

Glacial Water: A Dynamic Microbial Medium

 , , and
, , and 
Abstract
:1. Introduction
2. Glacial Hydrology
2.1. Seasonal Variation
2.1.1. Ablation Season
2.1.2. Seasonal Evolution
2.2. Hydrological Residence Times
3. Hydrology Influences on Glacial Nutrients and Microbial Communities
3.1. Deposition of Nutrients and Microorganisms on Glacial Surfaces
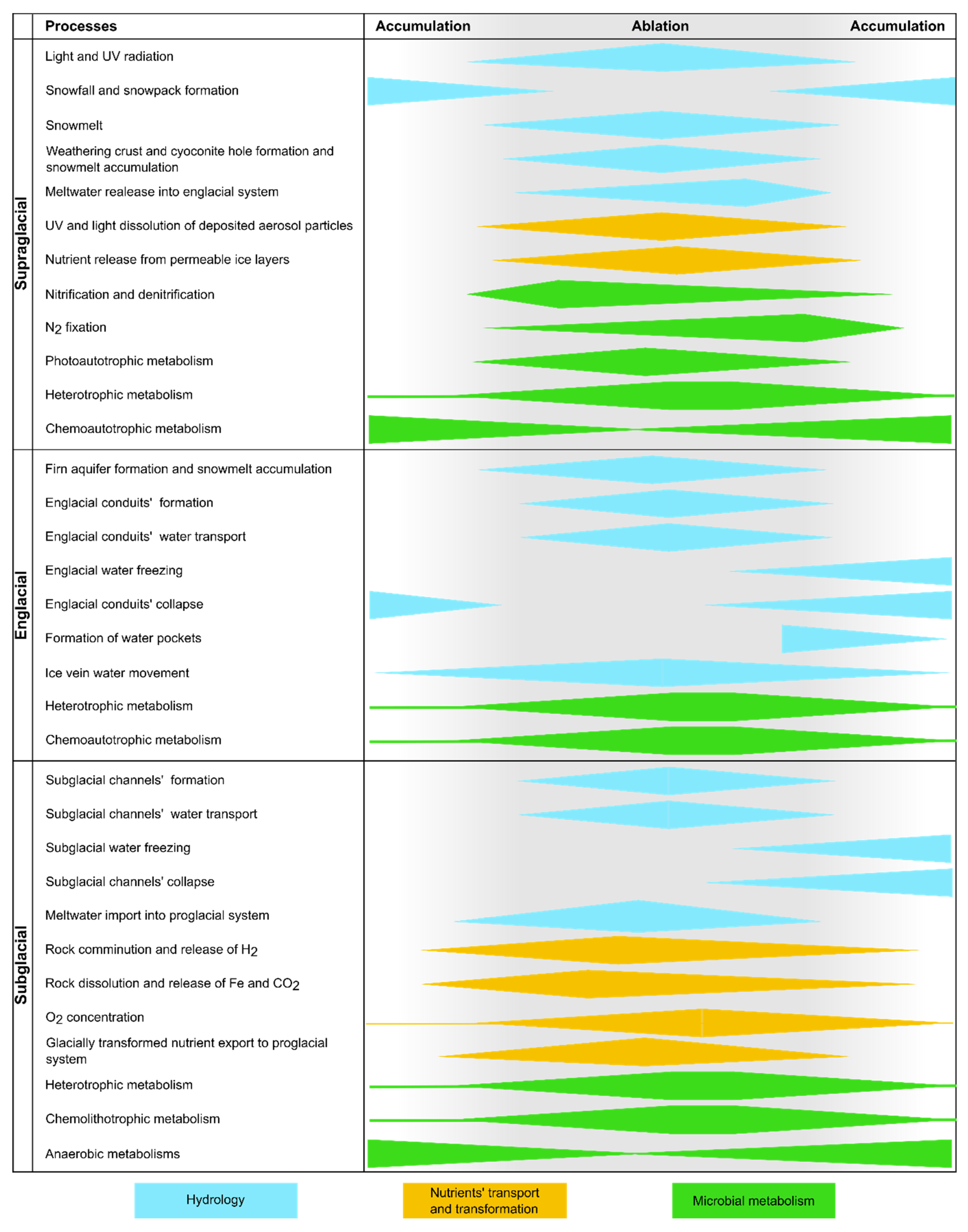
3.2. Microbial and Geochemical Dynamics during the Ablation Season
3.2.1. Supraglacial Realm
3.2.2. Englacial Realm
3.2.3. Subglacial Realm
3.3. Microbial and Geochemical Dynamics during the Accumulation Season
4. Glaciers and Ice Sheets as Bioreactors
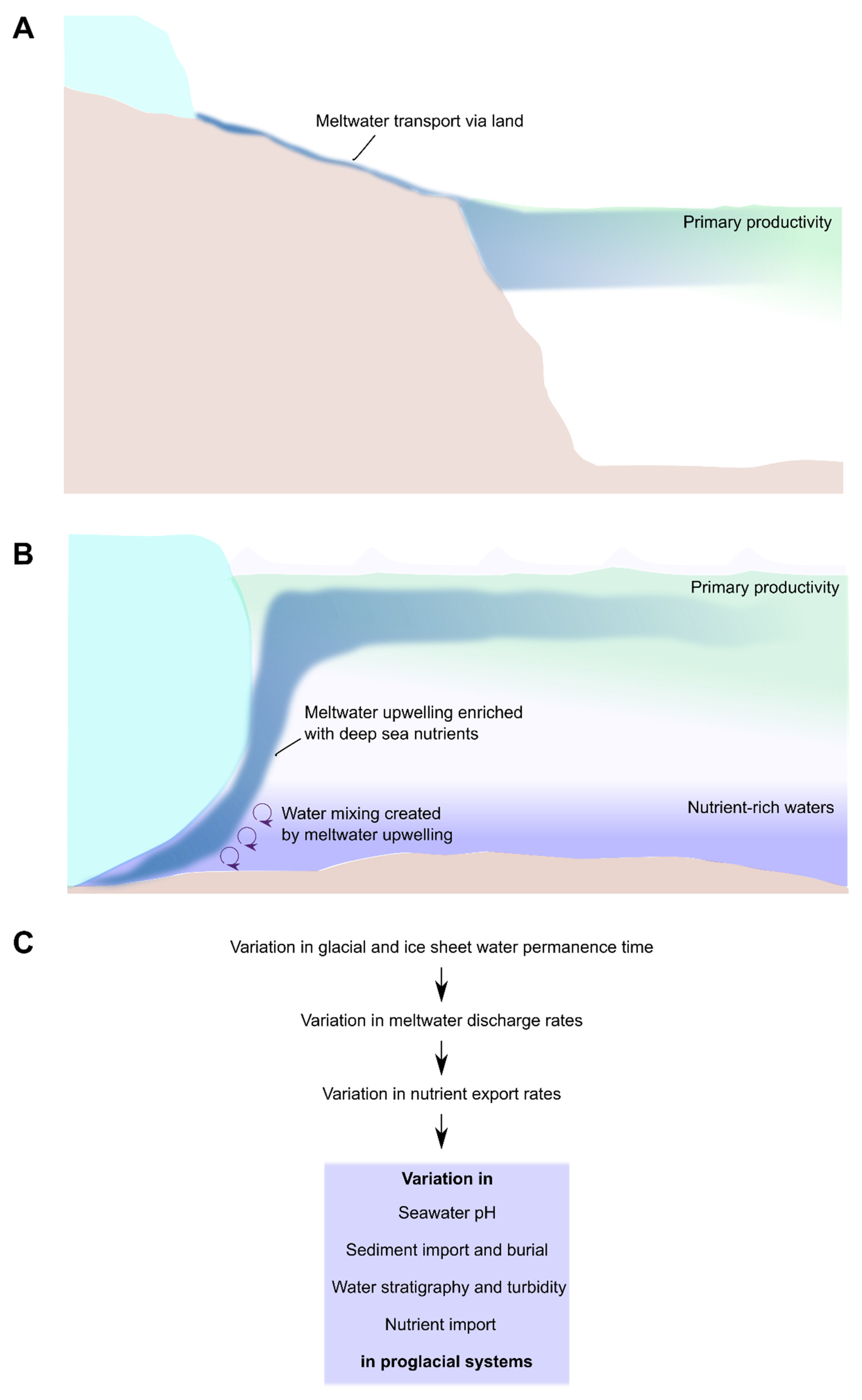
5. Proglacial Systems
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hooke, R. Principles of Glacier Mechanics, 3rd ed.; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Jones, D.B.; Harrison, S.; Anderson, K.; Whalley, W.B. Rock Glaciers and Mountain Hydrology: A Review. Earth-Sci. Rev. 2019, 193, 66–90. [Google Scholar] [CrossRef]
- Zemp, M.; Huss, M.; Thibert, E.; Eckert, N.; McNabb, R.; Huber, J.; Barandun, M.; Machguth, H.; Nussbaumer, S.U.; Gärtner-Roer, I.; et al. Global Glacier Mass Changes and Their Contributions to Sea-Level Rise from 1961 to 2016. Nature 2019, 568, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Hotaling, S.; Hood, E.; Hamilton, T.L. Microbial Ecology of Mountain Glacier Ecosystems: Biodiversity, Ecological Connections and Implications of a Warming Climate. Environ. Microbiol. 2017, 19, 2935–2948. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.A.; Müller, O.; Stibal, M.; Edwards, A.; Jacobsen, C.S. Glacial Microbiota Are Hydrologically Connected and Temporally Variable. Environ. Microbiol. 2020, 22, 3172–3187. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.A.; Stibal, M.; Hawkings, J.R.; Mikkelsen, A.B.; Telling, J.; Kohler, T.J.; Gözdereliler, E.; Zarsky, J.D.; Wadham, J.L.; Jacobsen, C.S. Meltwater Export of Prokaryotic Cells from the Greenland Ice Sheet: Microbial Export from the Greenland Ice Sheet. Environ. Microbiol. 2017, 19, 524–534. [Google Scholar] [CrossRef]
- Fegel, T.; Boot, C.M.; Broeckling, C.D.; Baron, J.S.; Hall, E.K. Assessing the Chemistry and Bioavailability of Dissolved Organic Matter From Glaciers and Rock Glaciers. J. Geophys. Res. Biogeosci. 2019, 124, 1988–2004. [Google Scholar] [CrossRef]
- Kohler, T.J.; Vinšová, P.; Falteisek, L.; Žárský, J.D.; Yde, J.C.; Hatton, J.E.; Hawkings, J.R.; Lamarche-Gagnon, G.; Hood, E.; Cameron, K.A.; et al. Patterns in Microbial Assemblages Exported From the Meltwater of Arctic and Sub-Arctic Glaciers. Front. Microbiol. 2020, 11, 669. [Google Scholar] [CrossRef]
- Chu, V.W. Greenland Ice Sheet Hydrology: A Review. Prog. Phys. Geogr. Earth Environ. 2014, 38, 19–54. [Google Scholar] [CrossRef]
- Simkins, L.M.; Greenwood, S.L.; Munevar Garcia, S.; Eareckson, E.A.; Anderson, J.B.; Prothro, L.O. Topographic Controls on Channelized Meltwater in the Subglacial Environment. Geophys. Res. Lett. 2021, 48, e2021GL094678. [Google Scholar] [CrossRef]
- Tranter, M.; Skidmore, M.; Wadham, J. Hydrological Controls on Microbial Communities in Subglacial Environments. Hydrol. Process. 2005, 19, 995–998. [Google Scholar] [CrossRef]
- Dubnick, A.; Kazemi, S.; Sharp, M.; Wadham, J.; Hawkings, J.; Beaton, A.; Lanoil, B. Hydrological Controls on Glacially Exported Microbial Assemblages: Subglacial Hydrology and Microbiology. J. Geophys. Res. Biogeosci. 2017, 122, 1049–1061. [Google Scholar] [CrossRef]
- Raiswell, R.; Hawkings, J.; Elsenousy, A.; Death, R.; Tranter, M.; Wadham, J. Iron in Glacial Systems: Speciation, Reactivity, Freezing Behavior, and Alteration During Transport. Front. Earth Sci. 2018, 6, 222. [Google Scholar] [CrossRef]
- Margesin, R.; Collins, T. Microbial Ecology of the Cryosphere (Glacial and Permafrost Habitats): Current Knowledge. Appl. Microbiol. Biotechnol. 2019, 103, 2537–2549. [Google Scholar] [CrossRef]
- Barker, J.D.; Sharp, M.J.; Fitzsimons, S.J.; Turner, R.J. Abundance and Dynamics of Dissolved Organic Carbon in Glacier Systems. Arct. Antarct. Alp. Res. 2006, 38, 163–172. [Google Scholar] [CrossRef]
- Dubnick, A.; Barker, J.; Sharp, M.; Wadham, J.; Lis, G.; Telling, J.; Fitzsimons, S.; Jackson, M. Characterization of Dissolved Organic Matter (DOM) from Glacial Environments Using Total Fluorescence Spectroscopy and Parallel Factor Analysis. Ann. Glaciol. 2010, 51, 111–122. [Google Scholar] [CrossRef]
- Anesio, A.M.; Laybourn-Parry, J. Ecology of Arctic Glaciers. In Arctic Ecology; Thomas, D.N., Ed.; Wiley: Hoboken, NJ, USA, 2021; pp. 133–158. ISBN 978-1-118-84654-4. [Google Scholar]
- Boyd, E.S.; Hamilton, T.L.; Havig, J.R.; Skidmore, M.L.; Shock, E.L. Chemolithotrophic Primary Production in a Subglacial Ecosystem. Appl. Environ. Microbiol. 2014, 80, 6146–6153. [Google Scholar] [CrossRef]
- Hodson, A.; Anesio, A.M.; Tranter, M.; Fountain, A.; Osborn, M.; Priscu, J.; Laybourn-Parry, J.; Sattler, B. Glacial Ecosystems. Ecol. Monogr. 2008, 78, 41–67. [Google Scholar] [CrossRef]
- Bradley, J.A.; Trivedi, C.B.; Winkel, M.; Mourot, R.; Lutz, S.; Larose, C.; Keuschnig, C.; Doting, E.; Halbach, L.; Zervas, A.; et al. Active and Dormant Microorganisms on Glacier Surfaces. Geobiology 2023, 21, 244–261. [Google Scholar] [CrossRef]
- Jain, S. Fundamentals of Physical Geology; Springer Geology; Springer India: New Delhi, India, 2014; ISBN 978-81-322-1538-7. [Google Scholar]
- Buytaert, W.; Moulds, S.; Acosta, L.; De Bièvre, B.; Olmos, C.; Villacis, M.; Tovar, C.; Verbist, K.M.J. Glacial Melt Content of Water Use in the Tropical Andes. Environ. Res. Lett. 2017, 12, 114014. [Google Scholar] [CrossRef]
- Milner, A.M.; Khamis, K.; Battin, T.J.; Brittain, J.E.; Barrand, N.E.; Füreder, L.; Cauvy-Fraunié, S.; Gíslason, G.M.; Jacobsen, D.; Hannah, D.M.; et al. Glacier Shrinkage Driving Global Changes in Downstream Systems. Proc. Natl. Acad. Sci. USA 2017, 114, 9770–9778. [Google Scholar] [CrossRef]
- Biemans, H.; Siderius, C.; Lutz, A.F.; Nepal, S.; Ahmad, B.; Hassan, T.; von Bloh, W.; Wijngaard, R.R.; Wester, P.; Shrestha, A.B.; et al. Importance of Snow and Glacier Meltwater for Agriculture on the Indo-Gangetic Plain. Nat. Sustain. 2019, 2, 594–601. [Google Scholar] [CrossRef]
- Catania, G.A.; Stearns, L.A.; Moon, T.A.; Enderlin, E.M.; Jackson, R.H. Future Evolution of Greenland’s Marine-Terminating Outlet Glaciers. J. Geophys. Res. Earth Surf. 2020, 125, e2018JF004873. [Google Scholar] [CrossRef]
- Slater, T.; Shepherd, A.; McMillan, M.; Leeson, A.; Gilbert, L.; Muir, A.; Munneke, P.K.; Noël, B.; Fettweis, X.; van den Broeke, M.; et al. Increased Variability in Greenland Ice Sheet Runoff from Satellite Observations. Nat. Commun. 2021, 12, 6069. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Moosdorf, N.; Hartmann, J.; Adkins, J.F.; West, A.J. Glacial Weathering, Sulfide Oxidation, and Global Carbon Cycle Feedbacks. Proc. Natl. Acad. Sci. USA 2017, 114, 8716–8721. [Google Scholar] [CrossRef]
- Delaney, I.; Adhikari, S. Increased Subglacial Sediment Discharge in a Warming Climate: Consideration of Ice Dynamics, Glacial Erosion, and Fluvial Sediment Transport. Geophys. Res. Lett. 2020, 47, e2019GL085672. [Google Scholar] [CrossRef]
- Stevens, I.T.; Irvine-Fynn, T.D.L.; Edwards, A.; Mitchell, A.C.; Cook, J.M.; Porter, P.R.; Holt, T.O.; Huss, M.; Fettweis, X.; Moorman, B.J.; et al. Spatially Consistent Microbial Biomass and Future Cellular Carbon Release from Melting Northern Hemisphere Glacier Surfaces. Commun. Earth Environ. 2022, 3, 275. [Google Scholar] [CrossRef]
- Anderson, S.P. Biogeochemistry of Glacial Landscape Systems. Annu. Rev. Earth Planet. Sci. 2007, 35, 375–399. [Google Scholar] [CrossRef]
