

Bacillus cereus in the Artisanal Cheese Production Chain in Southwestern Mexico
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification of Bacterial Strains
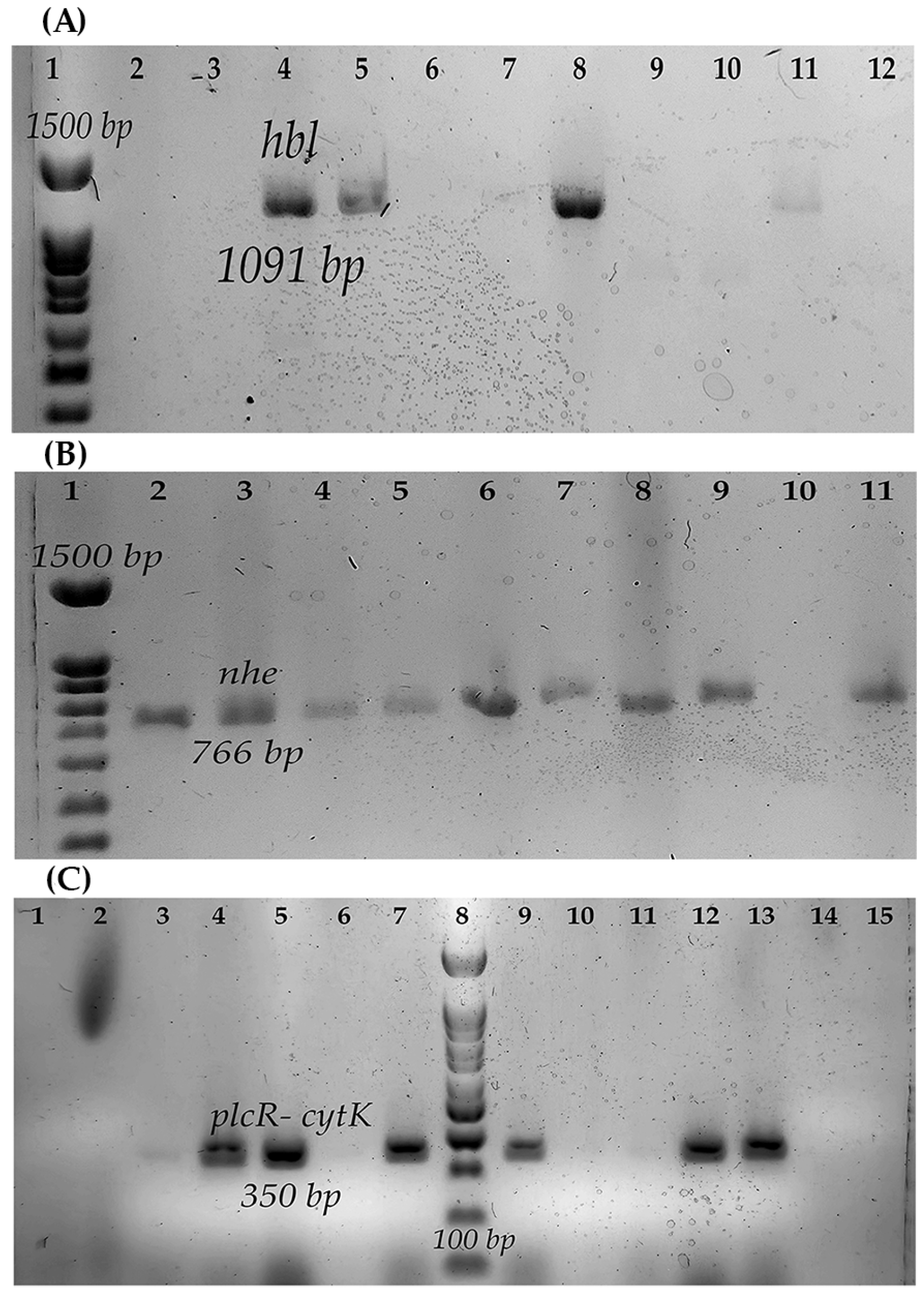
2.3. Genotyping and Enterotoxigenic Profile of B. cereus
2.4. Determination of B. cereus Biofilms
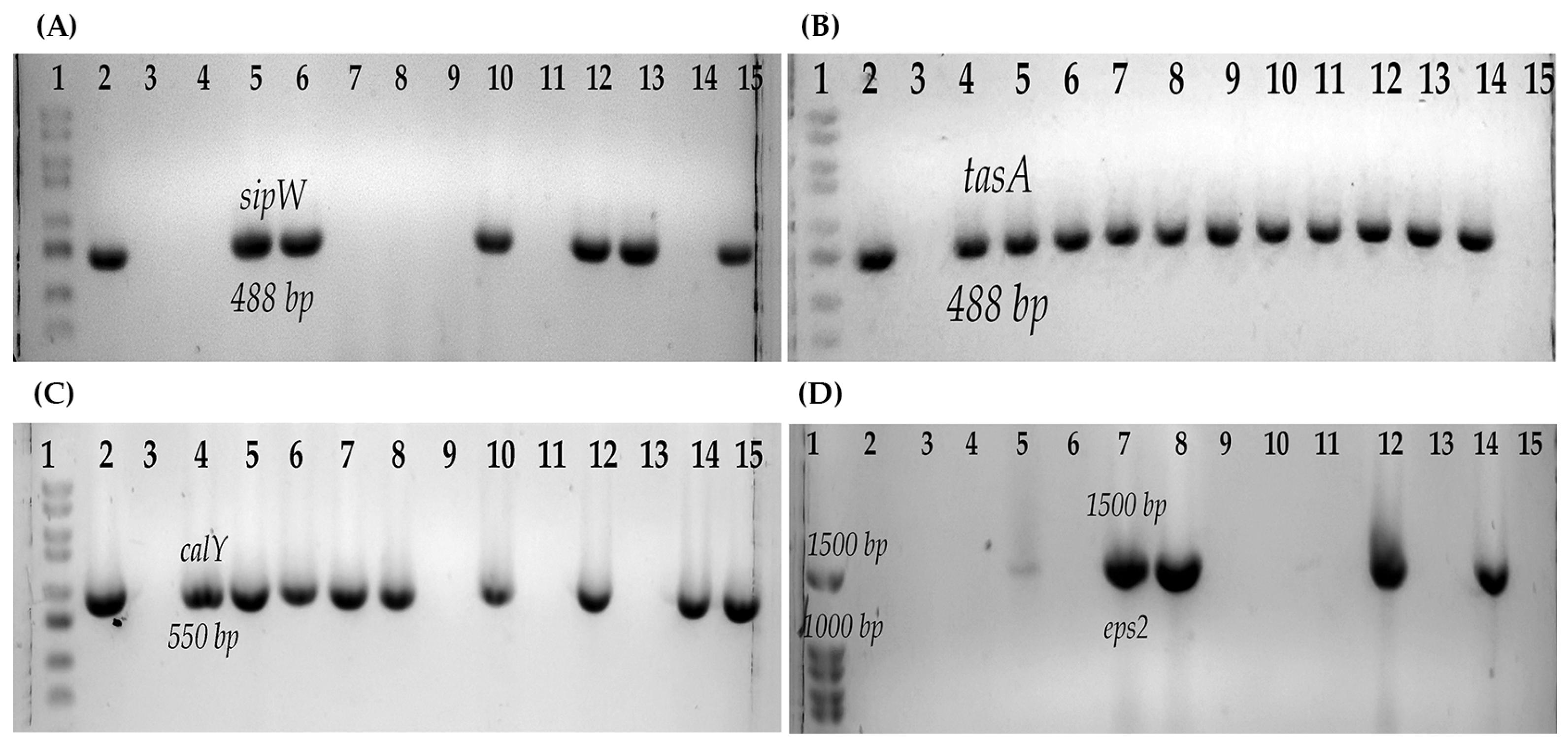
2.5. Determination of Genes Involved in the Formation of B. cereus Biofilms
2.6. Determination of Extracellular Enzymes and Cold Tolerance of B. cereus
2.7. Determination of Antimicrobial Resistance
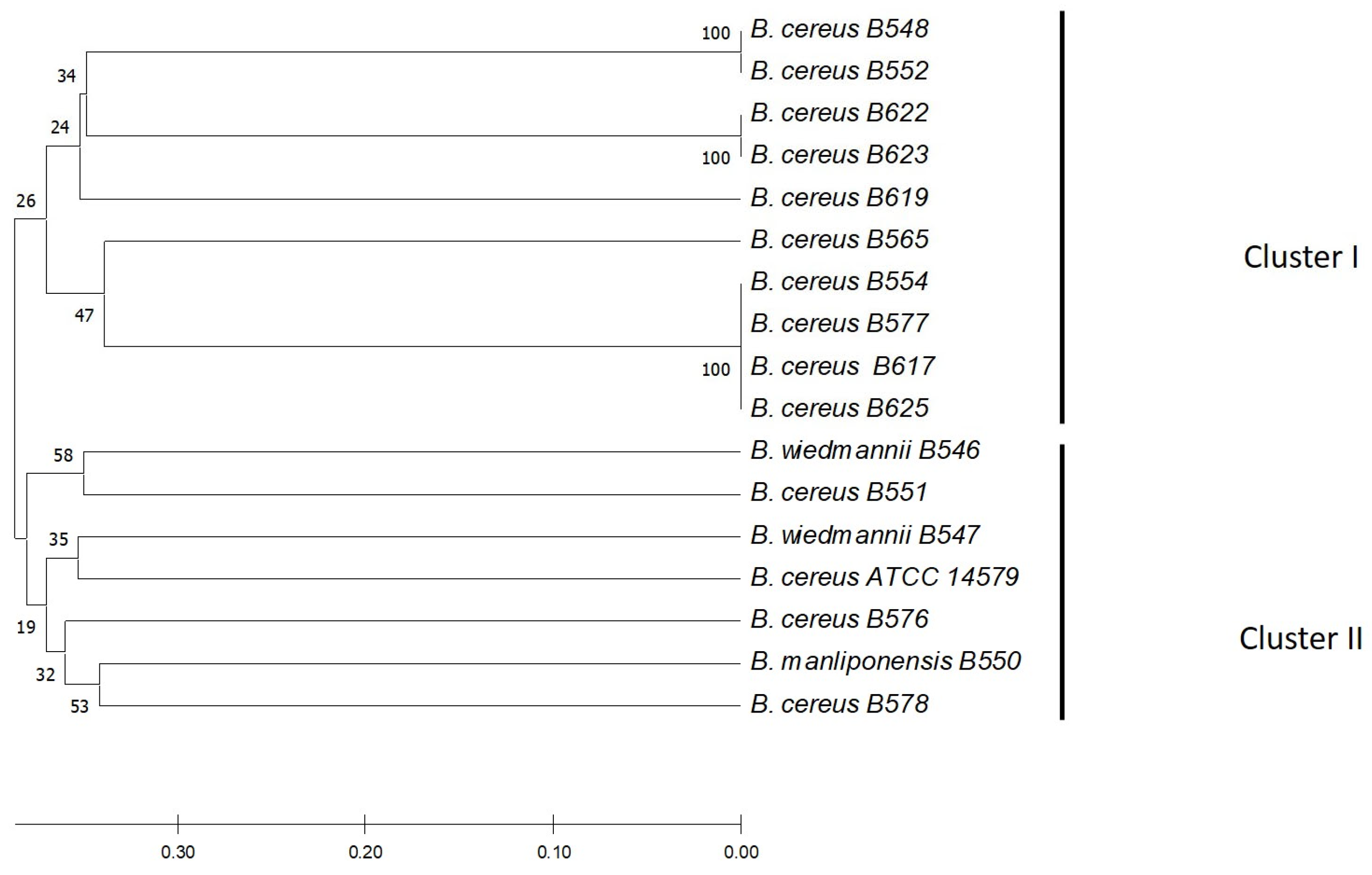
2.8. Bioinformatics Analysis
3. Results
3.1. Isolation and Identification of Bacterial Strains
3.2. Enterotoxigenic Profile of B. cereus
3.3. Determination of B. cereus Metabolic Traits
3.4. Determination of Biofilms and Gene Involved in the Biofilm Formation in Strains of B. cereus s.l.
3.5. Determination of Antimicrobial Resistance
3.6. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ankolekar, C.; Rahmati, T.; Labbé, R.G. Detection of toxigenic Bacillus cereus and Bacillus thuringiensis spores in U.S. Rice. Int. J. Food Microbiol. 2009, 128, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Frenzel, E.; Gohar, M. Food–bacteria interplay: Pathometabolism of emetic Bacillus cereus. Front. Microbiol. 2015, 6, 704. [Google Scholar] [CrossRef] [PubMed]
- Helgason, E.; Økstad, O.A.; Caugant, D.A.; Johansen, H.A.; Fouet, A.; Mock, M.; Hegna, I.; Kolstø, A.-B. Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis—One species on the basis of genetic evidence. Appl. Environ. Microbiol. 2000, 66, 2627–2630. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.; Altherr, M.; Han, C.; Ravel, J. Genomics of the Bacillus cereus group of organisms. FEMS Microbiol. Rev. 2005, 29, 303–329. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef]
