Targets for Antimicrobial Chemotherapy. Antibiotics 2020, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- van Hoek, M.L.; Hoang, K.V.; Gunn, J.S. Two-Component Systems in Francisella Species. Front. Cell Infect. Microbiol. 2019, 9, 198. [Google Scholar] [CrossRef]
- Yuan, J.; Jin, F.; Glatter, T.; Sourjik, V. Osmosensing by the bacterial PhoQ/PhoP two-component system. Proc. Natl. Acad. Sci. USA 2017, 114, E10792–E10798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuhiro Gotoh, Y.E.; Watanabe, T.; Okamoto, S.; Doi, A.; Utsumi, R. Two-component signal transduction as potential drug targets in pathogenic bacteria. Curr. Opin. Microbiol. 2010, 13, 232–239. [Google Scholar] [CrossRef]
- Huang, J.; Li, C.; Song, J.; Velkov, T.; Wang, L.; Zhu, Y.; Li, J. Regulating polymyxin resistance in Gram-negative bacteria: Roles of two-component systems PhoPQ and PmrAB. Future Microbiol. 2020, 15, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Lingzhi, L.; Haojie, G.; Dan, G.; Hongmei, M.; Yang, L.; Mengdie, J.; Chengkun, Z.; Xiaohui, Z. The role of two-component regulatory system in beta-lactam antibiotics resistance. Microbiol. Res. 2018, 215, 126–129. [Google Scholar] [CrossRef]
- Takada, H.; Yoshikawa, H. Essentiality and function of WalK/WalR two-component system: The past, present, and future of research. Biosci. Biotechnol. Biochem. 2018, 82, 741–751. [Google Scholar] [CrossRef]
- Cardona, S.T.; Choy, M.; Hogan, A.M. Essential Two-Component Systems Regulating Cell Envelope Functions: Opportunities for Novel Antibiotic Therapies. J. Membr. Biol. 2018, 251, 75–89. [Google Scholar] [CrossRef]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef] [Green Version]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Murret-Labarthe, C.; Kerhoas, M.; Dufresne, K.; Daigle, F. New Roles for Two-Component System Response Regulators of Salmonella enterica Serovar Typhi during Host Cell Interactions. Microorganisms 2020, 8, 722. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Du, C.; Huang, H.; Gu, H.; Dong, X.; Hu, Y. TCS response regulator OmpR plays a major role in stress resistance, antibiotic resistance, motility, and virulence in Edwardsiella piscicida. Aquaculture 2022, 559, 738441. [Google Scholar] [CrossRef]
- Shaw, C.; Hess, M.; Weimer, B.C. Two-component systems regulate bacterial virulence in response to the host gastrointestinal environment and metabolic cues. Virulence 2022, 13, 1666–1680. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Dhankhar, P.; Kumar, P. Bacterial histidine kinases as potential antibacterial drug targets. In Protein Kinase Inhibitors; Elsevier: Amsterdam, The Netherlands, 2022; pp. 711–734. [Google Scholar]
- Kwiecinski, J.M.; Jelani, D.A.; Fuentes, E.J.; Horswill, A.R. Therapeutic Inhibition of Staphylococcus aureus ArlRS Two-Component Regulatory System Blocks Virulence. Antimicrob. Agents Chemother. 2022, 66, e00187-22. [Google Scholar] [CrossRef]
- Kundu, M. The role of two-component systems in the physiology of Mycobacterium tuberculosis. IUBMB Life 2018, 70, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Schaefers, M.M. Regulation of Virulence by Two-Component Systems in Pathogenic Burkholderia. Infect. Immun. 2020, 88, e00927-19. [Google Scholar] [CrossRef]
- Choudhary, K.S.; Kleinmanns, J.A.; Decker, K.; Sastry, A.V.; Gao, Y.; Szubin, R.; Seif, Y.; Palsson, B.O. Elucidation of Regulatory Modes for Five Two-Component Systems in Escherichia coli Reveals Novel Relationships. mSystems 2020, 5, e00980-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, D.; Zhu, J.; Liu, W. Two-Component Signal Transduction Systems: A Major Strategy for Connecting Input Stimuli to Biofilm Formation. Front. Microbiol. 2018, 9, 3279. [Google Scholar] [CrossRef]
- Badal, D.; Jayarani, A.V.; Kollaran, M.A.; Kumar, A.; Singh, V. Pseudomonas aeruginosa biofilm formation on endotracheal tubes requires multiple two-component systems. J. Med. Microbiol. 2020, 69, 906–919. [Google Scholar] [CrossRef]
- Kera, K.; Yoshizawa, Y.; Shigehara, T.; Nagayama, T.; Tsujii, M.; Tochigi, S.; Uozumi, N. Hik36-Hik43 and Rre6 act as a two-component regulatory system to control cell aggregation in Synechocystis sp. PCC6803. Sci. Rep. 2020, 10, 19405. [Google Scholar] [CrossRef]
- Sionov, R.V.; Steinberg, D. Targeting the Holy Triangle of Quorum Sensing, Biofilm Formation, and Antibiotic Resistance in Pathogenic Bacteria. Microorganisms 2022, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, V.; Tiwari, M.; Joshi, R.; Tiwari, V. Therapeutic strategies against potential antibiofilm targets of multidrug-resistant Acinetobacter baumannii. J. Cell. Physiol. 2022, 237, 2045–2063. [Google Scholar] [CrossRef]