- Anesio, A.M.; Sattler, B.; Foreman, C.; Telling, J.; Hodson, A.; Tranter, M.; Psenner, R. Carbon Fluxes through Bacterial Communities on Glacier Surfaces. Ann. Glaciol. 2010, 51, 32–40. [Google Scholar] [CrossRef]
- Hawkings, J.; Wadham, J.; Tranter, M.; Telling, J.; Bagshaw, E.; Beaton, A.; Simmons, S.-L.; Chandler, D.; Tedstone, A.; Nienow, P. The Greenland Ice Sheet as a Hot Spot of Phosphorus Weathering and Export in the Arctic: THE GREENLAND ICE SHEET P CYCLE. Glob. Biogeochem. Cycles 2016, 30, 191–210. [Google Scholar] [CrossRef]
- Wadham, J.L.; Hawkings, J.R.; Tarasov, L.; Gregoire, L.J.; Spencer, R.G.M.; Gutjahr, M.; Ridgwell, A.; Kohfeld, K.E. Ice Sheets Matter for the Global Carbon Cycle. Nat. Commun. 2019, 10, 3567. [Google Scholar] [CrossRef]
- Boetius, A.; Anesio, A.M.; Deming, J.W.; Mikucki, J.A.; Rapp, J.Z. Microbial Ecology of the Cryosphere: Sea Ice and Glacial Habitats. Nat. Rev. Microbiol. 2015, 13, 677–690. [Google Scholar] [CrossRef]
- Anesio, A.M.; Lutz, S.; Chrismas, N.A.M.; Benning, L.G. The Microbiome of Glaciers and Ice Sheets. npj Biofilms Microbiomes 2017, 3, 10. [Google Scholar] [CrossRef]
- Fountain, A.G.; Walder, J.S. Water Flow through Temperate Glaciers. Rev. Geophys. 1998, 36, 299–328. [Google Scholar] [CrossRef]
- St Germain, S.L.; Moorman, B.J. Long-Term Observations of Supraglacial Streams on an Arctic Glacier. J. Glaciol. 2019, 65, 900–911. [Google Scholar] [CrossRef]
- Cook, S.J.; Swift, D.A.; Kirkbride, M.P.; Knight, P.G.; Waller, R.I. The Empirical Basis for Modelling Glacial Erosion Rates. Nat. Commun. 2020, 11, 759. [Google Scholar] [CrossRef]
- Irvine-Fynn, T.D.L.; Hodson, A.J.; Moorman, B.J.; Vatne, G.; Hubbard, A.L. Polythermal glacier hydrology: A review. Rev. Geophys. 2011, 49. [Google Scholar] [CrossRef]
- Clason, C.; Mair, D.W.F.; Burgess, D.O.; Nienow, P.W. Modelling the Delivery of Supraglacial Meltwater to the Ice/Bed Interface: Application to Southwest Devon Ice Cap, Nunavut, Canada. J. Glaciol. 2012, 58, 361–374. [Google Scholar] [CrossRef]
- Hambrey, M.J.; Glasser, N.F. Discriminating Glacier Thermal and Dynamic Regimes in the Sedimentary Record. Sediment. Geol. 2012, 251–252, 1–33. [Google Scholar] [CrossRef]
- Pittard, M.L.; Galton-Fenzi, B.K.; Roberts, J.L.; Watson, C.S. Organization of Ice Flow by Localized Regions of Elevated Geothermal Heat Flux. Geophys. Res. Lett. 2016, 43, 3342–3350. [Google Scholar] [CrossRef]
- Marmoni, G.M.; Martino, S.; Salvatore, M.C.; Gaeta, M.; Perinelli, C.; Scarascia Mugnozza, G.; Baroni, C. Numerical Modelling of Geothermal Heat Flux and Ice Velocity Influencing the Thermal Conditions of the Priestley Glacier Trough (Northern Victoria Land, Antarctica). Geomorphology 2021, 394, 107959. [Google Scholar] [CrossRef]
- Pettersson, R.; Jansson, P.; Blatter, H. Spatial Variability in Water Content at the Cold-Temperate Transition Surface of the Polythermal Storglaciären, Sweden: Spatial Variability in Water Content. J. Geophys. Res. 2004, 109, F02009. [Google Scholar] [CrossRef]
- Vatne, G.; Irvine-Fynn, T.D.L. Morphological Dynamics of an Englacial Channel. Hydrol. Earth Syst. Sci. 2016, 20, 2947–2964. [Google Scholar] [CrossRef]
- Badgeley, J.A.; Pettit, E.C.; Carr, C.G.; Tulaczyk, S.; Mikucki, J.A.; Lyons, W.B.; MIDGE Science Team. An Englacial Hydrologic System of Brine within a Cold Glacier: Blood Falls, McMurdo Dry Valleys, Antarctica. J. Glaciol. 2017, 63, 387–400. [Google Scholar] [CrossRef]
- Temminghoff, M.; Benn, D.I.; Gulley, J.D.; Sevestre, H. Characterization of the Englacial and Subglacial Drainage System in a High Arctic Cold Glacier by Speleological Mapping and Ground-Penetrating Radar. Geogr. Ann. Ser. A Phys. Geogr. 2019, 101, 98–117. [Google Scholar] [CrossRef]
- Hansen, L.U.; Piotrowski, J.A.; Benn, D.I.; Sevestre, H. A Cross-Validated Three-Dimensional Model of an Englacial and Subglacial Drainage System in a High-Arctic Glacier. J. Glaciol. 2020, 66, 278–290. [Google Scholar] [CrossRef]
- Bergstrom, A.; Gooseff, M.N.; Singley, J.G.; Cohen, M.J.; Welch, K.A. Nutrient Uptake in the Supraglacial Stream Network of an Antarctic Glacier. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005679. [Google Scholar] [CrossRef]
- Bagshaw, E.A.; Tranter, M.; Wadham, J.L.; Fountain, A.G.; Basagic, H. Dynamic Behaviour of Supraglacial Lakes on Cold Polar Glaciers: Canada Glacier, McMurdo Dry Valleys, Antarctica. J. Glaciol. 2010, 56, 366–368. [Google Scholar] [CrossRef]
- Miller, O.L.; Solomon, D.K.; Miège, C.; Koenig, L.S.; Forster, R.R.; Montgomery, L.N.; Schmerr, N.; Ligtenberg, S.R.M.; Legchenko, A.; Brucker, L. Hydraulic Conductivity of a Firn Aquifer in Southeast Greenland. Front. Earth Sci. 2017, 5, 38. [Google Scholar] [CrossRef]
- Montgomery, L.N.; Schmerr, N.; Burdick, S.; Forster, R.R.; Koenig, L.; Legchenko, A.; Ligtenberg, S.; Miège, C.; Miller, O.L.; Solomon, D.K. Investigation of Firn Aquifer Structure in Southeastern Greenland Using Active Source Seismology. Front. Earth Sci. 2017, 5, 10. [Google Scholar] [CrossRef]
- Kendrick, A.K.; Schroeder, D.M.; Chu, W.; Young, T.J.; Christoffersen, P.; Todd, J.; Doyle, S.H.; Box, J.E.; Hubbard, A.; Hubbard, B.; et al. Surface Meltwater Impounded by Seasonal Englacial Storage in West Greenland. Geophys. Res. Lett. 2018, 45, 10474–10481. [Google Scholar] [CrossRef]
- Fountain, A.G. Effect of snow and firn hydrology on the physical and chemical characteristics of glacial runoff. Hydrol. Process. 1996, 10, 509–521. [Google Scholar] [CrossRef]
- Cook, J.M.; Hodson, A.J.; Irvine-Fynn, T.D.L. Supraglacial Weathering Crust Dynamics Inferred from Cryoconite Hole Hydrology: Weathering Crust Hydrology Observed in Cryoconite Holes. Hydrol. Process. 2016, 30, 433–446. [Google Scholar] [CrossRef]
- Hoffman, M.J.; Fountain, A.G.; Liston, G.E. Near-Surface Internal Melting: A Substantial Mass Loss on Antarctic Dry Valley Glaciers. J. Glaciol. 2014, 60, 361–374. [Google Scholar] [CrossRef]
- Christner, B.C.; Lavender, H.F.; Davis, C.L.; Oliver, E.E.; Neuhaus, S.U.; Myers, K.F.; Hagedorn, B.; Tulaczyk, S.M.; Doran, P.T.; Stone, W.C. Microbial Processes in the Weathering Crust Aquifer of a Temperate Glacier. Cryosphere 2018, 12, 3653–3669. [Google Scholar] [CrossRef]
- Irvine-Fynn, T.D.L.; Edwards, A.; Stevens, I.T.; Mitchell, A.C.; Bunting, P.; Box, J.E.; Cameron, K.A.; Cook, J.M.; Naegeli, K.; Rassner, S.M.E.; et al. Storage and Export of Microbial Biomass across the Western Greenland Ice Sheet. Nat. Commun. 2021, 12, 3960. [Google Scholar] [CrossRef]
- Christianson, K.; Kohler, J.; Alley, R.B.; Nuth, C.; Pelt, W.J.J. Dynamic Perennial Firn Aquifer on an Arctic Glacier. Geophys. Res. Lett. 2015, 42, 1418–1426. [Google Scholar] [CrossRef]
- Stevens, I.T.; Irvine-Fynn, T.D.L.; Porter, P.R.; Cook, J.M.; Edwards, A.; Smart, M.; Moorman, B.J.; Hodson, A.J.; Mitchell, A.C. Near-Surface Hydraulic Conductivity of Northern Hemisphere Glaciers. Hydrol. Process. 2018, 32, 850–865. [Google Scholar] [CrossRef]
- Fountain, A.G.; Tranter, M.; Nylen, T.H.; Lewis, K.J.; Mueller, D.R. Evolution of Cryoconite Holes and Their Contribution to Meltwater Runoff from Glaciers in the McMurdo Dry Valleys, Antarctica. J. Glaciol. 2004, 50, 35–45. [Google Scholar] [CrossRef]
- MacDonell, S.; Fitzsimons, S. The Formation and Hydrological Significance of Cryoconite Holes. Progress. Phys. Geogr. Earth Environ. 2008, 32, 595–610. [Google Scholar] [CrossRef]
- Cook, J.; Edwards, A.; Takeuchi, N.; Irvine-Fynn, T. Cryoconite: The Dark Biological Secret of the Cryosphere. Progress. Phys. Geogr. Earth Environ. 2016, 40, 66–111. [Google Scholar] [CrossRef]
- Pitcher, L.H.; Smith, L.C. Supraglacial Streams and Rivers. Annu. Rev. Earth Planet. Sci. 2019, 47, 421–452. [Google Scholar] [CrossRef]