- Vilas-Boas, G.; Sanchis, V.; Lereclus, D.; Lemos, M.V.F.; Bourguet, D. Genetic differentiation between sympatric populations of Bacillus cereus and Bacillus thuringiensis. Appl. Environ. Microbiol. 2002, 68, 1414–1424. [Google Scholar] [CrossRef]
- Granum, P.E.; Lund, T. Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Lett. 2006, 157, 223–228. [Google Scholar] [CrossRef]
- Agata, N.; Ohta, M.; Mori, M.; Isobe, M. A novel dodecadepsipeptide, cereulide, is an emetic toxin of Bacillus cereus. FEMS Microbiol. Lett. 1995, 129, 17–19. [Google Scholar] [CrossRef]
- Agata, N.; Ohta, M.; Yokoyama, K. Production of Bacillus cereus emetic toxin (cereulide) in various foods. Int. J. Food Microbiol. 2002, 73, 23–27. [Google Scholar] [CrossRef]
- Shinagawa, K.; Konuma, H.; Sekita, H.; Sugii, S. Emesis of rhesus monkeys induced by intragastric administration with the HEp-2 vacuolation factor (cereulide) produced by Bacillus cereus. FEMS Microbiol. Lett. 1995, 130, 87–90. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Märtlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the CytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [PubMed]
- Granum, P. The sequence of the non-haemolytic enterotoxin operon from Bacillus cereus. FEMS Microbiol. Lett. 1999, 177, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; De Buyser, M.-L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef]
