
Microbiota and Cyanotoxin Content of Retail Spirulina Supplements and Spirulina Supplemented Foods

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Foods and Spirulina Dietary Supplements Examined
2.2. Measurement of pH
2.3. Microbiological Analysis and Isolation of Bacteria
2.4. Bacterial Identification Using Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry
2.5. Detection of Cyanotoxins Using Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Amplicon Sequencing Analysis
3. Results
3.1. pH Measurements
3.2. Microbiological Analysis Using Culture-Dependent Techniques
3.3. Microbiological Analysis by Amplicon Sequencing
3.4. Identification of Individual Isolates by MALDI-TOF
3.5. Cyanotoxins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lafarga, T.; Fernández-Sevilla, J.M.; González-López, C.; Acién-Fernández, F.G. Spirulina for the food and functional food industries. Food Res. Int. 2020, 137, 109356. [Google Scholar] [CrossRef] [PubMed]
- Muys, M.; Sui, Y.; Schwaiger, B.; Lesueur, C.; Vandenheuvel, D.; Vermeir, P.; Vlaeminck, S.E. High variability in nutritional value and safety of commercially available Chlorella and Spirulina biomass indicates the need for smart production strategies. Bioresour. Tech. 2019, 275, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Wu, Q.; Kuča, K. Spirulina. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 569–583. [Google Scholar] [CrossRef]
- Vonshak, A.; Tomaselli, L. Arthrospira (Spirulina): Systematics and EcophysioIogy. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 505–522. [Google Scholar] [CrossRef]
- Masojídek, J.; Torzillo, G. Mass Cultivation of Freshwater Microalgae. In Reference Module in Earth Systems and Environmental Sciences [Online Collection]; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Microalgae in medicine and human health: A historical perspective. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 195–210. [Google Scholar] [CrossRef]
- Lee, E.H.; Park, J.-E.; Choi, Y.-J.; Huh, K.-B.; Kim, W.-Y. A randomised study to establish the effects of spirulina in type 2 diabetes mellitus patients. Nutr. Res. Pract. 2008, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Cingi, C.; Conk-Dalay, M.; Cakli, H.; Bal, C. The effects of spirulina on allergic rhinitis. Eur. Arch. Otorhinolaryngol. 2008, 265, 1219–1223. [Google Scholar] [CrossRef]
- Mao, T.K.; Van de Water, J.; Gershwin, M.E. Effects of a Spirulina-based dietary supplement on cytokine production from allergic rhinitis patients. J. Med. Food 2005, 8, 27–30. [Google Scholar] [CrossRef]
- Hoekstra, D.T.; Volschenk, H.; Collins, M.; McMaster, L.D. An investigation of Clostridium species present in nutraceutical preparations of Arthrospira platensis (Spirulina) for human consumption. J. Appl. Phycol. 2011, 23, 777–787. [Google Scholar] [CrossRef]
- Vardaka, E.; Kormas, K.A.; Katsiapi, M.; Genitsaris, S.; Moustaka-Gouni, M. Molecular diversity of bacteria in commercially available “Spirulina” food supplements. PeerJ 2016, 1610, e1610. [Google Scholar] [CrossRef]
- Anon. Bacteriological Analytical Manual (BAM). US Food and Drug Administration. Available online: https://www.fda.gov/food/laboratory-methods-food/bacteriological-analytical-manual-bam (accessed on 23 March 2023).