- Dunham, E.C.; Dore, J.E.; Skidmore, M.L.; Roden, E.E.; Boyd, E.S. Lithogenic Hydrogen Supports Microbial Primary Production in Subglacial and Proglacial Environments. Proc. Natl. Acad. Sci. USA 2021, 118, e2007051117. [Google Scholar] [CrossRef] [PubMed]
- Bottrell, S.H.; Tranter, M. Sulphide Oxidation under Partially Anoxic Conditions at the Bed of the Haut Glacier d’Arolla, Switzerland. Hydrol. Process. 2002, 16, 2363–2368. [Google Scholar] [CrossRef]
- Wadham, J.L.; Tranter, M.; Skidmore, M.; Hodson, A.J.; Priscu, J.; Lyons, W.B.; Sharp, M.; Wynn, P.; Jackson, M. Biogeochemical Weathering under Ice: Size Matters: Glacial Biogeochemical Weathering. Glob. Biogeochem. Cycles 2010, 24, GB3025. [Google Scholar] [CrossRef]
- Maccario, L.; Sanguino, L.; Vogel, T.M.; Larose, C. Snow and Ice Ecosystems: Not so Extreme. Res. Microbiol. 2015, 166, 782–795. [Google Scholar] [CrossRef]
- Fountain, A.G.; Jacobel, R.W.; Schlichting, R.; Jansson, P. Fractures as the Main Pathways of Water Flow in Temperate Glaciers. Nature 2005, 433, 618–621. [Google Scholar] [CrossRef]
- McGrath, D.; Colgan, W.; Steffen, K.; Lauffenburger, P.; Balog, J. Assessing the Summer Water Budget of a Moulin Basin in the Sermeq Avannarleq Ablation Region, Greenland Ice Sheet. J. Glaciol. 2011, 57, 954–964. [Google Scholar] [CrossRef]
- Lampkin, D.J.; Amador, N.; Parizek, B.R.; Farness, K.; Jezek, K. Drainage from Water-filled Crevasses along the Margins of Jakobshavn Isbræ: A Potential Catalyst for Catchment Expansion. J. Geophys. Res. Earth Surf. 2013, 118, 795–813. [Google Scholar] [CrossRef]
- Llubes, M.; Lanseau, C.; Rémy, F. Relations between Basal Condition, Subglacial Hydrological Networks and Geothermal Flux in Antarctica. Earth Planet. Sci. Lett. 2006, 241, 655–662. [Google Scholar] [CrossRef]
- Fisher, A.T.; Mankoff, K.D.; Tulaczyk, S.M.; Tyler, S.W.; Foley, N.; The WISSARD Science Team. High Geothermal Heat Flux Measured below the West Antarctic Ice Sheet. Sci. Adv. 2015, 1, e1500093. [Google Scholar] [CrossRef]
- Lösing, M.; Ebbing, J.; Szwillus, W. Geothermal Heat Flux in Antarctica: Assessing Models and Observations by Bayesian Inversion. Front. Earth Sci. 2020, 8, 105. [Google Scholar] [CrossRef]
- Beem, L.H.; Jezek, K.C.; Van Der Veen, C.J. Basal Melt Rates beneath Whillans Ice Stream, West Antarctica. J. Glaciol. 2010, 56, 647–654. [Google Scholar] [CrossRef]
- Nanni, U.; Gimbert, F.; Roux, P.; Lecointre, A. Observing the Subglacial Hydrology Network and Its Dynamics with a Dense Seismic Array. Proc. Natl. Acad. Sci. USA 2021, 118, e2023757118. [Google Scholar] [CrossRef]
- Scholzen, C.; Schuler, T.V.; Gilbert, A. Sensitivity of Subglacial Drainage to Water Supply Distribution at the Kongsfjord Basin, Svalbard. Cryosphere 2021, 15, 2719–2738. [Google Scholar] [CrossRef]
- Ronayne, M.J.; Houghton, T.B.; Stednick, J.D. Field Characterization of Hydraulic Conductivity in a Heterogeneous Alpine Glacial Till. J. Hydrol. 2012, 458–459, 103–109. [Google Scholar] [CrossRef]
- Siegert, M.J.; Ross, N.; Le Brocq, A.M. Recent Advances in Understanding Antarctic Subglacial Lakes and Hydrology. Philos. Trans. R. Soc. A 2016, 374, 20140306. [Google Scholar] [CrossRef]
- Bowling, J.S.; Livingstone, S.J.; Sole, A.J.; Chu, W. Distribution and Dynamics of Greenland Subglacial Lakes. Nat. Commun. 2019, 10, 2810. [Google Scholar] [CrossRef]
- Jansson, P.; Hock, R.; Schneider, T. The Concept of Glacier Storage: A Review. J. Hydrol. 2003, 282, 116–129. [Google Scholar] [CrossRef]
- Vaňková, I.; Voytenko, D.; Nicholls, K.W.; Xie, S.; Parizek, B.R.; Holland, D.M. Vertical Structure of Diurnal Englacial Hydrology Cycle at Helheim Glacier, East Greenland. Geophys. Res. Lett. 2018, 45, 8352–8362. [Google Scholar] [CrossRef]
- Dunse, T.; Schellenberger, T.; Hagen, J.O.; Kääb, A.; Schuler, T.V.; Reijmer, C.H. Glacier-Surge Mechanisms Promoted by a Hydro-Thermodynamic Feedback to Summer Melt. Cryosphere 2015, 9, 197–215. [Google Scholar] [CrossRef]
- Cooper, M.G.; Smith, L.C.; Rennermalm, A.K.; Miège, C.; Pitcher, L.H.; Ryan, J.C.; Yang, K.; Cooley, S.W. Meltwater Storage in Low-Density near-Surface Bare Ice in the Greenland Ice Sheet Ablation Zone. Cryosphere 2018, 12, 955–970. [Google Scholar] [CrossRef]
- Thøgersen, K.; Gilbert, A.; Schuler, T.V.; Malthe-Sørenssen, A. Rate-and-State Friction Explains Glacier Surge Propagation. Nat. Commun. 2019, 10, 2823. [Google Scholar] [CrossRef] [PubMed]
- Harrison, W.D. Temperature of a Temperate Glacier. J. Glaciol. 1972, 11, 15–29. [Google Scholar] [CrossRef]
- Harrison, W.D.; Raymond, C.F. Impurities and Their Distribution in Temperate Glacier Ice. J. Glaciol. 1976, 16, 173–181. [Google Scholar] [CrossRef]
- Phillips, T.; Rajaram, H.; Steffen, K. Cryo-Hydrologic Warming: A Potential Mechanism for Rapid Thermal Response of Ice Sheets: Rapid Thermal Response of Ice Sheets. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Pitcher, L.H.; Smith, L.C.; Gleason, C.J.; Miège, C.; Ryan, J.C.; Hagedorn, B.; As, D.; Chu, W.; Forster, R.R. Direct Observation of Winter Meltwater Drainage From the Greenland Ice Sheet. Geophys. Res. Lett. 2020, 47, e2019GL086521. [Google Scholar] [CrossRef]
- Hindshaw, R.S.; Tipper, E.T.; Reynolds, B.C.; Lemarchand, E.; Wiederhold, J.G.; Magnusson, J.; Bernasconi, S.M.; Kretzschmar, R.; Bourdon, B. Hydrological Control of Stream Water Chemistry in a Glacial Catchment (Damma Glacier, Switzerland). Chem. Geol. 2011, 285, 215–230. [Google Scholar] [CrossRef]
- Foreman, C.M.; Sattler, B.; Mikucki, J.A.; Porazinska, D.L.; Priscu, J.C. Metabolic Activity and Diversity of Cryoconites in the Taylor Valley, Antarctica: Activity in Antarctic Cryoconites. J. Geophys. Res. 2007, 112, G04S32. [Google Scholar] [CrossRef]
- Forster, R.R.; Box, J.E.; van den Broeke, M.R.; Miège, C.; Burgess, E.W.; van Angelen, J.H.; Lenaerts, J.T.M.; Koenig, L.S.; Paden, J.; Lewis, C.; et al. Extensive Liquid Meltwater Storage in Firn within the Greenland Ice Sheet. Nat. Geosci. 2014, 7, 95–98. [Google Scholar] [CrossRef]
- Koenig, L.S.; Miège, C.; Forster, R.R.; Brucker, L. Initial in Situ Measurements of Perennial Meltwater Storage in the Greenland Firn Aquifer: Measurements of Greenland Aquifer. Geophys. Res. Lett. 2014, 41, 81–85. [Google Scholar] [CrossRef]
- Chu, W.; Schroeder, D.M.; Siegfried, M.R. Retrieval of Englacial Firn Aquifer Thickness From Ice-Penetrating Radar Sounding in Southeastern Greenland. Geophys. Res. Lett. 2018, 45, 11770–11778. [Google Scholar] [CrossRef]
- Miller, O.; Solomon, D.K.; Miège, C.; Koenig, L.; Forster, R.; Schmerr, N.; Ligtenberg, S.R.M.; Legchenko, A.; Voss, C.I.; Montgomery, L.; et al. Hydrology of a Perennial Firn Aquifer in Southeast Greenland: An Overview Driven by Field Data. Water Resour. Res. 2020, 56, e2019WR026348. [Google Scholar] [CrossRef]
- Seaberg, S.Z.; Seaberg, J.Z.; Hooke, R.L.; Wiberg, D.W. Character of the Englacial and Subglacial Drainage System in the Lower Part of the Ablation Area of Storglaciären, Sweden, as Revealed by Dye-Trace Studies. J. Glaciol. 1988, 34, 217–227. [Google Scholar] [CrossRef]
- Nienow, P.; Sharp, M.; Willis, I. Temporal Switching Between Englacial and Subglacial Drainage Pathways: Dye Tracer Evidence from the Haut Glacier D’arolla, Switzerland. Geogr. Ann. Ser. A Phys. Geogr. 1996, 78, 51–60. [Google Scholar] [CrossRef]
- Nye, J.F. The Geometry of Water Veins and Nodes in Polycrystalline Ice. J. Glaciol. 1989, 35, 17–22. [Google Scholar] [CrossRef]
- Fountain, A.G. Geometry and Flow Conditions of Subglacial Water at South Cascade Glacier, Washington State, U.S.A.; an Analysis of Tracer Injections. J. Glaciol. 1993, 39, 143–156. [Google Scholar] [CrossRef]