- Wong, A.C.L.; Beecher, D.J. Tripartite haemolysin BL: Isolation and characterization of two distinct homologous sets of components from a single Bacillus cereus isolate. Microbiology 2000, 146, 1371–1380. [Google Scholar] [CrossRef]
- Agata, N.; Ohta, M.; Arakawa, Y.; Mori, M. The BceT gene of Bacillus cereus encodes an enterotoxic protein. Microbiology 1995, 141, 983–988. [Google Scholar] [CrossRef]
- Asano, S.I.; Nukumizu, Y.; Bando, H.; Iizuka, T.; Yamamoto, T. Cloning of novel enterotoxin genes from Bacillus cereus and Bacillus thuringiensis. Appl. Environ. Microbiol. 1997, 63, 1054–1057. [Google Scholar] [CrossRef]
- Choma, C.; Granum, P.E. The enterotoxin T (BcET) from Bacillus cereus can probably not contribute to food poisoning. FEMS Microbiol. Lett. 2002, 217, 115–119. [Google Scholar] [CrossRef]
- Arslan, S.; Eyi, A.; Küçüksarı, R. Toxigenic genes, spoilage potential, and antimicrobial resistance of Bacillus cereus group strains from ice cream. Anaerobe 2014, 25, 42–46. [Google Scholar] [CrossRef]
- Němečková, I.; Pechačová, M.; Roubal, P. Problems with detection of proteolytic microorganisms and their undesirable activities in milk. Czech J. Food Sci. 2010, 27, 82–89. [Google Scholar] [CrossRef]
- Chen, L.; Daniel, R.M.; Coolbear, T. Detection and impact of protease and lipase activities in milk and milk powders. Int. Dairy J. 2003, 13, 255–275. [Google Scholar] [CrossRef]
- Bartoszewicz, M.; Hansen, B.M.; Swiecicka, I. The members of the Bacillus cereus group are commonly present contaminants of fresh and heat-treated milk. Food Microbiol. 2008, 25, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, virulence genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Front. Microbiol. 