- Fastner, J.; Flieger, I.; Neumann, U. Optimized extraction of microcystins from field samples—A comparison of different solvents and procedures. Water Res. 1998, 32, 3177–3181. [Google Scholar] [CrossRef]
- Törökné, A.; Asztalos, M.; Bánkiné, M.; Bickel, H.; Borbély, G.; Carmeli, S.; Codd, G.A.; Fastner, J.; Huang, Q.; Humpage, A.; et al. Interlaboratory comparison trial on cylindrospermopsin measurement. Anal. Biochem. 2004, 15, 280–284. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Bell, S.G.; Codd, G.A. Colorimetric immuno-protein phosphatase inhibition assay for specific detection of microcystins and nodularins of cyanobacteria. Appl. Environ. Microbiol. 2001, 67, 904–909. [Google Scholar] [CrossRef]
- Dietrich, D.; Hoeger, S. Guidance values for microcystins in water and cyanobacterial supplement products (blue-green algal supplements): A reasonable or misguided approach? Toxicol. Appl. Pharmacol. 2005, 15, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Humpage, A.R.; Falconer, I.R. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: Determination of no observed adverse effect level for deriving a drinking water guideline value. Environ. Toxicol. 2003, 18, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acid Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef]
- Fromme, H.; Köhler, A.; Krause, R.; Führling, D. Occurrence of cyanobacterial toxins—Microcystins and anatoxin-a—In Berlin water bodies with implications to human health and regulations. Environ. Toxicol. 2000, 15, 120–130. [Google Scholar] [CrossRef]
- Lechner, S.; Mayr, R.; Francis, K.P.; Prüss, B.M.; Kaplan, T.; Wiessner-Gunkel, E.; Stewart, G.S.; Scherer, S. Bacillus weihenstephanensis sp. nov. is a new psychrotolerant species of the Bacillus cereus group. Int. J. Syst. Bacteriol. 1998, 48, 1373–1382. [Google Scholar] [CrossRef]
- Ha, M.; Jo, H.-J.; Choi, E.-K.; Kim, Y.; Kim, J.; Cho, H.-J. Reliable identification of Bacillus cereus group species using low mass biomarkers by MALDI-TOF MS. J. Microbiol. Biothechnol. 2019, 29, 887–896. [Google Scholar] [CrossRef]
- Manzulli, V.; Rondinone, V.; Buchicchio, A.; Serrecchia, L.; Cipolletta, D.; Fasanella, A.; Parisi, A.; Difato, L.; Iatarola, M.; Aceti, A.; et al. Discrimination of Bacillus cereus group members by MALDI-TOF mass spectrometry. Microorganisms 2021, 9, 1202. [Google Scholar] [CrossRef] [PubMed]
- Pauker, V.I.; Thoma, B.R.; Grass, G.; Bleichert, P.; Hanczaruk, M.; Zöller, L.; Zange, S. Improved discrimination of Bacillus anthracis from closely related species in the Bacillus cereus sensu lato group based on matrix-assisted laser desorption ionization—Time of flight mass spectrometry. J. Clin. Microbiol. 2018, 56, e01900-17. [Google Scholar] [CrossRef] [PubMed]
