
Glutamate Transporters GltS, GltP and GltI Are Involved in Escherichia coli Tolerance In Vitro and Pathogenicity in Mouse Urinary Tract Infections

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cell Line
2.2. RNA Isolation, cDNA Construction, and Real-Time PCR Assay
2.3. Construction of GltS, GltP, and GltI Deletion Mutants in E. coli BW25113 and Uropathogenic E. coli UTI89
2.4. Complementation of E. coli Deletion Mutant Strains
2.5. Evaluation of Tolerance of the Constructed Mutants to Antibiotics and Various Stressors
2.6. Assays to Detect Bacterial Abilities to Adhere to and Invade Epithelial Cells
2.7. Animals and Urinary Tract Infection Model in Mice
2.8. Statistical Analysis
3. Results
3.1. E. coli GltS, GltP, and GltI were Highly Expressed in the Stationary Phase
3.2. Supplementation of Glutamate in Culture Medium Increased the Ability of E. coli Tolerance
3.3. Deletion of GltS, GltP, and GltI Genes in E. coli BW25113 Results in Decreased Tolerance to Antibiotics and Stress Conditions
3.4. Uropathogenic E. coli UTI89 Mutants ΔgltS, ΔgltP, and ΔgltI Exhibited Weakened Adhesion and Invasion to Human Bladder Epithelial Cells
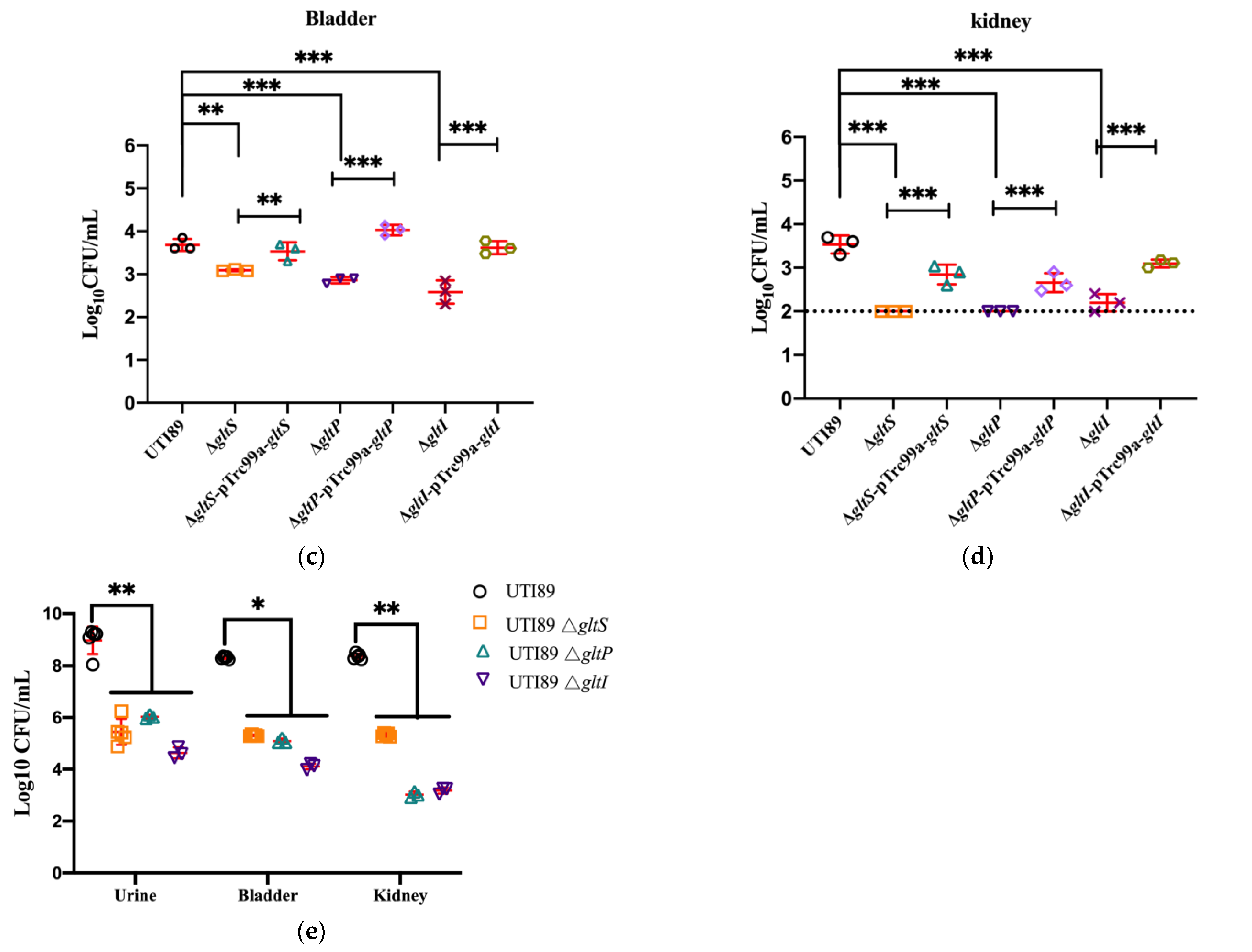
3.5. Loss of GltS, GltP, and GltI in Uropathogenic E. coli UTI89 Caused Markedly Reduced Survival in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blango, M.G.; Mulvey, M.A. Persistence of uropathogenic Escherichia coli in the face of multiple antibiotics. Antimicrob. Agents Chemother. 2010, 54, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yew, W.W.; Barer, M.R. Targeting Persisters for Tuberculosis Control. Antimicrob. Agents Chemother. 2012, 56, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. APMIS 2013, 121, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Gerdes, K.; Lewis, K.; McKinney, J.D. A problem of persistence: Still more questions than answers? Nat. Rev. Microbiol. 2013, 11, 587–591. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Persisters, persistent infections and the Yin–Yang model. Emerg. Microbes Infect. 2014, 3, e3. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Hayes, F.; Melderen, L.V. Toxins-antitoxins: Diversity, evolution and function. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 386–408. [Google Scholar] [CrossRef]
- Hauryliuk, V.; Atkinson, G.C.; Murakami, K.S.; Tenson, T.; Gerdes, K. Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 2015, 13, 298–309. [Google Scholar] [CrossRef]