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Fei, P.; Yuan, X.; Zhao, S.; Yang, T.; Xiang, J.; Chen, X.; Zhou, L.; Ji, M. Prevalence and genetic diversity of Bacillus cereus isolated from raw milk and cattle farm environments. Curr. Microbiol. 2019, 76, 1355–1360. [Google Scholar] [CrossRef]
- Adame-Gómez, R.; Muñoz-Barrios, S.; Castro-Alarcón, N.; Leyva-Vázquez, M.-A.; Toribio-Jiménez, J.; Ramírez-Peralta, A. Prevalence of the strains of Bacillus cereus group in artisanal mexican cheese. Foodborne Pathog. Dis. 2020, 17, 8–14. [Google Scholar] [CrossRef]
- Adame-Gómez, R.; Toribio-Jimenez, J.; Vences-Velazquez, A.; Rodríguez-Bataz, E.; Santiago Dionisio, M.C.; Ramirez-Peralta, A. Methicillin-resistant Staphylococcus aureus (MRSA) in artisanal cheeses in México. Int. J. Microbiol. 2018, 2018, 8760357. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef]
- Christison, C.A.; Lindsay, D.; von HOLY, A. Cleaning and handling implements as potential reservoirs for bacterial contamination of some ready-to-eat foods in retail delicatessen environments. J. Food Prot. 2007, 70, 2878–2883. [Google Scholar] [CrossRef]
- Shemesh, M.; Ostrov, I. Role of Bacillus species in biofilm persistence and emerging antibiofilm strategies in the dairy industry. J. Sci. Food Agric. 2020, 100, 2327–2336. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, X.; Dietrich, R.; Märtlbauer, E.; Cao, J.; Ding, S.; Zhu, K. Characterization of Bacillus cereus isolates from local dairy farms in China. FEMS Microbiol. Lett. 2016, 363, fnw096. [Google Scholar] [CrossRef]