- Jabbour, N.; Morello, E.; Camiade, E.; Lartigue, M.-F. Biofilm Formation in Streptococcus agalactiae Is Inhibited by a Small Regulatory RNA Regulated by the Two-Component System CiaRH. Microbiol. Spectr. 2022, 10, e0063522. [Google Scholar] [CrossRef]
- Ballén, V.; Cepas, V.; Ratia, C.; Gabasa, Y.; Soto, S.M. Clinical Escherichia coli: From Biofilm Formation to New Antibiofilm Strategies. Microorganisms 2022, 10, 1103. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, S.E.; Lamont, R.J. Tyrosine phosphorylation and bacterial virulence. Int. J. Oral Sci. 2012, 4, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzone, A.J. Role of protein phosphorylation on serine/threonine and tyrosine in the virulence of bacterial pathogens. J. Mol. Microbiol. Biotechnol. 2005, 9, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-component signal transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, A.A.; De Lange, L.C.M.; Bachovchin, W.W.; Robillard, G.T. Effect of phosphorylation on hydrogen-bonding interactions of the active site histidine of the phosphocarrier protein HPr of the phosphoenolpyruvate-dependent phosphotransferase system determined by nitrogen-15 NMR spectroscopy. Biochemistry 1990, 29, 8164–8171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reizer, J.; Romano, A.H.; Deutscher, J. The role of phosphorylation of HPr, a phosphocarrier protein of the phosphotransferase system, in the regulation of carbon metabolism in gram-positive bacteria. J. Cell Biochem. 1993, 51, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Mijakovic, I.; Poncet, S.; Galinier, A.; Monedero, V.; Fieulaine, S.; Janin, J.; Nessler, S.; Marquez, J.A.; Scheffzek, K.; Hasenbein, S.; et al. Pyrophosphate-producing protein dephosphorylation by HPr kinase/phosphorylase: A relic of early life? Proc. Natl. Acad. Sci. USA 2002, 99, 13442–13447. [Google Scholar] [CrossRef] [Green Version]
- LaPorte, D.C. The isocitrate dehydrogenase phosphorylation cycle: Regulation and enzymology. J. Cell. Biochem. 1993, 51, 14–18. [Google Scholar] [CrossRef]
- Laporte, D.C.; Stueland, C.S.; Ikeda, T.P. Isocitrate dehydrogenase kinase/phosphatase. Biochimie 1989, 71, 1051–1057. [Google Scholar] [CrossRef]
- Garnak, M.; Reeves, H.C. Phosphorylation of Isocitrate dehydrogenase of Escherichia coli. Science 1979, 203, 1111–1112. [Google Scholar] [CrossRef] [PubMed]
- Stueland, C.S.; Gorden, K.; LaPorte, D.C. The isocitrate dehydrogenase phosphorylation cycle. Identification of the primary rate-limiting step. J. Biol. Chem. 1988, 263, 19475–19479. [Google Scholar] [CrossRef] [PubMed]
- Mijakovic, I. Protein phosphorylation in bacteria. Microbe 2010, 5, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Soufi, B.; Gnad, F.; Jensen, P.R.; Petranovic, D.; Mann, M.; Mijakovic, I.; Macek, B. The Ser/Thr/Tyr phosphoproteome of Lactococcus lactis IL1403 reveals multiply phosphorylated proteins. Proteomics 2008, 8, 3486–3493. [Google Scholar] [CrossRef]
- Chao, J.D.; Wong, D.; Av-Gay, Y. Microbial protein-tyrosine kinases. J. Biol. Chem. 2014, 289, 9463–9472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellinzoni, M.; Wehenkel, A.M.; Duran, R.; Alzari, P.M. Novel mechanistic insights into physiological signaling pathways mediated by mycobacterial Ser/Thr protein kinases. Microbes Infect. 2019, 21, 222–229. [Google Scholar] [CrossRef]
- Getz, L.J.; Runte, C.S.; Rainey, J.K.; Thomas, N.A. Tyrosine Phosphorylation as a Widespread Regulatory Mechanism in Prokaryotes. J. Bacteriol. 2019, 201, e00205-19. [Google Scholar] [CrossRef] [Green Version]
- Nariya, H.; Inouye, S. Identification of a protein Ser/Thr kinase cascade that regulates essential transcriptional activators in Myxococcus xanthus development. Mol. Microbiol. 2005, 58, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.F.; Goss, L.; Dworkin, J. Eukaryote-like serine/threonine kinases and phosphatases in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 192–212. [Google Scholar] [CrossRef] [Green Version]
- Birk, M.S.; Charpentier, E.; Frese, C.K. Automated Phosphopeptide Enrichment for Gram-Positive Bacteria. J. Proteome Res. 2021, 20, 4886–4892. [Google Scholar] [CrossRef]