- Dörr, T.; Lewis, K.; Vulić, M.; Rosenberg, S.M. SOS Response Induces Persistence to Fluoroquinolones in Escherichia coli. PLoS Genet. 2009, 5, e1000760. [Google Scholar] [CrossRef]
- Völzing, K.G.; Brynildsen, M.P. Stationary-Phase Persisters to Ofloxacin Sustain DNA Damage and Require Repair Systems Only during Recovery. mBio 2015, 6, e00731-15. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Sim, S.; Shi, W.; Du, L.; Xing, D.; Zhang, Y. Energy production genes sucB and ubiF are involved in persister survival and tolerance to multiple antibiotics and stresses in Escherichia coli. FEMS Microbiol. Lett. 2010, 303, 33–40. [Google Scholar] [CrossRef]
- Li, J.; Ji, L.; Shi, W.; Xie, J.; Zhang, Y. Trans-translation mediates tolerance to multiple antibiotics and stresses in Escherichia coli. J. Antimicrob. Chemother. 2013, 68, 2477–2481. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Zhao, Z.; Li, Y.; Jin, Z.; Qi, M.; Zhao, Y.; Ke, Y.; Yun, Z.; Chen, H.; Baker, M.B. Enhanced Efflux Activity Facilitates Drug Tolerance in Dormant Bacterial Cells. Mol. Cell 2016, 62, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Orman, M.A.; Brynildsen, M.P. Establishment of a method to rapidly assay bacterial persister metabolism. Antimicrob. Agents Chemother. 2013, 57, 4398–4409. [Google Scholar] [CrossRef]
- Conover, M.S.; Hadjifrangiskou, M.; Palermo, J.J.; Hibbing, M.E.; Dodson, K.W.; Hultgren, S.J. Metabolic Requirements of Escherichia coli in Intracellular Bacterial Communities during Urinary Tract Infection Pathogenesis. mBio 2016, 7, e00104. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y. PhoU is a persistence switch involved in persister formation and tolerance to multiple antibiotics and stresses in Escherichia coli. Antimicrob. Agents Chemother. 2007, 51, 2092–2099. [Google Scholar] [CrossRef]
- Yee, R.; Cui, P.; Shi, W.; Feng, J.; Zhang, Y. Genetic Screen Reveals the Role of Purine Metabolism in Staphylococcus aureus Persistence to Rifampicin. Antibiotics 2015, 4, 627–642. [Google Scholar] [CrossRef]
- Feehily, C.; Karatzas, K.A. Role of glutamate metabolism in bacterial responses towards acid and other stresses. J. Appl. Microbiol. 2013, 114, 11–24. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K.; Keembiyehetty, C.N.; Chen, W.; Tabor, H. Polyamines Stimulate the Level of the σ38 Subunit (RpoS) of Escherichia coli RNA Polymerase, Resulting in the Induction of the Glutamate Decarboxylase-dependent Acid Response System via the gadE Regulon. J. Biol. Chem. 2015, 290, 17809–17821. [Google Scholar] [CrossRef]