- Dréan, P.; McAuley, C.M.; Moore, S.C.; Fegan, N.; Fox, E.M. Characterization of the spore-forming Bacillus cereus sensu lato group and clostridium perfringens bacteria isolated from the Australian dairy farm environment. BMC Microbiol. 2015, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Vissers, M.M.M.; Te Giffel, M.C.; Driehuis, F.; De Jong, P.; Lankveld, J.M.G. Minimizing the level of Bacillus cereus spores in farm tank milk. J. Dairy Sci. 2007, 90, 3286–3293. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vitela, M.R.; Mendoza-Bernardo, M.; Castro-Rosas, J.; Gomez-Aldapa, C.A.; Garay-Martinez, L.E.; Navarro-Hidalgo, V.; Villarruel-López, A. Incidence of Salmonella, Listeria monocytogenes, Escherichia coli O157:H7, and staphylococcal enterotoxin in two types of Mexican fresh cheeses. J. Food Prot. 2012, 75, 79–84. [Google Scholar] [CrossRef]
- Listeria Outbreak Linked to Hispanic-Style Fresh and Soft Cheeses|CDC. Available online: https://www.cdc.gov/listeria/outbreaks/hispanic-soft-cheese-02-21/index.html (accessed on 30 March 2023).
- Soto Beltran, M.; Gerba, C.P.; Porto Fett, A.; Luchansky, J.B.; Chaidez, C. Prevalence and characterization of Listeria monocytogenes, Salmonella and Shiga toxin-producing Escherichia coli isolated from small mexican retail markets of queso fresco. Int. J. Environ. Health Res. 2015, 25, 140–148. [Google Scholar] [CrossRef]
- Holle, M.J.; Ibarra-Sánchez, L.A.; Liu, X.; Stasiewicz, M.J.; Miller, M.J. Microbial analysis of commercially available US queso fresco. J. Dairy Sci. 2018, 101, 7736–7745. [Google Scholar] [CrossRef]
- Norma Oficial Mexicana 110 (NOM-110); Bienes y Servicios. Preparación y Dilución de Muestras de Alimentos para su Análisis Microbiológico. Secretaria de Salud (SSA), Diario Oficial de la Federacion: Mexico City, Mexico, 1994.