- Bonne Kohler, J.; Jers, C.; Senissar, M.; Shi, L.; Derouiche, A.; Mijakovic, I. Importance of protein Ser/Thr/Tyr phosphorylation for bacterial pathogenesis. FEBS Lett. 2020, 594, 2339–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.; Haller, L.; Blein-Nicolas, M.; Zivy, M.; Canette, A.; Verbrugghe, M.; Mezange, C.; Boulay, M.; Gardan, R.; Samson, S.; et al. Identification of Hanks-Type Kinase PknB-Specific Targets in the Streptococcus thermophilus Phosphoproteome. Front. Microbiol. 2019, 10, 1329. [Google Scholar] [CrossRef] [PubMed]
- Szoke, T.; Albocher, N.; Govindarajan, S.; Nussbaum-Shochat, A.; Amster-Choder, O. Tyrosine phosphorylation-dependent localization of TmaR that controls activity of a major bacterial sugar regulator by polar sequestration. Proc. Natl. Acad. Sci. USA 2021, 118, e2016017118. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Sammartino, J.C.; Costantino, L.; Gelain, A.; Meneghetti, F.; Villa, S.; Chiarelli, L.R. An Overview on the Potential Antimycobacterial Agents Targeting Serine/Threonine Protein Kinases from Mycobacterium tuberculosis. Curr. Top. Med. Chem. 2019, 19, 646–661. [Google Scholar] [CrossRef]
- Hirschfeld, C.; Gomez-Mejia, A.; Bartel, J.; Hentschker, C.; Rohde, M.; Maass, S.; Hammerschmidt, S.; Becher, D. Proteomic Investigation Uncovers Potential Targets and Target Sites of Pneumococcal Serine-Threonine Kinase StkP and Phosphatase PhpP. Front. Microbiol. 2019, 10, 3101. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Garcia, T.; Poncet, S.; Cuenot, E.; Douché, T.; Gianetto, Q.G.; Peltier, J.; Courtin, P.; Chapot-Chartier, M.-P.; Matondo, M.; Dupuy, B.; et al. Ser/Thr kinase-dependent phosphorylation of the peptidoglycan hydrolase CwlA controls its export and modulates cell division in Clostridioides difficile. bioRxiv 2020, 12, e00519-21. [Google Scholar] [CrossRef]
- Andrews, L.D.; Graham, J.; Snider, M.J.; Fraga, D. Characterization of a novel bacterial arginine kinase from Desulfotalea psychrophila. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 312–319. [Google Scholar] [CrossRef]
- Elsholz, A.K.; Turgay, K.; Michalik, S.; Hessling, B.; Gronau, K.; Oertel, D.; Mäder, U.; Bernhardt, J.; Becher, D.; Hecker, M. Global impact of protein arginine phosphorylation on the physiology of Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2012, 109, 7451–7456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Trentini, D.B.; Spiess, S.; Fuhrmann, J.; Ammerer, G.; Mechtler, K.; Clausen, T. Quantitative phosphoproteomics reveals the role of protein arginine phosphorylation in the bacterial stress response. Mol. Cell. Proteom. 2014, 13, 537–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trentini, D.B.; Suskiewicz, M.J.; Heuck, A.; Kurzbauer, R.; Deszcz, L.; Mechtler, K.; Clausen, T. Arginine phosphorylation marks proteins for degradation by a Clp protease. Nature 2016, 539, 48–53. [Google Scholar] [CrossRef]
- Junker, S.; Maaß, S.; Otto, A.; Hecker, M.; Becher, D.R. Toward the quantitative characterization of arginine phosphorylations in Staphylococcus aureus. J. Proteome Res. 2018, 18, 265–279. [Google Scholar] [PubMed]
- Junker, S.; Maaβ, S.; Otto, A.; Michalik, S.; Morgenroth, F.; Gerth, U.; Hecker, M.; Becher, D. Spectral library based analysis of arginine phosphorylations in Staphylococcus aureus. Mol. Cell. Proteom. 2018, 17, 335–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.; Choi, Y.; Lee, D.; Seo, J.K.; Kee, J.M. Distinct phosphorylation and dephosphorylation dynamics of protein arginine kinases revealed by fluorescent activity probes. Chem. Commun. 2019, 55, 7482–7485. [Google Scholar] [CrossRef]
- Suskiewicz, M.J.; Hajdusits, B.; Beveridge, R.; Heuck, A.; Vu, L.D.; Kurzbauer, R.; Hauer, K.; Thoeny, V.; Rumpel, K.; Mechtler, K. Structure of McsB, a protein kinase for regulated arginine phosphorylation. Nat. Chem. Biol. 2019, 15, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Semanjski, M.; Orlovetskie, N.; Bhattacharya, S.; Alon, S.; Argaman, L.; Jarrous, N.; Zhang, Y.; Macek, B.; Sinai, L. Arginine dephosphorylation propels spore germination in bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 14228–14237. [Google Scholar] [CrossRef] [Green Version]
- Ogbonna, E.C.; Anderson, H.R.; Schmitz, K.R. Identification of Arginine Phosphorylation in Mycolicibacterium smegmatis. Microbiol. Spectr. 2022, 10, e0204222. [Google Scholar] [CrossRef] [PubMed]
- Prust, N.; van Breugel, P.C.; Lemeer, S. Widespread Arginine Phosphorylation in Staphylococcus aureus. Mol. Cell. Proteom. 2022, 21, 100232. [Google Scholar] [CrossRef]
- Kolkman, A.; Daran-Lapujade, P.; Fullaondo, A.; Olsthoorn, M.M.A.; Pronk, J.T.; Slijper, M.; Heck, A.J.R. Proteome analysis of yeast response to various nutrient limitations. Mol. Syst. Biol. 2006, 2, 2006.0026. [Google Scholar] [CrossRef]