- Huang, Y.; Suo, Y.; Shi, C.; Szlavik, J.; Shi, X.M.; KnøChel, S. Mutations in gltB and gltC reduce oxidative stress tolerance and biofilm formation in Listeria monocytogenes 4b G. Int. J. Food Microbiol. 2013, 163, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Bearson, B.L.; Lee, I.S.; Casey, T.A. Escherichia coli O157:H7 glutamate- and arginine-dependent acid-resistance systems protect against oxidative stress during extreme acid challenge. Microbiology 2009, 155, 805. [Google Scholar] [CrossRef] [PubMed]
- Yee, R.; Cui, P.; Shi, W.; Feng, J.; Wang, J.; Zhang, Y. Identification of a novel gene argJ involved in arginine biosynthesis critical for persister formation in Staphylococcus aureus. Discov. Med. 2020, 29, 65–77. [Google Scholar]
- Biase, D.; Pennacchietti, E. Glutamate decarboxylase-dependent acid resistance in orally acquired bacteria: Function, distribution and biomedical implications of the gadBC operon. Mol. Microbiol. 2012, 86, 770–786. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Cui, P.; Shi, W.; Zhang, S.; Feng, J.; Wang, Y.; Sullivan, D.; Zhang, W.; Zhu, B.; Zhang, Y. Identification of Anti-Persister Activity against Uropathogenic Escherichia coli from a Clinical Drug Library. Antibiotics 2015, 4, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, J.; Cao, Q.; Li, F.; Niu, H. Identification of Novel Genes Involved in Escherichia coli Persistence to Tosufloxacin. Front. Cell. Infect. Microbiol. 2020, 10, 581986. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Martinez, J.J.; Mulvey, M.A.; Schilling, J.D.; Pinkner, J.S.; Hultgren, S.J. Type 1 pilus-mediated bacterial invasion of bladder epithelial cells. EMBO J. 2014, 19, 2803–2812. [Google Scholar] [CrossRef]
- Elsinghorst, E.A. Measurement of invasion by gentamicin resistance. Methods Enzymol. 1994, 236, 405–420. [Google Scholar]
- Cui, P.; Niu, H.; Shi, W.; Zhang, S.; Zhang, H.; Margolick, J.; Zhang, W.; Zhang, Y.; Cui, C. Disruption of membrane by colistin kills uropathogenic Escherichia coli persisters and enhances killing of other antibiotics. Antimicrob. Agents Chemother. 2018, 60, 6867–6871. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, 2437. [Google Scholar]
- Yee, R.; Yuan, Y.; Shi, W.; Brayton, C.; Tarff, A. Infection with persister forms of Staphylococcus aureus causes a persistent skin infection with more severe lesions in mice: Failure to clear the infection by the current standard of care treatment. Discov. Med. 2019, 28, 7–16. [Google Scholar]