- Stenfors, L.P.; Mayr, R.; Scherer, S.; Granum, P.E. Pathogenic potential of fifty Bacillus weihenstephanensis strains. FEMS Microbiol. Lett. 2002, 215, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Thorsen, L.; Hansen, B.M.; Nielsen, K.F.; Hendriksen, N.B.; Phipps, R.K.; Budde, B.B. Characterization of emetic Bacillus weihenstephanensis, a new cerelulide-producing bacterium. Appl. Environ. Microbiol. 2006, 72, 5118–5121. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef]
- Pomastowski, P.; Złoch, M.; Rodzik, A.; Ligor, M.; Kostrzewa, M.; Buszewski, B. Analysis of bacteria associated with honeys of different geographical and botanical origin using two different identification approaches: MALDI-TOF MS and 16S rDNA PCR technique. PLoS ONE 2019, 14, e0217078. [Google Scholar] [CrossRef]
- Daczkowska-Kozon, E.G.; Bednarczyk, A.; Biba, M.; Repich, K. Bacteria of Bacillus cereus group in cereals at retail. Pol. J. Food Nutr. Sci. 2009, 59, 53–59. [Google Scholar]
- Eijlander, R.T.; Breitenwieser, F.; de Groot, R.; Hoornstra, E.; Kamphuis, H.; Kokken, M.; Kuijpers, A.; de Mello, I.I.G.; de Rijdt, G.V.; Vadier, C.; et al. Enumeration and identification of bacterial spores in cocoa powders. J. Food Prot. 2020, 83, 1530–1539, Erratum in J. Food Prot. 2020, 83, 2030. [Google Scholar] [CrossRef]
- Hartantyo, S.H.P.; Chau, M.L.; Hoh, T.H.; Yap, M.; Yi, T.; Cao, D.Y.H.; Gutiérrez, R.A.; Ng, L.C. Foodborne Klebsiella pneumoniae: Virulence potential, antibiotic resistance, and risks to food safety. J. Food Prot. 2020, 83, 1096–1103. [Google Scholar] [CrossRef]
- Theocharidi, N.A.; Balta, I.; Houhoula, D.; Tsantes, A.G.; Lalliotis, G.P.; Polydera, A.C.; Stamatis, H.; Halvatsiotis, P. High prevalence of Klebsiella pneumoniae in Greek meat products: Detection of virulence and antimicrobial resistance genes by molecular techniques. Foods 2022, 11, 708. [Google Scholar] [CrossRef]
- Fung, C.P.; Lin, Y.T.; Lin, J.C.; Chen, T.L.; Yey, K.M.; Chang, F.Y.; Chuang, H.C.; Wu, H.S.; Tseng, C.P.; Siu, L.K. Klebsiella pneumoniae in gastrointestinal tract and pyogenic liver disease. Emerg. Infect. Dis. 2012, 18, 1322–1325. [Google Scholar] [CrossRef] [PubMed]
- Sabota, J.M.; Hoppes, W.L.; Ziegler, J.R.; DuPont, H.; Mathewson, J.; Rutecki, G.W. A new variant of food poisoning: Enteroinvasive Klebsiella pneumoniae and Escherichia coli sepsis from a contaminated hamburger. Am. J. Gastroenterol. 1998, 93, 118–119. [Google Scholar] [CrossRef] [PubMed]