- ISO. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Presumptive Bacillus cereus—Colony-Count Technique at 30 °C (ISO 7932:2004); International Organization for Standardization: Geneva, Switzerland, 2005. [Google Scholar]
- Bavykin, S.G.; Lysov, Y.P.; Zakhariev, V.; Kelly, J.J.; Jackman, J.; Stahl, D.A.; Cherni, A. Use of 16S rRNA, 23S rRNA, and gyrB gene sequence analysis to determine phylogenetic relationships of Bacillus cereus group microorganisms. J. Clin. Microbiol. 2004, 42, 3711–3730. [Google Scholar] [CrossRef] [PubMed]
- Ołtuszak-Walczak, E.; Walczak, P.; Modrak, R. Detection of enterotoxic Bacillus cereus producing hemolytic and non hemolytic enterotoxins by PCR test. Pol. J. Microbiol. 2006, 55, 113–118. [Google Scholar]
- Ehling-Schulz, M.; Guinebretiere, M.-H.; Monthán, A.; Berge, O.; Fricker, M.; Svensson, B. Toxin gene profiling of enterotoxic and emetic Bacillus cereus. FEMS Microbiol. Lett. 2006, 260, 232–240. [Google Scholar] [CrossRef]
- Adame-Gómez, R.; Cruz-Facundo, I.-M.; García-Díaz, L.-L.; Ramírez-Sandoval, Y.; Pérez-Valdespino, A.; Ortuño-Pineda, C.; Santiago-Dionisio, M.-C.; Ramírez-Peralta, A. Biofilm production by enterotoxigenic strains of Bacillus cereus in different materials and under different environmental conditions. Microorganisms 2020, 8, 1071. [Google Scholar] [CrossRef]
- Caro-Astorga, J.; Pérez-García, A.; de Vicente, A.; Romero, D. A genomic region involved in the formation of adhesin fibers in Bacillus cereus biofilms. Front. Microbiol. 2014, 5, 745. [Google Scholar] [CrossRef]
- Caro-Astorga, J.; Álvarez-Mena, A.; Hierrezuelo, J.; Guadix, J.A.; Heredia-Ponce, Z.; Arboleda-Estudillo, Y.; González-Munoz, E.; de Vicente, A.; Romero, D. Two genomic regions encoding exopolysaccharide production systems have complementary functions in B. cereus multicellularity and host interaction. Sci. Rep. 2020, 10, 1000. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; Clinical and Laboratory Standards Institute: Berwyn, PA, USA, 2015; Volume M45, ISBN 1-56238-918-1. [Google Scholar]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in dairy products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.; Christiansson, A.; Svensson, B. Bacillus cereus spores during housing of dairy cows: Factors affecting contamination of raw milk. J. Dairy Sci. 2007, 90, 2745–2754. [Google Scholar] [CrossRef] [PubMed]
- Adame- Gomez, R.; Guzman- Guzman, K.; Vences-Velazquez, A.; Leyva-Vazquez, M.-A.; Muñoz-Barrios, S.; Ramirez-Peralta, A. Prevalence and genetic diversity of the strains of Bacillus cereus groups in food for infants and young children in Mexico. Afr. J. Microbiol. Res. 2018, 12, 730–735. [Google Scholar] [CrossRef]
- Cruz-Facundo, I.; Adame-Gómez, R.; Vences-Velázquez, A.; Rodríguez-Bataz, E.; Muñoz-Barrios, S.; Pérez-Oláis, J.; Ramírez-Peralta, A. Bacillus cereus in eggshell: Enterotoxigenic profiles and biofilm production. Braz. J. Poult. Sci. 2022, 24, eRBCA-2021-1535. [Google Scholar] [CrossRef]
- Adame-Gomez, R.; Castro Alarcón, N.; Vences-Velázquez, A.; Rodríguez-Bataz, E.; Santiago-Dionisio, M.C.; Ramírez-Peralta, A. Prevalencia de cepas del grupo de Bacillus cereus productoras de biopelicula en helados comercializados en México. Kasmera 2019, 47, 115–122. [Google Scholar] [CrossRef]
- Rigourd, V.; Barnier, J.P.; Ferroni, A.; Nicloux, M.; Hachem, T.; Magny, J.F.; Lapillonne, A.; Frange, P.; Nassif, X.; Bille, E. Recent actuality about Bacillus cereus and human milk bank: A new sensitive method for microbiological analysis of aasteurized milk. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1297–1303. [Google Scholar] [CrossRef]