- Voisin, S.; Watson, D.C.; Tessier, L.; Ding, W.; Foote, S.; Bhatia, S.; Kelly, J.F.; Young, N.M. The cytoplasmic phosphoproteome of the Gram-negative bacterium Campylobacter jejuni: Evidence for modification by unidentified protein kinases. Proteomics 2007, 7, 4338–4348. [Google Scholar] [CrossRef]
- Schmidl, S.R.; Gronau, K.; Pietack, N.; Hecker, M.; Becher, D.; Stulke, J. The phosphoproteome of the minimal bacterium Mycoplasma pneumoniae: Analysis of the complete known Ser/Thr kinome suggests the existence of novel kinases. Mol. Cell Proteom. 2010, 9, 1228–1242. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.L.; Liao, J.H.; Lin, G.H.; Lin, M.H.; Chang, Y.C.; Liang, S.Y.; Yang, F.L.; Khoo, K.H.; Wu, S.H. Phosphoproteomic analysis reveals the effects of PilF phosphorylation on type IV pilus and biofilm formation in Thermus thermophilus HB27. Mol. Cell Proteom. 2013, 12, 2701–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, N.C.; Spat, P.; Mendez, J.A.; Nakedi, K.; Aranda, J.; Bou, G. Ser/Thr/Tyr phosphoproteome characterization of Acinetobacter baumannii: Comparison between a reference strain and a highly invasive multidrug-resistant clinical isolate. J. Proteom. 2014, 102, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-H.; Sugiyama, N.; Ishihama, Y. Systematic profiling of the bacterial phosphoproteome reveals bacterium-specific features of phosphorylation. Sci. Signal. 2015, 8, rs10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakedi, K.C.; Nel, A.J.; Garnett, S.; Blackburn, J.M.; Soares, N.C. Comparative Ser/Thr/Tyr phosphoproteomics between two mycobacterial species: The fast growing Mycobacterium smegmatis and the slow growing Mycobacterium bovis BCG. Front. Microbiol. 2015, 6, 237. [Google Scholar] [CrossRef] [Green Version]
- Fortuin, S.; Tomazella, G.G.; Nagaraj, N.; Sampson, S.L.; Gey van Pittius, N.C.; Soares, N.C.; Wiker, H.G.; de Souza, G.A.; Warren, R.M. Phosphoproteomics analysis of a clinical Mycobacterium tuberculosis Beijing isolate: Expanding the mycobacterial phosphoproteome catalog. Front. Microbiol. 2015, 6, 6. [Google Scholar] [CrossRef]
- de Keijzer, J.; Mulder, A.; de Beer, J.; de Ru, A.H.; van Veelen, P.A.; van Soolingen, D. Mechanisms of Phenotypic Rifampicin Tolerance in Mycobacterium tuberculosis Beijing Genotype Strain B0/W148 Revealed by Proteomics. J. Proteome Res. 2016, 15, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Potel, C.M.; Lin, M.H.; Heck, A.J.R.; Lemeer, S. Defeating Major Contaminants in Fe3+-Immobilized Metal Ion Affinity Chromatography (IMAC) Phosphopeptide Enrichment. Mol. Cell Proteom. 2018, 17, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.H.; Potel, C.M.; Tehrani, K.; Heck, A.J.R.; Martin, N.I.; Lemeer, S. A New Tool to Reveal Bacterial Signaling Mechanisms in Antibiotic Treatment and Resistance. Mol. Cell Proteom. 2018, 17, 2496–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semanjski, M.; Germain, E.; Bratl, K.; Kiessling, A.; Gerdes, K.; Macek, B. The kinases HipA and HipA7 phosphorylate different substrate pools in Escherichia coli to promote multidrug tolerance. Sci. Signal. 2018, 11, eaat5750. [Google Scholar] [CrossRef] [PubMed]
- Albeldas, C.; Ganief, N.; Calder, B.; Nakedi, K.C.; Garnett, S.; Nel, A.J.M.; Blackburn, J.M.; Soares, N.C. Global proteome and phosphoproteome dynamics indicate novel mechanisms of vitamin C induced dormancy in Mycobacterium smegmatis. J. Proteom. 2018, 180, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rioseras, B.; Shliaha, P.V.; Gorshkov, V.; Yagüe, P.; López-García, M.T.; Gonzalez-Quiñonez, N.; Kovalchuk, S.; Rogowska-Wrzesinska, A.; Jensen, O.N.; Manteca, A. Quantitative Proteome and Phosphoproteome Analyses of Streptomyces coelicolor Reveal Proteins and Phosphoproteins Modulating Differentiation and Secondary Metabolism. Mol. Cell Proteom. 2018, 17, 1591–1611. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Hou, L.; Liu, M.; Xu, X.; Gao, Q.; Deng, J.; Xiang, S.; Cao, Q.; Zhou, M.; Yang, Q.; et al. Phosphoproteomic Analysis of Spiroplasma eriocheiris and Crosstalk with Acetylome Reveals the Role of Post-Translational Modifications in Metabolism. Curr. Proteom. 2019, 17, 392–403. [Google Scholar] [CrossRef]
- Lim, S. Quantitative Bacterial Phosphoproteomics, Electronic, Scholarly Journal. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2015. [Google Scholar] [CrossRef]