- Calbo, E.; Freixas, N.; Xercavins, M.; Riera, M.; Nicolás, C.; Monistrol, O.; Solé Mdel, M.; Sala, M.R.; Vila, J.; Garau, J. Foodborne nosocomial outbreak of SHV1 and CTX-M-15-producing Klebsiella pneumoniae: Epidemiology and control. Clin. Infect. Dis. 2011, 52, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Zayet, S.; Lang, S.; Garnier, P.; Pierron, A.; Plantin, J.; Toko, L.; Royer, P.-Y.; Villemain, M.; Klopfenstein, T.; Gendrin, V. Leclercia adecarboxylata as emerging pathogen in human infections: Clinical features and antimicrobial susceptibility testing. Pathogens 2021, 10, 1399. [Google Scholar] [CrossRef]
- Al Shuhoumi, M.A.; Al Mhrooqi, A.; Al Rashdi, A.; Kumar, R.; Al Jabri, A.; Al Kalbani, A.; Al Jardani, A. First clinical case of VIM-1-producing Leclercia adecarboxylata: A case report and literature review. Med. Microecol. 2023, 15, 100075. [Google Scholar] [CrossRef]
- Kamboj, K.; Vasquez, A.; Balada-Lioasat, J.-M. Identification and significance of Weisella species infections. Front. Microbiol. 2015, 6, 1204. [Google Scholar] [CrossRef]
- Spiegelhauer, M.R.; Yusibova, M.; Rasmussen, I.K.B.; Fuglsang, K.A.; Thomsen, K.; Andersen, L.P. A case report of polymicrobial bacteremia with Weissella confusa and a comparison of previous treatment for successful recovery with a review of the literature. Access Microbiol. 2020, 2, acmi000119. [Google Scholar] [CrossRef]
- García-López, R.; Cornejo-Granados, F.; Lopez-Zavala, A.A.; Sánchez-López, F.; Cota-Huízar, A.; Sotelo-Mundo, R.R.; Guerrero, A.; Mendoza-Vargas, A.; Gómez-Gil, B.; Ochoa-Leyva, A. Doing more with less: A comparison of 16S hypervariable regions in search of defining the shrimp microbiota. Microorganisms 2020, 8, 134. [Google Scholar] [CrossRef]
- Sirichoat, A.; Sankuntaw, N.; Engchanil, C.; Buppasiri, P.; Faksri, K.; Namwat, W.; Chantratita, W.; Lulitanond, V. Comparison of different hypervariable regions of 16S rRNA for taxonomic profiling of vaginal microbiota using next-generation sequencing. Arch. Microbiol. 2021, 203, 1159–1166. [Google Scholar] [CrossRef]
- Saker, M.L.; Jungblut, A.D.; Neilan, B.A.; Rawn, D.F.; Vasconcelos, V.M. Detection of microcystin synthetase genes in health food supplements containing the freshwater cyanobacterium Aphanizomenon flos-aquae. Toxicon 2005, 46, 555–562. [Google Scholar] [CrossRef]
- Marsan, D.W.; Conrad, S.M.; Stutts, W.L.; Parker, C.H.; Deeds, J.R. Evaluation of microcystin contamination in blue-green algal dietary supplements using a protein phosphatase inhibition-based test kit. Heliyon 2018, 16, e00573. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.R.; Xiong, A.; Deeds, J.R.; Stutts, W.L.; Samdal, I.A.; Løvberg, K.E.; Miles, C.O. Microcystin toxins at potentially hazardous levels in algal dietary supplements revealed by a combination of bioassay, immunoassay, and mass spectrometric methods. J. Agric. Food Chem. 2020, 68, 8016–8025. [Google Scholar] [CrossRef]