- Richard, H.; Foster, J.W. Sodium regulates Escherichia coli acid resistance, and influences GadX- and GadW-dependent activation of gadE. Microbiology 2007, 153, 3154–3161. [Google Scholar] [CrossRef] [PubMed]
- Goode, O.; Smith, A.; Zarkan, A.; Cama, J.; Invergo, B.M.; Belgami, D.; Cano-Muniz, S.; Metz, J.; O’Neill, P.; Jeffries, A.; et al. Persister Escherichia coli Cells Have a Lower Intracellular pH than Susceptible Cells but Maintain Their pH in Response to Antibiotic Treatment. mBio 2021, 12, e0090921. [Google Scholar] [CrossRef] [PubMed]
- Foster, J. Escherichia coli acid resistance: Tales of an amateur acidophile. Nat. Rev. Microbiol. 2004, 2, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Biase, D.D.; Tramonti, A.; Bossa, F.; Visca, P. The response to stationary-phase stress conditions in Escherichia coli: Role and regulation of the glutamic acid decarboxylase system. Mol. Microbiol. 1999, 32, 1198–1211. [Google Scholar] [CrossRef]
- Welsh, D.T. Ecological significance of compatible solute accumulation by micro-organisms: From single cells to global climate. Fems Microbiol. Rev. 2000, 24, 263–290. [Google Scholar] [CrossRef]
- Yee, R.; Feng, J.; Wang, J.; Chen, J.; Zhang, Y. Identification of Genes Regulating Cell Death in Staphylococcus aureus. Front. Microbiol. 2019, 10, 2199. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, M.; Zhang, Y.; Zhang, Y.; Bhushan, S. Uropathogenic Escherichia coli virulence factor α hemolysin reduces histone acetylation to inhibit expression of pro-inflammatory cytokine genes. J. Infect. Dis. 2021, 223, 1040–1051. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | |
|---|---|---|
| For RT-PCR | gltS A-F | GTTCATTGAACGTTATGGCTTC |
| gltS A-R | CCGCCAATCAAGCCGCCCAG | |
| gltP A-F | GCAGTTCCCACGGCATTATG | |
| gltP A-R | CAGAGATGGAGCGGAACACG | |
| gltI A-F | CGATTTTGAATGTGGTTCTAC | |
| gltI A-R | GACTACGGCTTTGTCTTTCAG | |
| rrsA-F | GAAAGGGGAGTGGGGTAAAGG | |
| rrsA-R | CGGCTGAAGGTGATGGTGT | |
| For deletion mutant construction in coli BW25113 | gltS B-F | GATGAAGCGGCGGTAGAAGTGCCGCCGCAACAAAGACAAATGCCTGATGTGTAGGCTGGAGCTGCTTC |
| gltS B-R | ATCAGGCATTTGTCTTTGTTGCGGCGGCACTTCTACCGCCGCTTCATCGGTATGGGAATTAGCCATGGTCC | |
| gltP B-F | TTCTCGCGTTTCTGAACGGGGAACGGCGCTCCATTGAGGAAGTTATTCTGGTGTAGGCTGGAGCTGCTTC | |
| gltP B-R | AGTCAGGCATCCACACATTGCCGGGTGGATATCCCCCGGCAATCTTCAAATGGGAATTAGCCATGGTCC | |
| gltI B-F | TCACAACGGGTATCCATGCGTTCTTAACGCAGAAGATAAAGGAGTTGGATGTGTAGGCTGGAGCTGCTTC | |