- Werder, M.A.; Loye, A.; Funk, M. Dye Tracing a Jökulhlaup: I. Subglacial Water Transit Speed and Water-Storage Mechanism. J. Glaciol. 2009, 55, 889–898. [Google Scholar] [CrossRef]
- Bartholomew, I.; Nienow, P.; Mair, D.; Hubbard, A.; King, M.A.; Sole, A. Seasonal Evolution of Subglacial Drainage and Acceleration in a Greenland Outlet Glacier. Nat. Geosci. 2010, 3, 408–411. [Google Scholar] [CrossRef]
- Wingham, D.J.; Siegert, M.J.; Shepherd, A.; Muir, A.S. Rapid Discharge Connects Antarctic Subglacial Lakes. Nature 2006, 440, 1033–1036. [Google Scholar] [CrossRef]
- Tranter, M.; Fountain, A.G.; Fritsen, C.H.; Berry Lyons, W.; Priscu, J.C.; Statham, P.J.; Welch, K.A. Extreme Hydrochemical Conditions in Natural Microcosms Entombed within Antarctic Ice. Hydrol. Process. 2004, 18, 379–387. [Google Scholar] [CrossRef]
- Fountain, A.G. The Storage of Water in, and Hydraulic Characteristics of, the Firn of South Cascade Glacier, Washington State, USA. Ann. Glaciol. 1989, 13, 69–75. [Google Scholar] [CrossRef]
- van Wessem, J.M.; Steger, C.R.; Wever, N.; van den Broeke, M.R. Modelling Perennial Firn Aquifers in the Antarctic Peninsula (1979–2016); Snow/Snow Physics: Seattle, WA, USA, 2020. [Google Scholar]
- Barletta, R.E.; Priscu, J.C.; Mader, H.M.; Jones, W.L.; Roe, C.H. Chemical Analysis of Ice Vein Microenvironments: II. Analysis of Glacial Samples from Greenland and Antarctica. J. Glaciol. 2012, 58, 1109–1118. [Google Scholar] [CrossRef]
- Neff, J.C.; Holland, E.A.; Dentener, F.J.; McDowell, W.H.; Russell, K.M. The Origin, Composition and Rates of Organic Nitrogen Deposition: A Missing Piece of the Nitrogen Cycle? In The Nitrogen. Cycle at Regional to Global Scales; Boyer, E.W., Howarth, R.W., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 99–136. ISBN 978-90-481-6086-0. [Google Scholar]
- Hodson, A.J.; Mumford, P.N.; Kohler, J.; Wynn, P.M. The High Arctic Glacial Ecosystem: New Insights from Nutrient Budgets. Biogeochemistry 2005, 72, 233–256. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, J.; Wang, F.; Mark, L.; Xu, J.; Armstrong, D.; Li, C.; Zhang, Y.; Kang, S. Mercury Distribution and Deposition in Glacier Snow over Western China. Environ. Sci. Technol. 2012, 46, 5404–5413. [Google Scholar] [CrossRef] [PubMed]
- Björkman, M.; Kühnel, R.; Partridge, D.; Roberts, T.; Aas, W.; Mazzola, M.; Viola, A.; Hodson, A.; Ström, J.; Isaksson, E. Nitrate Dry Deposition in Svalbard. Tellus B Chem. Phys. Meteorol. 2013, 65, 19071. [Google Scholar] [CrossRef]
- McCutcheon, J.; Lutz, S.; Williamson, C.; Cook, J.M.; Tedstone, A.J.; Vanderstraeten, A.; Wilson, S.A.; Stockdale, A.; Bonneville, S.; Anesio, A.M.; et al. Mineral Phosphorus Drives Glacier Algal Blooms on the Greenland Ice Sheet. Nat. Commun. 2021, 12, 570. [Google Scholar] [CrossRef]
- Nanus, L.; Campbell, D.H.; Ingersoll, G.P.; Clow, D.W.; Alisa Mast, M. Atmospheric Deposition Maps for the Rocky Mountains. Atmos. Environ. 2003, 37, 4881–4892. [Google Scholar] [CrossRef]
- Ginoux, P.; Prospero, J.M.; Gill, T.E.; Hsu, N.C.; Zhao, M. Global-Scale Attribution of Anthropogenic and Natural Dust Sources and Their Emission Rates Based on MODIS Deep Blue Aerosol Products: Anthropogenic and Natural Dust Sources. Rev. Geophys. 2012, 50, RG3005. [Google Scholar] [CrossRef]
- Kühnel, R.; Björkman, M.P.; Vega, C.P.; Hodson, A.; Isaksson, E.; Ström, J. Reactive Nitrogen and Sulphate Wet Deposition at Zeppelin Station, Ny-Ålesund, Svalbard. Polar Res. 2013, 32, 19136. [Google Scholar] [CrossRef]
- Fortner, S.K.; Lyons, W.B. Dissolved Trace and Minor Elements in Cryoconite Holes and Supraglacial Streams, Canada Glacier, Antarctica. Front. Earth Sci. 2018, 6, 31. [Google Scholar] [CrossRef]
- Stibal, M.; Lawson, E.C.; Lis, G.P.; Mak, K.M.; Wadham, J.L.; Anesio, A.M. Organic Matter Content and Quality in Supraglacial Debris across the Ablation Zone of the Greenland Ice Sheet. Ann. Glaciol. 2010, 51, 1–8. [Google Scholar] [CrossRef]
- Takeuchi, N.; Fujisawa, Y.; Kadota, T.; Tanaka, S.; Miyairi, M.; Shirakawa, T.; Kusaka, R.; Fedorov, A.N.; Konstantinov, P.; Ohata, T. The Effect of Impurities on the Surface Melt of a Glacier in the Suntar-Khayata Mountain Range, Russian Siberia. Front. Earth Sci. 2015, 3. [Google Scholar] [CrossRef]
- Stibal, M.; Bradley, J.A.; Box, J.E. Ecological Modeling of the Supraglacial Ecosystem: A Process-Based Perspective. Front. Earth Sci. 2017, 5, 52. [Google Scholar] [CrossRef]
- Tedstone, A.J.; Bamber, J.L.; Cook, J.M.; Williamson, C.J.; Fettweis, X.; Hodson, A.J.; Tranter, M. Dark Ice Dynamics of the South-West Greenland Ice Sheet. Cryosphere 2017, 11, 2491–2506. [Google Scholar] [CrossRef]
- Franzetti, A.; Tatangelo, V.; Gandolfi, I.; Bertolini, V.; Bestetti, G.; Diolaiuti, G.; D’Agata, C.; Mihalcea, C.; Smiraglia, C.; Ambrosini, R. Bacterial Community Structure on Two Alpine Debris-Covered Glaciers and Biogeography of Polaromonas Phylotypes. ISME J. 2013, 7, 1483–1492. [Google Scholar] [CrossRef]
- Rime, T.; Hartmann, M.; Frey, B. Potential Sources of Microbial Colonizers in an Initial Soil Ecosystem after Retreat of an Alpine Glacier. ISME J. 2016, 10, 1625–1641. [Google Scholar] [CrossRef]
- Darcy, J.L.; King, A.J.; Gendron, E.M.S.; Schmidt, S.K. Spatial Autocorrelation of Microbial Communities atop a Debris-Covered Glacier Is Evidence of a Supraglacial Chronosequence. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Liu, Y.; Vick-Majors, T.J.; Priscu, J.C.; Yao, T.; Kang, S.; Liu, K.; Cong, Z.; Xiong, J.; Li, Y. Biogeography of Cryoconite Bacterial Communities on Glaciers of the Tibetan Plateau. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Lutz, S.; Anesio, A.M.; Edwards, A.; Benning, L.G. Linking Microbial Diversity and Functionality of Arctic Glacial Surface Habitats: Arctic Glacial Surface Habitats. Environ. Microbiol. 2017, 19, 551–565. [Google Scholar] [CrossRef]
- Segawa, T.; Yonezawa, T.; Edwards, A.; Akiyoshi, A.; Tanaka, S.; Uetake, J.; Irvine-Fynn, T.; Fukui, K.; Li, Z.; Takeuchi, N. Biogeography of Cryoconite Forming Cyanobacteria on Polar and Asian Glaciers. J. Biogeogr. 2017, 44, 2849–2861. [Google Scholar] [CrossRef]
- Havig, J.R.; Hamilton, T.L. Snow Algae Drive Productivity and Weathering at Volcanic Rock-Hosted Glaciers. Geochim. Et Cosmochim. Acta 2019, 247, 220–242. [Google Scholar] [CrossRef]
- Kim, K.; Choi, W.; Hoffmann, M.R.; Yoon, H.-I.; Park, B.-K. Photoreductive Dissolution of Iron Oxides Trapped in Ice and Its Environmental Implications. Environ. Sci. Technol. 2010, 44, 4142–4148. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.T.; Williamson, C.J.; Sgouridis, F.; Tedstone, A.J.; McCutcheon, J.; Cook, J.M.; Poniecka, E.; Yallop, M.L.; Tranter, M.; Anesio, A.M.; et al. Dissolved Organic Nutrients Dominate Melting Surface Ice of the Dark Zone (Greenland Ice Sheet). Biogeosciences 2019, 16, 3283–3296. [Google Scholar] [CrossRef]
- Wharton, R.A.; McKay, C.P.; Simmons, G.M.; Parker, B.C. Cryoconite Holes on Glaciers. BioScience 1985, 35, 499–503. [Google Scholar] [CrossRef]
- Bagshaw, E.A.; Tranter, M.; Fountain, A.G.; Welch, K.; Basagic, H.J.; Lyons, W.B. Do Cryoconite Holes Have the Potential to Be Significant Sources of C, N, and P to Downstream Depauperate Ecosystems of Taylor Valley, Antarctica? Arct. Antarct. Alp. Res. 2013, 45, 440–454. [Google Scholar] [CrossRef]
- Bagshaw, E.A.; Tranter, M.; Wadham, J.L.; Fountain, A.G.; Mowlem, M. High-Resolution Monitoring Reveals Dissolved Oxygen Dynamics in an Antarctic Cryoconite Hole. Hydrol. Process. 2011, 25, 2868–2877. [Google Scholar] [CrossRef]
- Poniecka, E.A.; Bagshaw, E.A.; Tranter, M.; Sass, H.; Williamson, C.J.; Anesio, A.M.; Black and Bloom Team. Rapid Development of Anoxic Niches in Supraglacial Ecosystems. Arct. Antarct. Alp. Res. 2018, 50, S100015. [Google Scholar] [CrossRef]