- Chang, Y.; Xie, Q.; Yang, J.; Ma, L.; Feng, H. The prevalence and characterization of Bacillus cereus isolated from raw and pasteurized buffalo milk in Southwestern China. J. Dairy Sci. 2021, 104, 3980–3989. [Google Scholar] [CrossRef]
- Radmehr, B.; Zaferanloo, B.; Tran, T.; Beale, D.J.; Palombo, E.A. Prevalence and characteristics of Bacillus cereus group isolated from raw and pasteurised milk. Curr. Microbiol. 2020, 77, 3065–3075. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, J.; Fei, P.; Feng, H.; Wang, Y.; Ali, M.A.; Li, S.; Jing, H.; Yang, W. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J. Dairy Sci. 2020, 103, 3994–4001. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Zhang, R.; Dong, L.; Hu, H.; Liu, H.; Zheng, N.; Wang, J.; Cheng, J. Characterization and spoilage potential of Bacillus cereus isolated from farm environment and raw milk. Front. Microbiol. 2022, 13, 940611. [Google Scholar] [CrossRef] [PubMed]
- Carlin, F.; Fricker, M.; Pielaat, A.; Heisterkamp, S.; Shaheen, R.; Salkinojasalonen, M.; Svensson, B.; Nguyenthe, C.; Ehlingschulz, M. Emetic toxin-producing strains of Bacillus cereus show distinct characteristics within the Bacillus cereus group. Int. J. Food Microbiol. 2006, 109, 132–138. [Google Scholar] [CrossRef]
- Peng, J.-S.; Tsai, W.-C.; Chou, C.-C. Inactivation and removal of Bacillus cereus by sanitizer and detergent. Int. J. Food Microbiol. 2002, 77, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Candela, T.; Fagerlund, A.; Buisson, C.; Gilois, N.; Kolstø, A.; Økstad, O.A.; Aymerich, S.; Nielsen-Leroux, C.; Lereclus, D.; Gohar, M. CalY is a major virulence factor and a biofilm matrix Protein. Mol. Microbiol. 2019, 111, 1416–1429. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.S.; Kwon, M.; Oh, D.-H. Impact of manganese and heme on biofilm formation of Bacillus cereus food isolates. PLoS ONE 2018, 13, e0200958. [Google Scholar] [CrossRef]
- Banerjee, M.; Sarkar, P.K. Antibiotic resistance and susceptibility to some food preservative measures of spoilage and pathogenic microorganisms from spices. Food Microbiol. 2004, 21, 335–342. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Accession Number on GenBank | Identification | Closest Related Bacteria | Similarity |
|---|---|---|---|---|
| B546 | OQ507680 | B. wiedmannii | B. wiedmannii FSL W8-0169 | 99.75 |
| B547 | OQ507681 | B. wiedmannii | B. wiedmannii FSL W8-0169 | 99.88 |
| B548 | OQ507682 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| B550 | OQ507685 | B. manliponensis | B. manliponensis BL4-6 | 89.55 |
| B551 | OQ507683 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| B552 | OQ507686 | B. cereus | B. cereus ATCC 14579 | 99.69 |
| B554 | OQ507687 | B. cereus | B. cereus ATCC 14579 | 99.16 |
| B565 | OQ507688 | B. cereus | B. cereus ATCC 14579 | 99.86 |
| B576 | OQ507688 | B. cereus | B. cereus ATCC 14579 | 99.87 |
| B577 | OQ507690 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| B578 | OQ507691 | B. cereus | B. cereus ATCC 14579 | 99.87 |
| B617 | OQ507692 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| B619 | OQ507693 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| B622 | OQ507684 | B. cereus | B. cereus ATCC 14579 | 99.86 |
| B623 | OQ507694 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| B625 | OQ507695 | B. cereus | B. cereus ATCC 14579 | 100.00 |