- Del Olmo Iruela, M.; Del Mar Aparicio-Muriana, M.; Lara, F.J.; Garcia-Campaña, A.M. Determination of multiclass cyanotoxins in spirulina-based dietary supplements using a SLE-Tandem-SPE procedure followed by HILIC-MS/MS. Biol. Life Sci. Forum 2022, 14, 42. [Google Scholar] [CrossRef]
- Van Hassel, W.H.R.; Ahn, A.C.; Huybrechts, B.; Masquelier, J.; Wilmotte, A.; Andjelkovic, M. LC-MS/MS validation and quantification of cyanotoxins in algal food supplements from the Belgium market and their molecular origins. Toxins 2022, 14, 513. [Google Scholar] [CrossRef]
- Okamoto, K.; Fleming, L.E.; Wexler, P. Algae. In Encyclopedia of Toxicology, 2nd ed.; Oxford University Press: Oxford, UK, 2005; Volume 1, pp. 68–76. [Google Scholar]
- Papadimitriou, T.; Kormas, K.; Vardaka, E. Cyanotoxin contamination in commercial Spirulina food supplements. J. Consum. Prot. Food Saf. 2021, 16, 227–235. [Google Scholar] [CrossRef]
- Singh, S.; Kate, B.N.; Banerjee, U.C. Bioactive compounds from cyanobacteria and microalgae: An overview. Crit. Rev. Biotechnol. 2005, 25, 73–95. [Google Scholar] [CrossRef] [PubMed]
- Cousins, L.T.; Bealing, D.J.; James, H.A.; Sutton, A. Biodegradation of microcystin-LR by indigenous mixed bacterial populations. Water Res. 1996, 30, 481–485. [Google Scholar] [CrossRef]
- WHO. Cyanobacterial Toxins: Microcystin-LR, Guidelines for Drinking-Water Quality; World Health Organization: Geneva, Switzerland, 1998; pp. 95–110. [Google Scholar]
- Gilroy, D.J.; Kauffman, K.W.; Hall, R.A.; Huang, X.; Chu, F.S. Assessing potential health risks from microcystin toxins in blue-green algae dietary supplements. Environ. Health Perspect. 2000, 108, 435–439. [Google Scholar] [CrossRef]
- Codd, G.A.; Metcalf, J.S.; Beattie, K.A. Retention of Microcystis aeruginosa and microcystin by salad lettuce (Lactuca sativa) after spray irrigation with water containing cyanobacteria. Toxicon 1999, 37, 1181–1185. [Google Scholar] [CrossRef]
- Fawell, J.K.; Mitchell, R.E.; Everett, D.J.; Hill, R.E. The toxicity of cyanobacterial toxins in the mouse: I microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.L.; Rodrigues, J.A.; Azevedo, J.; Vasconcelos, V.; Eiras, E.; Campos, M.G. Hepatotoxicity induced by paclitaxel interaction with turmeric in association with a microcystin from a contaminated dietary supplement. Toxicon 2018, 150, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Grobbelaar, J.U. Quality control and assurance: Crucial for the sustainability of the applied phycology industry. J. Appl. Phycol. 2003, 15, 209–215. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target Organisms | Plating Medium | Inoculation Technique | Incubation |