| gltI B-R | TGCTACGTAACAATCGAGAGGGCTGGAATTTCCGCCCCTGGTTCTTGTAAATGGGAATTAGCCATGGTCC | |
| For deletion mutant construction in E. coli UTI89 | gltS C-F | GTTACTCGAATGCGTAAAAAGCGGCGGTGAGAAGACCGCCGCTTCATCGGGTGTAGGCTGGAGCTGCTTC |
| gltS C-R | GATGAAGTATGACGAGTATGAAAGAGTGATGCGGACACAAAGGAGTAACTATGGGAATTAGCCATGGTCC | |
| gltP C-F | TTCTCGCGTTTCTGAACGGGGAACGGCGCTCCATTGAGGAAGTTATTCTGGTGTAGGCTGGAGCTGCTTC | |
| gltP C-R | AGTCAGGCATCCACACATTGCCGGGTGGATATCCCCCGGCAATCTTCAAATGGGAATTAGCCATGGTCC | |
| gltI C-F | TGCTACGTAACAATCGAGAGGGCTGGAATTTCCGCCCCTGGTTCTTGTAAGTGTAGGCTGGAGCTGCTTC | |
| gltI C-R | TCACAACGGGTATCCATGCGTTTTTTAACGCAGAAGATAAAGGAGTTGGATATGGGAATTAGCCATGGTCC | |
| For complementation of deletion mutants | gltS D-F | CGGAATTCATGTCGATACTTTAGCAACGCTTGT (Ecor I) |
| gltS D-R | TCGAAGCTTTTACCAGCGCATTGACGATA (BamH I) | |
| gltP D-F | CCGGAATTCATGCGTAACGAACTGAACGG (EcoR I) | |
| gltP D-R | CGCGGATCCTTATTTTCAATCAACTGGATCAGG (BamH I) | |
| gltI D-F | GGAATTCATGCAATTACGTAAACCTGC (EcoR I) | |
| gltI D-R | CGGGATCCTTAGTTCAGTGCCTTGTC (BamH I) | |
| Strains | Levofloxacin (μM) | Gentamicin (μM) | Tosufloxacin (μM) |
|---|---|---|---|
| BW25113 | 0.03 | 2.75 | 0.03 |
| ΔgltS | 0.03 | 2.75 | 0.015 |
| ΔgltP | 0.015 | 2.75 | 0.0075 |
| ΔgltI | 0.015 | 2.75 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, H.; Li, T.; Du, Y.; Lv, Z.; Cao, Q.; Zhang, Y. Glutamate Transporters GltS, GltP and GltI Are Involved in Escherichia coli Tolerance In Vitro and Pathogenicity in Mouse Urinary Tract Infections. Microorganisms 2023, 11, 1173. https://doi.org/10.3390/microorganisms11051173
Niu H, Li T, Du Y, Lv Z, Cao Q, Zhang Y. Glutamate Transporters GltS, GltP and GltI Are Involved in Escherichia coli Tolerance In Vitro and Pathogenicity in Mouse Urinary Tract Infections. Microorganisms. 2023; 11(5):1173. https://doi.org/10.3390/microorganisms11051173
Chicago/Turabian StyleNiu, Hongxia, Tuodi Li, Yunjie Du, Zhuoxuan Lv, Qianqian Cao, and Ying Zhang. 2023. "Glutamate Transporters GltS, GltP and GltI Are Involved in Escherichia coli Tolerance In Vitro and Pathogenicity in Mouse Urinary Tract Infections" Microorganisms 11, no. 5: 1173. https://doi.org/10.3390/microorganisms11051173
APA StyleNiu, H., Li, T., Du, Y., Lv, Z., Cao, Q., & Zhang, Y. (2023). Glutamate Transporters GltS, GltP and GltI Are Involved in Escherichia coli Tolerance In Vitro and Pathogenicity in Mouse Urinary Tract Infections. Microorganisms, 11(5), 1173. https://doi.org/10.3390/microorganisms11051173