- Yague, P.; Gonzalez-Quinonez, N.; Fernanez-Garcia, G.; Alonso-Fernandez, S.; Manteca, A. Goals and Challenges in Bacterial Phosphoproteomics. Int. J. Mol. Sci. 2019, 20, 5678. [Google Scholar] [CrossRef] [Green Version]
- Potel, C.M.; Lin, M.-H.; Heck, A.J.R.; Lemeer, S. Widespread bacterial protein histidine phosphorylation revealed by mass spectrometry-based proteomics. Nat. Methods 2018, 15, 187–190. [Google Scholar] [CrossRef]
- Baros-Steyl, S.S.; Nakedi, K.C.; Ganief, T.A.; Okendo, J.O.; Tabb, D.L.; Soares, N.C.; Blackburn, J.M. Phosphoproteomics reveals new insights into the role of PknG during the persistence of pathogenic mycobacteria in host macrophages. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hu, Q.; Yao, L.; Liao, X.; Zhang, L.-S.; Li, H.-T.; Li, T.-T.; Jiang, Q.-G.; Tan, M.-F.; Li, L.; Draheim, R.R.; et al. Comparative Phenotypic, Proteomic, and Phosphoproteomic Analysis Reveals Different Roles of Serine/Threonine Phosphatase and Kinase in the Growth, Cell Division, and Pathogenicity of Streptococcus suis. Microorganisms 2021, 9, 2442. [Google Scholar] [CrossRef]
- Luu, L.D.W.; Zhong, L.; Kaur, S.; Raftery, M.J.; Lan, R. Comparative Phosphoproteomics of Classical Bordetellae Elucidates the Potential Role of Serine, Threonine and Tyrosine Phosphorylation in Bordetella Biology and Virulence. Front. Cell. Infect. Microbiol. 2021, 11, 660280. [Google Scholar] [CrossRef]
- Prust, N.; van der Laarse, S.; van den Toorn, H.W.P.; van Sorge, N.M.; Lemeer, S. In-Depth Characterization of the Staphylococcus aureus Phosphoproteome Reveals New Targets of Stk1. Mol. Cell Proteom. 2021, 20, 100034. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Jers, C.; Ganief, T.A.; Shi, L.; Senissar, M.; Kohler, J.B.; Macek, B.; Mijakovic, I. Phosphoproteome Study of Escherichia coli Devoid of Ser/Thr Kinase YeaG during the Metabolic Shift from Glucose to Malate. Front. Microbiol. 2021, 12, 657562. [Google Scholar] [CrossRef]
- Alonso-Fernández, S.; Arribas-Díez, I.; Fernández-García, G.; González-Quiñónez, N.; Jensen, O.N.; Manteca, A. Quantitative phosphoproteome analysis of Streptomyces coelicolor by immobilized zirconium (IV) affinity chromatography and mass spectrometry reveals novel regulated protein phosphorylation sites and sequence motifs. J. Proteom. 2022, 269, 104719. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, T.; Douché, T.; Giai Gianetto, Q.; Poncet, S.; El Omrani, N.; Smits, W.K.; Cuenot, E.; Matondo, M.; Martin-Verstraete, I. In-Depth Characterization of the Clostridioides difficile Phosphoproteome to Identify Ser/Thr Kinase Substrates. Mol. Cell. Proteom. 2022, 21, 100428. [Google Scholar] [CrossRef] [PubMed]
- Šarić, E.; Quinn, G.A.; Nalpas, N.; Paradžik, T.; Kazazić, S.; Filić, Ž.; Šemanjski, M.; Herron, P.; Hunter, I.; Maček, B.; et al. Phosphoproteome Dynamics of Streptomyces rimosus during Submerged Growth and Antibiotic Production. mSystems 2022, 7, e0019922. [Google Scholar] [CrossRef]
- Smits Wiep, K.; Mohammed, Y.; de Ru Arnoud, H.; Cordo, V.; Friggen Annemieke, H.; van Veelen Peter, A.; Hensbergen Paul, J. Clostridioides difficile Phosphoproteomics Shows an Expansion of Phosphorylated Proteins in Stationary Growth Phase. mSphere 2022, 7, e0091121. [Google Scholar] [CrossRef]
- Licona-Cassani, C.; Lim, S.; Marcellin, E.; Nielsen, L.K. Temporal dynamics of the Saccharopolyspora erythraea phosphoproteome. Mol. Cell Proteom. 2014, 13, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.H.; Yang, J.T.; Chern, J.; Chen, T.L.; Wu, W.L.; Liao, J.H.; Tsai, S.F.; Liang, S.Y.; Chou, C.C.; Wu, S.H. Comparative Phosphoproteomics Reveals the Role of AmpC β-lactamase Phosphorylation in the Clinical Imipenem-resistant Strain Acinetobacter baumannii SK17. Mol. Cell Proteom. 2016, 15, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Basell, K.; Otto, A.; Junker, S.; Zuhlke, D.; Rappen, G.M.; Schmidt, S.; Hentschker, C.; Macek, B.; Ohlsen, K.; Hecker, M.; et al. The phosphoproteome and its physiological dynamics in Staphylococcus aureus. Int. J. Med. Microbiol. 2014, 304, 121–132. [Google Scholar] [CrossRef]