|---|---|---|---|
| Mesophilic microorganisms & Bacillus spores | Plate count agar (PCA) 1 | Surface spread, quarter plate, 20 μL | 30 °C, 72 h |
| Clostridium & Clostridium spores | Reinforced Clostridial Agar (RCA) 1 | Surface spread, quarter plate, 20 μL | 30 °C, 72 h |
| Lactic acid bacteria | De Man, Rogosa and Sharpe (MRSA)1 | Surface spread, quarter plate, 20 μL | 30 °C, 72 h, candle jars |
| Thermotolerant enterococci & streptococci | KF streptococcus medium (KF) 2 | Surface spread, quarter plate, 20 μL | 37 °C, 4 h then 44 °C, 44 h. |
| Staphylococcus aureus | Mannitol Salt Agar (MSA) 1 | Surface spread, quarter plate, 20 μL | 37 °C, 48 h |
| Yeasts and moulds | Yeast glucose chloramphenicol agar (YGC) 3 | Surface spread, 100 μL | 25 °C, 3–5 days |
| Listeria spp. | Oxford agar 1 | Surface spread, 500 μL | 30 °C, 48 h |
| Coliforms | Violet red bile agar (VRBA) 1 | Pour plate, 1 mL, with overlay | 37 °C, 48 h |
| Pseudomonas spp. | Cetrimide Agar medium 4 + 10 mL glycerol 3 | Surface spread, quarter plate, 20 μL | 37 °C, 48 h |
| Vibrio spp. |
| 10 g of sample in 90 mL Plates were streaked with isolation technique | 37 °C, 24 h 37 °C, 24 h |
| Salmonella spp. |
| 10 g of sample in 90 mL pre-enrichment broth 1 mL of pre-enrichment broth in 10 mL enrichment broths Plates were streaked with isolation technique | 37 °C, 24 h 37 °C, 24 h 37 °C, 24 h |
| Analysis | Powder | Tablets | Lemon Cereal Bar | Tahini with Honey | Cocoa-Praline |
|---|---|---|---|---|---|
| log cfu/g ± Standard Deviation | |||||
| Aerobic plate count | 3.2 ± 0.17 | 1.9 ± 0.25 | 2.2 ± 0.06 | 3.1 ± 0.11 | 2.5 ± 0.18 |
| Anaerobic plate count | 2.7 ± 0.43 | <1.0 | <1.0 | 3.1 ± 0.01 | 2.1 ± 0.12 |
| Aerobic spore count | 2.6 ± 0.41 | 1.7 ± 0.12 | 1.7 ± 0.14 | 2.7 ± 0.51 | <2.0 |
| Anaerobic spore count | 2.7 ± 0.81 | <1.0 | <1.0 | 2.1 ± 0.12 | <2.0 |
| Pseudomonas spp. | 2.8 ± 0.36 | <2.7 | <2.7 | <2.7 | <2.7 |
| Lactic acid bacteria | <2.7 | <2.7 | <2.7 | 3.1 ± 0.12 | <2.7 |
| Yeasts and Moulds | 3.0 ± 0.28 | <2.7 | <2.7 | <2.7 | <2.7 |
| Taxonomic Level | Name | Relative Abundance (%) |
|---|---|---|
| Phylum | Proteobacteria Firmicutes Actinobacteria | 33.8 32.4 13.5 |
| Family | Bacillaceae Rhodobacteraceae Enterobacteriaceae Enterococcaceae Lactobacillaceae Microbacteriaceae Micrococcaceae | 13.5 9.5 6.8 5.4 5.4 4.1 4.1 |
| Product | Isolation Medium | Number of Colonies | Species Identified |
|---|---|---|---|