- Williamson, C.J.; Cameron, K.A.; Cook, J.M.; Zarsky, J.D.; Stibal, M.; Edwards, A. Glacier Algae: A Dark Past and a Darker Future. Front. Microbiol. 2019, 10, 524. [Google Scholar] [CrossRef]
- Nash, M.V.; Anesio, A.M.; Barker, G.; Tranter, M.; Varliero, G.; Eloe-Fadrosh, E.A.; Nielsen, T.; Turpin-Jelfs, T.; Benning, L.G.; Sánchez-Baracaldo, P. Metagenomic Insights into Diazotrophic Communities across Arctic Glacier Forefields. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef]
- Varliero, G.; Anesio, A.M.; Barker, G.L.A. A Taxon-Wise Insight Into Rock Weathering and Nitrogen Fixation Functional Profiles of Proglacial Systems. Front. Microbiol. 2021, 12, 627437. [Google Scholar] [CrossRef]
- Telling, J.; Anesio, A.M.; Tranter, M.; Irvine-Fynn, T.; Hodson, A.; Butler, C.; Wadham, J. Nitrogen Fixation on Arctic Glaciers, Svalbard. J. Geophys. Res. 2011, 116, G03039. [Google Scholar] [CrossRef]
- Telling, J.; Stibal, M.; Anesio, A.M.; Tranter, M.; Nias, I.; Cook, J.; Bellas, C.; Lis, G.; Wadham, J.L.; Sole, A.; et al. Microbial Nitrogen Cycling on the Greenland Ice Sheet. Biogeosciences 2012, 9, 2431–2442. [Google Scholar] [CrossRef]
- Clark, S.C.; Barnes, R.T.; Oleksy, I.A.; Baron, J.S.; Hastings, M.G. Persistent Nitrate in Alpine Waters with Changing Atmospheric Deposition and Warming Trends. Environ. Sci. Technol. 2021, 55, 14946–14956. [Google Scholar] [CrossRef]
- Stibal, M.; Anesio, A.M.; Blues, C.J.D.; Tranter, M. Phosphatase Activity and Organic Phosphorus Turnover on a High Arctic Glacier. Biogeosciences 2009, 6, 913–922. [Google Scholar] [CrossRef]
- Stibal, M.; Tranter, M.; Telling, J.; Benning, L.G. Speciation, Phase Association and Potential Bioavailability of Phosphorus on a Svalbard Glacier. Biogeochemistry 2008, 90, 1–13. [Google Scholar] [CrossRef]
- Smith, H.J.; Foster, R.A.; McKnight, D.M.; Lisle, J.T.; Littmann, S.; Kuypers, M.M.M.; Foreman, C.M. Microbial Formation of Labile Organic Carbon in Antarctic Glacial Environments. Nat. Geosci. 2017, 10, 356–359. [Google Scholar] [CrossRef]
- Sanyal, A.; Antony, R.; Samui, G.; Thamban, M. Microbial Communities and Their Potential for Degradation of Dissolved Organic Carbon in Cryoconite Hole Environments of Himalaya and Antarctica. Microbiol. Res. 2018, 208, 32–42. [Google Scholar] [CrossRef]
- Andrews, M.G.; Jacobson, A.D.; Osburn, M.R.; Flynn, T.M. Dissolved Carbon Dynamics in Meltwaters From the Russell Glacier, Greenland Ice Sheet. J. Geophys. Res. Biogeosci. 2018, 123, 2922–2940. [Google Scholar] [CrossRef]
- Samui, G.; Antony, R.; Thamban, M. Chemical Characteristics of Hydrologically Distinct Cryoconite Holes in Coastal Antarctica. Ann. Glaciol. 2018, 59, 69–76. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, T.; Kang, S.; Sprenger, M.; Tao, S.; Du, W.; Yang, J.; Wang, F.; Meng, W. Effects of Black Carbon and Mineral Dust on Glacial Melting on the Muz Taw Glacier, Central Asia. Sci. Total Environ. 2020, 740, 140056. [Google Scholar] [CrossRef]
- Benning, L.G.; Anesio, A.M.; Lutz, S.; Tranter, M. Biological Impact on Greenland’s Albedo. Nat. Geosci. 2014, 7, 691. [Google Scholar] [CrossRef]
- Hotaling, S.; Lutz, S.; Dial, R.J.; Anesio, A.M.; Benning, L.G.; Fountain, A.G.; Kelley, J.L.; McCutcheon, J.; Skiles, S.M.; Takeuchi, N.; et al. Biological Albedo Reduction on Ice Sheets, Glaciers, and Snowfields. Earth-Sci. Rev. 2021, 220, 103728. [Google Scholar] [CrossRef]
- Hawkings, J.R.; Wadham, J.L.; Tranter, M.; Lawson, E.; Sole, A.; Cowton, T.; Tedstone, A.J.; Bartholomew, I.; Nienow, P.; Chandler, D.; et al. The Effect of Warming Climate on Nutrient and Solute Export from the Greenland Ice Sheet. Geochem. Persp. Let. 2015, 94–104. [Google Scholar] [CrossRef]
- Musilova, M.; Tranter, M.; Bennett, S.A.; Wadham, J.; Anesio, A.M. Stable Microbial Community Composition on the Greenland Ice Sheet. Front. Microbiol. 2015, 6, 193. [Google Scholar] [CrossRef] [PubMed]
- Varliero, G.; Holland, A.; Barker, G.L.A.; Yallop, M.L.; Fountain, A.G.; Anesio, A.M. Glacier Clear Ice Bands Indicate Englacial Channel Microbial Distribution. J. Glaciol. 2021, 67, 811–823. [Google Scholar] [CrossRef]
- Martinez-Alonso, E.; Pena-Perez, S.; Serrano, S.; Garcia-Lopez, E.; Alcazar, A.; Cid, C. Taxonomic and Functional Characterization of a Microbial Community from a Volcanic Englacial Ecosystem in Deception Island, Antarctica. Sci. Rep. 2019, 9, 12158. [Google Scholar] [CrossRef]
- Bakermans, C.; Skidmore, M.L. Microbial Metabolism in Ice and Brine at −5°C: Microbial Metabolism in Ice and Brine at −5°C. Environ. Microbiol. 2011, 13, 2269–2278. [Google Scholar] [CrossRef]
- Dani, K.G.S.; Mader, H.M.; Wolff, E.W.; Wadham, J.L. Modelling the Liquid-Water Vein System within Polar Ice Sheets as a Potential Microbial Habitat. Earth Planet. Sci. Lett. 2012, 333–334, 238–249. [Google Scholar] [CrossRef]
- Mader, H.M.; Pettitt, M.E.; Wadham, J.L.; Wolff, E.W.; Parkes, R.J. Subsurface Ice as a Microbial Habitat. Geology 2006, 34, 169. [Google Scholar] [CrossRef]
- Rohde, R.A.; Price, P.B. Diffusion-Controlled Metabolism for Long-Term Survival of Single Isolated Microorganisms Trapped within Ice Crystals. Proc. Natl. Acad. Sci. USA 2007, 104, 16592–16597. [Google Scholar] [CrossRef]
- Li, Y.; Cha, Q.-Q.; Dang, Y.-R.; Chen, X.-L.; Wang, M.; McMinn, A.; Espina, G.; Zhang, Y.-Z.; Blamey, J.M.; Qin, Q.-L. Reconstruction of the Functional Ecosystem in the High Light, Low Temperature Union Glacier Region, Antarctica. Front. Microbiol. 2019, 10, 2408. [Google Scholar] [CrossRef]
- Mitchell, A.C.; Lafrenière, M.J.; Skidmore, M.L.; Boyd, E.S. Influence of Bedrock Mineral Composition on Microbial Diversity in a Subglacial Environment. Geology 2013, 41, 855–858. [Google Scholar] [CrossRef]
- Skidmore, M.; Anderson, S.P.; Sharp, M.; Foght, J.; Lanoil, B.D. Comparison of Microbial Community Compositions of Two Subglacial Environments Reveals a Possible Role for Microbes in Chemical Weathering Processes. Appl. Environ. Microbiol. 2005, 71, 6986–6997. [Google Scholar] [CrossRef]
- Tranter, M.; Sharp, M.J.; Lamb, H.R.; Brown, G.H.; Hubbard, B.P.; Willis, I.C. Geochemical Weathering at the Bed of Haut Glacier d’Arolla, Switzerland?A New Model. Hydrol. Process. 2002, 16, 959–993. [Google Scholar] [CrossRef]
- Anderson, S.P. Glaciers Show Direct Linkage between Erosion Rate and Chemical Weathering Fluxes. Geomorphology 2005, 67, 147–157. [Google Scholar] [CrossRef]
- Macdonald, M.L.; Wadham, J.L.; Telling, J.; Skidmore, M.L. Glacial Erosion Liberates Lithologic Energy Sources for Microbes and Acidity for Chemical Weathering Beneath Glaciers and Ice Sheets. Front. Earth Sci. 2018, 6, 212. [Google Scholar] [CrossRef]
- Thór Marteinsson, V.; Rúnarsson, Á.; Stefánsson, A.; Thorsteinsson, T.; Jóhannesson, T.; Magnússon, S.H.; Reynisson, E.; Einarsson, B.; Wade, N.; Morrison, H.G.; et al. Microbial Communities in the Subglacial Waters of the Vatnajökull Ice Cap, Iceland. ISME J. 2013, 7, 427–437. [Google Scholar] [CrossRef]
- Achberger, A.M.; Michaud, A.B.; Vick-Majors, T.J.; Christner, B.C.; Skidmore, M.L.; Priscu, J.C.; Tranter, M. Microbiology of Subglacial Environments. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 83–110. ISBN 978-3-319-57057-0. [Google Scholar]
- Elberling, B. Temperature and Oxygen Control on Pyrite Oxidation in Frozen Mine Tailings. Cold Reg. Sci. Technol. 2005, 41, 121–133. [Google Scholar] [CrossRef]
- Nixon, S.L.; Telling, J.P.; Wadham, J.L.; Cockell, C.S. Viable Cold-Tolerant Iron-Reducing Microorganisms in Geographically Diverse Subglacial Environments. Biogeosciences 2017, 14, 1445–1455. [Google Scholar] [CrossRef]
- Hamilton, T.L.; Peters, J.W.; Skidmore, M.L.; Boyd, E.S. Molecular Evidence for an Active Endogenous Microbiome beneath Glacial Ice. ISME J. 2013, 7, 1402–1412. [Google Scholar] [CrossRef]