- Lin, M.H.; Hsu, T.L.; Lin, S.Y.; Pan, Y.J.; Jan, J.T.; Wang, J.T.; Khoo, K.H.; Wu, S.H. Phosphoproteomics of Klebsiella pneumoniae NTUH-K2044 reveals a tight link between tyrosine phosphorylation and virulence. Mol. Cell Proteom. 2009, 8, 2613–2623. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, A.; Sugiyama, N.; Tomita, M.; Swarup, S.; Ishihama, Y. Ser/Thr/Tyr phosphoproteome analysis of pathogenic and non-pathogenic Pseudomonas species. Proteomics 2009, 9, 2764–2775. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ge, F.; Xiao, C.-L.; Yin, X.-F.; Ge, R.; Zhang, L.-H.; He, Q.-Y. Phosphoproteomic Analysis Reveals the Multiple Roles of Phosphorylation in Pathogenic Bacterium Streptococcus pneumoniae. J. Proteome Res. 2010, 9, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Prisic, S.; Dankwa, S.; Schwartz, D.; Chou, M.F.; Locasale, J.W.; Kang, C.-M.; Bemis, G.; Church, G.M.; Steen, H.; Husson, R.N. Extensive phosphorylation with overlapping specificity by Mycobacterium tuberculosis serine/threonine protein kinases. Proc. Natl. Acad. Sci. USA 2010, 107, 7521–7526. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.L.; Jones, A.M.; Serazetdinova, L.; Saalbach, G.; Bibb, M.J.; Naldrett, M.J. Analysis of the phosphoproteome of the multicellular bacterium Streptomyces coelicolor A3(2) by protein/peptide fractionation, phosphopeptide enrichment and high-accuracy mass spectrometry. Proteomics 2010, 10, 2486–2497. [Google Scholar] [CrossRef]
- Misra, S.K.; Milohanic, E.; Ake, F.; Mijakovic, I.; Deutscher, J.; Monnet, V.; Henry, C. Analysis of the serine/threonine/tyrosine phosphoproteome of the pathogenic bacterium Listeria monocytogenes reveals phosphorylated proteins related to virulence. Proteomics 2011, 11, 4155–4165. [Google Scholar] [CrossRef] [PubMed]
- Manteca, A.; Ye, J.; Sanchez, J.; Jensen, O.N. Phosphoproteome analysis of Streptomyces development reveals extensive protein phosphorylation accompanying bacterial differentiation. J. Proteome Res. 2011, 10, 5481–5492. [Google Scholar] [CrossRef]
- Ge, R.; Sun, X.; Xiao, C.; Yin, X.; Shan, W.; Chen, Z.; He, Q.Y. Phosphoproteome analysis of the pathogenic bacterium Helicobacter pylori reveals over-representation of tyrosine phosphorylation and multiply phosphorylated proteins. Proteomics 2011, 11, 1449–1461. [Google Scholar] [CrossRef]
- Bai, X.; Ji, Z. Phosphoproteomic investigation of a solvent producing bacterium Clostridium acetobutylicum. Appl. Microbiol. Biotechnol. 2012, 95, 201–211. [Google Scholar] [CrossRef]
- Hu, C.-W.; Lin, M.-H.; Huang, H.-C.; Ku, W.-C.; Yi, T.-H.; Tsai, C.-F.; Chen, Y.-J.; Sugiyama, N.; Ishihama, Y.; Juan, H.-F.; et al. Phosphoproteomic Analysis of Rhodopseudomonas palustris Reveals the Role of Pyruvate Phosphate Dikinase Phosphorylation in Lipid Production. J. Proteome Res. 2012, 11, 5362–5375. [Google Scholar] [CrossRef]
- Takahata, Y.; Inoue, M.; Kim, K.; Iio, Y.; Miyamoto, M.; Masui, R.; Ishihama, Y.; Kuramitsu, S. Close proximity of phosphorylation sites to ligand in the phosphoproteome of the extreme thermophile Thermus thermophilus HB8. Proteomics 2012, 12, 1414–1430. [Google Scholar] [CrossRef]
- Yang, M.K.; Qiao, Z.X.; Zhang, W.Y.; Xiong, Q.; Zhang, J.; Li, T.; Ge, F.; Zhao, J.D. Global phosphoproteomic analysis reveals diverse functions of serine/threonine/tyrosine phosphorylation in the model cyanobacterium Synechococcus sp. strain PCC 7002. J. Proteome Res. 2013, 12, 1909–1923. [Google Scholar] [CrossRef]
- Soares, N.C.; Spat, P.; Krug, K.; Macek, B. Global dynamics of the Escherichia coli proteome and phosphoproteome during growth in minimal medium. J. Proteome Res. 2013, 12, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, V.; Shi, L.; Krug, K.; Derouiche, A.; Jers, C.; Cousin, C.; Kobir, A.; Mijakovic, I.; Macek, B. Quantitative phosphoproteome analysis of Bacillus subtilis reveals novel substrates of the kinase PrkC and phosphatase PrpC. Mol. Cell Proteom. 2014, 13, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Ouidir, T.; Jarnier, F.; Cosette, P.; Jouenne, T.; Hardouin, J. Extracellular Ser/Thr/Tyr phosphorylated proteins of Pseudomonas aeruginosa PA14 strain. Proteomics 2014, 14, 2017–2030. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.K.; Moussan Désirée Aké, F.; Wu, Z.; Milohanic, E.; Cao, T.N.; Cossart, P.; Deutscher, J.; Monnet, V.; Archambaud, C.; Henry, C. Quantitative proteome analyses identify PrfA-responsive proteins and phosphoproteins in Listeria monocytogenes. J. Proteome Res. 2014, 13, 6046–6057. [Google Scholar] [CrossRef]