| Spirulina powder | PCA, RCA, KF PCA PCA PCA MSA | 6 4 1 1 1 | Enterococcus faecium Bacillus badius Bacillus subtilis Bacillus pumilus Bacillus mojavensis |
| Spirulina tablets | SS XLD XLD TCBS PCA Oxford TCBS RCA RCA | 1 1 1 1 1 1 1 1 1 | Enterobacter cloacae Enterobacter kobei Enterobacter asburiae Enterococcus faecium Micrococcus luteus Bacillus cereus Bacillus pumilus Curtobacterium albidum Pseudomonas synxantha |
| Lemon cereal bar | PCA PCA, RCA PCA PCA PCA PCA PCA PCA | 3 2 1 1 1 1 1 1 | Staphylococcus xylosus Staphylococcus hominis Micrococcus luteus Bacillus cereus Bacillus subtilis Bacillus mojavensis Bacillus altitudinis Pseudomonas rhodesiae |
| Tahini with honey | Oxford, PCA, RCA PCA, RCA MRS, RCA MSA PCA PCA PCA | 4 3 2 1 1 1 1 | Bacillus cereus Leucanostoc mesenteroides Bacillus subtilis Bacillus atrophaeus Bacillus vallismortis Brevibacterium casei Micrococcus luteus |
| Cocoa-praline | PCA, RCA, Oxford MSA, Oxford PCA | 6 2 1 | Bacillus cereus Bacillus licheniformis Enterococcus faecium |
| MCYST | CYN | STX | |
|---|---|---|---|
| Product | ng g−1 | ||
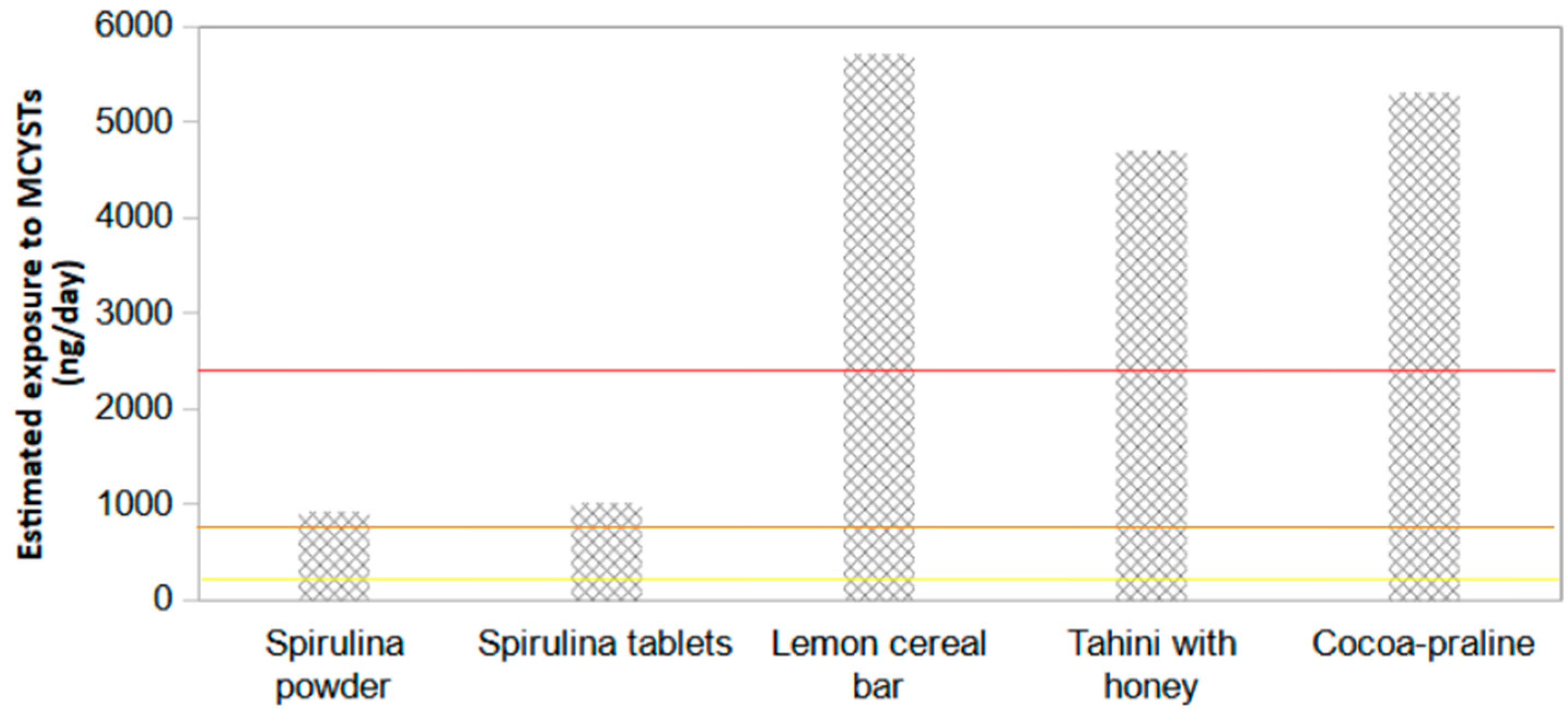
| Spirulina powder | 285.8 | <0.040 | <0.015 |
| Spirulina tablets | 321.3 | <0.040 | <0.015 |
| Lemon cereal bar | 143.3 | <0.040 | <0.015 |
| Tahini with honey | 188.1 | <0.040 | <0.015 |
| Cocoa-praline | 212.2 | <0.040 | <0.015 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhoades, J.; Fotiadou, S.; Paschalidou, G.; Papadimitriou, T.; Ordóñez, A.Á.; Kormas, K.; Vardaka, E.; Likotrafiti, E. Microbiota and Cyanotoxin Content of Retail Spirulina Supplements and Spirulina Supplemented Foods. Microorganisms 2023, 11, 1175. https://doi.org/10.3390/microorganisms11051175
Rhoades J, Fotiadou S, Paschalidou G, Papadimitriou T, Ordóñez AÁ, Kormas K, Vardaka E, Likotrafiti E. Microbiota and Cyanotoxin Content of Retail Spirulina Supplements and Spirulina Supplemented Foods. Microorganisms. 2023; 11(5):1175. https://doi.org/10.3390/microorganisms11051175
Chicago/Turabian StyleRhoades, Jonathan, Stamatia Fotiadou, Georgia Paschalidou, Theodoti Papadimitriou, Avelino Álvarez Ordóñez, Konstantinos Kormas, Elisabeth Vardaka, and Eleni Likotrafiti. 2023. "Microbiota and Cyanotoxin Content of Retail Spirulina Supplements and Spirulina Supplemented Foods" Microorganisms 11, no. 5: 1175. https://doi.org/10.3390/microorganisms11051175
APA StyleRhoades, J., Fotiadou, S., Paschalidou, G., Papadimitriou, T., Ordóñez, A. Á., Kormas, K., Vardaka, E., & Likotrafiti, E. (2023). Microbiota and Cyanotoxin Content of Retail Spirulina Supplements and Spirulina Supplemented Foods. Microorganisms, 11(5), 1175. https://doi.org/10.3390/microorganisms11051175