- Wynn, P.M.; Hodson, A.; Heaton, T. Chemical and Isotopic Switching within the Subglacial Environment of a High Arctic Glacier. Biogeochemistry 2006, 78, 173–193. [Google Scholar] [CrossRef]
- Wynn, P.M.; Hodson, A.J.; Heaton, T.H.E.; Chenery, S.R. Nitrate Production beneath a High Arctic Glacier, Svalbard. Chem. Geol. 2007, 244, 88–102. [Google Scholar] [CrossRef]
- Liermann, L.J.; Hausrath, E.M.; Anbar, A.D.; Brantley, S.L. Assimilatory and Dissimilatory Processes of Microorganisms Affecting Metals in the Environment. J. Anal. At. Spectrom. 2007, 22, 867. [Google Scholar] [CrossRef]
- Muyzer, G.; Stams, A.J.M. The Ecology and Biotechnology of Sulphate-Reducing Bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Stibal, M.; Wadham, J.L.; Lis, G.P.; Telling, J.; Pancost, R.D.; Dubnick, A.; Sharp, M.J.; Lawson, E.C.; Butler, C.E.H.; Hasan, F.; et al. Methanogenic Potential of Arctic and Antarctic Subglacial Environments with Contrasting Organic Carbon Sources. Glob. Change Biol. 2012, 18, 3332–3345. [Google Scholar] [CrossRef]
- Anesio, A.M.; Hodson, A.J.; Fritz, A.; Psenner, R.; Sattler, B. High Microbial Activity on Glaciers: Importance to the Global Carbon Cycle. Glob. Chang. Biol. 2009, 15, 955–960. [Google Scholar] [CrossRef]
- Price, P.B. A Habitat for Psychrophiles in Deep Antarctic Ice. Proc. Natl. Acad. Sci. USA 2000, 97, 1247–1251. [Google Scholar] [CrossRef]
- Mikucki, J.A.; Pearson, A.; Johnston, D.T.; Turchyn, A.V.; Farquhar, J.; Schrag, D.P.; Anbar, A.D.; Priscu, J.C.; Lee, P.A. A Contemporary Microbially Maintained Subglacial Ferrous "Ocean". ” Science 2009, 324, 397–400. [Google Scholar] [CrossRef]
- Lyons, W.B.; Mikucki, J.A.; German, L.A.; Welch, K.A.; Welch, S.A.; Gardner, C.B.; Tulaczyk, S.M.; Pettit, E.C.; Kowalski, J.; Dachwald, B. The Geochemistry of Englacial Brine From Taylor Glacier, Antarctica. J. Geophys. Res. Biogeosci. 2019, 124, 633–648. [Google Scholar] [CrossRef]
- Vick-Majors, T.J.; Achberger, A.M.; Michaud, A.B.; Priscu, J.C. Metabolic and Taxonomic Diversity in Antarctic Subglacial Environments. In Life in Extreme Environments; di Prisco, G., Edwards, H.G.M., Elster, J., Huiskes, A.H.L., Eds.; Cambridge University Press: Cambridge, UK, 2020; pp. 279–296. ISBN 978-1-108-68331-9. [Google Scholar]
- Telling, J.; Boyd, E.S.; Bone, N.; Jones, E.L.; Tranter, M.; MacFarlane, J.W.; Martin, P.G.; Wadham, J.L.; Lamarche-Gagnon, G.; Skidmore, M.L.; et al. Rock Comminution as a Source of Hydrogen for Subglacial Ecosystems. Nat. Geosci. 2015, 8, 851–855. [Google Scholar] [CrossRef]
- Greening, C.; Biswas, A.; Carere, C.R.; Jackson, C.J.; Taylor, M.C.; Stott, M.B.; Cook, G.M.; Morales, S.E. Genomic and Metagenomic Surveys of Hydrogenase Distribution Indicate H2 Is a Widely Utilised Energy Source for Microbial Growth and Survival. ISME J. 2016, 10, 761–777. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Y.; Lv, Y.; Yan, W.; Xiao, X.; Sun, B.; Ma, H. H2 Metabolism Revealed by Metagenomic Analysis of Subglacial Sediment from East Antarctica. J. Microbiol. 2019, 57, 1095–1104. [Google Scholar] [CrossRef]
- Holland, A.T.; Bergk Pinto, B.; Layton, R.; Williamson, C.J.; Anesio, A.M.; Vogel, T.M.; Larose, C.; Tranter, M. Over Winter Microbial Processes in a Svalbard Snow Pack: An Experimental Approach. Front. Microbiol. 2020, 11, 1029. [Google Scholar] [CrossRef]
- Kranzler, C.; Kessler, N.; Keren, N.; Shaked, Y. Enhanced Ferrihydrite Dissolution by a Unicellular, Planktonic Cyanobacterium: A Biological Contribution to Particulate Iron Bioavailability: Bio-Dissolution of Ferrihydrite by Cyanobacteria. Environ. Microbiol. 2016, 18, 5101–5111. [Google Scholar] [CrossRef]
- Koffman, B.G.; Yoder, M.F.; Methven, T.; Hanschka, L.; Sears, H.B.; Saylor, P.L.; Wallace, K.L. Glacial Dust Surpasses Both Volcanic Ash and Desert Dust in Its Iron Fertilization Potential. Glob. Biogeochem. Cycles 2021, 35. [Google Scholar] [CrossRef]
- Hood, E.; Battin, T.J.; Fellman, J.; O’Neel, S.; Spencer, R.G.M. Storage and Release of Organic Carbon from Glaciers and Ice Sheets. Nat. Geosci. 2015, 8, 91–96. [Google Scholar] [CrossRef]
- Kellerman, A.M.; Hawkings, J.R.; Wadham, J.L.; Kohler, T.J.; Stibal, M.; Grater, E.; Marshall, M.; Hatton, J.E.; Beaton, A.; Spencer, R.G.M. Glacier Outflow Dissolved Organic Matter as a Window Into Seasonally Changing Carbon Sources: Leverett Glacier, Greenland. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005161. [Google Scholar] [CrossRef]
- Feng, L.; An, Y.; Xu, J.; Kang, S. Characteristics and Sources of Dissolved Organic Matter in a Glacier in the Northern Tibetan Plateau: Differences between Different Snow Categories. Ann. Glaciol. 2018, 59, 31–40. [Google Scholar] [CrossRef]
- Säwström, C.; Mumford, P.; Marshall, W.; Hodson, A.; Laybourn-Parry, J. The Microbial Communities and Primary Productivity of Cryoconite Holes in an Arctic Glacier (Svalbard 79°N). Polar Biol. 2002, 25, 591–596. [Google Scholar] [CrossRef]
- Mindl, B.; Anesio, A.M.; Meirer, K.; Hodson, A.J.; Laybourn-Parry, J.; Sommaruga, R.; Sattler, B. Factors Influencing Bacterial Dynamics along a Transect from Supraglacial Runoff to Proglacial Lakes of a High Arctic Glacieri: Bacterial Dynamics in a Glacial Environment. FEMS Microbiol. Ecol. 2007, 59, 307–317. [Google Scholar] [CrossRef]
- Irvine-Fynn, T.D.L.; Edwards, A.; Newton, S.; Langford, H.; Rassner, S.M.; Telling, J.; Anesio, A.M.; Hodson, A.J. Microbial Cell Budgets of an Arctic Glacier Surface Quantified Using Flow Cytometry. Environ. Microbiol. 2012, 14, 2998–3012. [Google Scholar] [CrossRef] [PubMed]
- Gaidos, E.; Marteinsson, V.; Thorsteinsson, T.; Jóhannesson, T.; Rúnarsson, Á.R.; Stefansson, A.; Glazer, B.; Lanoil, B.; Skidmore, M.; Han, S.; et al. An Oligarchic Microbial Assemblage in the Anoxic Bottom Waters of a Volcanic Subglacial Lake. ISME J. 2009, 3, 486–497. [Google Scholar] [CrossRef] [PubMed]
- Christner, B.C.; Priscu, J.C.; Achberger, A.M.; Barbante, C.; Carter, S.P.; Christianson, K.; Michaud, A.B.; Mikucki, J.A.; Mitchell, A.C.; Skidmore, M.L.; et al. A Microbial Ecosystem beneath the West Antarctic Ice Sheet. Nature 2014, 512, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Mikucki, J.A.; Lee, P.A.; Ghosh, D.; Purcell, A.M.; Mitchell, A.C.; Mankoff, K.D.; Fisher, A.T.; Tulaczyk, S.; Carter, S.; Siegfried, M.R.; et al. Subglacial Lake Whillans Microbial Biogeochemistry: A Synthesis of Current Knowledge. Philos. Trans. R. Soc. A 2016, 374, 20140290. [Google Scholar] [CrossRef]
- Hood, E.; Fellman, J.; Spencer, R.G.M.; Hernes, P.J.; Edwards, R.; D’Amore, D.; Scott, D. Glaciers as a Source of Ancient and Labile Organic Matter to the Marine Environment. Nature 2009, 462, 1044–1047. [Google Scholar] [CrossRef]
- Bhatia, M.P.; Kujawinski, E.B.; Das, S.B.; Breier, C.F.; Henderson, P.B.; Charette, M.A. Greenland Meltwater as a Significant and Potentially Bioavailable Source of Iron to the Ocean. Nat. Geosci. 2013, 6, 274–278. [Google Scholar] [CrossRef]
- Hawkings, J.R.; Wadham, J.L.; Tranter, M.; Raiswell, R.; Benning, L.G.; Statham, P.J.; Tedstone, A.; Nienow, P.; Lee, K.; Telling, J. Ice Sheets as a Significant Source of Highly Reactive Nanoparticulate Iron to the Oceans. Nat. Commun. 2014, 5, 3929. [Google Scholar] [CrossRef]
- Hawkings, J.R.; Benning, L.G.; Raiswell, R.; Kaulich, B.; Araki, T.; Abyaneh, M.; Stockdale, A.; Koch-Müller, M.; Wadham, J.L.; Tranter, M. Biolabile Ferrous Iron Bearing Nanoparticles in Glacial Sediments. Earth Planet. Sci. Lett. 2018, 493, 92–101. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Statham, P.J.; Tranter, M.; Wadham, J.L. Glacial Flours as a Potential Source of Fe(II) and Fe(III) to Polar Waters. Biogeochemistry 2014, 118, 443–452. [Google Scholar] [CrossRef]