- Liu, T.; Tian, C.F.; Chen, W.X. Site-Specific Ser/Thr/Tyr Phosphoproteome of Sinorhizobium meliloti at Stationary Phase. PLoS ONE 2015, 10, e0139143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, A.; Soufi, B.; Ravikumar, V.; Soares, N.C.; Krug, K.; Smith, Y.; Macek, B.; Ben-Yehuda, S. Phosphoproteome dynamics mediate revival of bacterial spores. BMC Biol. 2015, 13, 76. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Marcellin, E.; Jacob, S.; Nielsen, L.K. Global dynamics of Escherichia coli phosphoproteome in central carbon metabolism under changing culture conditions. J. Proteom. 2015, 126, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Spät, P.; Maček, B.; Forchhammer, K. Phosphoproteome of the cyanobacterium Synechocystis sp. PCC 6803 and its dynamics during nitrogen starvation. Front. Microbiol. 2015, 6, 248. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.; Pinto, S.M.; Patil, A.H.; Advani, J.; Subba, P.; Kumar, M.; Sharma, J.; Dey, G.; Ravikumar, R.; Buggi, S.; et al. Quantitative Proteomic and Phosphoproteomic Analysis of H37Ra and H37Rv Strains of Mycobacterium tuberculosis. J. Proteome Res. 2017, 16, 1632–1645. [Google Scholar] [CrossRef]
- Qu, J.; Shen, L.; Zhao, M.; Li, W.; Jia, C.; Zhu, H.; Zhang, Q. Determination of the Role of Microcystis aeruginosa in Toxin Generation Based on Phosphoproteomic Profiles. Toxins 2018, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Tatli, M.; Hebert, A.S.; Coon, J.J.; Amador-Noguez, D. Genome Wide Phosphoproteome Analysis of Zymomonas mobilis under Anaerobic, Aerobic, and N(2)-Fixing Conditions. Front. Microbiol. 2019, 10, 1986. [Google Scholar] [CrossRef] [PubMed]
- Errington, J.; Aart, L.T.V. Microbe Profile: Bacillus subtilis: Model organism for cellular development, and industrial workhorse. Microbiology 2020, 166, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.I.; Rahman, M.M.; Salam, M.A.; Khan, M.A.R.; Islam, M.T. Identification of marine sponge-associated bacteria of the Saint Martin’s island of the Bay of Bengal emphasizing on the prevention of motile Aeromonas septicemia in Labeo rohita. Aquaculture 2021, 545, 737156. [Google Scholar] [CrossRef]
- Rahman, M.M.; Paul, S.I.; Akter, T.; Tay, A.C.Y.; Foysal, M.J.; Islam, M.T. Whole-Genome Sequence of Bacillus subtilis WS1A, a Promising Fish Probiotic Strain Isolated from Marine Sponge of the Bay of Bengal. Microbiol. Resour. Announc. 2020, 9, e00641-20. [Google Scholar] [CrossRef]
- Sriskandan, S.; Slater, J.D. Invasive disease and toxic shock due to zoonotic Streptococcus suis: An emerging infection in the East? PLoS Med. 2006, 3, e187. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Mateus, A.; Hevler, J.; Bobonis, J.; Kurzawa, N.; Shah, M.; Mitosch, K.; Goemans, C.V.; Helm, D.; Stein, F.; Typas, A.; et al. The functional proteome landscape of Escherichia coli. Nature 2020, 588, 473–478. [Google Scholar] [CrossRef]
- Riggs, P.J.; Chelius, M.; Iniguez, A.L.; Kaeppler, S.; Triplett, E. Enhanced maize productivity by inoculation with diazotrophic bacteria. Aust. J. Plant Physiol. 2001, 28, 829–836. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Organism | Strain | Year | P-pro. (ea) | P-pep. (ea) | P-site (ea) | Ser (%) | Thr (%) | Tyr (%) | Arg (%) | His (%) | Refs. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C. jejuni | 11168 | 2007 | 36 | 58 | 35 | 30.3 | 72.7 | 9.1 | [73] | ||
| B. subtilis | 168 | 2007 | 78 | 103 | 78 | 69.2 | 20.5 | 10.3 | [3] | ||
| L. lactis | Il1403 | 2008 | 63 | 102 | 79 | 46.5 | 50.6 | 2.7 | [48] | ||
| E. coli K12 | MG1665 | 2008 | 79 | 105 | 81 | 67.9 | 23.5 | 8.6 | [4] | ||
| K. pneumoniae | K2044 | 2009 | 81 | 117 | 93 | 31.2 | 15.4 | 25.8 | [102] | ||
| P. putida | MK25 | 2009 | 40 | 56 | 53 | 52.8 | 39.6 | 7.5 | [103] | ||
| P. aeruginosa | PAO1 | 2009 | 23 | 57 | 55 | 52.7 | 32.7 | 14.5 | [103] | ||
| M. pneumoniae | M129 | 2010 | 63 | 16 | 16 | 53.3 | 46.7 | 0 | [74] | ||
| S. pneumoniae | D39 | 2010 | 84 | 102 | 163 | 47.2 | 43.8 | 9 | [104] | ||
| M. tuberculosis | H37Rv | 2010 | 301 | 381 | 506 | 40 | 60 | 0 | [105] | ||
| S. coelicolor | A3(2) | 2010 | 40 | 44 | 46 | 34.1 | 52.3 | 13.6 | [106] | ||
| L. monocytogenes | EGDe | 2011 | 112 | 155 | 143 | 93 | 43 | 7 | [107] | ||
| S. coelicolor | M145 | 2011 | 127 | 260 | 289 | 46.8 | 48 | 5.2 | [108] | ||
| H. pylori | 26695 | 2011 | 67 | 80 | 124 | 42.8 | 38.7 | 18.5 | [109] | ||
| C. acetobutylicum | ATCC824 | 2012 | 61 | 82 | 107 | 42 | 47.6 | 10.6 | [110] | ||
| R. palustris(Ch) | CGA010 | 2012 | 54 | 100 | 63 | 63.3 | 16.1 | 19.4 | [111] | ||
| R. palustris(Ph) | CGA010 | 2012 | 42 | 74 | 59 | 58.9 | 23.2 | 17.9 | [111] | ||
| T. thermophilus | HB8 | 2012 | 48 | 52 | 46 | 30 | 12 | 4 | [112] | ||
| T. thermophilus | HB27 | 2013 | 53 | 93 | 67 | 57 | 36 | 7 | [75] | ||
| Synechococcus sp. | PCC7002 | 2013 | 245 | 280 | 410 | 43.9 | 42.4 | 13.6 | [113] | ||
| E. coli K12 | BW25113 | 2013 | 133 | 150 | 108 | 75.9 | 16.7 | 7.4 | [114] | ||