| Strain | Identification | Source | Diarrheagenic | Emetic | ||
|---|---|---|---|---|---|---|
| hbl | nhe | cytk | ces | |||
| B546 | B. wiedmannii | Air | − | + | − | − |
| B547 | B. wiedmannii | Air | + | + | + | − |
| B548 | B. cereus | Air | + | + | + | − |
| B550 | B. manliponensis | Air | − | + | + | − |
| B551 | B. cereus | Air | − | + | + | − |
| B552 | B. cereus | Air | + | + | + | − |
| B554 | B. cereus | Air | − | + | + | − |
| B565 | B. cereus | Air | − | + | − | − |
| B576 | B. cereus | Air | − | + | − | − |
| B577 | B. cereus | Air | + | + | − | − |
| B578 | B. cereus | Air | − | − | − | − |
| B617 | B. cereus | QF 1 | − | + | + | − |
| B619 | B. cereus | QF | − | + | − | − |
| B622 | B. cereus | QF | − | + | + | − |
| B623 | B. cereus | QF | − | + | − | − |
| B625 | B. cereus | QF | − | + | + | − |
| Strain | Lecithinase (mm) | Protease | Amylase |
|---|---|---|---|
| B546 | 0.47 | + | − |
| B547 | 0.42 | + | − |
| B548 | 0.55 | + | + |
| B550 | 0.45 | + | + |
| B551 | 0.50 | + | − |
| B552 | 0.67 | + | − |
| B554 | 0.73 | + | − |
| B565 | 0.48 | + | − |
| B576 | 0.55 | + | − |
| B577 | 0.48 | + | − |
| B578 | 0.52 | + | − |
| B617 | 0.65 | + | − |
| B619 | 0.60 | + | − |
| B622 | 0.45 | + | − |
| B623 | 0.57 | + | − |
| B625 | 0.62 | + | + |
| Strain | Psychrotrophic Growth (DT) | Biofilm | tasA | sipW | calY | eps2 |
|---|---|---|---|---|---|---|
| B546 | 1 | − | − | − | + | − |
| B547 | 1 | + | + | + | + | − |
| B548 | 1 | + | + | + | + | − |
| B550 | 1 | + | − | − | + | + |
| B551 | 1 | ++ | − | − | + | − |
| B552 | 1 | + | + | + | + | + |
| B554 | 1 | + | + | − | + | + |
| B565 | 1 | + | + | + | + | − |
| B576 | 1 | ++ | + | + | + | − |
| B577 | 1 | + | − | − | + | − |
| B578 | 1 | + | + | + | + | + |
| B617 | 5 | ++ | + | − | − | − |
| B619 | 5 | + | + | − | − | − |
| B622 | 1 | ++ | + | − | + | − |
| B623 | 5 | − | − | − | − | − |
| B625 | 5 | + | + | − | − | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Facundo, I.-M.; Toribio-Jiménez, J.; Castro-Alarcón, N.; Leyva-Vázquez, M.-A.; Rodríguez-Ruíz, H.-A.; Pérez-Olais, J.-H.; Adame-Gómez, R.; Rodríguez-Bataz, E.; Reyes-Roldán, J.; Muñoz-Barrios, S.; et al. Bacillus cereus in the Artisanal Cheese Production Chain in Southwestern Mexico. Microorganisms 2023, 11, 1290. https://doi.org/10.3390/microorganisms11051290
Cruz-Facundo I-M, Toribio-Jiménez J, Castro-Alarcón N, Leyva-Vázquez M-A, Rodríguez-Ruíz H-A, Pérez-Olais J-H, Adame-Gómez R, Rodríguez-Bataz E, Reyes-Roldán J, Muñoz-Barrios S, et al. Bacillus cereus in the Artisanal Cheese Production Chain in Southwestern Mexico. Microorganisms. 2023; 11(5):1290. https://doi.org/10.3390/microorganisms11051290
Chicago/Turabian StyleCruz-Facundo, Itzel-Maralhi, Jeiry Toribio-Jiménez, Natividad Castro-Alarcón, Marco-Antonio Leyva-Vázquez, Hugo-Alberto Rodríguez-Ruíz, José-Humberto Pérez-Olais, Roberto Adame-Gómez, Elvia Rodríguez-Bataz, Joel Reyes-Roldán, Salvador Muñoz-Barrios, and et al. 2023. "Bacillus cereus in the Artisanal Cheese Production Chain in Southwestern Mexico" Microorganisms 11, no. 5: 1290. https://doi.org/10.3390/microorganisms11051290
APA StyleCruz-Facundo, I. -M., Toribio-Jiménez, J., Castro-Alarcón, N., Leyva-Vázquez, M. -A., Rodríguez-Ruíz, H. -A., Pérez-Olais, J. -H., Adame-Gómez, R., Rodríguez-Bataz, E., Reyes-Roldán, J., Muñoz-Barrios, S., & Ramírez-Peralta, A. (2023). Bacillus cereus in the Artisanal Cheese Production Chain in Southwestern Mexico. Microorganisms, 11(5), 1290. https://doi.org/10.3390/microorganisms11051290