- Kim, I.; Kim, G.; Choy, E.J. The Significant Inputs of Trace Elements and Rare Earth Elements from Melting Glaciers in Antarctic Coastal Waters. Polar Res. 2015, 34, 24289. [Google Scholar] [CrossRef]
- Tockner, K.; Malard, F.; Uehlinger, U.; Ward, J.V. Nutrients and Organic Matter in a Glacial River-Floodplain System (Val Roseg, Switzerland). Limnol. Oceanogr. 2002, 47, 266–277. [Google Scholar] [CrossRef]
- Warner, K.A.; Saros, J.E.; Simon, K.S. Nitrogen Subsidies in Glacial Meltwater: Implications for High Elevation Aquatic Chains: Nitrogen Subsidies in Glacial Meltwater. Water Resour. Res. 2017, 53, 9791–9806. [Google Scholar] [CrossRef]
- Martin, J.B.; Pain, A.J.; Martin, E.E.; Rahman, S.; Ackerman, P. Comparisons of Nutrients Exported From Greenlandic Glacial and Deglaciated Watersheds. Glob. Biogeochem. Cycles 2020, 34, e2020GB006661. [Google Scholar] [CrossRef]
- Miller, J.B.; Frisbee, M.D.; Hamilton, T.L.; Murugapiran, S.K. Recharge from Glacial Meltwater Is Critical for Alpine Springs and Their Microbiomes. Environ. Res. Lett. 2021, 16, 064012. [Google Scholar] [CrossRef]
- Paulsen, M.L.; Robson, B.A. Glaciers and Land-to-Ocean Flux of Carbon. In Encyclopedia of Water; Maurice, P., Ed.; Wiley: Hoboken, NJ, USA, 2019; pp. 1–12. ISBN 978-1-119-30075-5. [Google Scholar]
- Zumsteg, A.; Luster, J.; Göransson, H.; Smittenberg, R.H.; Brunner, I.; Bernasconi, S.M.; Zeyer, J.; Frey, B. Bacterial, Archaeal and Fungal Succession in the Forefield of a Receding Glacier. Microb. Ecol. 2012, 63, 552–564. [Google Scholar] [CrossRef]
- Alfaro, F.D.; Salazar-Burrows, A.; Bañales-Seguel, C.; García, J.-L.; Manzano, M.; Marquet, P.A.; Ruz, K.; Gaxiola, A. Soil Microbial Abundance and Activity across Forefield Glacier Chronosequence in the Northern Patagonian Ice Field, Chile. Arct. Antarct. Alp. Res. 2020, 52, 553–562. [Google Scholar] [CrossRef]
- Wojcik, R.; Eichel, J.; Bradley, J.A.; Benning, L.G. How Allogenic Factors Affect Succession in Glacier Forefields. Earth-Sci. Rev. 2021, 218, 103642. [Google Scholar] [CrossRef]
- Heckmann, T.; McColl, S.; Morche, D. Retreating Ice: Research in pro-Glacial Areas Matters: Research in pro-Glacial Areas. Earth Surf. Process. Landf. 2016, 41, 271–276. [Google Scholar] [CrossRef]
- Saros, J.E.; Rose, K.C.; Clow, D.W.; Stephens, V.C.; Nurse, A.B.; Arnett, H.A.; Stone, J.R.; Williamson, C.E.; Wolfe, A.P. Melting Alpine Glaciers Enrich High-Elevation Lakes with Reactive Nitrogen. Environ. Sci. Technol. 2010, 44, 4891–4896. [Google Scholar] [CrossRef]
- Fegel, T.S.; Baron, J.S.; Fountain, A.G.; Johnson, G.F.; Hall, E.K. The Differing Biogeochemical and Microbial Signatures of Glaciers and Rock Glaciers: Defining Glacier-Type Signatures. J. Geophys. Res. Biogeosci. 2016, 121, 919–932. [Google Scholar] [CrossRef]
- Slemmons, K.E.H.; Rodgers, M.L.; Stone, J.R.; Saros, J.E. Nitrogen Subsidies in Glacial Meltwaters Have Altered Planktonic Diatom Communities in Lakes of the US Rocky Mountains for at Least a Century. Hydrobiologia 2017, 800, 129–144. [Google Scholar] [CrossRef]
- Peter, H.; Sommaruga, R. Shifts in Diversity and Function of Lake Bacterial Communities upon Glacier Retreat. ISME J. 2016, 10, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; John, S.G.; Zhang, J.; Ren, J.; Wu, Y.; Zhu, Z.; Liu, S.; Zhu, X.; Marsay, C.M.; Wenger, F. Transport and Reaction of Iron and Iron Stable Isotopes in Glacial Meltwaters on Svalbard near Kongsfjorden: From Rivers to Estuary to Ocean. Earth Planet. Sci. Lett. 2015, 424, 201–211. [Google Scholar] [CrossRef]
- Chifflard, P.; Fasching, C.; Reiss, M.; Ditzel, L.; Boodoo, K.S. Dissolved and Particulate Organic Carbon in Icelandic Proglacial Streams: A First Estimate. Water 2019, 11, 748. [Google Scholar] [CrossRef]
- Hodson, A.; Nowak, A.; Sabacka, M.; Jungblut, A.; Navarro, F.; Pearce, D.; Ávila-Jiménez, M.L.; Convey, P.; Vieira, G. Climatically Sensitive Transfer of Iron to Maritime Antarctic Ecosystems by Surface Runoff. Nat. Commun. 2017, 8, 14499. [Google Scholar] [CrossRef]
- Meire, L.; Mortensen, J.; Meire, P.; Juul-Pedersen, T.; Sejr, M.K.; Rysgaard, S.; Nygaard, R.; Huybrechts, P.; Meysman, F.J.R. Marine-Terminating Glaciers Sustain High Productivity in Greenland Fjords. Glob. Chang. Biol. 2017, 23, 5344–5357. [Google Scholar] [CrossRef]
- Torsvik, T.; Albretsen, J.; Sundfjord, A.; Kohler, J.; Sandvik, A.D.; Skarðhamar, J.; Lindbäck, K.; Everett, A. Impact of Tidewater Glacier Retreat on the Fjord System: Modeling Present and Future Circulation in Kongsfjorden, Svalbard. Estuar. Coast. Shelf Sci. 2019, 220, 152–165. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Carroll, D.; Dunse, T.; Hodson, A.; Holding, J.M.; Iriarte, J.L.; Ribeiro, S.; Achterberg, E.P.; Cantoni, C.; Carlson, D.F.; et al. Review Article: How Does Glacier Discharge Affect Marine Biogeochemistry and Primary Production in the Arctic? Cryosphere 2020, 14, 1347–1383. [Google Scholar] [CrossRef]
- Stuart-Lee, A.E.; Mortensen, J.; van der Kaaden, A.-S.; Meire, L. Seasonal Hydrography of Ameralik: A Southwest Greenland Fjord Impacted by a Land-Terminating Glacier. J. Geophys. Res. Ocean. 2021, 126, e2021JC017552. [Google Scholar] [CrossRef]
- Lyons, W.B.; Dailey, K.R.; Welch, K.A.; Deuerling, K.M.; Welch, S.A.; McKnight, D.M. Antarctic Streams as a Potential Source of Iron for the Southern Ocean. Geology 2015, 43, 1003–1006. [Google Scholar] [CrossRef]
- Bhatia, M.P.; Waterman, S.; Burgess, D.O.; Williams, P.L.; Bundy, R.M.; Mellett, T.; Roberts, M.; Bertrand, E.M. Glaciers and Nutrients in the Canadian Arctic Archipelago Marine System. Glob. Biogeochem. Cycles 2021, 35, e2021GB006976. [Google Scholar] [CrossRef]
- Williams, P.L.; Burgess, D.O.; Waterman, S.; Roberts, M.; Bertrand, E.M.; Bhatia, M.P. Nutrient and Carbon Export From a Tidewater Glacier to the Coastal Ocean in the Canadian Arctic Archipelago. JGR Biogeosci. 2021, 126, e2021JG006289. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Bacon, S.; Arendt, K.; Connelly, D.P.; Statham, P.J. Glacial Meltwater from Greenland Is Not Likely to Be an Important Source of Fe to the North Atlantic. Biogeochemistry 2015, 124, 1–11. [Google Scholar] [CrossRef]
- Cape, M.R.; Straneo, F.; Beaird, N.; Bundy, R.M.; Charette, M.A. Nutrient Release to Oceans from Buoyancy-Driven Upwelling at Greenland Tidewater Glaciers. Nat. Geosci. 2019, 12, 34–39. [Google Scholar] [CrossRef]
- Halbach, L.; Vihtakari, M.; Duarte, P.; Everett, A.; Granskog, M.A.; Hop, H.; Kauko, H.M.; Kristiansen, S.; Myhre, P.I.; Pavlov, A.K.; et al. Tidewater Glaciers and Bedrock Characteristics Control the Phytoplankton Growth Environment in a Fjord in the Arctic. Front. Mar. Sci. 2019, 6, 254. [Google Scholar] [CrossRef]
- Tremblay, J.-É.; Anderson, L.G.; Matrai, P.; Coupel, P.; Bélanger, S.; Michel, C.; Reigstad, M. Global and Regional Drivers of Nutrient Supply, Primary Production and CO2 Drawdown in the Changing Arctic Ocean. Progress. Oceanogr. 2015, 139, 171–196. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Carroll, D.; Browning, T.J.; Meire, L.; Mortensen, J.; Krisch, S.; Achterberg, E.P. Non-Linear Response of Summertime Marine Productivity to Increased Meltwater Discharge around Greenland. Nat. Commun. 2018, 9, 3256. [Google Scholar] [CrossRef]
- Lam, P.J.; Bishop, J.K.B. The Continental Margin Is a Key Source of Iron to the HNLC North Pacific Ocean: Continental Margin and the North Pacific. Geophys. Res. Lett. 2008, 35, L07608. [Google Scholar] [CrossRef]
- Laufkötter, C.; Stern, A.A.; John, J.G.; Stock, C.A.; Dunne, J.P. Glacial Iron Sources Stimulate the Southern Ocean Carbon Cycle. Geophys. Res. Lett. 2018, 45, 13377–13385. [Google Scholar] [CrossRef]