| S. aureus | COL | 2014 | 108 | 68 | 50 | 25 | 15 | 10 | [101] | ||
| A. baumannii | AbH120A2 | 2014 | 70 | 80 | 70.8 | 25.2 | 3.8 | [76] | |||
| A. baumannii | 17978 | 2014 | 41 | 48 | 68.9 | 24.1 | 5.2 | [76] | |||
| B. subtilis | 168 | 2014 | 177 | 155 | 74.6 | 18.6 | 7.3 | [115] | |||
| S. erythraea | NRRL2338 | 2014 | 88 | 109 | 47 | 45 | 8 | 5.3 | [99] | ||
| P. aeruginosa | PA14 | 2014 | 28 | 43 | 59 | 49 | 24 | 27 | [116] | ||
| L. monocytogenes | ∆PrfA | 2014 | 191 | 256 | 242 | 155 | 75 | 12 | [117] | ||
| S. meliloti | CCBAU | 2015 | 77 | 88 | 96 | 63 | 28 | 5 | [118] | ||
| B. subtilis | Spore | 2015 | 124 | 155 | 77.41 | 22.6 | [119] | ||||
| B. subtilis | 168 | 2015 | 175 | 441 | 339 | 74.8 | 17.7 | 7.1 | [77] | ||
| E. coli K12 | BW25113 | 2015 | 392 | 1212 | 1088 | 69.5 | 21.8 | 7.7 | [77] | ||
| E. coli K12 | MG1655 | 2015 | 71 | 82 | [120] | ||||||
| K. pneumoniae | K2044 | 2015 | 286 | 663 | 559 | 72.9 | 13.7 | 12.9 | [77] | ||
| Synechocystis sp. | PCC 6803 | 2015 | 188 | 242 | 262 | [121] | |||||
| M. tuberculosis | SAW5527 | 2015 | 214 | 303 | 414 | 38 | 59 | 3 | [79] | ||
| M. smegmatis | mc2155 | 2015 | 2462 | 464 | 185 | 39.5 | 57.1 | 3.5 | [78] | ||
| M. bovis BCG | 1173P2 | 2015 | 1765 | 402 | 442 | 35 | 61.6 | 3.1 | [78] | ||
| M. tuberculosis | B0/W148 | 2016 | 132 | 180 | 191 | 22 | 76 | 2 | [80] | ||
| A. baumannii | SK17-S | 2016 | 248 | 351 | 410 | 47 | 27.6 | 12.4 | 4.9 | [100] | |
| A. baumannii | SK17-R | 2016 | 211 | 240 | 285 | 41.4 | 29.5 | 17.5 | 4.9 | [100] | |
| M. tuberculosis | H37Ra | 2017 | 257 | 512 | 29 | 68 | 3 | [122] | |||
| M. smegmatis | mc2155 | 2018 | 154 | 222 | 242 | 24.8 | 74.0 | 1.2 | [84] | ||
| M. aeruginosa | FACHB-469 | 2018 | 37 | 59 | [123] | ||||||
| M. aeruginosa | FACHB-905 | 2018 | 18 | 26 | [123] | ||||||
| S. coelicolor | M145 | 2018 | 48 | 92 | 85 | 50.6 | 47.4 | 2 | [85] | ||
| E. coli K12 | MG1665 | 2018 | 632 | 1178 | 1183 | [83] | |||||
| E. coli K12 | W3110 | 2018 | 861 | 2446 | 57.2 | 25.3 | 8.5 | 9 | [81] | ||
| E. coli K12 | W3110 | 2018 | 781 | 2057 | 2129 | 1220 | 501 | 162 | 246 | [89] | |
| E. coli K12 | W3110 | 2018 | 2248 | 56 | 20 | 13 | 5 | 5 | [82] | ||
| Z. mobilis | ZM4,31821 | 2019 | 125 | 177 | 73 | 21 | 6 | [124] | |||
| S. thermophilus | LMD9 | 2019 | 106 | 410 | 161 | 43 | 33 | 23 | [56] | ||
| S. eriocheiris | M207170 | 2019 | 245 | 465 | [86] | ||||||
| E. coli K12 | 1655, ∆yea | 2021 | 83 | 127 | 67.7 | 28.3 | 3.9 | [94] | |||
| B. subtilis | 168 | 2021 | 146 | 283 | 267 | 73 | 12.7 | 7.5 | 6.7 | [93] | |
| S. aureus | USA300 | 2021 | 859 | 3800 | 3771 | 55.2 | 29.6 | 7.3 | 7.8 | [93] | |
| B. subtilis | 168 | 2021 | 153 | 214 | 67 | 28 | 5 | [54] | |||
| S. pyogenes | M1 | 2021 | 205 | 449 | 41 | 55 | 4 | [54] | |||
| L. monocytogenes | EGDe | 2021 | 241 | 420 | 56 | 35 | 9 | [54] | |||
| B. pertussis | L1423 | 2021 | 45 | 53 | 54 | 72 | 17 | 11 | [92] | ||
| B. bronchiseptica | RB50 | 2021 | 23 | 28 | 29 | 69 | 21 | 10 | [92] | ||
| B. parapertussis | 12822 | 2021 | 42 | 50 | 50 | 80 | 12 | 8 | [92] | ||
| M. bovis | BCG, ΔPknG | 2021 | 914 | 1371 | 1401 | 85.3 | 13.4 | 1.3 | [90] | ||
| S. suis | WT, Δstp | 2021 | 50 | 73 | [91] | ||||||
| S. suis | WT, Δstk | 2021 | 67 | 87 | [91] | ||||||
| S. aureus | NE98, ΔSdrE | 2022 | 953 | 4407 | 45.5 | 24 | 5 | 20.2 | 5.4 | [71] | |
| S. aureus | NE217, ΔStk1 | 2022 | 903 | 3779 | 48.1 | 22 | 6.7 | 18 | 5.2 | [71] | |
| S. aureus | NE1919, ΔStp1 | 2022 | 951 | 4085 | 40.2 | 21.2 | 6.1 | 26 | 6.5 | [71] | |
| C. difficile | 630WT | 2022 | 700 | 2994 | 1759 | 75 | 20 | 5 | [98] | ||
| C. difficile | 630WT, Δ erm | 2022 | 504 | 1061 | 117 | 76.6 | 17.8 | 5.6 | [96] | ||
| S. rimosus | G7, 10970 | 2022 | 230 | 273 | 417 | 41.3 | 53.5 | 5.3 | [97] | ||
| S. coelicolor | A3(2) | 2022 | 187 | 351 | 361 | 41 | 56.2 | 2.8 | [95] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S. A Review of the Bacterial Phosphoproteomes of Beneficial Microbes. Microorganisms 2023, 11, 931. https://doi.org/10.3390/microorganisms11040931
Lim S. A Review of the Bacterial Phosphoproteomes of Beneficial Microbes. Microorganisms. 2023; 11(4):931. https://doi.org/10.3390/microorganisms11040931
Chicago/Turabian StyleLim, Sooa. 2023. "A Review of the Bacterial Phosphoproteomes of Beneficial Microbes" Microorganisms 11, no. 4: 931. https://doi.org/10.3390/microorganisms11040931
APA StyleLim, S. (2023). A Review of the Bacterial Phosphoproteomes of Beneficial Microbes. Microorganisms, 11(4), 931. https://doi.org/10.3390/microorganisms11040931