- Arrigo, K.R.; Dijken, G.L.; Castelao, R.M.; Luo, H.; Rennermalm, Å.K.; Tedesco, M.; Mote, T.L.; Oliver, H.; Yager, P.L. Melting Glaciers Stimulate Large Summer Phytoplankton Blooms in Southwest Greenland Waters. Geophys. Res. Lett. 2017, 44, 6278–6285. [Google Scholar] [CrossRef]
- Cauvy-Fraunié, S.; Dangles, O. A Global Synthesis of Biodiversity Responses to Glacier Retreat. Nat. Ecol. Evol. 2019, 3, 1675–1685. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, M.J.; Connelly, D.P.; Arendt, K.E.; Juul-Pedersen, T.; Stinchcombe, M.C.; Meire, L.; Esposito, M.; Krishna, R. Seasonal Changes in Fe along a Glaciated Greenlandic Fjord. Front. Earth Sci. 2016, 4. [Google Scholar] [CrossRef]
- Lewis, S.M.; Smith, L.C. Hydrologic Drainage of the Greenland Ice Sheet. Hydrol. Process. 2009, 23, 2004–2011. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, S.; Zhao, L.; Li, Z.; Kang, S. Global Warming Weakening the Inherent Stability of Glaciers and Permafrost. Sci. Bull. 2019, 64, 245–253. [Google Scholar] [CrossRef]
- Fransson, A.; Chierici, M.; Nomura, D.; Granskog, M.A.; Kristiansen, S.; Martma, T.; Nehrke, G. Effect of Glacial Drainage Water on the CO2 System and Ocean Acidification State in an Arctic Rctic Tidewater-glacier Fjord during Two Contrasting Years. J. Geophys. Res. Ocean. 2015, 120, 2413–2429. [Google Scholar] [CrossRef]
- Murray, C.; Markager, S.; Stedmon, C.A.; Juul-Pedersen, T.; Sejr, M.K.; Bruhn, A. The Influence of Glacial Melt Water on Bio-Optical Properties in Two Contrasting Greenlandic Fjords. Estuar. Coast. Shelf Sci. 2015, 163, 72–83. [Google Scholar] [CrossRef]
- Buongiorno, J.; Herbert, L.C.; Wehrmann, L.M.; Michaud, A.B.; Laufer, K.; Røy, H.; Jørgensen, B.B.; Szynkiewicz, A.; Faiia, A.; Yeager, K.M.; et al. Complex Microbial Communities Drive Iron and Sulfur Cycling in Arctic Fjord Sediments. Appl. Environ. Microbiol. 2019, 85, e00949-19. [Google Scholar] [CrossRef]
- Cantoni, C.; Hopwood, M.J.; Clarke, J.S.; Chiggiato, J.; Achterberg, E.P.; Cozzi, S. Glacial Drivers of Marine Biogeochemistry Indicate a Future Shift to More Corrosive Conditions in an Arctic Fjord. J. Geophys. Res. Biogeosci. 2020, 125. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Laufer, K.; Michaud, A.B.; Wehrmann, L.M. Biogeochemistry and Microbiology of High Arctic Marine Sediment Ecosystems—Case Study of Svalbard Fjords. Limnol. Oceanogr. 2021, 66, S273–S292. [Google Scholar] [CrossRef]
- Szeligowska, M.; Trudnowska, E.; Boehnke, R.; Dąbrowska, A.M.; Dragańska-Deja, K.; Deja, K.; Darecki, M.; Błachowiak-Samołyk, K. The Interplay between Plankton and Particles in the Isfjorden Waters Influenced by Marine- and Land-Terminating Glaciers. Sci. Total. Environ. 2021, 780, 146491. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Location | Water Presence in Accumulation Season | Residence Time and Water Flow | Important References |
|---|---|---|---|
| Ice-lidded cryoconite holes | Yes, but some can completely freeze during accumulation season | Years, but occasionally connected to the rest of the system during accumulation season. Estimates say that ~50% of the cryoconite holes hydrologically connect to the supraglacial system every accumulation season. | [61,91] |
| Open cryoconite holes | No | During accumulation season, they have a higher connectivity to the glacial system than ice-lidded cryoconite holes, showing lower permanence times. | [39,63] |
| Weathering crust | Yes, if insulated by a snowpack. | Several days, water is released to the glacial when the system is saturated. Water flow is slow in the interstitial space. | [55,58,60] |
| Supraglacial streams | No | Depending on the ablation stage of the system, water can flow at different rates. | [64] |
| Supraglacial ponds and lakes | Yes. Lakes can form ice lids and maintain water during accumulation season. | Years; water is released to the glacial when the system is saturated. Smaller ponds can be drained by meltwater streams. | [39,50,61,64] |
| Firn aquifer | Yes | Perennial; water is released to the glacial when the system is saturated. | [52,92,93,94,95] |
| Englacial conduits | Yes, when the presence of solutes and particles lowers water freezing temperature. | Fast-flowing systems with permanence of hours up to a day. However, water can be present all year round and even for multiple years if water pockets are formed by collapsed conduits. Depending on the ablation stage of the system, water can flow at different rates. | [88,96,97] |
| Ice veins | Yes | Residence times in the ice veins are unknown. However, due to the low water flowing rate, we assume it to be in the order of years (at the very least). | [98] |
| Subglacial cavities | Yes | Days to months, and water is usually present all year round. | [99,100,101] |
| Subglacial channels | No | Hours, water is usually present only during peak ablation season. | [99,100,101] |
| Subglacial till | Yes | Potentially all-year round. | [36,78] |
| Subglacial lakes | Yes | Years; some systems have estimated water residence times of millions of years. | [79,80,102] |
| Cell Concentration | Source | Sampling Ablation Season | Reference |
|---|---|---|---|
| 1.0–4.5 × 104 cells mL−1 (3.97–12.7 × 104 VLP mL−1) | Cryoconite holes, Midtre Lovénbreen | 2000 and 2001 | [186] |
| 1.38 × 104–4.84 × 104 cells mL−1 | Supraglacial meltwater runoff and cryoconite holes, Midtre Lovénbreen | 2004 | [187] |
| 5.4 ± 1.6 × 104 cells mL−1 | Cryoconite holes, Austre Broggerbreen | 2005 | [31] |
| 3.4 ± 1.2 × 104 cells mL−1 | Cryoconite holes, Midtre Lovénbreen | 2005 | [31] |
| 4.1 ± 3.8 × 104 cells mL−1 | Cryoconite holes, Rotmoosferner | 2004 | [31] |
| 3.7 ± 1.4 × 104 cells mL−1 | Cryoconite holes, Stubacher Sonnblickkees | 2007 | [31] |
| 1.3 ± 8.2 × 104 cells mL−1 | Cryoconite holes, blue ice close to Patriot Hills | 2002 | [31] |
| 4.4 ± 2.4 × 104 cells mL−1 | Cryoconite holes, Canada, Commonwealth, and Taylor glaciers | 2005 | [31] |
| 2 × 104 cells mL−1 | Supraglacial meltwater runoff, Midtre Lovénbreen | 2010 | [188] |
| 8.38 × 103 ± 9.85 × 103 cells mL−1 | Supraglacial meltwater runoff, Russell glacier | 2012 | [6] |
| 2.2 × 104 ± 5.5 × 104 cells mL−1 | Weathering crust, Northern Hemisphere glaciers | 2014, 2015 and 2016 | [29] |
| 6 × 104 cells ml−1 | Subglacial brine, Blood Falls | 2004 | [174] |
| 106–108 cells mL−1 | Ice vein water | / | [154] |
| 4.7–5.7 × 105 cells mL−1 | Subglacial water, Skaftá subglacial lake | 2006 | [189] |
| 4.4 ± 2.2 × 105 cells mL−1 | Subglacial water, East Skaftárkatlar subglacial lakes | 2007 | [162] |
| 1.3 × 105 cells mL−1 | Subglacial water, subglacial lake Whillans | 2013 | [190] |
| 1.3 × 105 cells mL−1 | Subglacial water, subglacial lake Whillans | 2013 | [191] |
| 1.15 × 105 ± 1.38 × 105 cells mL−1 | Subglacial meltwater runoff, Leverett glacier | 2012 | [6] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varliero, G.; Lebre, P.H.; Frey, B.; Fountain, A.G.; Anesio, A.M.; Cowan, D.A. Glacial Water: A Dynamic Microbial Medium. Microorganisms 2023, 11, 1153. https://doi.org/10.3390/microorganisms11051153
Varliero G, Lebre PH, Frey B, Fountain AG, Anesio AM, Cowan DA. Glacial Water: A Dynamic Microbial Medium. Microorganisms. 2023; 11(5):1153. https://doi.org/10.3390/microorganisms11051153
Chicago/Turabian StyleVarliero, Gilda, Pedro H. Lebre, Beat Frey, Andrew G. Fountain, Alexandre M. Anesio, and Don A. Cowan. 2023. "Glacial Water: A Dynamic Microbial Medium" Microorganisms 11, no. 5: 1153. https://doi.org/10.3390/microorganisms11051153
APA StyleVarliero, G., Lebre, P. H., Frey, B., Fountain, A. G., Anesio, A. M., & Cowan, D. A. (2023). Glacial Water: A Dynamic Microbial Medium. Microorganisms, 11(5), 1153. https://doi.org/10.3390/microorganisms11051153